



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
从UB-2孔硅藻记录看末次冰期至早全新世日本海古海洋环境
[黄玥1  , 刘焱光
, 刘焱光2 , 蒋辉4 ]
, 刘焱光|
|

第一作者简介:黄玥,男,1980年生,博士,2009年毕业于华东师范大学地理系,现为云南大学讲师,主要从事硅藻与气候变化研究.通讯地址:云南省昆明市翠湖北路2号云南大学资源环境与地球科学学院;邮政编码:650091;联系电话:0871-4114918;E-mail:sohryu2016@hotmail.com.
日本海南部郁陵海盆的 UB-2孔沉积物硅藻记录,反映了 11000-- 28000 a BP 该区古海洋和古气候的变化.对比日本海和格陵兰冰芯( GRIP)δ 18O数据, UB-2孔硅藻暖水种比率及淡水--海滨种含量变化清楚地记录了氧同位素 3期( MIS3),末次盛冰期( LGM),波令--阿勒罗德暖期( B/A)以及新仙女木冷事件( YD)等古气候事件.淡水--海滨种硅藻含量在末次盛冰期阶段明显升高及暖水种数量的显著减少,表明此时日本海为低温,低盐的古海洋环境,这可归因于气候变冷,海平面下降造成的日本海当时相对封闭的海洋环境.自 15200 a BP 起,Paralia sulcata含量逐渐上升,这可能与海平面上升引起的古环境变化有关,可以作为对马海峡开启的标志.全球气候变化引起的海平面变化是该区域古气候变化的主要控制因素.
About the first author Huang Yue, born in 1980, doctor, graduated from the Department of Geography, East China Normal University in 2009. Now he is a lecturer at Yunnan University, and is mainly engaged in researches of diatom and global climate change. Address: School of Resources, Environments and Earth Science, Yunnan University, Kunming, 650091. Phone: 0871-4114918. E-mail: sohryu2016@hotmail.com.
A diatom record of core UB-2 from the Ulleung Basin in the southern part of Japan Sea reveals palaeoenvironmental changes through 11000--28000 a BP. Comparing with the δ18O records of Japan Sea and the Greenland icecore Project (GRIP), the Marine Isotope Stages 3(MIS3), the Last Glacial Maximum(LGM), the Bølling-Allerød warm phase(B/A)and the Younger Dryas cold event(YD)are clearly distinguished by the changes in percentage of warm water species and in abundance of fresh-littoral species. High abundance of fresh-littoral species during the LGM conforms a low temperature and low salinity palaeoceanographical environment due to the closed marine environment. Raising of the sea-level resulted in high abundance of Paralia sulcata from 15200 a BP, which refers to the opening of the Tsushima Strait. Climate-induced palaeogeographic variations may be the main contributor to the palaeoenvironmental changes.
日本海作为西北太平洋的一个边缘海, 是一个半封闭的海盆, 其独特的地理环境, 使得海平面变化对海洋环境变化的影响尤为剧烈.特别是在末次盛冰期(LGM), 日本海海平面降至现今海平面之下130 m(Itaki et al., 2004), 使其成为与外洋隔绝的封闭海盆, 形成与现代日本海大为不同的海洋环境.对末次冰期--冰消期日本海古环境和古气候记录的研究, 其意义远不限于解决其自身的古海洋学问题, 更可为认识东亚古环境和古气候在关键地质时段的演变提供重要依据.
日本海的古海洋学研究开始于上世纪中后期, 近10年来研究进展迅速.有关地球化学(Crusius et al., 1999; Tada et al., 1999; Khim et al., 2007), 稳定同位素(Gorbarenko and Southon, 2000; Dalai et al., 2005; Park et al., 2006), 海洋沉积记录(Bahk et al., 2000, 2001), 年代学(Park et al., 2003; Chun et al., 2007)和微体古生物(Itaki et al., 2004)等研究为重建晚第四纪特别是末次冰期以来日本海古海洋环境演化历史提供了详细的科学依据.
硅藻作为一种生活在有水或潮湿环境中的微体生物, 其种类, 数量及组合特征会因水体物理, 化学及水动力条件的差异有所不同, 故可以通过沉积物中硅藻化石的特征来推断相应的古海洋环境和古气候条件(Sancetta, 1982; Stoermer and Smol, 1999).许多学者在日本海曾开展过大量硅藻研究(Khim et al., 2005; Ryu et al., 2005, 2008; Koizumi et al., 2006), 确立了大量具有环境指示意义的硅藻属种, 为利用沉积物中的硅藻研究日本海古海洋环境提供了理论依据.
