
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
华南泥盆纪生物礁演化及其控制因素
[吴义布1, 2  , 龚一鸣
, 龚一鸣1, 2, 3 , 张立军1, 2 , 冯启1, 2 ]
, 龚一鸣|
|

第一作者简介:吴义布,男,1985年生,硕士研究生,主要从事生物礁及沉积学研究.通讯地址:武汉市洪山区鲁磨路388号中国地质大学(武汉)研究生院2008级硕2班;邮政编码:430074.电话:15927243757;E-mail:ttwyb123@yahoo.com.cn.
通讯作者简介:龚一鸣,男,教授,主要从事泥盆纪,遗迹化石和沉积地质研究.
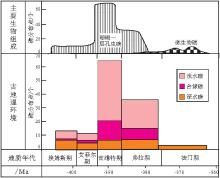
通过对华南泥盆纪生物礁现有资料和作者获得的实际材料的分析研究,编制了华南早,中,晚泥盆世生物礁分布图.华南泥盆纪生物礁最早出现在埃姆斯期晚期,分布层位由西南向北东逐渐变新.华南泥盆纪生物礁分布和演化具有 3方面的特征:( 1)不同类型生物礁演化差异很大:浅水礁变化最大,台缘礁变化次之,深水礁几乎没有变化;( 2)主要造礁生物随时间演替明显:床板珊瑚(埃姆斯期)→床板珊瑚,四射珊瑚和层孔虫(艾菲尔期)→层孔虫和四射珊瑚(吉维特期)→层孔虫(弗拉期)→菌藻类(法门期);( 3)生物礁数量和规模在吉维特期出现峰值.研究表明:华南泥盆纪生物礁的发育受区域和全球性因素的共同制约,其中温度,大气中 CO2分压(
About the first author Wu Yibu, born in 1985, is a candidate for his master degree in China University of Geosciences(Wuhan), and is mainly engaged in researches of bioreefs and sedimentology.
About the corresponding author Gong Yiming is a professor of China University of Geosciences(Wuhan), and is mainly engaged in researches of Devonian, trace fossils and sedimentary geology.
Based on the data of the Devonian bioreefs, we compiled the distribution maps of the Early, Middle and Late Devonian bioreefs in South China. The earliest Devonian bioreefs appeared in the Late Emsinian. The spatiotemporal distribution of the Early, Middle and Late Devonian bioreefs shows that development of the Devonian bioreefs gradually migrated from the southwest to the northeast in South China. Their distribution and evolution are characterized by:(1)Different bioreefs have obviously different evolution processes. The greatest and second-greatest changes occurred in the shallow-water bioreef and platform marginal bioreef, respectively, while almost no changes occurred in deep-water bioreef during the Devonian. (2)The replacement of major reef-builders were obvious from the Early Devonian to the Late Devonian: Tabulate corals(Emsian) to tabulate and rugose corals and stromatoporoids(Eifelian) to rugose corals and stromatoporoids(Givetian) to rugose corals and stromatoporoids(Frasnian) to cyanobacteria(Famennian).(3)Bioreefs reached apex in size and quantity during the Middle Devonian Givetian Age. The main controlling factors of the Devonian bioreefs in South China include both local and global factors. The former are considered to be transgression-regression cycles and terrigenous clastic supply. The latter are considered to be changes of paleotemperature,
生物礁是油气有利富集体(张明书和何起祥, 1988), 也是研究生物与环境之间关系及其演化的最好载体之一.因此, 生物礁很早就受到关注, 研究历史近200年.尽管如此, 对生物礁的定义和分类等基本问题, 至今仍然众说纷纭(范嘉松和张维, 1985; Riding, 2002; 钟建华等, 2005).因此, 文中首先对生物礁的定义, 分类等基本概念加以讨论和界定.
生物礁的定义有很多种, 比较有代表性的有Dunham 于1970年提出的生态礁, Stanton 于1967年提出的碳酸盐建隆(buildup), 以及Wood (1999)和Riding(2002)对礁的定义的重新厘定等.文中采用的定义为: 生物礁是由原地生物构建的隆起, 与周围同期沉积物有别的碳酸盐地质体(Wood, 1999; Kiessling et al., 2002).
生物礁分类是一个比较复杂的问题(范嘉松和张维, 1985; Riding, 2002; 钟建华等, 2005), 涉及沉积学, 古生物学和岩石学等诸多方面, 如生物类型, 碳酸盐组分, 原地堆积和生物礁的内部结构等(Wood, 1999; Kiessling et al., 2002; Riding, 2002; 钟建华等, 2005).在选择礁分类方案时, 往往要考虑其实用性和广泛性.根据华南泥盆纪生物礁的特征, 文中采用以下两种分类方案:
第1类:根据主要的造礁生物, 泥盆纪的生物礁可以分为层孔虫礁, 珊瑚礁, 层孔虫珊瑚礁和菌藻微生物礁(图 1-1, 1-2).这种方案的优点是可以结合礁灰岩的结构特征(骨架岩, 障积岩和粘结岩)进一步细分, 如层孔虫骨架礁(图 1-5, 1-6); 同时能很好地反映生物礁的生物组成及其地史演化, 易与非礁相地层进行对比.
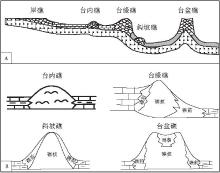
第2类:根据其发育的古地理位置, 华南泥盆纪生物礁可分为岸礁, 台内礁, 台缘礁, 斜坡礁, 台盆礁(金善燏和鞠天吟, 1998)(图 2; 图1-3, 1-5, 1-6, 1-7).该方案有利于探索生物礁的展布规律与古地理环境, 控制因素及其与油气的关系.
