






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
下扬子地区下三叠统蠕虫状灰岩及其成因*
[张杰1, 2, 3  , 童金南
, 童金南3 ]
, 童金南|
|

第一作者简介:张杰,男,1983年生,硕士,中国石油杭州地质研究院助理工程师,主要从事碳酸盐岩沉积学及储集层研究.通讯地址:浙江省杭州市西湖区西溪路920号杭州地质研究院;邮编:310023;电话:0571-81765658;E-mail:zhangjiecug@sohu.com.
华南下三叠统中上部普遍发育蠕虫状灰岩,宏观上可以分为深色"蠕体"和浅色"基质".对于其成因的认识一直存在很大的争议,主要的观点有化学凝聚说,生物扰动说和机械成因说等.笔者通过对下扬子地区 5条典型剖面进行野外详细分层描述,采样,薄片观察以及实验分析,并与中上扬子地区进行对比研究,对蠕虫状灰岩进行了详细的定义,并将蠕虫状灰岩分为 4类:连续成层蠕虫状灰岩,生物爬迹 /钻孔蠕虫状灰岩,呈似层状 /杂乱条带状排列的蠕虫状灰岩和混染状蠕虫状灰岩,其在剖面上具有典型的变化规律.蠕虫状灰岩发育的层段少见宏体化石,遗迹化石,微生物的分布与蠕虫状灰岩之间具有一定的联系.早三叠世缺氧, H2S和 CO2含量增加,海水呈碱性,喜氧生物匮乏等为蠕虫状灰岩的形成创造了前提条件,灰泥在早三叠世特殊的海洋化学条件下开始沉积分异,后经历了同沉积成岩作用和不同程度的宏体生物,微生物作用,因此形成了不同类型的蠕虫状灰岩.蠕虫状灰岩发育时为海洋缺氧,有毒气体释放的后期,海水 X( Mg) /X( Ca)值开始降低,碳同位素值回升,随后生物逐渐活跃起来,环境也逐渐改善.因此蠕虫状灰岩可以被认为是生物复苏,环境改善的标志.
About the first author:Zhang Jie,born in 1983,master,is an assistant engineer of Hangzhou Institute of Geology and is engaged in the study of carbonate sedimentology and reservoir.Adress: Hangzhou Institute of Geology,Xixi Road 920,Xihu District,Hangzhou,Zhejiang Province,310023;Tel: 0571-81765658;E-mail: zhangjiecug@sohu.com.
The vermicular limestone is widespread in the middle-upper part of Lower Triassic of South China.It is a kind of mudstone and is composed of dark "vermicuform bodies" and tint "matrix".So far, a lot of studies have been carried out on the origin of vermicular limestone in various parts,but dispute still exists.Various hypotheses have been proposed to interpret its origin,including chemical coacervation,bioturbation and mechanical hypotheses. We made detailed stratification description,sampling,thin-section study and some experiment analysis on 5 typical sections in the Lower Yangtze Region and compared with some sections in the Middle-Upper Yangtze Region.We particularly defined the vermicular limestone and classified them as 4 types: Laminar vermicular limestone,trace/boring,laminar-like banding vermicular limestone,and contaminated vermicular limestone.Very few macro-fossils can be observed.The distribution of ichnofossils and microbe is related with vermicular limestone.In the Early Triassic,the poor oxygen,increased proportion of H2S and CO2,increased alkalinity and salinity of the sea water,and lacking of organism provides premise condition.The mud started to become diverse with special chemical conditions of the ocean in the Early Triassic and then underwent the synsedimentary lithification and different extent effect of macro life-form and animalcules.And then different kinds of vermicular limestone were formed.The vermicular limestone was developed in the later period of anoxic events and the release of poisonous gas events,when the X(Mg)/X(Ca) began to fall and the values of carbon isotope returned to higher levels.Subsequently,the environment improved gradually and the biota became active.As a result,the vermicular limestone could be considered as the indicator of the improvement of environment and biotic recovery.
二叠纪--三叠纪(P--T)之交的生物集群灭绝事件被认为是显生宙海洋无脊椎动物史上最大的灭绝事件, 造成海洋中90%, 陆地上70%以上的生物在地球上永远的消失, 之后的生物复苏却表现出迟缓性, 直到中三叠世生物才逐渐兴盛起来, 之间相隔了几百万年到近千万年的时间, 有些类别的复苏甚至用了更长的时间(童金南, 1997).生物复苏阶段跨越了整个早三叠世, 并且被生物种属极其贫乏的海底生物群控制, 其中包括一些全球性生物和机会分子.
除了特殊的环境变化以及生物组合缺乏多样性以外, 极端异常的沉积记录也是下三叠统的一个特点, 包括缺失礁, 燧石和煤的沉积(Pruss et al., 2005; Baud et al., 2007).除此之外, 在中国发现的主要的非正常沉积现象还有:蠕虫状灰岩, 潮下皱纹构造, 微生物岩, 海底碳酸盐胶结扇和泥质条带泥晶灰岩等(赵小明等, 2008).最近几年, 国内外地质学家对上扬子地区的非正常沉积现象进行了更加深入细致的研究, 认为二叠纪末的生物灭绝事件对早三叠世的生态系及海洋环境造成了巨大的干扰, 长期的环境恶化和频繁波动导致生物的复苏迟缓(方宗杰, 2004; Pruss and Bottjer, 2004; Pruss et al., 2006), 同时在整个早三叠世留下了非正常沉积的踪迹.
蠕虫状灰岩为中下三叠统一种典型的非正常沉积现象.对生物大灭绝后及复苏前出现的这种特殊的蠕虫状灰岩的研究有助于反映当时缺乏宏体生物的海洋环境条件以及宏体生物与微生物在生物复苏前的生存关系, 了解蠕虫状灰岩对滞后的生物复苏及海洋条件恢复的指示意义, 但迄今为止对蠕虫状灰岩的研究仅限于描述性的称谓, 其定义, 分类和成因均无定论.