笔者尝试对日本海南部郁陵海盆UB-2孔活塞柱状样进行沉积物硅藻分析鉴定:(1)研究日本海UB-2孔末次冰期至早全新世硅藻组合面貌及特征种变化; (2)探讨硅藻数据与表层海水古温度, 古盐度之间的关系; (3)重建日本海UB-2孔末次冰期至早全新世古地理环境和古气候.
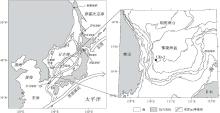
日本海介于32° 42'N至52° 14'N的中纬度地带, 呈不规则菱形, 东北窄, 中, 南部宽(图 1), 平均水深1350 m.日本海基本上以日本群岛与太平洋分隔, 通过宗谷海峡, 津轻海峡, 对马海峡和鞑靼海峡与外水域相通.这些海峡一般都狭而浅(< 130 m)(Itaki et al., 2004), 因此日本海与邻近海域的水体交换相当有限(崔琰琳等, 2006), 主要来源于黑潮暖流的对马暖流经对马海峡进入日本海, 是日本海尤其是南部海域表层和中层水体热量与盐分的主要来源.黎曼寒流则从鄂霍次克海经鞑靼海峡进入日本海.在二者的共同作用下, 日本海表层海水通常以逆时针方向流动(李志国, 1993).
 | 图1 日本海UB-2孔位置Fig.1 Position of core UB-2 in Japan Sea |
郁陵海盆是日本海南部的一个碗状的盆地, 西靠东朝鲜半岛, 北至朝鲜海台, 东接日本岛弧和隐岐海岭, 南邻对马海峡, 是对马暖流进入日本海后的必经之地.晚第四纪以来, 郁陵海盆上陆坡区频繁发生海底滑坡, 滑塌, 而碎屑流和浊流沉积等在下陆坡比较普遍(Lee et al., 1996).
日本海UB-2孔位于郁陵海盆西南侧(36° 33.6'N, 130° 11.6'E), 水深2165 m(图 1), 由韩国2002年"大地航次(R/V Onnuri)"获得.硅藻样品取自156~520, cm段, 其中156~200, cm段采样间距为2, cm, 200~520, cm段采样间距为8, cm, 共采集硅藻样品63个.
所有硅藻样品的处理均依据Hå kansson(1984)方法进行, 主要处理步骤如下:(1)去钙质.在装有硅藻样品的试管中加入浓度为10%~15%的稀盐酸, 待样品与盐酸初步反应后搅拌均匀并静置12~24 h, 然后用蒸馏水清洗3次.(2)去有机质.加入浓度为30%的双氧水, 待样品与双氧水初步反应后, 置于恒温(70 ℃ )水浴锅中加热1~2 h, 其后将样品从水浴锅中取出, 同样用蒸馏水清洗3次.(3)制片.用玻棒将样品均匀涂于载玻片上, 滴上Naphrax胶(dn=1.73), 盖上盖玻片, 然后在电热板上加热至150~200 ℃ , 待气泡排出后冷却, 保存于样品盒中.
硅藻鉴定与计数均在1000倍徕卡油镜下进行, 每个样品统计硅藻壳面数至少300个(不包括Chaetoceros休眠孢子).大部分硅藻鉴定到种, 少数到属.鉴定过程依据相关参考资料进行(Hustedt, 1959; Hendey, 1964; 布拉谢尔, 1986; 地质矿产部海洋地质综合大队, 1989; 郝诒纯等, 1989; Hasle and Syvertsen, 1997; Koizumi et al., 2006).
日本海的地理纬度较高, 碳酸盐补偿深度变化剧烈, 深海盆沉积物中钙质生物壳体难以很好地保存, 因此在日本海建立详细的地层年代框架相对困难.利用火山灰标志层和地层特征对比是常用的方法.
一方面, 由于日本海地区拥有大量火山, 火山灰广布于整个日本海区域, 这为沉积物年龄框架的建立提供了一个重要的指示标记.对郁陵海盆海洋沉积物的研究, 也多依靠火山灰层建立年龄框架(Park et al., 2003, 2006; Kim et al., 2006).
另一方面, 由于海洋沉积环境的频繁变化, 东亚地区气候的冷暖, 干湿变化影响到日本海的陆源物质输入和东海沿岸流的强弱变化, 导致日本海沉积物颜色呈现厘米--分米级厚度的浅色贫有机质纹层和深色富有机质纹层交替变化(Tada et al., 1992, 1999); 特别是一些深色纹层沉积(TL layers)在日本海半远洋沉积物中均有发现, 并与沉积时代有着很好的一致性, 这为沉积物年龄框架的建立提供了可靠的依据.在郁陵海盆, 也不乏利用纹层建立年龄框架的研究(Lee et al., 1996; Park et al., 2007; Khim et al., 2008a).