根据实际情况和研究目的, 文中着重以第2类分类方案进行编图和展开分析.
 | 图1 华南泥盆纪几种常见生物礁类型 1, 2--广西六景上泥盆统法门阶融县组菌藻微生物礁及其中的Izhella; 3--四川北川甘溪中泥盆统金宝石组台内礁中造礁群体四射珊瑚, 床板珊瑚和块状层孔虫; 4--北川甘溪中泥盆统金宝石组台内礁中造礁群体四射珊瑚特写(照片3中白色方框放大, 黄色圆形比例尺直径为1, cm); 5--桂林杨堤中泥盆统东岗岭组斜坡礁及内部层孔虫; 6--贵州独山上泥盆统望城坡组台内礁中的抱球状层孔虫; 7--广西南 丹六寨中泥盆统东岗岭组台缘礁远景; 8--广西六景中泥盆统民塘组礁前亚相中的珊瑚--层孔虫角砾灰岩Fig.1 Common types of the Devonian bioreefs in South China |
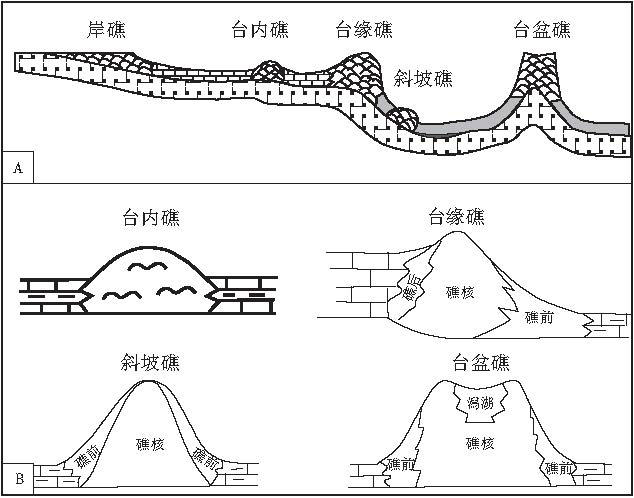 | 图2 华南泥盆纪生物礁古地理分类简图(A)及其内部结构图(B)Fig.2 Palaeogeographic classification(A)and interior structures(B) of the Devonian bioreefs in South China |
华南古生代生物礁的研究历史悠久, 已发表的成果也很多.但是在上世纪80年代中晚期之前, 研究成果主要集中在单个生物礁或小区域生物礁, 缺乏较大范围的生物礁之间的对比研究.至1988年, 曾鼎乾等编著了《中国各地质历史时期生物礁》, 这是中国第一本全面系统论述化石礁的著作; 进入90年代之后, 对生物礁系统研究的著作开始"大量"出现(如, 金善燏和鞠天吟, 1998), 这些著作对中国古生代和三叠纪生物礁的时空分布特征和演化规律进行了大尺度的总结及对比研究; 21世纪以来, 人们对化石礁的研究热情似乎有所下降, 但仍不乏有代表作问世(如, 柳祖汉等, 2000; 齐文同, 2002).
值得注意的是, 董兆雄(1987)首次对南华海泥盆纪生物礁时空分布及其控制因素进行了初步探讨, 但当时所发现的泥盆纪生物礁分布点仅有50来个.到目前为止, 泥盆纪生物礁已报导的分布点多达100余个(齐文同, 2002).
论文以作者的实际工作为基础, 侧重对前人资料的总结与对比分析, 编制了中国南方早, 中, 晚泥盆世生物礁的时空分布图.在此基础上, 对华南泥盆纪生物礁时空分布特征和组成及其演化规律进行初步总结, 并探讨其主控因素.
加里东运动之后, 东南加里东造山带隆升, 华南板块主体抬升为陆.在泥盆纪初期, 除桂东南钦防地区存在的残余海槽为次深海--深海沉积外, 华南其他地区以剥蚀为主, 仅在桂东南和川西局部地区发育有滨浅海相碎屑岩.从早泥盆世开始, 华南地区自西南滇黔桂地区逐渐向北东方向发生海侵; 早泥盆世中期区域性海侵扩大, 陆地面积缩小, 除钦防海槽外, 大部分沉积区仍以滨, 浅海碎屑岩为主, 并发育有少量灰岩; 自早泥盆世晚期(埃姆斯期)开始, 海盆中出现浅水碳酸盐台地(象州型)和条带状深水硅, 泥和泥灰质台盆及台沟沉积(南丹型), 生物相发生分异, 在滇黔桂浅海--深水台盆, 台沟区, 桂东滨浅海区及川西浅海等海域, 开始发育生物礁.
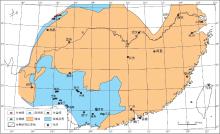
根据野外观察实测以及相关地质资料, 对埃姆斯期生物礁的出露位置及类型进行了统计和整理(表 1).在此基础上, 编制了华南早泥盆世埃姆斯期生物礁分布图(图 3).
