





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
河北平泉中元古界高于庄组生境型及古环境
[郭华1, 2  , 杜远生
, 杜远生1, 2 , 黄俊华1, 2 , 杨江海2 , 黄虎2 , 陈玉2 , 周瑶2 ]
, 杜远生, 黄俊华]
|
|

第一作者简介:郭华,女,1986年生,博士研究生,主要从事沉积学研究.通讯地址:中国地质大学(武汉)地球科学学院;邮编:430074;E-mail: didaguohua@163.com.
通讯作者简介: 杜远生,男,教授,从事沉积地质学研究.E-mail: dxyyz@cug.edu.cn.
河北平泉中元古界高于庄组海相碳酸盐岩地层中发育大量不同形态类型的微生物岩,主要包括柱状叠层石,层纹状叠层石,核形石,纹理石以及凝块石,它们各自不同的宏体和微观构造特征代表了不同的水体深度和环境水动力条件.通过野外实测观察,室内偏光显微镜下254块岩石薄片的微相分析以及碳,氧同位素的分析测定,研究了整个高于庄组沉积时期菌藻类生物的演化过程.在综合考虑岩性岩相特征,生物碎屑的含量,颗粒的保存状态以及微生物岩的宏体和微观特征等因素的基础上,将整个高于庄组划分为18个生境型,沉积相带演化从潮上带至内陆架的上部.河北平泉中元古界高于庄组地层序列中菌藻类生物含量的变化以及碳同位素的迁移规律清楚地记录了高于庄组沉积时期大规模的海进--海退的旋回沉积过程以及短期内频繁的海平面升降变化.通过分析生境型在地层剖面的分布特征以及生物与环境之间的相互作用关系,为今后研究生物与环境之间的协同演化以及更有效地进行烃源岩的评价提供了良好的基础资料.
About the first author Guo Hua, born in 1986, is a Ph.D. candidate of palaeontology and stratigraphy, and is engaged in sedimentology. Corresponding address: Faculty of Earth Science, China University of Geosciences(Wuhan); Postcode: 430074; E-mail: didaguohua@163.com.
About the corresponding author Du Yuansheng is a professor and he is engaged in sedimentology. E-mail: duyyz@cug.edu.cn.
Various types of morphological microbialites were developed in the marine carbonate rocks of the Mesoproterozoic Gaoyuzhuang Formation in Pingquan of Hebei Province. They mainly include five different types, namely, columnar stromatolites,laminated stromatolites, oncolites, biolaminates and thrombolites, which are characterized by their own special macro- and micro-structures, and thus can be taken as an indicator of varied water-depth and depositional environments. In this paper, we have conducted the field measurement of the Gaoyuzhuang Formation, analyzed 254 thin sections and C-O isotopes and made researches on the evolution processes of microbe and algae during the depositional period of the Gaoyuzhuang Formation. On the basis of lithologic facies features, the bioclastic magnitude, the grain preservation situation and the macro- and micro-characters of various microbialites, the whole Gaoyuzhuang Formation was divided into 18 habitat types, mainly occurring from supertidal to the upper shelf settings. The changes in bioclastic magnitude and C-isotope in the Gaoyuzhuang Formation illustrate clearly a transgression-regression cycle. Analyses of distribution feature of habitat types in the depositional sequence and the interaction between organism and environment have provided basic materials for further study of the coordinated evolution of organism and environment, and better evaluation of hydrocarbon source rocks.
对前寒武纪, 特别是中新元古代生命与环境协同演化的研究始终是地质科学中一个古老而引人入胜的命题(孙枢, 2005).近年来, 一些学者通过对微体化石(Lee and Golubic, 1999; Lee et al., 1999; 梅冥相, 2007), 微生物形成的各种原生沉积构造(Porada and Bouougri, 2007), 生物标志化合物(Peng et al., 1998; Li et al., 2003), 同位素(Chu et al., 2007)以及微量元素(Azmy et al., 2009)的分析, 在前寒武纪关键地层研究中取得了突破性进展, 为进一步探讨地球早期生命演化与环境的相互作用关系奠定了基础.
正如2003年起由Blackwell出版的Geobiology杂志上所列举的, 地球早期生命起源演化以及生物与环境相互作用关系的研究, 也是当前地球生物学领域一个重要的研究方面(Yin et al., 2008).作为地球科学与生物学相结合的学科领域, 它强调生命系统和地球系统的相互作用, 包括生物与环境相互作用的机制, 过程和演化历史等(Bottjer, 2005; 谢树成等, 2006).生境型是地球生物学研究中广泛应用的一个术语, 是确定地球生物相的主要参数之一.关于生境型的概念和系统的描述在殷鸿福等(1995)的相关专著中已有述及, 从潮上带到深海, 根据水深, 距海岸线的距离或者在陆架--斜坡--盆地体系中所处的位置划分为7个主要的生境型和25个亚型(Yin et al., 2008).通过深入研究生境型在地层剖面上的分布特征, 不仅可以提供生物和环境协同演化研究的基础资料, 而且对进一步恢复沉积有机质和埋藏有机质进而确定烃源岩和优质烃源岩的分布, 也具有重要的应用价值(杜远生等, 2007).作者旨在通过研究河北平泉高于庄组的生物地层序列, 菌藻类生物的演化过程以及碳同位素迁移规律, 确定高于庄组主要的生境型和沉积环境, 以期为研究中元古代生物与环境之间协同演化的关系提供一些有价值的信息, 并为将来有效地进行烃源岩的评价提供重要约束.