笔者根据U-Oki和AT火山灰层以及TL-1和TL-2深色纹层的发生和沉积年代, 建立了UB-2孔的年龄框架(表 1).
| 表1 UB-2孔火山灰层和深色纹层日历年龄 Table 1 Calibrated ages of tephra layers and TL layers in core UB-2 |
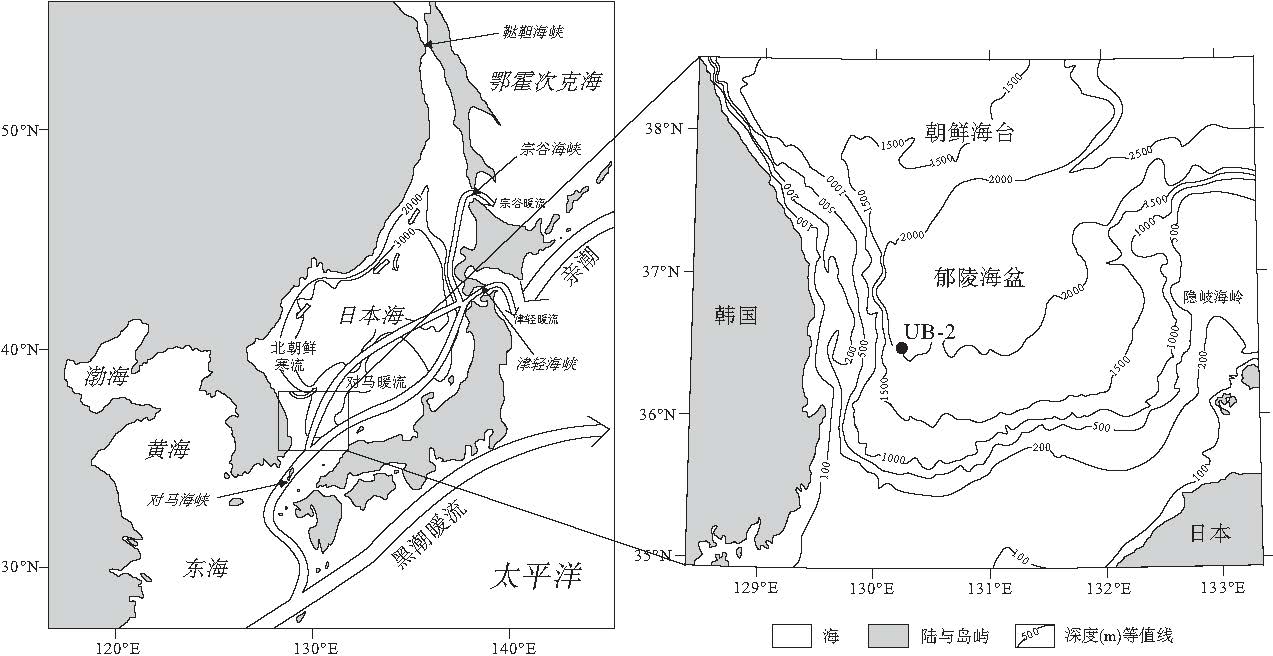
根据硅藻组合特征(图 2), 在240~440, cm段, 冷水种Rhizosolenia hebetata和Denticulopsis seminae, 热带海洋种以及暖水种Nitzschia interruptestriata和Fragilariopsis doliolus均有出现, 而相对难以溶解的R. hebetata和R. setigera在该层位大多以较强的硅化状态出现, 这说明本层沉积混合了来自更早的氧同位素3期(MIS3)的沉积物.聚类分析结果也显示该层位硅藻组合面貌与上, 下层位有较大的变化(图 2).韩国釜山大学海洋科学系Khim教授提供的本孔放射虫研究结果也表明, 在240~440, cm段出现大量暖水种(未发表数据), 故笔者认为该层位为浊流层, 即由上陆坡海底滑坡, 滑塌物质再沉积形成的层位.删除此层位后, 最终的深度--年代模式图详见图3.