总的来说, 埃姆斯期生物礁发育数量较少, 分布比较零星, 单个礁体规模较小, 厚度一般小于20 m.从图3可以看出, 生物礁分布在川西海域北川--江油一带, 桂东南部北流, 贵县蒙公, 云南广南, 隆林, 富宁和广西德保钦甲, 南丹大厂.造礁生物以Squameofavosites, Favosites属为代表的床板珊瑚为主, 少量层孔虫; 附礁生物主要为单体四射珊瑚, 腕足类, 海百合等.
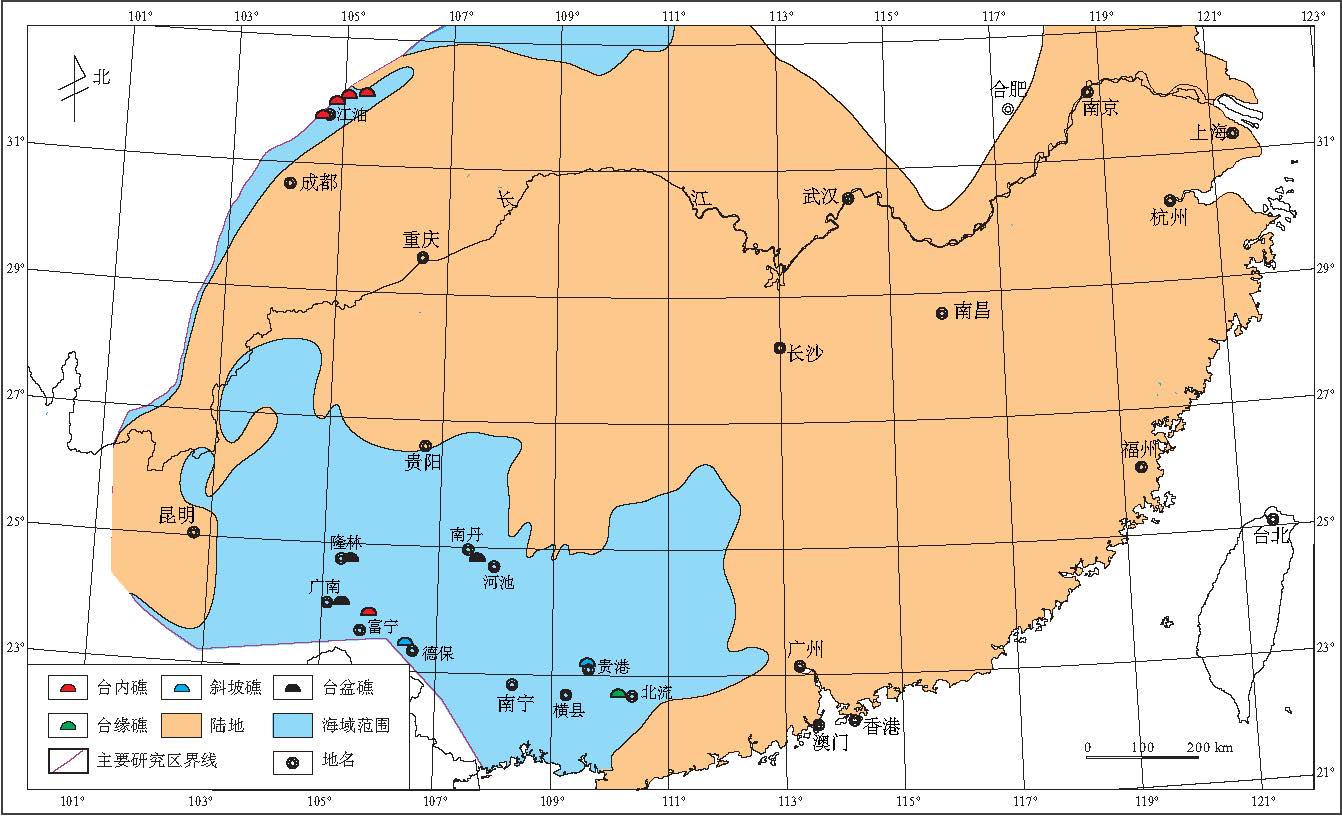 | 图3 华南早泥盆世埃姆斯期生物礁分布Fig.3 Distribution of the Early Devonian Emsian bioreefs in South China |
| 表1 华南早泥盆世埃姆斯期生物礁分布点资料统计 Table 1 Statistics of distribution of the Early Devonian Emsian bioreefs in South China |
综上所述, 华南埃姆斯期生物礁特征和分布呈现如下规律:
1)几乎所有的生物礁都发育在与外海相邻近的海域----南华海南部和川西海域北部, 而这些海域与外海连通性较好, 水流畅通.
2)台内礁主要发育在川西海域, 南华海主要发育台盆礁, 斜坡礁及台缘礁, 几乎没有台内礁发育, 整个华南海域没有岸礁发育(图 3).
3)不管是哪种类型的生物礁, 主要造礁生物一般都以Favosites, Squameofavosites, Heliotites等属的床板珊瑚为主, 主要造礁群落可以简单表示为Favosites-Heliotites群落.
华南埃姆斯期生物礁规律性的分布, 为研究其控制因素提供了良好的基础.华南早泥盆世埃姆斯期生物礁的主控因素可以概括为两方面:
1)川西海域: 在龙门山地区, 通过观察, 发现二台子组生物礁的每个旋回上部往往覆盖了潮上--潮间带沉积物; 同时, 还发现部分床板珊瑚礁被含浮游类竹节石和薄壳腕足类的黑色泥岩(深水/缺氧沉积物)所覆盖.这些现象都充分说明该区海盆基底或海平面垂向升降频繁, 从而导致生物礁生长速率跟不上海平面升降的频率, 致使生物礁暴露而死或溺死.
2)南华海:生物礁的控制因素相对复杂, 海平面变化也是一个重要的因素, 如德保钦甲礁向上变深的序列(吴诒, 1989); 生物礁主要发育在南华海南部(与外海邻近)及南丹海槽等开阔海域(图3), 整个华南没有岸礁发育, 说明盐度异常(海水循环), 陆源碎屑供给和营养物质输入对生物礁发育可能带来的负面影响是不容忽视的.
中泥盆世, 全球显生宙生物礁数量和规模达到了峰值(Copper, 1994; Kiessling et al., 1999), 到中泥盆世晚期(吉维特期), 华南生物礁也实现了首次"大爆发", 达到了显生宙生物礁分布的第一个峰值.
中泥盆世早期(艾菲尔期)基本上继承了早泥盆世晚期的古地理格局, 海域向北东方向湖南等地略有扩张, 生物礁分布与埃姆斯期基本相似: 1)钦防海槽和黔南滨浅海, 至今未发现生物礁; 2)生物礁主要分布在川西浅海碳酸盐台地, 滇黔桂深水台盆--台沟区及其中的孤立碳酸盐台地; 3)生物礁的规模一般也比较小.仅仅不同的是, 艾菲尔期以层孔虫, 床板珊瑚和四射珊瑚为主要造礁生物(图 1-3, 1-4).
中泥盆世晚期(吉维特期), 华南沉积古地理面貌发生了重大的改变.古陆进一步被夷平, 南华海随着北东向海侵, 海域迅速扩大, 与中泥盆世早期相比, 碳酸盐岩分布面积极大地增加----几乎增大了一倍, 华南出现了第一个生物礁高峰期.