 | 图1 河北省承德市平泉剖面位置Fig.1 Location of Pingquan section in Chengde of Hebei Province |
研究区位于河北省承德市平泉县境内(图 1), 大地构造位置属于华北克拉通北缘, 是中国中上元古界发育较为完整的地区之一.吕梁运动之后, 华北克拉通构造体制发生了根本变化, 从碰撞造山转向大规模的伸展裂陷阶段, 在基本固结和刚性化之后趋于稳定的华北克拉通太古界基底之上, 自下而上不整合沉积了长城系, 蓟县系和青白口系的巨厚盖层, 总厚度约10 000 m(Wan et al., 2003).
河北平泉剖面中元古界发育良好, 自南东向北西依次出露常州沟组--雾迷山组, 各组之间均为整合接触, 沉积岩石序列为碎屑岩--黏土岩--碳酸盐岩(河北省地质矿产局, 1989).传统概念中高于庄组一直是长城系最顶部的一个组, 随着高精度锆石SHRIMP 定年技术在华北前寒武纪地层学中的成功应用, 在几个关键层位定年研究中取得了实质性进展(高林志等, 2007, 2008; 乔秀夫等, 2007).根据已有的资料, 将高于庄组从原来的长城系的顶部调整到了蓟县系的底部, 并将其年龄限定在1600--1500, Ma之间, 使得华北中元古代地层格架发生了较大变化(乔秀夫等, 2007).高于庄组为一套近千米厚的海相碳酸盐岩沉积, 根据其岩性特征及生物--沉积构造等将其分为4段:1段底部为薄层状褐黄色砂质板岩, 泥质板岩, 向上渐变为含硅质白云岩, 藻纹层白云岩及厚层泥晶白云岩, 厚约130, m.2段由纹层状微晶白云岩, 含硅质条带白云岩, 薄层钙质泥岩, 泥质白云岩组成, 局部藻纹层发育.该段泥质含量自下而上逐渐增多, 表现为一个海进的沉积过程, 水体向上逐渐加深, 厚度为245, m.3段表现为中厚层状含硅质条带微晶白云岩, 薄层钙质泥岩, 泥质页岩的韵律式沉积特征, 局部见有柱状叠层石, 厚度为323, m.4段以厚层状细晶白云岩, 藻纹层白云岩, 砂质白云岩为主, 夹少量硅质团块和硅质条带, 上部帐篷构造发育, 厚约245 m.
微生物岩是由各种底栖微生物群落与环境相互作用形成的生物沉积岩(Burne and Moore, 1987).各种微生物特别是细菌和蓝细菌等通过自身生长, 新陈代谢活动引起沉积质点粘结和圈捕, 和/或表面矿物沉淀, 和/或生物矿化作用等方式形成各种不同的生物沉积构造(Wang et al., 2005; Bosak et al., 2009), 而这种具有特殊形态的生物建造在前寒武纪以微生物为主导的地层中尤为发育(Noffke, 2009).
据梅冥相(2007)新修订的微生物岩的划分方案, 将其分为叠层石, 凝块石, 核形石, 均一石, 树形石和纹理石6大类.河北平泉高于庄组微生物岩分布广泛, 类型各样, 由微生物在不同沉积环境中通过不同机制形成的微生物岩表现出各种不同的宏体形态和微观构造特征, 对微生物岩沉积特征的分析将对生境型的划分具有重要的指示意义.
地质历史时期, 叠层石几乎占据了地球上每个主要的生态领域(Riding, 2000), 而尤以元古代时期发育最广, 数量最丰富, 曾被称为元古代微生物碳酸盐岩中的"标准化石"(曹瑞骥和袁训来, 2006).对叠层石生长模式的研究表明, 生物群落的演替对叠层石纹层形态的发育有重要影响, 而环境条件的变化则可能是控制叠层石宏体形态发生变化的最主要的因素(Peter and Mikhail, 2001; Andres and Reid, 2006; Dupraz et al., 2006), 不同形态的叠层石中往往蕴含着不同的环境信息.在河北平泉高于庄组近千米厚的海相碳酸盐岩中, 主要分布有两种形态的叠层石----柱状叠层石和层纹状叠层石.