作者对下扬子地区下三叠统蠕虫状灰岩沉积序列发育较完整的安徽广德牛头山, 宿松坐山, 池州乌石山, 江苏镇江大力山和小力山5条剖面进行了实测, 并将其与中上扬子地区的湖北兴山大峡口, 五峰, 贵州青岩, 土城和四川鱼洞梁等8条剖面进行了对比研究, 通过野外分层描述, 采样, 薄片观察, 揭片, 光片, 酸处理及X衍射, 荧光分析等实验方法研究, 对蠕虫状灰岩进行了详细地定义和分类, 以期对蠕虫状灰岩有更为深入全面的了解, 探讨其成因及宏体生物, 微生物在其形成中的作用及其与当时海水化学条件和生物复苏之间的关系.
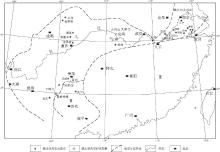
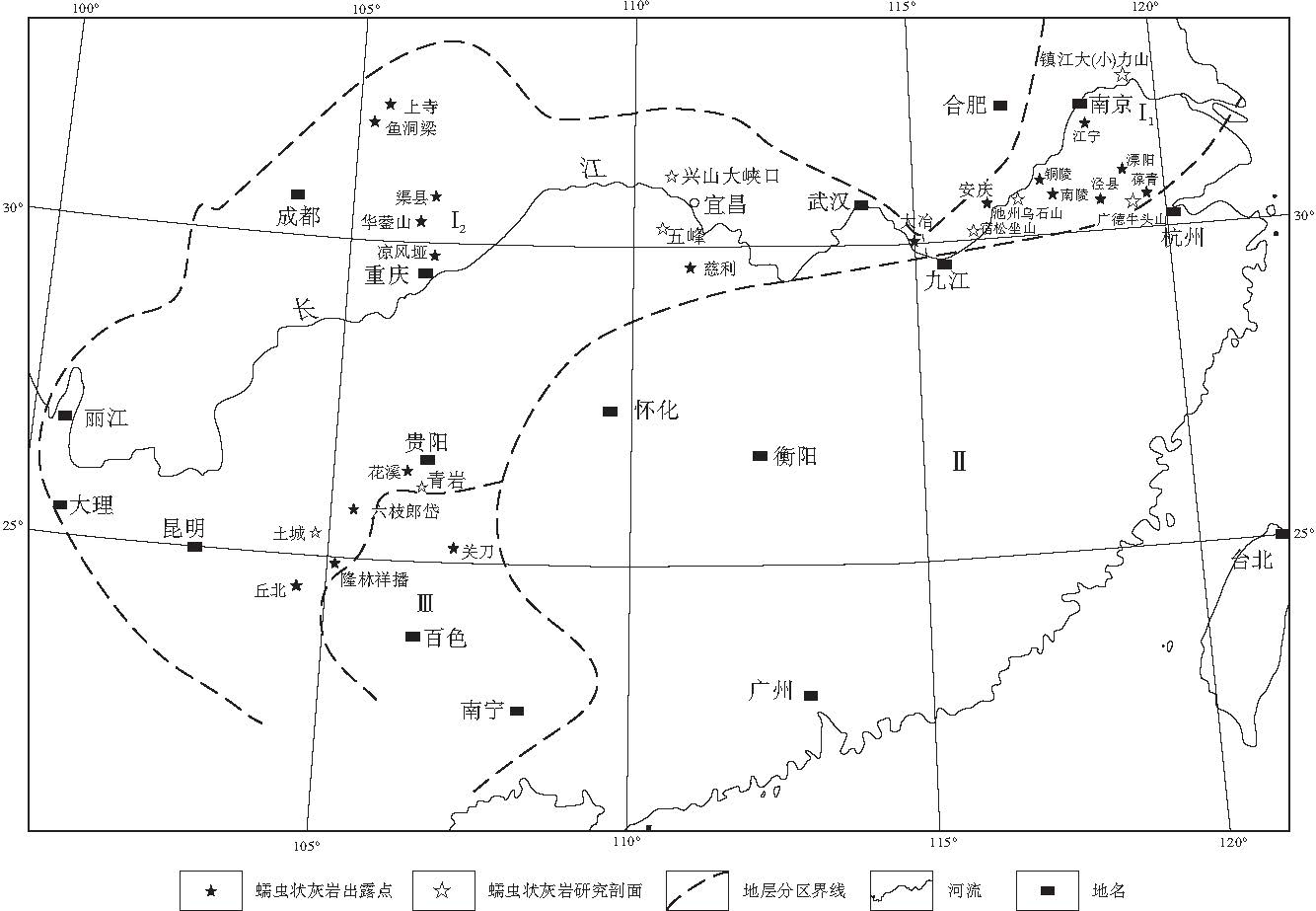 | 图1 中国南方早--中三叠世蠕虫状灰岩分布(地层分区底图据冯增昭等, 1997)(I1--下扬子地区; I2--上扬子地区; Ⅱ --华南区; Ⅲ --右江区)Fig.1 Distribution of vermicular limestone of the Early-Middle Triassic in South China (stratigraphic division after Feng et al., 1997) |
蠕虫状灰岩, 又称蠕粒状灰岩① , 蠕体石灰岩, 虫管灰岩, 生物扰动灰岩, 虫迹灰岩等, 英文名为vermicular limestone(使用较普遍), myrmekitic limestone或wormy limestone, 因其含有形似蠕虫的粒状体(即"蠕体"或"蠕粒")而得名.蠕虫状灰岩在各种研究文献中仅作为一种描述性的称谓, 至今还没有明确的定义和分类.蠕虫状灰岩在中国南方下三叠统中广泛分布(图 1), 有的地方延续到了中三叠统下部(表 1, 图2).蠕虫状灰岩在华南出现的层位为:下扬子地区和龙山组, 南陵湖组(少数地区包括东马鞍山组, 周冲村组下部), 中上扬子地区大冶组(飞仙关组), 永宁镇组(嘉陵江组), 偶见于中三叠统下部巴东组(雷口坡组).有文献记载的中国南方蠕虫状灰岩分布地区主要有:下扬子地区安徽中南部的铜陵, 贵池, 宿松, 广德等地, 浙江长兴葆青, 江苏镇江大力山; 上扬子地区湖南慈利, 黔中--黔南的贵州青岩, 花溪, 关岭永宁乡--六枝郎岱以及云南丘北腻脚, 四川广安县华蓥山, 渠县龙门峡和重庆凉风垭等地.