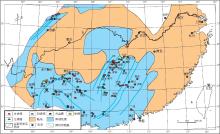
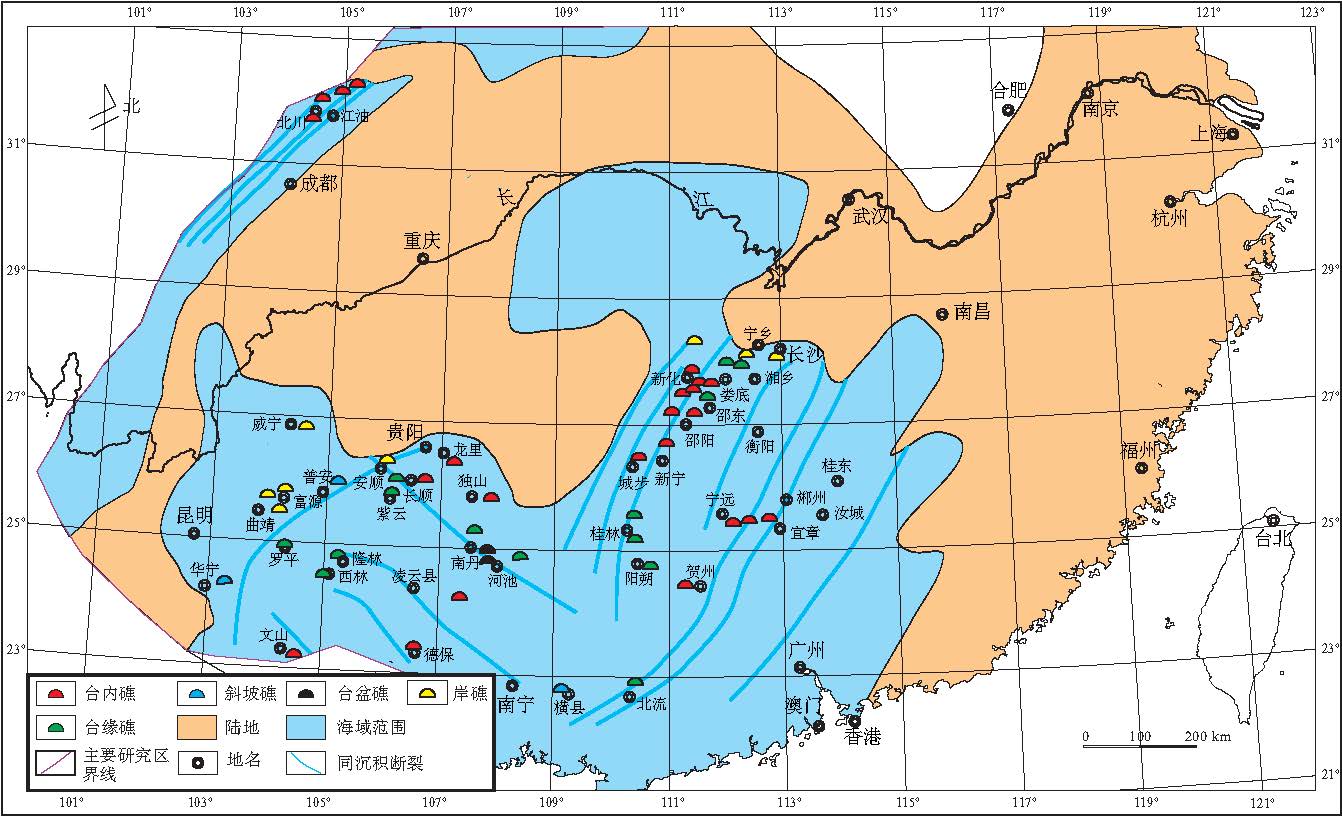 | 图4 华南中泥盆世吉维特期生物礁分布(同沉积断裂据董榕生, 1992, 有修改)Fig.4 Distribution of the Middle Devonian Givetian bioreefs in South China(Syndepositional fractures modified from Dong, 1992) |
| 表2 华南中泥盆世吉维特期生物礁分布点资料 Table 2 Data of the Middle Devonian Givetian bioreefs in South China |
根据野外观察实测以及前人地质资料, 对生物礁的分布位置和类型进行了总结(表 2), 编制了中国南方中泥盆世吉维特期生物礁分布图(图 4).
从图4可以看出, 吉维特期新增生物礁分布点多, 以台内礁为主, 台缘礁次之.这些礁体的广泛发育带来了华南显生宙生物礁的首次高峰期, 并形成了几个生物礁富集区: 川西北川--江油, 雁门坝一带, 滇中东部曲靖--富源一带, 贵州安顺--长顺--独山--桂北南丹一带和湘中南新化--新邵--邵东--涟源--韶山一带; 另外, 桂林地区台缘礁和宁远地区台内礁也较为发育.造礁生物主要以Stromatopora, Actinostroma等属为代表的层孔虫为主, 次为四射珊瑚和床板珊瑚; 附礁生物主要为腕足类, 枝状层孔虫, 单体四射珊瑚, 海百合和腹足类等.
华南中泥盆世吉维特期生物礁的特征和分布, 与埃姆斯期--艾菲尔期相比, 发生了巨大的变化:
1)新增生物礁主要为台内礁和岸礁, 其主要分布在 "浅水近岸"海域(图 4), 如开阔碳酸盐台地(局限台地一般很少发育), 但这些生物礁在吉维特期末期几乎全部灭绝了.
2)各种类型生物礁的造礁生物以层孔虫为主, 次为四射珊瑚和床板珊瑚; 菌藻类在造礁过程中起到了重要辅助造礁作用, 如环江北山, 南丹六寨生物礁等.Stromatopora-Actinostroma层孔虫群落是吉维特期大部分生物礁的重要造礁群落之一.
3)生物礁的分布与同沉积断裂密切相关, 特别是台缘礁--斜坡礁--台盆礁的分布点的连线几乎与断裂走向平行(图 4).因此, 台缘礁--斜坡礁--台盆礁的分布不仅与沉积环境参数, 如温度, 盐度, 浊度和水深密切相关, 也能示踪同沉积断裂的特征.
4)相对于埃姆斯期--艾菲尔期, 整个南华海吉维特期生物礁的富集区明显向北迁移, 迁移方向是从南华海南部(埃姆斯期)到南华海北部(吉维特期)(对比图3, 图4).
至今为止, 华南吉维特期生物礁"大爆发"的原因, 并没有引起足够的关注.本研究为解读吉维特期生物礁发育和分布规律及其控制因素, 提供了如下线索:
1)中泥盆世晚期华南海大规模海侵导致海域迅速扩大, 促使碳酸盐台地广泛分布, 为造礁生物定殖--繁盛提供了合适的环境, 如湘中南新化--新邵--邵东--韶山一带生物礁富集区的出现.
2)珊瑚对陆源物质的抵抗能力一般强于层孔虫(Kershaw, 1998; Leinfelder et al., 2005).陆源物质的减少, 促使层孔虫(吉维特期)繁盛, 并取代床板珊瑚(埃姆斯期)成为主要造礁生物, 为较大规模生物礁的发育提供了必要的条件; 但在某些地区, 如湘粤桂滨浅海碳酸盐台地, 陆源物质流入可能仍是阻碍生物礁发育的主导因素.
3)吉维特期同沉积断裂活动, 导致南华海一系列北西--南东向和北东--南西向台盆或台沟形成并扩大, 对生物礁发育和分布有重要影响: 不仅使早期的台缘礁, 斜坡礁, 台盆礁的规模增大, 还出现了新的分布点; 由于台盆或台沟存在, 海流可以沿此进入南华海内部(董兆雄, 1987), 使埃姆斯期--艾菲尔期不适合生物礁发育的滨浅海碳酸盐台地, 到吉维特期成为生物礁广泛发育的乐土.
4)盐度可能仍然对生物礁的发育起重要作用, 主要依据是该时期台内礁主要分布在开阔台地上, 在局限台地中几乎不发育, 同时附礁生物以腕足类, 四射珊瑚等窄盐性生物为主.
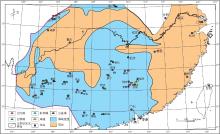
晚泥盆世早期是中国南方泥盆纪海侵最大时期, 海水继续向北东方向推进.此期古地理格架基本继承了吉维特期的特征(金善燏和鞠天吟, 1998).根据对前人资料的整理统计(表 3), 以及野外观察实测, 编制了华南晚泥盆世弗拉期生物礁分布图(图 5).