 | 图2 河北平泉地区高于庄组微生物岩的主要类型 A, B--高于庄组3段下部柱状叠层石, 野外照片, 其中B图中箭头所指为柱状叠层石的檐; C--高于庄组4段层纹状叠层石, 野外照片; D--高于庄组2段顶部核形石, 内部发育致密的微波状细纹层理, 野外照片; E--高于庄组2段纹理石, 内部纹层多弯曲, 单层厚度约0.5~1.5, mm, 野外照片; F--高于庄组2段底部凝块石, 野外照片, 箭头所指为富含有机质的凝块状构造Fig.2 Main types of microbialites of the Gaoyuzhuang Formation in Pingquan of Hebei Province |
柱状叠层石 叠层体为不分叉的次圆柱状, 轮廓清晰, 单体或相互平行排列, 直立生长, 柱体直径4~6, cm, 高8~14, cm, 某些叠层石有明显的檐.层理薄而密集, 厚度为0.5~1, mm, 在柱体中心或边缘部分厚度无明显变化, 呈平缓穹形或半球穹形, 接近柱体边缘明显向下弯曲, 具有显著的壁(图 2-A, 2-B).根据对叠层石形成机理的研究, 通常认为其形成与蓝绿藻和细菌微生物的生理及生化作用有关(Takeshi and Hideyuki, 2006), 多产于安静舒适透光性较好的潮下带(Wang et al., 2009).河北平泉柱状叠层石发育的层位, 其围岩为具水平微波状层理的泥晶白云岩, 含少量陆源泥砂质成分, 生物碎屑含量较少, 为水动力条件较弱的局限潮下带环境, 主要发育于高于庄组3段中下部.
层纹状叠层石 中新元古代层纹状叠层石多半形成于浅水的周期性暴露或半暴露的环境(曹瑞骥和袁训来, 2006), 叠层体由较平整或略有起伏的细纹层理叠置而成, 层理厚1~2 mm(图 2-C), 显微镜下显示为明暗相间的纹带状构造, 亮带主要由细晶白云石组成, 暗带则为富含有机质的纹层, 由于前寒武纪微生物岩中钙化作用较弱, 菌藻类生物化石保存较差(图 3-A).这种层纹状叠层石多出现在高于庄组4段, 且常与帐篷构造密切共生, 为潮上带--潮间带上部环境的标志.
核形石作为另一类重要的微生物岩在前寒武纪以及显生宙重大地质转折期较为发育(Peryt, 1981; Whalen et al., 2002; 梅冥相, 2005), 主要以规则--半规则的球状或椭球状密集产出, 直径有大有小, 从几毫米到数十厘米.已有的研究表明, 核形石的形态和分布特征主要受当时海平面变化及沉积速率等环境条件的制约(Vé drine et al., 2007), 因此, 其形态学特征能够有效地指示其形成环境.
这类微生物岩在河北平泉剖面高于庄组2段顶部较为发育, 主要以较为规则的球状和椭球状形态产出, 垂向厚度约1.5 m.颗粒直径多在2~4, cm之间, 内部发育致密的水平微波状细纹层理.在岩层层面上, 核形石内部纹层在相互临近的颗粒之间似乎表现为连续分布的特点(图 2-D), 颗粒之下围岩的沉积纹层多被压弯而不穿过核形石颗粒, 说明它们与围岩是同时形成的, 为原生沉积组构(王尚彦等, 2002).根据Vé drine 等(2007)以及Whalen等(2002)对不同粒径和形态的核形石形成环境条件的分析推测, 高于庄组3段底部发育的大型核形石应代表最大海泛期深水沉积的标志.核形石发育的层位, 其围岩为泥质含量较高的细晶白云岩, 局部见有浪成交错层理, 为海侵背景下内陆架环境中形成的一套微生物碳酸盐岩沉积.
纹理石的概念最早由Gerdes和 Krumbein(1987)提出, 用来描述有微生物活动参与的, 具有纹层构造的, 富含有机质的生物沉积岩, 其主要通过微生物的障积, 捕获和/或粘结沉积物颗粒形成, 代表了未受改造的席底构造(Noffke et al., 2002; 段凯波等, 2008), 多形成于微生物席发育的浅水潮间带--潮下带上部环境(Bouougri and Porada, 2007).
在河北平泉地区, 纹理石主要见于高于庄组2段中部, 纹层多弯曲, 单层厚度约0.5~1.5 mm(图 2-E), 显微镜下为具有明暗相间的纹层状构造, 暗带为藻迹, 藻斑点构成的有机质纹层, 亮带主要是一些细晶的碳酸盐岩颗粒(图 3-B), 与现代潮坪区环境中形成的纹理石相似(Noffke et al., 2002), 为潮间带--浅潮下带环境的标志.
凝块石以具有肉眼可见的斑块状构造而与显微形态的凝块状组构相区别, 类似于"凝聚颗粒"中的"团块"(Flü gel, 2004).河北平泉剖面凝块石仅在高于庄组2段最下部相应层位出现, 大小多为毫米级, 形态不规则, 由暗色富有机质的隐晶质所构成(图 2-F), 菌藻类生物化石呈散布的球粒状或相互粘结成凝块状(图 3-C), 凝块石之间为泥晶胶结, 组构贫乏.根据对现代凝块石形成环境的研究, 其形成的水体深度通常大于叠层石, 一般形成于潮下带环境(Feldmann and Mckenzie, 1998; 刘建波等, 2007).