 | 图2 日本海UB-2孔浊流层Fig.2 Turbidite layer of core UB-2 in Japan Sea |
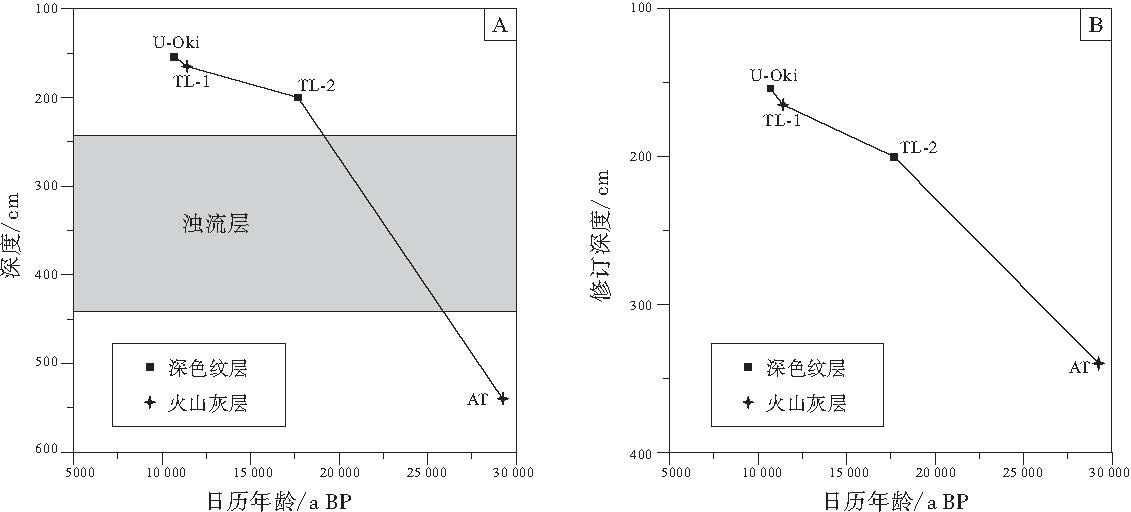 | 图3 日本海UB-2孔修订前(A)与修订后(B)深度--年代模式图Fig.3 Uncorrected(A)and corrected(B)depth-age diagram of core UB-2 in Japan Sea |
在UB-2孔63个沉积物硅藻样品中, 共鉴定出硅藻属种158个.其中Achnanthes spp., Cocconeis spp., Cyclotella spp., Diploneis spp., Denticulopsis seminae, Fragilariopsis doliolus, Paralia sulcata, Rhizosolenia hebetata, Thalassionema nitzschioides, Thalassiosira oestrupii是主要组成属种.硅藻组合以浮游种类为主; 淡水和海滨种变化明显; 寒带和热带浮游硅藻种属数量不多, 但都能很好地反映该区域古环境变化.
T. nitzschioides是UB-2孔沉积物硅藻中含量最为丰富的属种, 此种为世界性种, 除两极地区以外, 自赤道至高纬地区海洋均有分布(Hasle and Syvertsen, 1997), 且数量较大, 是常见的广布性浅海种类(Hasle, 1960).以该种为代表的大量浅海浮游硅藻种类聚集于UB-2孔, 这是因为日本海被日本岛弧环绕, 连接西太平洋和东海的海峡狭窄, 海水与周围海洋水体交换较弱, 水体性质相对均一.其变种T. nitzschioides var. parva通常被用作暖水指示种(Romero et al., 2005), 但在UB-2孔数量稀少(少于4%), 这与Tanimura(1999)研究结果一致.
Achnanthes spp.包括以A. lutheri和A. minutissima为主的若干种, 主要分布在河口, 海港等相对低盐度环境下(Hendey, 1964).在日本海海域, Koizumi等(2006)将Achnanthes spp.作为淡水硅藻.在UB-2孔, 该属主要集中出现在末次盛冰期(LGM).
Cocconeis spp.主要包括C. costata, C. disculus, C. peltoides和C. scutellum.据Hendey(1964)的研究, 其中C. costata和C. peltoides为北欧沿岸常见的海滨种, C. disculus则常出现于沿岸半咸水环境中, 而C. scutellum是世界广布性沿岸种, 在日本海将其归并为海滨种; Cyclotella spp.主要包括C. striata和C. stylorum; Diploneis spp.以D. bombus, D. oculata, D. smithii和D. weissflogii为主, Koizumi等(2006)认为它们是典型的海滨种, 生活在低盐环境下, 在日本海海域可作为海滨环境的指示.以上硅藻种属在UB-2孔广泛分布, 为主要的海滨种.
Jousé 等(1971)和Muhina(1971)分析了太平洋200个表层硅藻样品, 自北向南区分出7个硅藻组合, 其中, 热带硅藻组合和赤道硅藻组合可代表北赤道流和它的分支黑潮暖流的硅藻特征.这2个组合的主要成分在UB-2孔发现的有5种:Azpeitia nodulifera, A. neocrenulata, Hemidiscus cuneiformis, Alveus marinus和Rhizosolenia bergonii.尽管这些属种的绝对含量变化不大, 但却可以用于追寻外洋暖水以及黑潮暖流的踪迹(Jiang et al., 2006).故笔者将其合并后作为热带远洋种(Tropic oceanic species), 以指示外洋暖水的入侵.