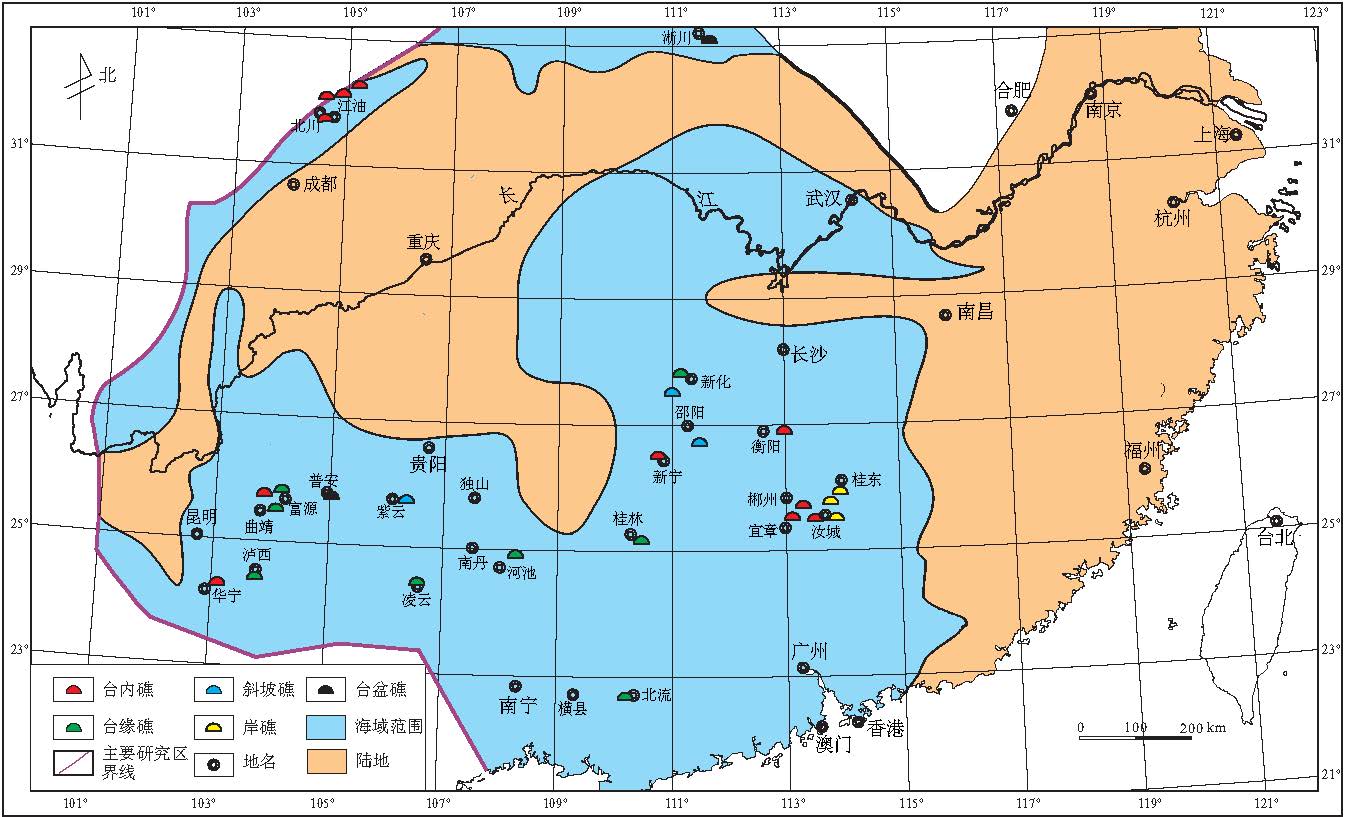 | 图5 华南晚泥盆世弗拉期生物礁分布图Fig.5 Distribution of the Late Devonian Frasnian bioreefs in South China |
| 表3 华南晚泥盆世弗拉期生物礁分布点资料 Table 3 Data of the Late Devonian Frasnian bioreefs in South China |
弗拉期华南新增生物礁分布点仍比较多, 但是, 吉维特期生物礁"大爆发"新增分布点很少被"继承"下来, 而且这些被"淘汰"的生物礁主要是台内礁.从图5可以看出, 弗拉期生物礁主要分布在滇中东部曲靖--富源一带, 贵州紫云, 普安等地区, 湘中南新宁--邵阳--新化一带, 湘东南宜章--汝城--郴州--桂东地区和川西唐王寨--甘溪--江油--马角坝地区.造礁生物主要为层孔虫, 其次为四射珊瑚和菌藻类, 在湘中, 黔南布寨等局部地区, 床板珊瑚也是重要的造礁生物, 附礁生物为枝状层孔虫, 菌藻类等.
除此之外, 弗拉期出现了一些"特色"生物礁.弗拉期晚期, 在南华海桂林地区海域开始出现菌藻礁(微生物礁), 主要造礁生物为菌藻类等微生物, 次为葵盘石, 其中后者多被前者缠绕粘结而共同构成礁骨架; 与此同时, 在秦岭海槽河南淅川地区首次发现了生物礁.
至今为止, 华南尚未发现法门期珊瑚层孔虫礁.但在弗拉期末期--法门期, 在桂林地区形成了菌藻礁富集区(殷德伟等, 1990a, 1990b), 同期在广西六景也有菌藻礁发育(陶业斌等, 1986; 张哲等, 2007), 这些菌藻礁一般分布在台地边缘--台前斜坡, 主要造礁生物为菌藻类.
虽然弗拉期古地理特征与吉维特期相似, 但是华南弗拉期生物礁面貌却发生了很大的变化:
1)吉维特期的生物礁富集区, 如湘中南, 黔南等地区, 在吉维特期末期--弗拉期由于台内礁的大规模衰退, 导致这些区域生物礁急剧减少, 这可能指示了吉维特期末期--弗拉期初期生物礁发生了一次大衰退.与台内礁相比, 台缘礁, 斜坡礁和台盆礁在吉维特期末期--弗拉期的减少趋势并不明显(图 6).总的来说, 从吉维特期到弗拉期, 生物礁体的规模明显减小.
2)弗拉期生物礁虽然总体上发生衰退, 但在湘东南宜章--汝城--郴州--桂东地区等吉维特期"无礁区", 在弗拉期岸礁和台内礁广泛发育, 形成了该期新的生物礁富集区.