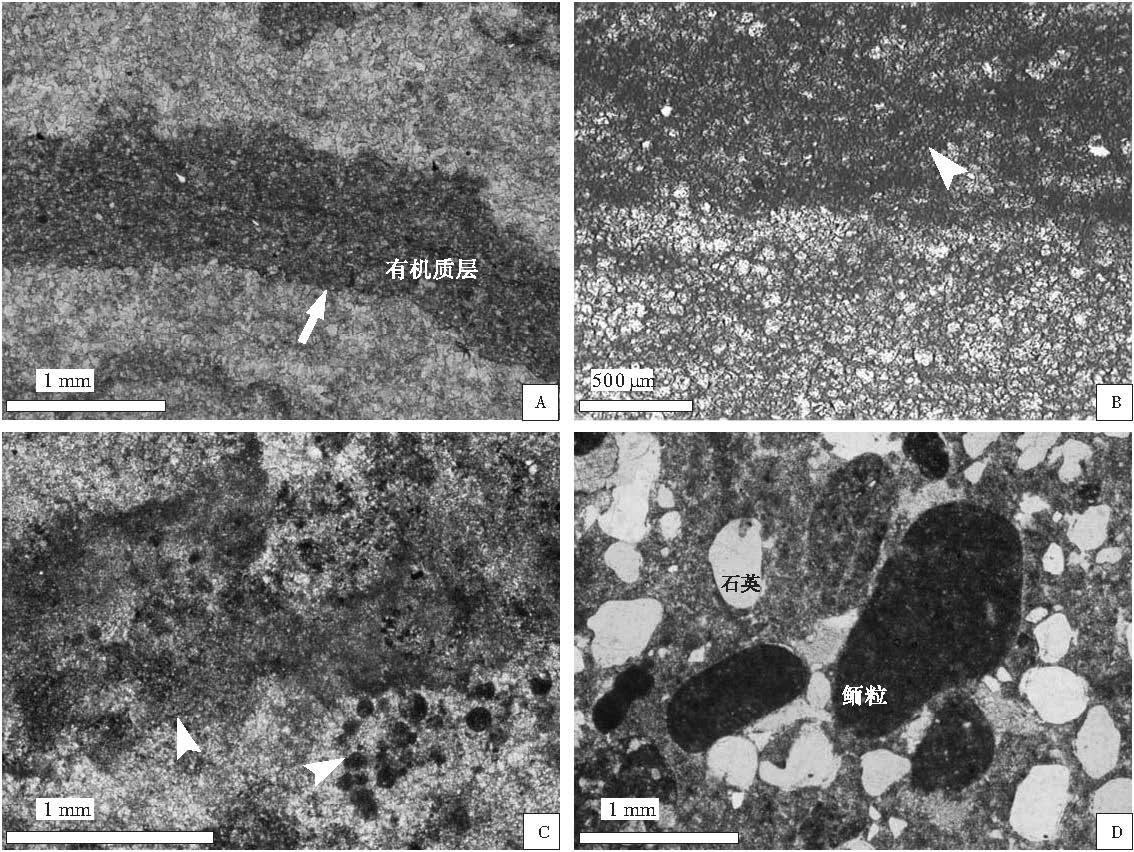 | 图3 河北平泉剖面中元古界高于庄组碳酸盐岩的微观特征 A--单偏光镜下层纹状叠层石中明暗相间的纹带状显微构造; B--单偏光镜下纹理石中明暗相间的纹层构造, 暗带为藻迹, 藻斑点构成的有机质纹层, 亮带主要是一些微晶的碳酸盐岩颗粒; C--单偏光镜下凝块石中保存的菌藻类生物化石; D--单偏光 镜下高于庄组1段碳酸盐岩显微构造Fig.3 Microphotographs showing microcosmic characters of carbonate rocks of the Mesoproterozoic Gaoyuzhuang Formation at Pingquan section in Hebei Province |
生境型的划分是建立在生物群落结构分析的基础之上的, 主要通过分析群落中的优势种, 特征种, 生物组合或生物之间的依存关系, 恢复生物生活的环境, 即生境型(杜远生等, 2007; Yan et al., 2008).关于生境型的划分, Boucot(1981)以底栖生物组合划分了6 个不同水深的典型的生境型, 殷鸿福等(1995)对Boucot 的生境型作了进一步细分, 将生境型划分为7 类25个亚类.河北平泉高于庄组沉积时期因缺乏特征的生物建隆及对应的礁前斜坡而形成以碳酸盐缓坡为特色的生境型模式(图 4).