| 表1 华南蠕虫状灰岩出现地层分布(据冯增昭等, 1997; 童金南, 2005) Table1 Stratigraphic distribution of vermicular limestone in South China(after Feng et al., 1997; Tong, 2005) |
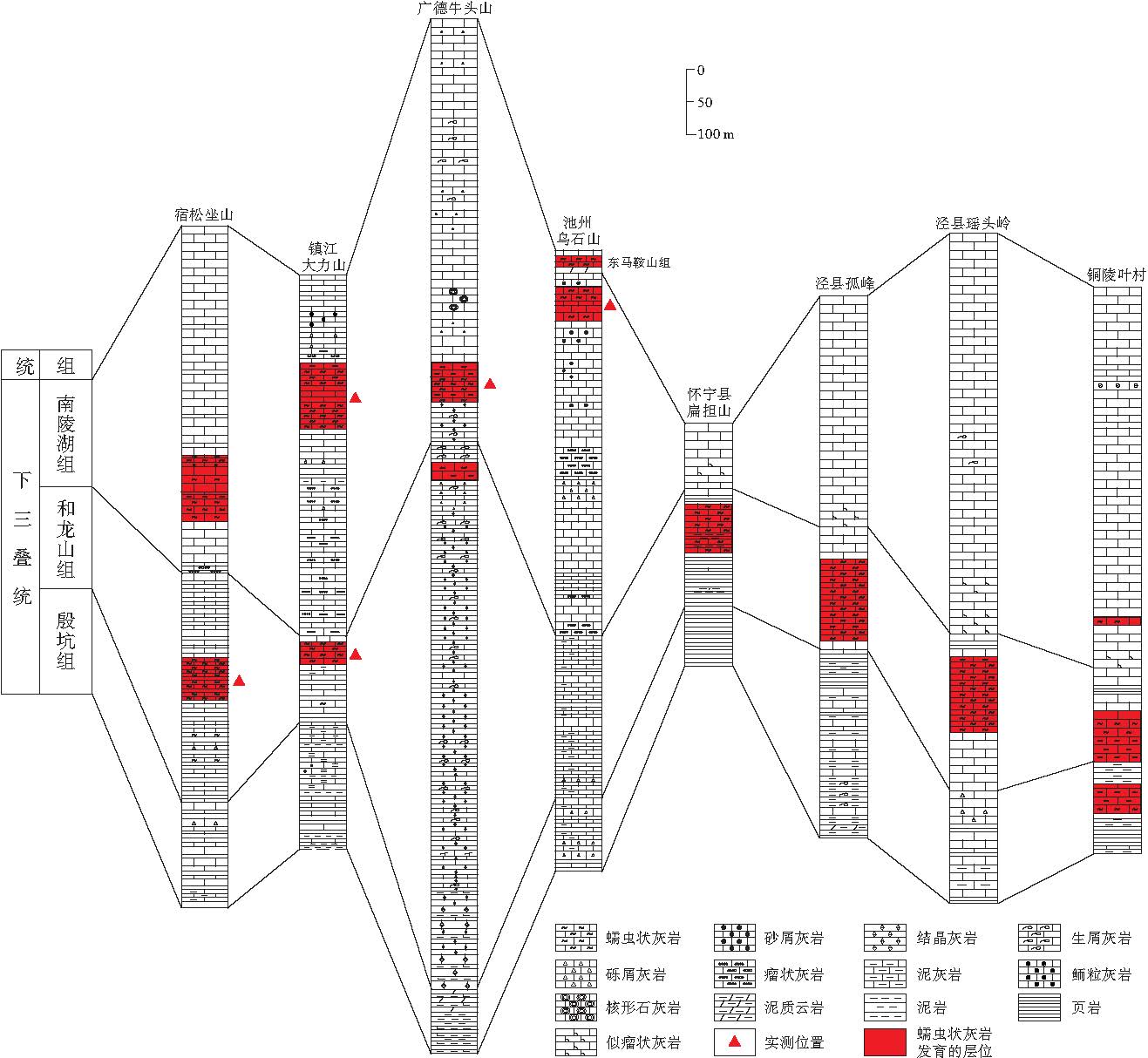 | 图2 下扬子地区部分剖面下三叠统柱状对比图(据实测及仇福康和陈才德, 1983; 汪贵翔, 1984)Fig.2 Correlation column of the Lower Triassic of some sections in Lower Yangtze Region(after measured data, Qiu and Chen, 1983; Wang, 1984) |
牛头山下三叠统剖面位于安徽省广德县东北部(图1), 毗邻浙江省长兴县.该地区三叠纪时处于江南隆起北部外缘, 属下扬子地台苏浙皖陆表海的中央一隅, 下三叠统发育不完全, 根据岩性及组合特征自下而上可划分为殷坑组(T1y), 和龙山组(T1h)及南陵湖组(T1n).牛头山剖面最典型的蠕虫状灰岩段位于和龙山组上部和南陵湖组下部(图 2).由于南陵湖组下部的蠕虫状灰岩段长而较完整, 出露良好, 因此笔者仅对南陵湖组下部厚约80, m的地层进行了实测(图 3).