3)菌藻类造礁作用进一步加强, 并首次出现了华南泥盆纪菌藻礁----广西桂林葵盘石菌藻礁; 与此同时, 层孔虫的造礁能力有所下降, 而造礁珊瑚已经发生了衰退, 代表了华南泥盆纪时期主要造礁生物首次由层孔虫和珊瑚向菌藻类微生物的转变(图 6).
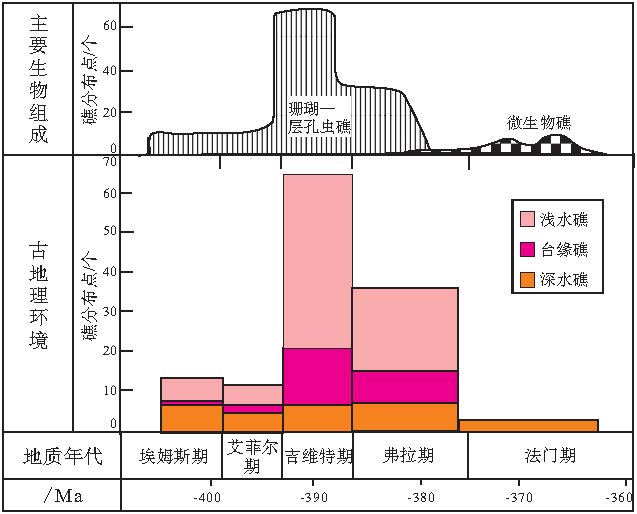 | 图6 华南泥盆纪生物礁类型和组成与古地理环境的关系Fig.6 Relationships between types and composition of the Devonian bioreefs and palaeogeographic environments in South China |
4)弗拉期末期, 华南生物礁遭到了毁灭性的打击----弗拉期--法门期(F--F)灭绝事件, 截至目前, 尚未在华南发现跨过F--F界线的泥盆纪珊瑚--层孔虫礁.
如果说吉维特期的古地理面貌促使了生物礁的广泛发育, 而弗拉期在继承了吉维特期古地理面貌的同时, 生物礁的面貌却发生了很大的变化, 这说明弗拉期生物礁有其独特的控制因素, 这主要表现在如下两个方面:
1)弗拉期是华南泥盆纪海平面上升到最高点的时期(龚一鸣等, 1997).海平面快速上升, 可能是导致弗拉期大范围台内礁和岸礁衰退(溺死)的最重要, 最直接的原因之一; 但是, 海平面上升促使部分局限台地海水循环更为畅通, 在一些"无礁区"(吉维特期)开始发育生物礁, 进而导致弗拉期出现新的生物礁富集区, 如湘东南宜章--汝城--郴州--桂东地区.
2)在以底栖固着生物为主的生态系中, 各种生物之间的竞争, 如对食物, 光线等的竞争, 主要表现为对空间的竞争, 而在礁生态系的"有限空间"内, 这种竞争在造礁生物之间表现尤为激烈, 在一定的环境条件下(如海水营养状况突变), 甚至会进入"你死我亡"的境地.因此, 晚泥盆世近岸海域海水恶化, 如营养盐含量增加, 海水富营养化, 菌藻微生物繁盛(Martin, 1996; 龚一鸣等, 2002, 2005; 徐冉等, 2006; Feng et al., 2010), 菌藻类造礁作用加剧(表 4), 可能是造礁层孔虫及珊瑚在吉维特期末期--弗拉期发生衰退和在弗拉期末期发生大灭绝的重要原因.
3)弗拉期南华海表层洋流流经的地方(杜远生等, 2009), 往往出现生物礁密集区, 说明洋流可能对生物礁有重要影响.
| 表4 华南泥盆纪主要造礁生物演化及对比 Table 4 Comparison and evolution of major bioreef-builders of the Devonian in South China |
华南泥盆纪生物礁可以分为4个演化阶段:
1)床板珊瑚礁初始发育期, 生物礁发育时限为埃姆斯期--艾菲尔期.这是中国南方泥盆纪的第一个成礁期.生物礁主要分布在南华海南部隆林--广南--富宁--德保--北流一带(图 3)和川西浅海龙门山地区.造礁生物以床板珊瑚为主, 次为层孔虫.
2)四射珊瑚--层孔虫礁"爆发"期, 即吉维特期.生物礁主要发育在南华海北部, 自西向东: 滇东曲靖, 沾益, 普安和富源等地--黔南桂北龙里, 独山, 南丹, 环江和桂林等地--湘中南城步, 隆回, 邵东, 新邵和涟源等地, 形成东西向展布的弧形带状生物礁富集区, 基本涵盖了南华海北缘(图 4); 与此同时, 川西浅海龙门山地区成为了生物礁富集区.主要造礁生物为层孔虫和四射珊瑚, 少量床板珊瑚(局部地区).
3)珊瑚--层孔虫礁衰灭期, 时限为吉维特期末期--弗拉期.此期基本继承了吉维特期的古地理格局, 生物礁发育的主要海域与吉维特期相似(图 5), 但生物礁面貌却发生了重大改变, 主要表现在吉维特期末期--弗拉期南华海台内礁大幅度衰退, 弗拉期末期珊瑚--层孔虫礁基本灭绝, 但在桂林出现了菌藻礁.造礁生物主要为层孔虫, 少量菌藻类和四射珊瑚.
4)菌藻礁广泛发育期, 时限为法门期.到目前为止, 在广西桂林地区(殷德伟等, 1990a)和横县六景地区(陶业斌等, 1986)已发现了菌藻礁富集区.
很明显, 在前3个阶段(埃姆斯期--艾菲尔期, 吉维特期, 弗拉期), 南华海泥盆纪生物礁分布层位由西南到北东逐渐升高, 很好地反映了整个海区由西南向北东侵进, 生物礁向北和北东方向迁移的过程; 与此同时, 生物礁的主要生物组成也呈现出有规律的变化.二者具体表现如下:
一是生物礁的古地理特征: 不同古地理单元生物礁分布点差异很大, 浅水礁(特指台内礁和岸礁)变化最大, 过渡带(特指台缘礁)变化次之, 深水礁(特指台盆礁和斜坡礁)几乎没有变化(图 6, 表5).因此, 从浅水到深水, 华南泥盆纪生物礁的"稳定性"增强; 浅水礁变化最大, 说明泥盆纪华南海域浅水区(主要是碳酸盐台地相)的环境变化大.