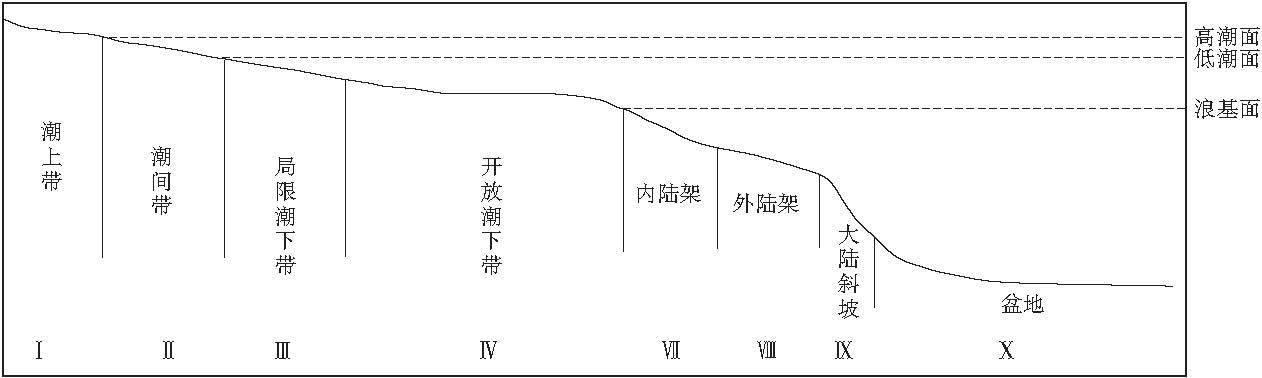 | 图4 河北平泉高于庄组生境型模式图(据杜远生等, 2007)Fig.4 Models showing habitat types of the Gaoyuzhuang Formation in Pingquan of Hebei Province(after Du et al., 2007) |
中元古代地层中缺乏后生高等生物及钙化的微生物, 生物类群以低等的菌藻类为主(Li et al., 2003; Sharma and Sergeev, 2004), 在这套以微生物建造为基础的地层单元中, 通过传统的生物类群结构分析方法恢复生境型是非常困难的.前人研究表明, 在正常浅水碳酸盐岩中, 一般从浅水到深水, 颗粒含量呈递减趋势变化, 因此, 岩石薄片中颗粒的含量可以作为判断生境型的一个有效的辅助方法.文中通过对河北平泉剖面野外实测观察, 在室内偏光显微镜下对254块岩石薄片进行了碳酸盐岩微相分析, 研究了高于庄组沉积期菌藻类生物的演化过程, 综合分析地层剖面中颗粒的含量及保存状态, 微生物岩类型, 伴生的沉积构造等因素, 将整个高于庄组划分为18个生境型, 沉积相带从潮上带至内陆架均有分布, 并显示出各自不同的特征(图 5).
高于庄组1段底部是在继大红峪组粉红色石英砂岩基础上形成的一套薄层砂泥质板岩, 变质石英砂岩, 厚约3.5 m.菌藻类生物难以鉴别, 且含有较多陆源泥砂质成分, 推测高于庄组沉积初期为受潮汐控制较弱的潮上带上部环境.向上逐渐过渡为藻纹层发育的细晶白云岩, 含砂质白云岩, 颗粒类型以鲕粒, 藻团粒为主, 含量约20%~30%, 亮晶胶结(图 3-D).陆源石英颗粒磨圆好, 分选较差, 局部见有鸟眼构造, 为沉积间歇暴露标志.综合岩性, 岩相和生物化石特征等因素, 推测为浅水动荡的潮间带上部环境, 相当于生境型Ⅱ 上部.
高于庄组2段以深灰色中厚层--薄层状的藻纹层微晶白云岩, 薄层钙质泥岩, 泥质白云岩的韵律式沉积为特征, 表现为总体海进, 并伴随有短期频繁的海平面升降变化.自下而上包含5个生境型:最下部是以发育凝块石为特征的含藻迹, 藻斑点的泥晶白云岩, 显微镜下藻丝体相互粘结形成凝块状或明暗相间的层纹状组构, 波状层理发育, 菌藻类含量为10%~30%, 为浅潮下带环境, 相当于生境型Ⅳ 上部; 向上逐渐被钙质泥岩, 泥质白云岩相所取代, 纹层以平直为主, 生物碎屑稀少, 为水流不畅的局限潮下带环境, 生境型为Ⅲ ; 之后海平面下降, 形成代表潮间--浅潮下带沉积的纹理石灰岩; 该段沉积中后期, 海平面上升, 沉积相带向深水区过渡, 至2段沉积末期, 基本演化为波状层理发育的泥质白云岩, 含核形石细晶白云岩, 大量菌藻类生物已基本销声匿迹, 生境型从最初的浅潮下带演化为内陆架环境.
高于庄组3段的下部继承了前期深水沉积相带的特征, 岩性表现为薄层泥质白云岩, 钙质泥页岩夹泥质透镜体, 菌藻类含量极少或无, 发育浪成交错层理(图 6-A), 为内陆架上部环境; 之后海平面下降, 生态环境逐渐适宜菌藻类的生长, 在生物灭绝之后首先繁盛起来的是建造叠层石的生物体, 在该层位发育柱状叠层石(图 2-A, 2-B), 根据前述分析推测, 沉积环境为局限潮下带上部, 相当于生境型Ⅲ ; 之后海平面频繁波动, 以竹叶状砾屑白云岩与水平纹层泥质白云岩的韵律式沉积为特征, 竹叶状砾屑颗粒长约2~4, cm, 分选, 磨圆较差, 形状不规则, 颗粒近于平行层面排列(图 6-B), 为深潮下带环境中风暴扰动作用形成(丁玉等, 2008); 泥质白云岩水平层理发育, 颗粒含量少(图 6-C), 代表水动力条件较弱的局限潮下带沉积.该段共划分为8个生境型.