F. doliolus与T. oestrupii生活于暖水环境下(Hasle and Syvertsen, 1997), 在日本海, 连同N. interruptestriata, 均为典型的海洋浮游暖水种(Koizumi et al., 2006).R. hebetata为北方冷水种(Hasle and Syvertsen, 1997), 连同D. seminae, Koizumi等(2006)将其作为典型的海洋浮游冷水种.在UB-2孔, 它们是代表表层海水温度的典型硅藻属种.
根据UB-2孔硅藻组合聚类分析结果, 可划分出3个硅藻组合带(图 4)(参照Grimm, 1987).由于240~440, cm段为浊流沉积, 故图4中删去了这一部分.
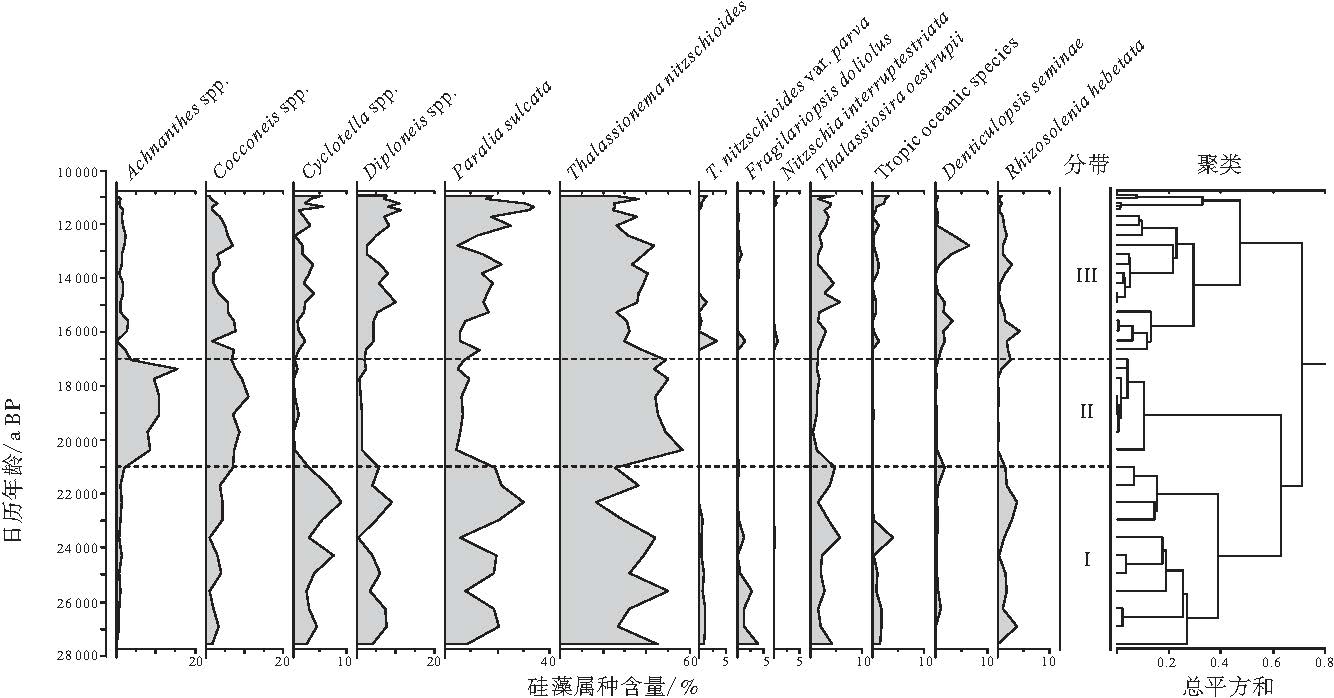 | 图4 日本海UB-2孔主要硅藻百分含量Fig.4 Percentage distributions of selected species and species groups of diatoms in core UB-2 in Japan Sea |
Ⅰ 带(28000--21000 a BP).本带海滨种Cyclotella spp.含量达到峰值, 另2个海滨种Cocconeis spp.与Diploneis spp.和半浮游种P.sulcata含量也较多, 而淡水种Achnanthes spp.含量较少.热带远洋种及暖水硅藻N. interruptestriata, F. doliolus, T. oestrupii和T. nitzschioides var. parva均有出现.这表明该时期处于一个相对温暖的阶段, 沿岸流较强, 海水盐度不高, 对应氧同位素3期(MIS3).
Ⅱ 带(21000--17000 a BP).淡水种Achnanthes spp.含量达到峰值, 海滨种Cyclotella spp.与Diploneis spp.含量有所降低.热带远洋种以及浮游暖水种N. interruptestriata与F. doliolus完全消失, T. oestrupii含量也降至最低, 该时期对应着一个低温低盐环境, 为末次盛冰期.由于海平面下降, 一方面使得UB-2孔位置与海岸线距离变近, 来自陆地的淡水更容易影响本区域, 造成淡水种含量上升; 另一方面, 低海平面也造成日本海与周边海域联系中断, 因而热带远洋种以及浮游种含量也随之降低.而浅海浮游种T. nitzschioides含量极高, 这也从另一个角度说明了此时为封闭的古海洋环境.