蠕虫状灰岩在结构上由两部分组成, 即深灰色--灰黑色"蠕体(蠕粒)"及浅灰色"基质"(图4-1; 图5).牛头山剖面蠕虫状灰岩中的蠕体一般宽几毫米, 长度不定, 形状有粒状, 豆状, 管状, 长条状和藕节状等, 更多的呈不规则颗粒状和条带状(图 4-1, 4-4)等.蠕体四周一般下凹, 说明与基质相比, 蠕体抗风化能力较强.新鲜面上蠕体为深灰色--灰黑色, 基质为泥黄色--深灰黄色.有的蠕体顺层面分布, 有的蠕体切穿层面.
牛头山剖面蠕虫状灰岩从横切层面方向观察具有点斑状, 点线状, 条带状及不规则状等形态特征.其三维分布经连续切割发现为面状, 棍状和球状等.该剖面中的蠕虫状灰岩垂向分布(图3)具有规律性变化, 自下而上为:
1)宽而平直的深灰色蠕体条带与窄的颜色较浅的基质条带交互出现.蠕体条带宽几毫米到1, cm, 有时具微波状起伏, 抗风化能力强, 三维空间上呈面状展布, 向上逐渐变得弯曲.
2)波状蠕体条带与基质条带交互出现.蠕体条带宽几毫米到1, cm, 向上逐渐变得时断时续.
3)蠕体条带呈似层状不连续地分布于基质中, 横切面呈透镜状和蝌蚪状等, 宽仅毫米级, 基本成层展布.向上逐渐开始变得杂乱.
4)蠕体呈混染状杂乱分布于基质中, 大小约1, cm, 呈各种不规则颗粒状, 点斑状分布, 向上个体越来越小, 密密麻麻分布.
5)杂乱的蠕虫状灰岩消失, 取而代之的是均质的灰泥岩.
一个完整的序列包括了以上从1)至5)的5种类型, 但有的只有其中的几种; 一个完整的变化序列有的可达几十厘米甚至1, m, 有的仅有几厘米.
 | 图3 安徽广德牛头山剖面下三叠统南陵湖组下段实测柱状图Fig.3 Column of lower member of the Lower Triassic Nanlinghu Formation at Niutoushan section in Guangde of Anhui Province |
 | 图4 下扬子地区下三叠统蠕虫状灰岩宏观特征照片(1--广德牛头山剖面的蠕虫状灰岩, 蠕体呈深灰色, 条带状, 颗粒状, 0.2~1, cm不等, 呈似层状杂乱排列, 含量达到80%以上, 基质以泥质为主围绕其分布, 有的被溶蚀, 具压溶现象和缝合线, 方解石脉垂直穿过, T1n; 2--镇江大力山剖面的蠕虫状灰岩变化序列:下部1~3, mm的深灰色蠕体呈杂乱, 混染状分布于浅灰色--灰白色基质中, 向上变为宽1~2, mm, 呈顺层, 似层状排列的暗色蠕体, T1h上部; 3--宿松坐山剖面蠕虫状灰岩, 底部为泥晶灰岩, 不含蠕体; 下部深灰色--灰黑色蠕体大小约2~5, mm, 呈颗粒状, 短条带状分布于浅灰色基质中, 二者界线清晰, 局部见压实和缝合线; 上部蠕体较少, 呈似层状断续分布; 方解石脉普遍分布, T1h; 4--广德牛头山剖面的蠕虫状灰岩, 蠕体断续成层, 局部膨大, 四周被基质形成的纹理包围, 可见明显的拖曳/压溶现象(箭头所指), T1n; 5--镇江大力山剖面蠕虫状灰岩, 蠕体密集排列组成特殊现象(箭头所指):四周为斑点/条带状蠕体组成, 平行或交叉, 中部颜色较浅, 不含蠕体, 具有一定的扭曲, 垂直/斜交岩层面, T1n下部; 6--广德牛头山剖面蠕虫状灰岩中的扰动现象:杂乱状蠕虫状灰岩与层状蠕虫状灰岩互层, 前者中具缝合线, 后者中具下凹的"U"形扰动现象, T1n)Fig.4 Photos showing macro characters of vermicular limestone of the Lower Triassic in Lower Yangtze Region |
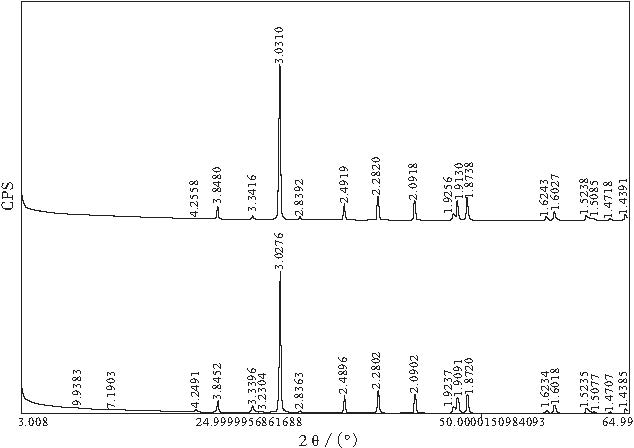 | 图5 安徽广德牛头山剖面蠕体(上)与基质(下)X 衍射图Fig.5 X-diffraction diagram of vermicuform body and matrix in Niutoushan section, Guangde of Anhui Province |
 | 图6 扬子地区下三叠统蠕虫状灰岩的显微照片(1--镇江大力山剖面蠕虫状灰岩层面上大量的遗迹化石Planolites:无分枝或偶有分枝的潜穴, 无清楚的内部构造, 横切面呈椭圆形, 直径1~3, mm, 粗细一致, 群生或相互交切, 水平; 墙壁清晰无衬里, T1h上部; 2--贵州青岩蠕虫状灰岩, 蠕体呈断续成层--杂乱排列, 光面扫描, T1l; 3--宿松坐山剖面蠕虫状灰岩蠕体中的藻纹层, T1h 上部SS-4 层顶, 单偏光; 4--广德牛头山剖面蠕虫状灰岩中的石膏假晶, T1n 底部NTS-4 层, 单偏光; 5--兴山大峡口剖面蠕虫状灰岩中的黄铁矿, 呈立方体晶形, T1d3, 单偏光; 6--广德牛头山剖面蠕虫状灰岩中的藻斑点, T1n 底部NTS-2 层, 单偏光)Fig.6 Microphotos of vermicular limestone of the Lower Triassic in Yangtze Region |