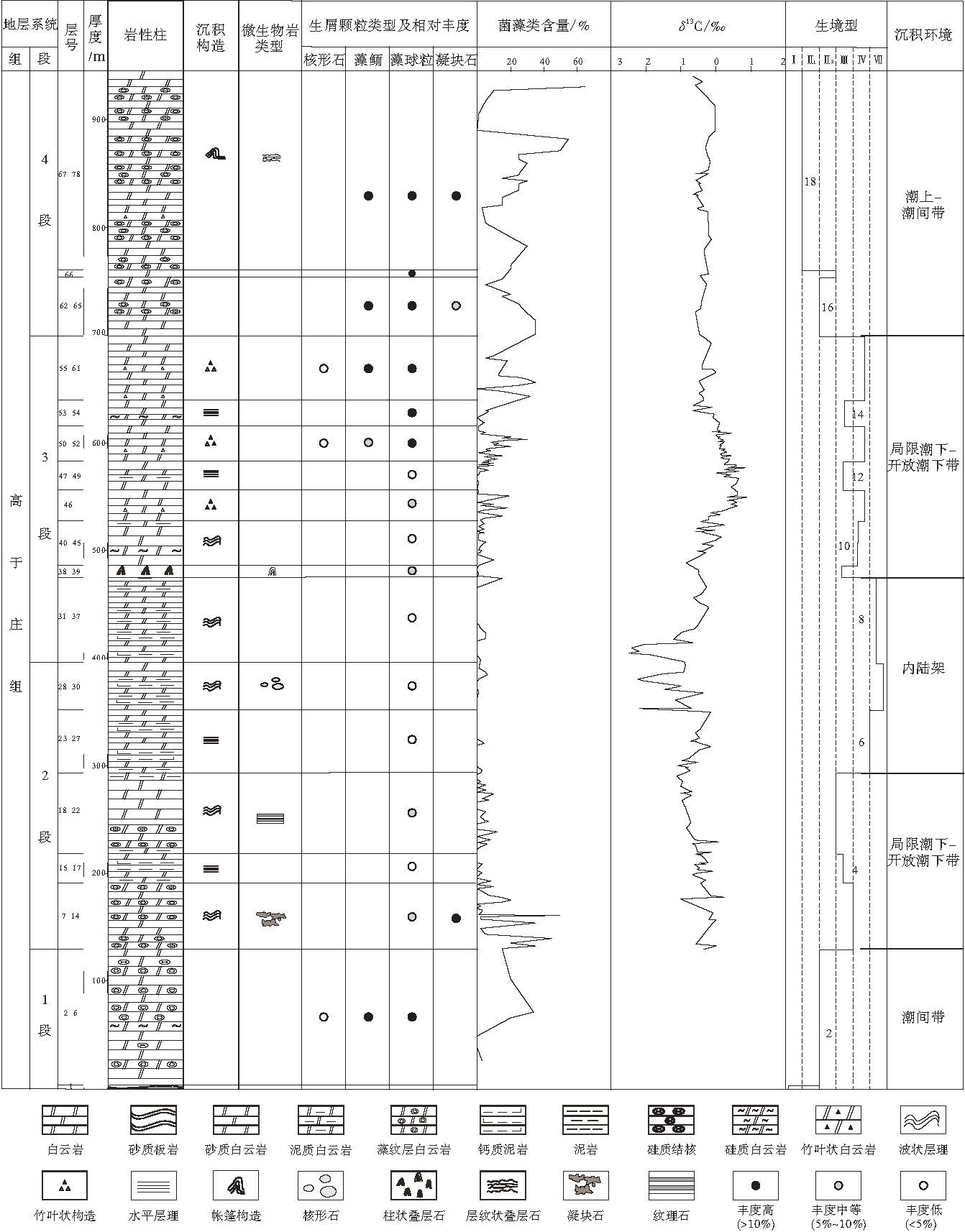 | 图5 河北平泉高于庄组生境型柱状图Fig.5 Stratigraphic column and habitat types of the Gaoyuzhuang Formation in Pingquan of Hebei Province |
 | 图6 河北平泉高于庄组碳酸盐岩的宏观面貌 A--高于庄组3段下部浪成交错层理; B--高于庄组3段上部竹叶状砾屑白云岩, 砾屑颗粒长约2~4, cm, 分选, 磨圆差, 近于平行层面排列; C--高于庄组3段上部泥质白云岩层; D--高于庄组4段帐篷构造Fig.6 Photographs showing macro characters of carbonate rocks of the Gaoyuzhuang Formation in Pingquan of Hebei Province |
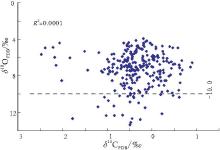
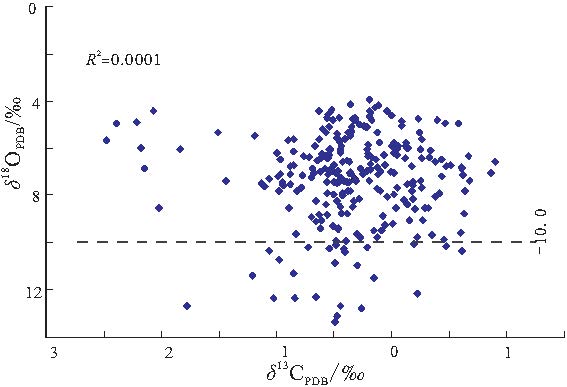 | 图7 河北平泉高于庄组碳酸盐岩δ 13C, δ 18O分布图Fig.7 Distribution of δ 13C and δ 18O values of carbonate rocks in the Gaoyuzhuang Formation in Pingquan of Hebei Province |
高于庄组4段总体代表了相对稳定的浅水潮间--潮上带沉积, 包含3个生境型.下部主要由中厚层状含砂质白云岩, 藻纹层发育的细晶白云岩组成, 局部夹石英砂砾岩层, 镜下观察显示, 颗粒以藻鲕, 凝块石为主, 含量15%~35%, 陆源石英分选, 磨圆较好, 其特征与高于庄组1段相似, 形成于水动力条件较强的潮间带环境; 中部为中薄层的细晶白云岩, 发育小型交错层理, 代表了短暂的海平面上升过程; 之后一直到高于庄组沉积末期, 以发育中厚层状的藻纹层细晶白云岩为特征, 部分层位见有竹叶状构造以及与周期性暴露作用有关的帐篷构造(图 6-D)(Kendall and Warren, 1987; 刘芊等, 2007), 代表潮上--潮间带沉积环境, 相当于生境型Ⅰ -Ⅱ .