Ⅲ 带(17000--11000 a BP).淡水种Achnanthes spp.含量逐渐减少, 海滨种Cyclotella spp., Cyclotella spp., Diploneis spp.含量逐渐增多, 热带远洋种含量也逐渐增加, 表示本带对应着一个逐渐回暖的过程.在13400--12300 a BP, 冷水种D.seminae含量骤增(该种在末次盛冰期未见, 这是由于日本海较为封闭造成的), 相应的, 热带远洋种含量有所降低, 显示出这是一个突然降温的过程, 对应着新仙女木冷期(YD).从11300--11100 a BP, P.sulcata含量达到峰值, 明显反映了一次冲淡水事件.
根据Hendey(1964), Hasle和Syvertsen(1997), Koizumi等(2006)以及Jiang等(2006)对硅藻生态的研究, 作者在UB-2孔硅藻组合中挑选具有明显环境指示意义的硅藻属种, 并依照生态习性, 将其分为淡水种, 海滨种及浮游种, 而浮游种又可细分为浮游暖水种与浮游冷水种(表 2).
| 表2 UB-2孔具有环境指示意义的硅藻属种 Table 2 Ecological affinity of diatoms in core UB-2 |
浮游暖水种在浮游种中所占的比率是指浮游暖水种含量在所有浮游种(浮游暖水种含量+浮游冷水种含量)中所占的小数比, 将其作为海水温度变化的指示物(图 5-A).在28000--24000 a BP期间, 浮游暖水种比率较高, 随后迅速下降, 直至15000 a BP, 均维持在一个较低的水平.15000 a BP以来, 暖水种比率又逐渐上升, 在13400--12300 a BP经历了一个突然降低的过程.
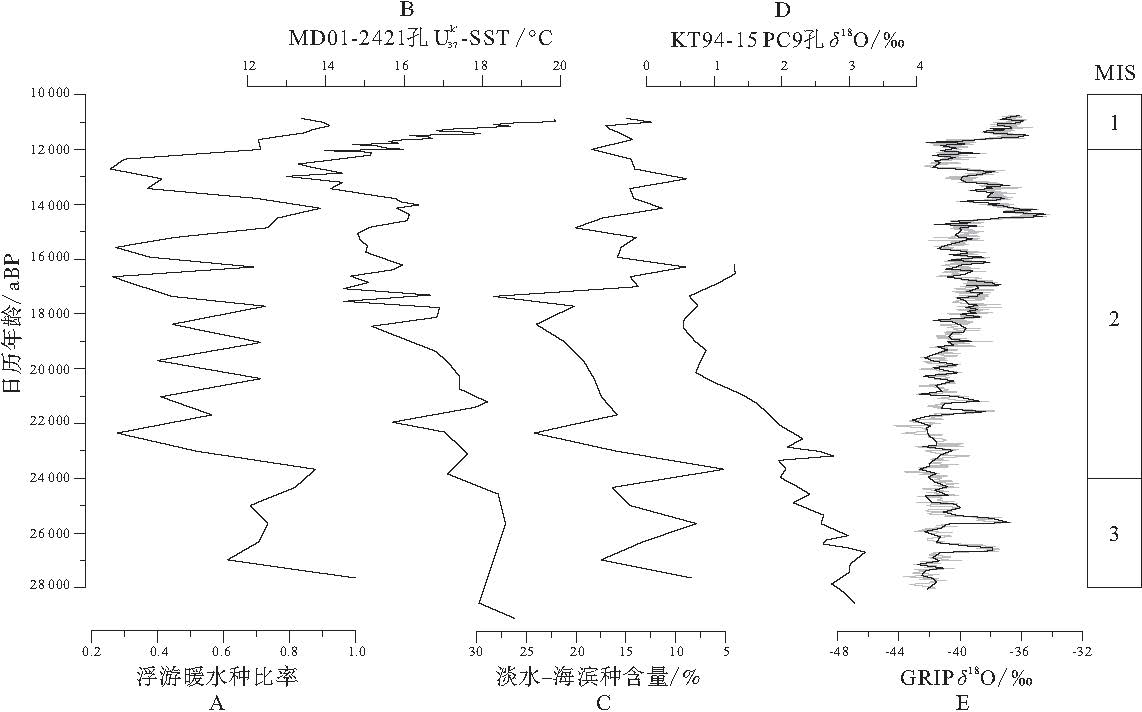 | 图5 日本海UB-2孔硅藻的各种测定指标 A--日本海UB-2孔硅藻浮游暖水种比率; B--日本东岸MD01-2421孔长链不饱和酮 |
笔者将淡水种与海滨种合并, 并以淡水--海滨种含量来表示淡水及沿岸流影响的强弱, 进而代表该地区的盐度变化(图 5-C).28000--24000 a BP期间, 淡水--海滨种含量不高; 在24000到17000 a BP段, 淡水--海滨种含量大幅度上升; 从17000到11000 a BP, 淡水--海滨种含量又有所降低.