二是主要造礁生物为: 床板珊瑚(埃姆斯期)→ 床板, 四射珊瑚和层孔虫(艾菲尔期)→ 层孔虫和四射珊瑚(吉维特期)→ 层孔虫(弗拉期)→ 菌藻类微生物(法门期)(表 5).
| 表5 华南泥盆纪生物礁的时空分布格局 Table 5 Spatiotemporal distribution pattern of the Devonian bioreefs in South China |
泥盆纪埃姆斯期至法门期, 华南板块主要位于赤道附近, 隶属于热带--亚热带, 处于全球生物礁富集区(Copper and Scotese, 2003); 而华南板块内部特殊的构造背景导致了复杂古地理面貌的形成.二者使华南泥盆纪生物礁的时空分布具有"全球性"和"地方性"双重特征.
时间上, 生物礁在中泥盆世吉维特期"大爆发", 在吉维特期末期--弗拉期初期之间"大衰退"和在弗拉期--法门期(F--F)之交大灭绝, 这3个事件具有全球一致性(Copper, 2002a); 生物组成上, 珊瑚--层孔虫为主(埃姆斯期--弗拉期)→ 菌藻类微生物(法门期), 这与国外是可以对比的, 如澳大利亚西部Canning盆地(Shen et al., 2008)和加拿大西部(Copper, 2002a).
华南泥盆纪生物礁也极具"地方性"特色: 吉维特期生物礁大爆发是以台内礁(浅水礁)的发育为主要特征, 层状礁较为发育(单个礁体的厚度一般都不大); 礁垂向序列间歇性发育, 以多旋回为特征, 不少礁的相带分异不明显, 即生物礁特征"不典型"; 与国外同期相比, 如加拿大和俄罗斯, 华南泥盆纪生物礁规模总体偏小.
因此, 虽然华南泥盆纪生物礁的控制因素复杂, 但可以简单分为"全球性"和"地方性"(区域性)两大类控制因素.
通过对前人资料的总结和对比, 华南泥盆纪各个阶段生物礁主要"地方性"控制因素有很大的不同, 必须分阶段进行讨论, 这方面前文已经详细阐述, 在此仅着重讨论"全球性"控制因素.
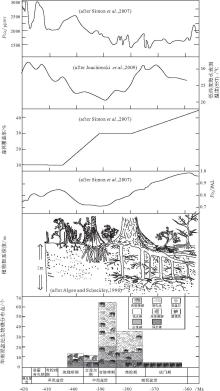
华南泥盆纪生物礁吉维特期的繁盛, 吉维特期--弗拉期之交大衰退以及弗拉期末大灭绝3次事件, 与全球泥盆纪同期其他重大地质事件在发生时间上恰好一致, 下面将主要讨论温度, 大气中CO2和O2的分压(
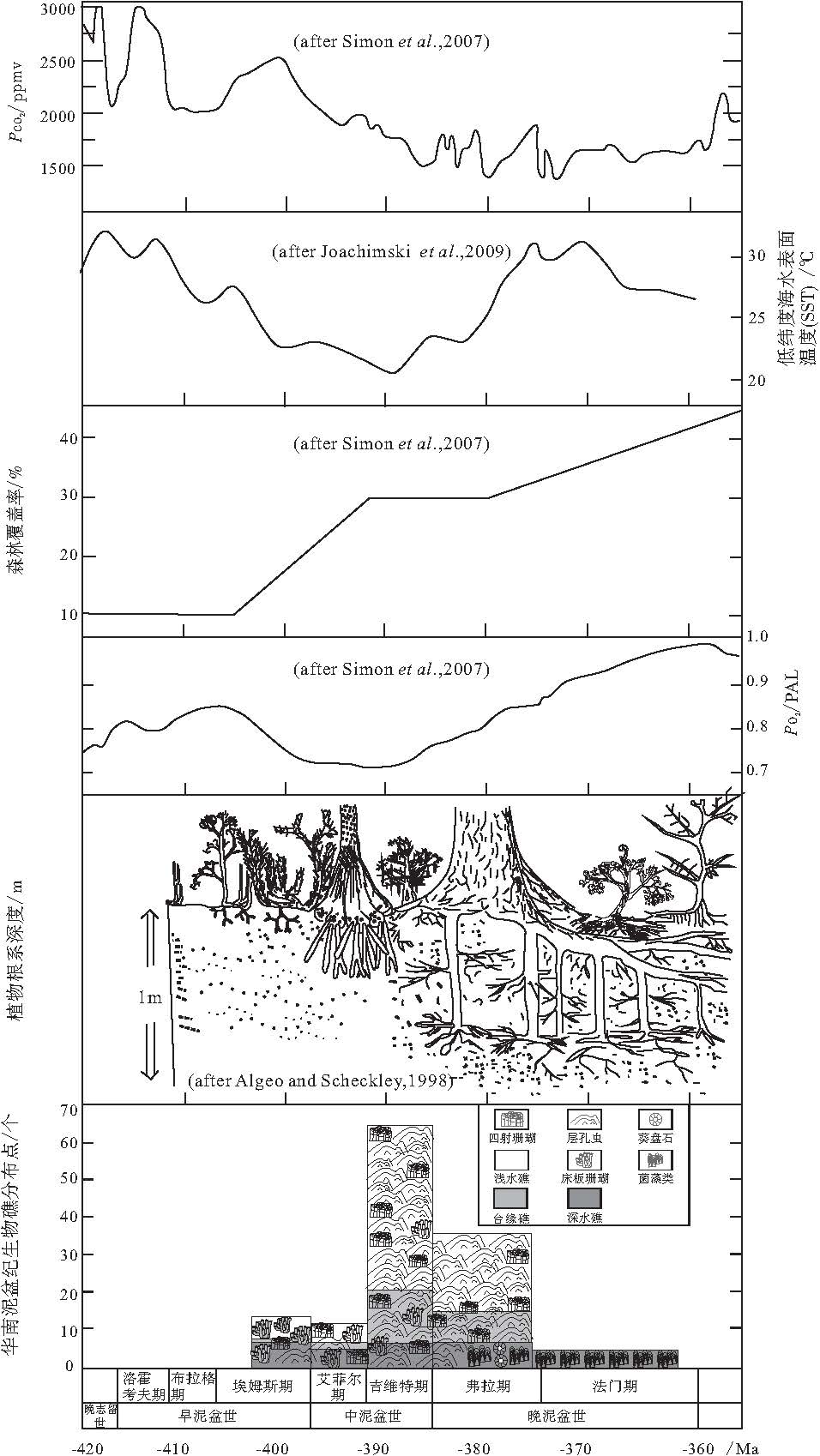 | 图7 华南泥盆纪生物礁演化与全球环境参数变化的耦合关系PAL--"present atmospheric level"的缩写, 即"现今大气中的含量水平"Fig.7 Sketch showing coupling relationships between evolution of the Devonian bioreefs in South China and changes of global environmental parameters |
5.2.1 温度
目前, 全球变暖引发的海水表面温度(SST)升高导致现代热带--亚热带海洋中珊瑚礁白化的现象受到了广泛的关注(Celliers and Schleyer, 2002; Douglas, 2003; Phinney et al., 2006; Crabbe and James, 2008).化石礁和其他浅海陆架碳酸盐沉积, 常被当作是地史时期热带和亚热带良好的指示标志(Copper and Scotese, 2003); 生物礁的纬度分布范围被认为能够指示古气候, 特别是古温度(Frakes et al., 1993; Johnson et al., 1996); 生物礁的组成也被作为古气候重建的重要指标(Kiessling, 2001; Joachimski et al., 2009).因此, 温度与生物礁的发育及其纬度分布密切相关, 是生物礁的重要控制因素.