一般来讲, 海相碳酸盐岩δ 13C值主要受海平面变化以及有机质埋藏通量的影响, 与生境型的演化关系密切, 是研究地质历史时期生物地球化学循环与环境演化的有效途径( 储雪蕾等, 2003; Dimitri et al., 2004; Roberto et al., 2004; Zuo et al., 2006).大量研究表明, δ 18O值受沉积期后大气和热水流体影响显著, 是成岩蚀变作用的灵敏指示剂.当δ 18O值不大于-10‰ 时, 其氧同位素较原始组成可能已发生明显改变(Kaufman and Knoll, 1995), δ 18O值在-5‰ ~-10‰ 之间的样品, 其氧同位素较原始组成可能稍有变化, 但碳同位素组成变化不大(Derry et al., 1994).研究样品的δ 18O值在 -13.3‰ ~-3.9‰ 之间, 主体分布区间为-4.0‰ ~-9.0‰ , 在利用样品测定的δ 13C, δ 18O值所作的散点图中(图 7), 碳, 氧同位素线性相关性较弱(R2=0.0001), 表明样品受后期成岩作用影响不大, 所得数据能够较为真实地反映原岩信息.
河北平泉高于庄组碳酸盐岩的δ 13C值变化区间为-2.48‰ ~0.9‰ , 主体分布于-1.0‰ ~0.7‰ , 平均值为-0.34‰ .从整个高于庄组来看, δ 13C值表现为"稳定--下降--上升--下降--稳定"的变化规律(图 5).高于庄组沉积早期, δ 13C值基本处于平均值附近(-0.3‰ ), 经历一段时期的稳定之后开始缓慢下降, 在高于庄组2段和3段交界处达到最低值-2.48‰ , 3段底部有一个快速上升的过程, 且变化幅度较大, 从-2.48‰ 急剧升高至 -0.6‰ , 并在此基础上缓慢向正值方向漂移, 至3段中部达到极大值0.9‰ , 随后又开始缓慢下降, 3段上部降低至-0.3‰ , 之后一直到高于庄组顶部, δ 13C值始终在该值附近波动.总体来看, 高于庄组2段δ 13C值处于缓慢下降趋势, 3段表现为"负--正--负"的波动规律, 高于庄组4段, δ 13C值一直在-0.3‰ 附近徘徊, 无明显的波动(图 5).
从图5中可以看出, δ 13C值的变化主要受菌藻类生物的繁盛程度以及当时海平面升降的制约, 与菌藻类含量变化曲线, 生境型曲线表现出较为明显的同步变化规律.高于庄组沉积早期, δ 13C值的相对高值很可能与菌藻类的大量繁殖和生长有关, 由于生物体从海水中选择性地吸收12C, 从而导致了海洋碳酸盐对重碳同位素(13C)的相对富集(Elrick et al., 2009); 高于庄组沉积中期, 海平面大规模上升, 相对较深的水体环境抑制了菌藻类生物的生长和繁衍, 生物产率降低, δ 13C表现为极低的负值; 而在高于庄组沉积后期, 相对较浅的海洋环境导致菌藻类生物的大量繁盛, 海洋生物生产力较高, 而δ 13C值相对于下部层位略微降低的现象可能与近岸环境中有机质周期性暴露与氧化分解有关.高于庄组δ 13C值的演化特征反映了高于庄组不同沉积时期的生物面貌以及海平面的变化, 是生物与环境协同演变研究的重要载体.