通过对包括安徽广德牛头山剖面在内的下扬子地区5条剖面蠕虫状灰岩的研究, 笔者发现蠕体多呈连续成层, 条带状, 似层状, 蠕虫状, 团块状, 蝌蚪状或絮状, 云雾状, 不规则颗粒状和斑点状等(图 4-1, 4-2, 4-3; 图6-2), 形态不规则, 大小差别悬殊, 小者小于1, mm, 大者长轴方向可达十几厘米.大小不等的蠕体呈混染状悬浮于基质中或与基质以缝合线接触, 基质纹理经常在透镜状蠕体两端产生收敛合拢(图 4-4).与盐酸反应时蠕体较快, 基质较慢, 且基质中的酸不溶物主要为黏土矿物和少量石英.显微镜下蠕体中的方解石颗粒较粗大(25~60, μ m), 结晶程度较好, 较纯; 基质成分也为方解石, 但颗粒较细, 结晶程度较差, 可能是沉积期后黏土杂质阻碍重结晶作用所致.X衍射分析(图5)发现, 蠕体的主要矿物成分为方解石(> 95%)和少量石英, 基质成分较复杂, 但仍以方解石为主(85%~90%), 残余许多不溶的黏土矿物(伊利石, 蒙脱石, 伊/蒙混层), 石英及少量绿泥石.
蠕虫状灰岩发育的层段少见宏体化石, 只在部分层面发现有厚度仅为毫米级的少量遗迹化石Planolites(漫移迹)等(图 6-1; 图7).薄片鉴定发现:蠕虫状灰岩的基质中含有少量广盐度的介形虫, 双壳类, 有孔虫碎片以及少量腕足类, 棘皮类等生物碎屑, 蠕体中生屑偶见, 但生物类型极其单调.藻类遗迹普遍, 主要有藻迹, 藻斑点和藻线纹等(图 6-3), 其中藻斑点形态不规则, 有的呈点线状排列(图 6-5).细小的黄铁矿颗粒呈散布状(图 6-6), 一般黄铁矿附近藻类遗迹较多.牛头山剖面薄片中所见到的少量硫酸盐矿物在大多数剖面中呈假晶, 成分为亮晶方解石的多晶集合体, 呈板柱状, 柱状, 大小为0.3~3 mm(图 6-4).
蠕虫状灰岩中还存在一些其他类型的沉积组构, 如在蠕体的周围分布着一些与藻同心纹层相似的纹层, 水平的蠕体--基质被扰动呈下凹的"U"形扰动斑块(图 4-6)等.下扬子地区许多剖面中的蠕虫状灰岩段还发育一种特殊的沉积现象(图 4-5), 表现为暗色蠕体密集排列, 垂直/斜交岩层面, 四周为斑点状蠕体组成, 平行或交叉, 中部浅色为基质, 其成因有待更深入的研究.
为方便研究, 综合诸多研究者之见, 并结合蠕虫状灰岩的特征, 将蠕虫状灰岩划分为连续成层的蠕虫状灰岩, 生物爬迹/钻孔蠕虫状灰岩和呈似层状, 杂乱状排列的蠕虫状灰岩和混染状蠕虫状灰岩4种.
迄今为止, 许多学者从不同方面对蠕虫状灰岩进行了研究, 并进行了不同的分类, 但至今没有定论; 对于其成因, 一直存在争议, 主要的观点有化学凝聚说, 机械成因说, 生物扰动说和复合成因说等.
化学凝聚说:罗正华(1977)① 认为蠕体是胶体成因, 即尚处于塑性状态的凝胶在弱动荡水动力条件下受冲刷或拖曳而形成.乐昌硕和田成(1981)② , 黄思静(1984), 罗璋和陈学时(1988)认为蠕虫状灰岩形成于水流滞缓, 环境闭塞, 海水盐度较高的潮坪环境, 由化学凝聚作用而形成.
机械成因说:鲍志东(1997), 胡广成等(2009)认为似纹层蠕虫状灰岩为静水条件下的正常沉积产物; 断链状蠕体应是似纹层蠕虫状灰岩遭受后期压溶作用的产物.华东地质研究所钾盐组(1977)认为蠕体是内碎屑成因, 即在中等强度的水动力条件下, 弱固结的泥晶方解石沉积物经扰动破碎形成.朱井泉(1988)认为蠕虫状灰岩为沉积分异和压实(溶)调整双重作用的结果.钱守荣(1995, 1996)认为蠕虫状灰岩与风暴作用有关, 是高能条件下快速堆积的产物, 实际上是一种较为特殊的风暴(浊积)岩.
 | 图7 湖北宜昌大峡口剖面蠕虫状灰岩段遗迹化石变化(1--仅在蠕虫状灰岩层表面的遗迹化石; 2--遗迹化石开始深入到岩层内部; 3--遗迹化石在岩层内普遍分布, 杂乱排列)Fig.7 Ichnofossil changes of the vermicular limestone member in Daxiakou section, Yichang of Hubei Province |
生物扰动说:许靖华(1980)③ 认为四川峨眉山嘉陵江组3段中的蠕虫状灰岩是藻席被虫子吃掉了部分, 剩下的部分形成了蠕虫构造.对于蠕虫状灰岩中的虫管和潜穴构造, 大多数研究者承认其为生物扰动形成的.