笔者将硅藻组合中浮游暖水种比率和淡水--海滨种含量与日本东岸MD01-2421孔(36° 01.4'N, 141° 46.8'E)长链不饱和酮
日本海UB-2孔硅藻组合显示, 在28000--17000 a BP期间浮游暖水种比率较低, 除了在氧同位素3期(MIS3, 28000--24000 a BP)达到0.8左右之外, 基本在0.5上下浮动(图 5-A); 淡水--海滨种含量较高, 在10%以上(图 5-C), 说明此时日本海处于一个低温低盐的环境.日本东岸MD01-2421孔长链不饱和酮
4.1.1 氧同位素
3期(28 000--24 000 a BP) 从28000到24000 a BP浮游暖水种呈现较高的比率, 此间淡水--海滨种含量不多, 约为10%~15%.日本东岸MD01-2421孔长链不饱和酮
4.1.2 末次盛冰期(21000--17000 a BP)
随着气温的持续降低, 海平面也逐渐下降.在21000--17000 a BP期间, 日本海温度较低, 而表层海水受封闭的海洋环境影响盐度也较低, UB-2孔沉积物硅藻组合很好地反映了这一特点.
此时浮游暖水种比率达到最低, 代表着一个低温的海洋环境, 对应于末次盛冰期.Oba和Marayama(1995)认为日本海末次盛冰期表层海水温度较低, 一直稳定在4.4± 1 ℃ . 日本东岸MD01-2421孔长链不饱和酮
但是, 利用日本海长链不饱和酮
随着研究的进一步开展, 大多数古海洋学家均认定日本海末次盛冰期时并不存在高温现象(Seki et al., 2004; Fujine et al., 2006; Yokoyama et al., 2007; Khim et al., 2008b), 尤以Yokoyama等(2007)的研究最为细致.他们整合了大量研究结果, 发现周边海区末次盛冰期温度较低; 而日本海除长链不饱和酮
UB-2孔硅藻淡水--海滨种含量在末次盛冰期阶段明显升高, 日本海东部KT94-15 PC9孔δ 18O数据也骤然降低(Crusius et al., 1999), 也说明此时海水盐度较低.隐岐海岭的氧同位素在末次盛冰期变轻, 认为是表层海水盐度降低的信号, 并估计在此期间表层海水盐度从33‰ 下降到20‰ (Oba and Marayama, 1995), Mutsui等(1998)的盐度平衡模式实验也支持这一假设.
近年来学者们对日本海末次盛冰期古环境进行了大量研究(Crusius et al., 1999; Gorbarenko and Southon, 2000; Ishiwatari et al., 2001; Itaki et al., 2004; Dalai et al., 2005; Oba et al., 2006; Fujine et al., 2006; Khim et al., 2008b), 普遍认为该时期表层海水发生淡化.一方面, 末次盛冰期时海平面下降到现今海平面之下130 m(Itaki et al., 2004), 对马海峡(水深小于130, m)基本关闭, 无法为对马暖流提供进入日本海的路径, 此时日本海处于一个封闭的环境, 与外洋高盐度海水的交换大大减少, 而受陆地河流淡水影响较大(Lee, 2007).另一方面, 此时气候急剧变冷, 蒸发量显著减少, 使得降雨量高于蒸发量(Yokoyama et al., 2007), 进而形成表层海水低盐度的独特现象.
浮游暖水种比率的不断上升是该时期硅藻组合中最为显著的变化, 格陵兰冰芯(GRIP)δ 18O数据表明, 温度升高是此时最主要的气候特征(Dansgaard et al., 1993).Itaki等(2004)认为, 17000 a BP气温开始回升, 进入末次冰消期.而淡水--海滨种硅藻含量的降低, 说明此时海水盐度也有所上升.