高于庄组沉积时期河北平泉地区处于大规模海侵开始背景下形成的陆表海环境(Li et al., 2003), 由于水面范围大, 地形切割不深, 径流补给量低, 又地处低纬度, 蒸发量大, 因此水体维持咸化, 生物群落为耐盐的低级菌藻类(孙淑芬, 2006).
文中通过对高于庄组岩相特征以及菌藻类生物群落兴衰演化的分析表明, 高于庄组沉积时期总体经历了一个海进--海退的沉积旋回, 并伴随着短期内频繁的海平面升降变化.这一认识与杨进辉等(1995)在平泉地区对高于庄组风暴岩的研究得出的结论一致.高于庄组沉积伊始, 河北平泉地区以发育大套的厚层藻纹层砂屑白云岩, 含沥青质白云岩及灰质白云岩为特征, 菌藻类生物繁盛, 碳酸盐岩颗粒类型以鲕粒, 藻团粒以及陆源石英为主, 颗粒磨圆好, 分选较差, 代表水动力较强的浅水环境, 为碳酸盐岩缓坡相区潮间带沉积.高于庄组大规模的海侵发生在2段沉积中后期和3段沉积早期, 伴随着海平面的缓慢上升, 菌藻类生物逐渐减少, 并在高于庄组最大海侵期发生灾变性灭绝, 地层中生物记录几乎为零.与此同时, 无机碳同位素也在海平面上升过程中表现出巨大的负向漂移, 至高于庄组2, 3段界线附近降至-2.48‰ , 成为沉积环境演变的又一个见证者.该阶段的地层岩相特征表现为薄层泥晶白云岩, 钙质泥岩, 泥质白云岩的韵律式沉积, 菌藻类生物逐渐衰退, 沉积相带从局限潮下带, 开放潮下带演化至内陆架上部, 代表一个向上变深的沉积旋回.自高于庄组3段沉积中期开始, 平泉地区又转而进入了一个海退的沉积阶段, 在海平面下降, 生态环境逐步适宜生物生存和繁衍的情况下, 叠层石的生物建造者代表了大灭绝之后首次繁盛起来的菌藻类生态系, 高于庄组3段地层中发育的柱状叠层石证明了这一点.高于庄组沉积中后期代表了菌藻类生物从复苏到繁盛的阶段, 温暖舒适的浅水环境为生物的繁衍和生长提供了良好的条件, 成为菌藻类生物发展的又一个"黄金阶段", 全球碳循环向平衡状态发展, 生态环境由潮下带逐步向潮间, 潮上带演化, 形成一个向上变浅的沉积旋回.
河北平泉中元古界高于庄组生物地层序列, 碳同位素的波动以及菌藻类生物含量的变化, 清楚地记录了高于庄组沉积时期大规模的海进--海退的旋回沉积过程以及短期内频繁的海平面升降现象, 自高于庄组沉积时期开始的陆表海环境, 经历了从潮间带至潮下带, 内陆架又至潮间带的韵律式沉积过程, 是一个以潮坪为主的陆表海沉积环境.
通过对河北平泉剖面高于庄组地层沉积序列特征, 菌藻类生物群落面貌, 碳同位素演变和沉积环境等方面进行详细研究, 获得了以下4点认识:
1)河北平泉高于庄组微生物岩类型多样, 以叠层石, 核形石, 纹理石, 凝块石为代表, 其中叠层石主要发育柱状和层纹状两种形态.它们各自不同的宏体和微观特征代表了不同的水体深度和环境水动力条件.
2)综合分析河北平泉剖面整个高于庄组的生物地层序列, 菌藻类生物的演化过程以及碳同位素迁移规律等因素, 将整个高于庄组划分为18个生境型, 沉积相带分布从潮上带至内陆架的上部.
3)高于庄组沉积时期海洋环境中菌藻类生物受海平面升降影响, 经历了从"繁盛--衰落--复苏--繁盛"的过程, 反映了生物与环境之间协同演化的相互作用关系.
4)河北平泉地区高于庄组沉积时期总体经历了一个海进--海退的沉积旋回, 并伴随着短期内频繁的海平面升降变化, 沉积环境演变由潮间带→ 潮下带→ 内陆架→ 潮下带→ 潮间带上部.
致谢 野外工作得到了冯庆来教授, 谢树成教授和史晓颖教授的悉心指导, 刘建波, 梅冥相两位评审专家为论文提供了部分参考文献和宝贵的建议, 在此致以诚挚的谢意!
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|