复合成因说:认为蠕虫状灰岩成因不是单一的, 不同类型的蠕虫状灰岩具有不同的成因.朱洪发等(1990), 于炳松(1991)及朱洪发和王恕一(1992)认为, 交替沉积的灰质与泥质经成岩期差异压实和压溶作用而成为不规则纹层状, 断续纹层状直至透镜状蠕虫状灰岩, 杂乱排列的云朵状, 不规则状蠕虫状灰岩的形成与早期生物扰动有关.冯增昭和吴胜和(1988), 黄明康等(1988), 刘泽均等(1988)及冯增昭等(1997)认为纹层状蠕体为静水条件下正常沉积的产物; 弯曲管状, 分叉管状和切割纹层的典型蠕虫状灰岩为生物扰动形成; 排列成交错层理的蠕体受过机械作用影响; 顺层断续蠕体多由不甚规则的纹层状蠕体经后期压溶作用改造而成, 与原始灰泥沉积物中的有机质演化有较密切的关系; 杂乱分布的不规则斑状蠕体和短条状蠕体, 为化学凝聚和机械作用共同作用而成.王英华等(1988)认为顺层断续和纹层状蠕虫状灰岩是继沉积分异后差异成岩作用的产物; 异形蠕虫状灰岩应为生物扰动成因; 顺层蠕体在食泥生物改造下可转化为异形蠕体.姜月华等(1992)认为沉积作用, 沉积分异作用是形成蠕虫状灰岩的重要前提, 而生物扰动, 水流作用和压实, 压溶作用则是形成各种蠕虫状灰岩的关键.赵小明等(2008)认为层状, 似层状(椭球状)蠕虫状灰岩分别为成岩分异作用, 化学作用和机械作用成因, 粒状蠕虫状灰岩为化学凝聚/微生物凝聚成因, 柱状蠕虫状灰岩中蠕体是生物潜管和扰动斑点, 为生物成因.
综上所述, 蠕虫状灰岩的成因很复杂, 到目前为止还没有公认的模式可以解释, 具体表现为:
1)生物成因?究竟是什么样的生物作用造成沉积了如此特殊的岩石类型?而且当时的生物类型极其单一, 且在蠕虫状灰岩发育段很少见到典型的较完整的生物化石, 通过很少见到的遗迹化石Planolites等无法推知具体的生物作用类型, 蠕体的形成与生物之间也不具有直接关系.
2)微生物成因?蠕虫状灰岩出现层位为下三叠统中上部--中三叠统下部, 位于生物大灭绝后即出现的微生物岩沉积之后, 之中虽不乏藻类斑点和藻线纹等, 但无典型的微生物藻类化石形态, 仅有斑点及一些藻纹层, 藻斑点内部纵纹, 基本无法鉴定, 其分布亦未发现具普遍的规律性, 因此单纯微生物成因的观点也值得商榷.
3)非生物成因?一般认为的非生物成因是指化学凝聚成因, 然而化学凝聚成因的说法很笼统, 正如黄思静(1984)所述, "化学凝聚成因的机理尚不清楚, 亦无实验证实, 一般认为与盐度的略微升高有关.蠕虫状灰岩中生物数量少, 种属单调, 缺乏窄盐度生物群以及颜色普遍偏深, 并常有细分散状黄铁矿伴生, 可能是水体循环差, 盐度偏高的佐证".于炳松(1991)认为蠕虫状灰岩与瘤状灰岩成因具有明显的一致性.笔者通过对安徽广德牛头山剖面这两类岩石的研究发现:二者的形成具有一定的相似性, 所不同的是后者的瘤体与基质是在不同时期沉积而成的灰泥和黏土, 这从成分上可以分辨; 此外, 二者的形成环境也不同(表 2).蠕虫状灰岩段夹有瘤状灰岩, 但二者却不共生, 亦从另一个方面说明二者形成环境的不同以及二者成因的不相容性.
| 表2 瘤状灰岩与蠕虫状灰岩特征对比 Table2 Correlation of characters of nodule limestone and vermicular limestone |
3.2.1 蠕虫状灰岩的形成环境及分布规律
很明显, 蠕虫状灰岩的形成应是多种条件综合作用的结果.通过对下扬子地区蠕虫状灰岩的研究, 可以得出一些认识:
1)蠕虫状灰岩形成于潮间带及潮下带水体较浅, 盐度较高, 相对比较安静的环境.
2)下扬子地区各剖面的蠕虫状灰岩都具有平直的蠕体条带→ 波状蠕体条带→ 断续成层蠕体条带→ 杂乱分布的混染状蠕虫体→ 均质的灰泥岩→ 平直的蠕体条带的垂向变化序列(张杰等, 2007).说明蠕虫状灰岩的沉积环境具有周期性变化.底部连续成层的蠕虫状灰岩为缺乏深入到沉积物内部的低水平生物扰动条件下灰质与泥质分异沉积形成的.成层的蠕体条带向上逐渐断开并且变杂乱, 可能因为后来经历了海水化学条件的变化和生物扰动.