4.2.1 波令--阿勒罗德暖期(B/A)(14 800--13 400 a BP)
在14800--13400 a BP期间, 浮游暖水种硅藻比率较高, 表明海水温度比末次冰期有所增加.日本东岸MD01-2421孔长链不饱和酮
4.2.2 新仙女木冷事件(YD)(13400--12300 a BP)
从13400到12300 a BP, 浮游暖水种比率大幅度下降, 达到末次冰期的水平, 尤其是浮游冷水种D. seminea含量突增, 淡水--海滨种含量也随之下降, 对应了一个突然变冷, 变干的海洋环境.日本东岸MD01-2421孔长链不饱和酮
4.2.3 Paralia sulcata含量上升
自15200 a BP起, 半浮游种(近岸浅海种)P.sulcata含量明显上升, 由末次冰期的平均不到10%, 提升至15%以上.尤其在11300--11100 a BP期间, P.sulcata含量达到30%以上的峰值.随后, 该种含量逐渐降低.
Ryu等(2005)在研究郁陵海盆(Ullueng Basin)2个站位沉积物硅藻时, 发现早全新世P.sulcata含量的迅速增长.随后, Ryu等(2008)综合了韩国半岛沿岸几个站位和新的郁陵海盆站位沉积物硅藻研究结果, 认为P.sulcata含量的变化, 可以用于指示对马海峡的开启与关闭.
末次盛冰期, 对马海峡封闭, P.sulcata含量也很少, 只有不到10%.直至15200 a BP, 其含量开始增加, 此时海平面约为 -110 m(Hanebuth et al., 2000; Steinke et al., 2006), 外洋水已可通过平均水深120, m的对马海峡进入日本海.对表层硅藻研究表明, P.sulcata在日本海含量不高, 但在东海50~100, m水深的海区含量非常丰富(蒋辉, 1987).故末次冰消期海平面的上升, 导致对马海峡开启, 使得P.sulcata可随东海沿岸流进入日本海.
在11300--11100 a BP期间P.sulcata达到峰值, 这是一个明显的冲淡水事件, 对应着10445 a BP的融冰冲淡水事件(mwp-IB)(Fairbanks, 1989).深色富有机质纹层TL-1的形成(Crusius et al., 1999), 也证明了该时期有大量淡水涌入日本海.
UB-2孔沉积物硅藻组合显示末次冰期至早全新世阶段古海洋环境变化显著.气候变化, 海平面波动及其所导致的古地理变化是古环境变化的主要原因.
末次冰期相对较低的气温, 导致海平面降低, 而日本海独特的地理环境, 使得海平面变化对古环境变化的影响更为敏感.尤其在末次盛冰期阶段, 海平面下降使得日本海成为与外洋隔绝的封闭海盆.从UB-2孔硅藻组合中可发现, 在此时期, 代表外洋暖水的硅藻基本消失.一方面, 陆上淡水的注入降低了日本海表层海水的盐度, 氧同位素也表现出异常的高值(Gorbarenko and Southon, 2000; Oba et al., 2006; Yokoyama et al., 2007); 另一方面, 由于表层海水盐度的降低, 水体因密度差异而出现分层现象, 底层水的缺氧停滞状况达到极致, 形成特殊的深色富有机质纹层(Tada et al., 1992).日本海UB-2孔硅藻组合在末次盛冰期浮游暖水种比率较低, 淡水--海滨种含量较高的结果很好地显示出该时期低温, 低盐的海洋环境.
随着气温的回升, 海平面逐渐上升, P.sulcata含量在末次冰消期的明显上升, 从一个侧面说明了对马海峡的开启.
位于日本海南部郁陵海盆的UB-2孔沉积物硅藻记录反映了11000--28000 a BP古海洋和古气候的变化, 包括深海氧同位素3期(MIS3), 末次盛冰期(LGM), 波令--阿勒罗德暖期(B/A)以及新仙女木冷事件(YD).
1)使用聚类分析得出3个硅藻组合带.其中17000--21000 a BP的分带, 清楚地划分出了末次盛冰期的界限.依照不同的生态习性, 将硅藻合并为暖水种, 冷水种, 淡水种以及海滨种, 其中暖水种比率与淡水--海滨种含量反映了11000--28000 a BP来该区域的古温度, 古盐度变化.
2)末次盛冰期日本海成为封闭的海盆, 其特殊的地理条件造成低温, 低盐的海洋环境, 暖水种比率与淡水--海滨种含量很好地反映出该时期古气候, 古环境的变化.
3)自15200 a BP起, Paralia sulcata含量逐渐上升, 这可能与海平面上升引起的古环境变化有关, 可以作为对马海峡开启的标志.早全新世出现的P.sulcata峰值, 对应着融冰冲淡水事件(mwp-IB).
4)气候引起的海平面变化是古地理环境变化的主要控制因素:低温导致的低海平面, 使得日本海与外洋隔绝, 成为封闭的海盆, 沿岸淡水的注入降低了表层海水盐度; 气温的回升带来海平面的上升, 高温高盐的外洋海水进入日本海, 对该区域海水温度, 盐度产生影响.
The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|