根据腕足类壳体 δ 18O(Van Geldem et al., 2006)和牙形石δ 18O(Joachimski et al., 2009)资料, 对泥盆纪低纬度海水表面温度(SST)进行重建, 得到了相对一致的温度变化趋势: 早泥盆世(埃姆斯期)和晚泥盆世大多的时间内都是相对温暖的气候, 而中泥盆世明显为相对低温期(图 7).SST的变化趋势与华南泥盆纪生物礁的变化趋势关系密切: 生物礁在相对较低温时期繁盛, 而高温时生物礁发育不好或者发生大的衰退(图 7).
华南板块在泥盆纪(埃姆斯期--法门期)位于赤道附近, 隶属于热带(Copper and Scotese, 2003; 曾雄伟等, 2007); 而华南泥盆纪珊瑚--层孔虫礁和温度的关系, 与现代热带--亚热带珊瑚同温度的关系恰好一致, 即SST增高导致生物礁衰退.Talent(1988)和Copper(2002b)认为泥盆纪造礁珊瑚可能也有与现代造礁珊瑚相似的光养共生生物----动物黄藻(zooxanthellate).因此, 华南泥盆纪生物礁与温度之间的关系可与现代进行对比: 一定范围内(如热带--亚热带海域), SST降低会导致生物礁的繁盛(中泥盆世), 而SST的升高则抑制生物礁发育和生长(晚泥盆世), 进而导致生物礁衰退和灭绝.
值得注意的是, 现代礁型珊瑚发育的最佳温度范围是23~28℃ (Fagerstrom, 1987), 而艾菲尔期SST降低到23 ℃ 之下, 到吉维特期SST开始上升至23~25 ℃ (图 7), 这可能是华南艾菲尔期生物礁数量稍有降低和吉维特期生物礁"大爆发"的重要原因.
5.2.2 植被--森林生态系
众所周知, 森林是陆地上最重要的生态系统之一.从植物首次登陆到泥盆纪森林生态系统的形成, 植物在不断适应贫瘠的陆地环境的同时也对环境产生了缓慢而重要的改造作用, 进而对其他生态系统产生重大影响.作为海洋中"热带雨林"的生物礁生态系更是首当其冲, 在植物登陆至森林广泛出现的过程中, 发生了极大的变化.植被--森林生态系对生物礁生态系的影响可能主要表现在以下两方面:(1)植被--森林对大气组分有重要的影响, 进而对气候产生影响; (2)植被--森林对陆地风化作用的影响, 导致生物风化和化学风化作用加剧, 促进土壤的形成, 使大量营养物质流入海水当中, 进而改变海洋的营养状况.
早泥盆世植物群被称为工蕨(Zosterophyllum)植物群(Cai and Wang, 1995), 植被仅仅局限分布在潮湿地区, 无法摆脱对水的依赖, 植被覆盖率小.植物的主要特点: 个体小, 无根或者根系发育极差(Algeo and Scheckler, 1998), 叶子发育也很差, 茎叶未分化.植被对土壤的形成, 陆地风化作用以及大气组分的改造是极其有限的.因此, 该期植被对生物礁生态系的影响也不大.
中泥盆世植物群被称为原始鳞木(Protolepidodendron)植物群(Cai and Wang, 1995; 蔡重阳, 2000).植物开始出现比较深的根系, 对水的依赖性减小, 并开始能够摆脱水而独立生存了, 但仍只能生活在近水陆地区; 根茎叶等器官的分化也在这个时期完成(杨楠和李承森, 2009); 相对于早泥盆世, 植物的个体也有所增大.因此, 植被促进了土壤的形成以及陆地化学风化作用的进行; 树木叶子的光合作用对当时大气的组分产生了重大影响, 早泥盆世末期--中泥盆世
晚泥盆世植物群被称为薄皮木--古羊齿(Leptophloeum-Archaeopteris)植物群(Cai and Wang, 1995), 开始出现种子, 已经可以完全独立于水而生存, 能够在内陆地区发育和分布(Mosbrugger, 1990), 覆盖面积有了极大增加(图 7), 该时期真正意义上的森林开始出现.植物根系进一步发达, 促进了大陆的化学风化, 生物风化作用和土壤的形成, 大量的营养物质流入碳酸盐台地等近岸浅水区(Martin, 1996), 促使礁相菌藻类繁盛(Riding, 2006), 进而出现微生物礁, 极大地抑制了更喜欢寡营养环境的造礁层孔虫(Kershaw, 1998); 除此之外, 菌藻微生物繁荣并最终导致频繁的赤潮及海洋水团缺氧(龚一鸣等, 2002, 2005), 给宏体造礁生物带来了毁灭性的打击.因此, 在华南地区, 大量台内礁在吉维特末期--弗拉期之交发生了一次大衰退, 而且在F--F之交珊瑚--层孔虫礁发生了大灭绝(图 7).
近期的研究表明(曾剑威等, 2010): F--F之交在华南和世界范围内存在广泛的热液和火山事件, 该事件造成的海水酸化无疑会对晚泥盆世的海洋生态系造成重大创伤, 并直接导致钙质骨骼生物, 如层孔虫--珊瑚礁生态系彻底崩溃, 这一现象也是值得关注的.
综上可知, 华南泥盆纪生物礁与植被--森林生态系之间关系密切.这种关系可以简单地表示为以下两个方面:
1)早泥盆世--中泥盆世:植被繁盛----
2)中泥盆世--晚泥盆世:森林繁盛----化学风化, 生物风化作用和土壤的形成加剧----海水(近岸浅水区)中营养物质过剩, 菌藻类繁盛----珊瑚--层孔虫礁衰退.
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|