3)华南地区早三叠世的台地和盆地碳酸盐岩含有丰富的海底同沉积成岩作用的证据(Wignall and Twitchett, 1999), 且许多证据表明早三叠世海水缺氧, 并处于文石海时期(Sandberg, 1983), 海水碱性增强, 不利于带壳生物生存.与此相反, 微生物在早三叠世大量繁盛(Baud et al., 1997)
3.2.2 海水化学条件
早三叠世的海水化学条件究竟有什么特殊性?结合前人的研究成果, 认为海水的化学条件主要表现为:
1)还原, 普遍缺氧/贫氧的海洋环境.二叠纪--三叠纪生物大灭绝后, 微生物在一个相对氧化的环境下发展起来, 而后由于海水由氧化的环境变为还原, 普遍缺氧的环境(Wignall and Hallam, 1992; Wignall and Twitchett, 1996), 微生物的发展受到影响.
2)同位素数据表明当时环境恶劣.早三叠世碳同位素值一直偏低(左景勋等, 2003), 表明当时环境不适于生物生存, 以致不能形成较强的生物扰动.此外, 早三叠世氧同位素也是负偏的(左景勋等, 2003), 硫同位素亦是负偏(Yoshimichi et al., 1995).硫酸盐剧烈减少导致了H2S 和CO2 的增多, 富含H2S 和CO2 的盆地水定期向陆架上涌毒害了后生生物.硫酸盐的还原产生了重碳酸盐(碳酸氢钙), 从而使得海水碱性增强.海水碱性增强(Pruss et al., 2005)在蠕虫状灰岩形成中具有很重要的作用:海水中的碱水上涌使得海水毒化; 有毒的条件在二叠纪末期生物大灭绝之后至复苏之前存在了几百万年, 这不仅导致了这些非正常沉积物的形成, 还抑制了海洋生物的复苏.
缺氧, H2S和CO2含量的增加, 海水碱性增强, 喜氧生物匮乏, 文石海(Lowenstein et al., 2001)等构成了早三叠世海洋环境的主要特征.当时的环境条件为蠕虫状灰岩的形成创造了前提条件.
3.2.3 蠕虫状灰岩的成因分析
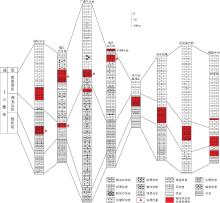
环境的变化影响了生物的扰动, 并导致了该扰动不能深入到岩石内部, 因而造成许多地区简单的漫移迹Planolites 普遍发育, 这与当时氧含量低的古环境条件一致.低水平垂向生物扰动使得开始沉积时形成了水平的条带状灰泥灰岩, 缺乏深入到岩石内部的生物扰动使得薄层灰岩得以保存, 为形成蠕虫状灰岩奠定了基础.从蠕虫状灰岩中发现的少量生物遗迹(图 6-1)和镜下观察到的藻类斑点(图 6-3, 6-6), 可知蠕虫状灰岩的形成不仅有宏体生物的作用, 亦有微生物的参与, 二者为此消彼长的关系.因此, 条带状灰泥灰岩在早三叠世特殊的海洋化学条件下开始沉积分异, 经历了同沉积成岩作用和不同程度的宏体生物, 微生物作用, 形成了不同类型的蠕虫状灰岩(图 8).
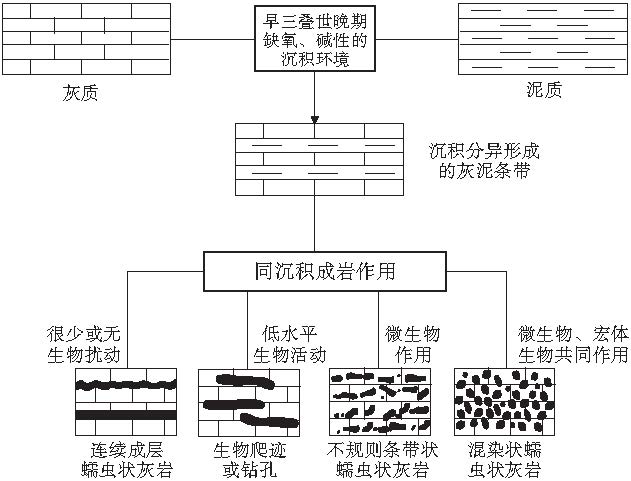 | 图8 蠕虫状灰岩形成模式Fig.8 Formation model of vermicular limestone |
蠕虫状灰岩具有平直的蠕体条带→ 波状蠕体条带→ 断续成层蠕体条带→ 杂乱分布的混染状蠕体→ 均质的灰泥岩→ 平直的蠕体条带的垂向变化序列(张杰等, 2007; 张杰, 2008), 说明生物活动对沉积物的影响曾一次次增强, 但仍迫于环境压力而未能深入到沉积物内部, 以致垂向生物遗迹不发育.这从另一个方面说明, 当环境逐渐恢复正常时, 蠕虫状灰岩中这种低水平的生物扰动必被深入的, 受到生物活动强烈改造的其他类型岩石所代替.
另外, 压实作用对蠕体形态也具有改造作用.基质因压实作用而发生的体积收缩应比蠕体大得多, 这可能就是基质纹理经常在透镜状蠕体两端产生收敛合拢的原因, 也导致蠕虫状灰岩中各种类型缝合线的形成.
部分剖面(如宜昌大峡口剖面)中的蠕虫状灰岩段发育丘状交错层理和鲕粒盖层, 其形成可能与风暴作用有一定的关系(张杰, 2008).
可见, 单一的条件并不足以形成蠕虫状灰岩, 其形成有宏体生物的作用, 亦有微生物的参与, 二者为此消彼长的关系.当时特殊的海水环境在蠕虫状灰岩形成过程中更起到了关键性的作用.此外, 宏体生物扰动和微生物的作用影响了海水性质, 对海水化学条件具有一定的反作用, 并导致了沉积环境和沉积物成分的变化(图 9).
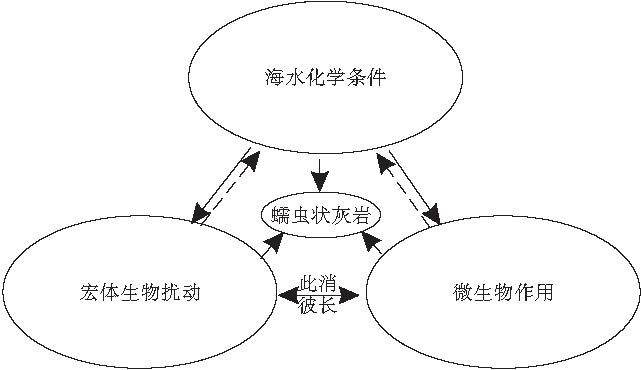 | 图9 蠕虫状灰岩形成的影响因素及其相互关系(实线表示决定性作用, 虚线表示反作用)Fig.9 Affecting factors and their relationship of vermicular limestone |
二叠纪末海洋中发生了大规模海水缺氧事件(Wignall and Hallam, 1992; Wignall and Twitchett, 1996, 2002), 持续时间约6~10, Ma; 海洋底层存在大量的CO, CO2, H2S和CH4等水溶性有毒气体(李玉成和周忠泽, 2002), 其持续存在导致了宏体生物灭绝, 微生物岩随后出现(杨浩等, 2007).在这种有毒气体释放的后期, 环境逐渐恢复正常, 海水 X(Mg)/X(Ca) 值开始下降(Wilkinson and Algeo, 1989; Horita et al., 2002; 颜佳新和伍明, 2006), 此时出现的蠕虫状灰岩预示着生物复苏即将全面开始.在下扬子地区奥伦尼克期, 碳同位素值经历了第2个"U"型谷后逐渐回升为正值, 碳同位素值的上升与负偏反映生物的复苏与灭绝(左景勋等, 2003), 因此碳同位素值的回升也表明生物已经全面复苏.
关于早三叠世牙形石演化的研究(丁连生和包德宪, 1989; 王成源, 1993)发现, 在蠕虫状灰岩沉积结束后, 牙形石的多样性大幅增加, 这也表明了生物的复苏及环境的逐步改善.许多关于当时生物复苏形式与环境变化之间关系的研究也表明, 随着蠕虫状灰岩沉积的结束, 生物逐渐活跃起来, 环境变得适于生物生存(Horita et al., 2002).
至早三叠世晚期蠕虫状灰岩沉积逐渐减少也可作为当时全球海洋环境逐步得到改善的一个标志.从宜昌大峡口剖面可以明显发现造迹生物作用有逐渐增强的趋势, 从开始所发现的遗迹化石很少且仅在表面分布直到深入到岩层内部并普遍分布(图 7), 说明该时期开始宏体生物活动受到限制, 后来环境对生物的影响逐渐减弱.
因此, 蠕虫状灰岩的消失可以作为早三叠世死寂生物面貌时代的结束, 生物全面复苏的开始以及环境正常化的重要标志.
作为生物大灭绝后出现的一种重要的非正常沉积, 蠕虫状灰岩的出现对于早三叠世荒凉的生物界具有重要的指示意义.
1)蠕虫状灰岩在华南下三叠统中上部--中三叠统底部普遍分布, 宏观上可以分为深色"蠕体"和浅色"基质".蠕体具有连续成层, 条带状, 似层状, 不规则颗粒状等外观形态, 宏观上与基质界线清楚.蠕体成分主要为方解石和少量石英, 基质成分较蠕体复杂, 但仍以方解石为主, 残余许多不溶的黏土矿物等.基质纹理经常在透镜状蠕体两端产生收敛合拢.蠕虫状灰岩发育的层段少见宏体化石, 仅有少量厚约1~2, mm的遗迹化石, 镜下生物类型单调, 仅具有广盐度的生物, 藻类遗迹普遍.
2)蠕虫状灰岩具有平直的蠕体条带→ 波状蠕体条带→ 断续成层蠕体条带→ 杂乱分布蠕体→ 均质的灰泥岩变化序列.按照形态及成因, 可以分为连续成层的蠕虫状灰岩, 生物爬迹或钻孔, 条带状蠕虫状灰岩和混染状蠕虫状灰岩4种.
3)单一的条件并不足以形成蠕虫状灰岩, 4种蠕虫状灰岩的形成既有宏体生物的作用, 亦有微生物的参与, 二者为此消彼长的关系.文石海, 缺氧, H2S和CO2含量的增加, 海水碱性增强, 喜氧生物匮乏等当时的环境条件为蠕虫状灰岩的形成创造了前提条件.灰泥在早三叠世特殊的海洋化学条件下开始沉积分异, 后经历了同沉积成岩作用和不同程度的宏体生物, 微生物作用, 因此形成了不同类型的蠕虫状灰岩.
4)蠕虫状灰岩发育时为海洋缺氧, 有毒气体释放的后期, 碳同位素值回升, 海水X(Mg)/X(Ca) 值从峰值开始降低, 随后生物逐渐活跃起来, 环境也逐渐改善.因此, 蠕虫状灰岩可以被认为是生物复苏, 环境改善的标志.
致谢 论文工作系作者在中国地质大学(武汉)学习期间完成, 是中国地质大学"转折期研究"群体成果之一.感谢《古地理学报》主编冯增昭老师, 审稿人刘建波老师, 吴胜和老师和朱井泉老师对论文修改提出的宝贵意见!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|