




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
河南焦作地区下二叠统太原组遗迹化石及其沉积环境*
[胡斌1, 2  , 周方
, 周方1 , 宋慧波1, 2 ]
, 周方|
|

第一作者简介:胡斌,男,1952年生,河南理工大学资源环境学院教授,博士生导师,主要从事遗迹学与沉积学的教学和研究工作.E-mail: hub@hpu.edu.cn.
河南省焦作地区下二叠统太原组发育一套陆表海碳酸盐岩以及海湾--潟湖,潮坪--沼泽和障壁岛(砂质滩,坝)相碎屑岩和煤层,其中保存有大量完好的遗迹化石,已识别出的有 10个遗迹属 11个遗迹种,包括Zoophycos brianeus,Zoophycos villae, Teichichnus rectus,Nereites isp.,Chondrites isp.,Rhizocorallium isp.,Thalassinoides suevicus,Ophiomorpha nodosa,Gordia marina,Taenidium satanassi和Planolites isp.等.这些遗迹化石多为全浮痕保存,少数为半浮痕表迹保存,其丰度较高.根据遗迹化石的组成与产状特征及其围岩的沉积特征,可识别出 3种遗迹化石组合:( 1)Zoophycos -Nereites遗迹组合,出现在水体相对较深,浪基面之下且沉积底层内氧气贫乏的远岸浅海沉积环境;( 2)Zoophycos -Taenidium 遗迹组合,产生于水体相对较浅的浪基面之上的近岸浅海沉积环境;( 3)Ophiomorpha-Thalassinoides遗迹组合,发育于海岸带潮坪或海湾,潟湖沉积环境,遗迹化石往往保存在泥质灰岩或钙质粉砂岩和薄层细砂岩中.基于遗迹化石组合的分布特点,提出了焦作地区太原组遗迹化石的沉积环境模式.
About the first author:Hu Bin was born in 1952.He is a professor and Ph.D.supervisor of Henan Polytechnic University and is mainly engaged in teaching and researches of ichnology and sedimentology.E-mail: hub@hpu.edu.cn.
The Lower Permian Taiyuan Formation in the Jiaozuo area of Henan Province is composed mainly of carbonate,clastic rocks and coal beds, which are deposited in such sedimentary settings as epicontinental sea,bay-lagoon,tidal flat-shore swamp and barrier islands(sandy shoal and bar).This unit contains abundant ichnofossils including 10 ichnogenus and 11 ichnospecies, such as Zoophycos brianeus, Zoophycos villae, Teichichnus rectus, Nereites isp., Chondrites isp., Rhizocorallium isp., Thalassinoides suevicus, Ophiomorpha nodosa, Gordia marina, Taenidium satanassi, Planolites isp.and so on.These ichnofossils,displaying higher abundance,were preserved mostly in full relief and also partly in semi relief.According to composition and occurrence characteristics of ichnofossils and sedimentary features of the host rock,three ichnoassemblages were recognized:(1) Zoophycos-Nereites ichnoassemblage,appeared in the shallow marine environment below wave base,and the substrate is in the relatively deeper water and anoxic environment.(2) Zoophycos-Taenidium ichnoassemblage occurred in the shallow marine environment above wave base.(3) Ophiomorpha-Thalassinoides ichnoassemblage is formed in the marlite or calcareous siltstone and thin-bedded fine sandstone generated within soft ground or semi-consolidation ground in the tidal flat and lagoon sedimentary environments.Based on the distribution characteristics of the ichnoassemblages,this paper suggests the sedimentary environment model of the ichnofossils of the Taiyuan Formation in this study area.
文中所研究的下二叠统太原组剖面位于河南省焦作市西部的刘庄附近(图 1), 地层出露良好, 总厚度80~110, m, 为一套海陆交互相沉积, 含灰岩8~9层, 灰岩层间夹厚度不等的灰色细砂岩, 灰白色石英砂岩, 深灰色粉砂岩, 砂质泥岩, 黑色泥岩和薄煤层.根据垂向上岩性的变化特征, 该区太原组可进一步分为下部灰岩段, 中部碎屑岩段和上部灰岩段(图 2), 与下伏上石炭统本溪组和上覆中二叠统山西组呈整合接触.应该指出的是, 在该露头剖面的下部灰岩段(L2-L5)中发现化石Sphaeroschwagerina moelleri, 其地质时代相当于俄罗斯地台和南乌拉尔地区下二叠统阿瑟尔阶(Asselian)和美国西德克萨斯地区下二叠统的Pseudoschwagerina 带(姚庚云, 1989).因而焦作地区太原组应划归下二叠统, 其上覆山西组依据植物化石组合带而划归中二叠统(张海清等, 2008).这一划分与整个华北晚古生代地层的划分大体一致, 可与太原西山煤田的太原组和山西组对比, 只是太原组底部碎屑岩段在不同地区地质时代的归属有所不同, 有的归早二叠世, 有的归晚石炭世--早二叠世(张鹏飞等, 2001; 汪啸风等, 2005).
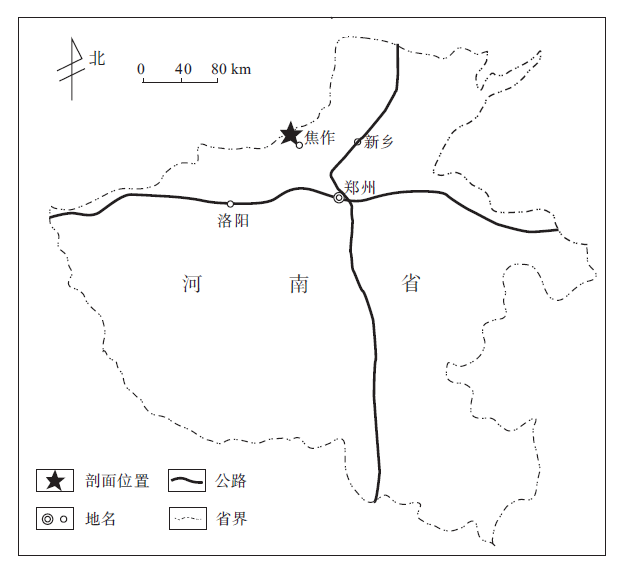 | 图1 研究区地理位置图Fig.1 Location of study area |
焦作地区太原组主要是一套浅海相碳酸盐岩和陆源碎屑岩的混合沉积, 是晚古生代华北陆表海盆地或碳酸盐岩台地沉积的一部分.基于对该组剖面的详细观察, 对岩性与沉积特征以及遗迹化石的研究, 将其划分为以下5种沉积相带(图 3):
 | 图2 河南焦作地区太原组沉积相及遗迹化石分布特征(DS--远岸浅海相带; PS--近岸浅海相带; BI--障壁岛相带; BL--海湾--潟湖相带; TS--潮坪--沼泽相带)Fig.2 Distribution characteristics of sedimentary facies and ichnofossils in the Taiyuan Formation in Jiaozuo area, Henan Province |
 | 图3 河南焦作地区太原组沉积相带分布模式Fig.3 Distribution model of sedimentary facies of the Taiyuan Formation in Jiaozuo area, Henan Province |
远岸浅海相带属台地相区浪基面之下, 水体相对较深的沉积区域.在该相带内, 沉积特征以深灰色薄--中厚层状泥晶生物屑灰岩, 生物屑泥晶灰岩为主, 发育不规则的丘状交错层理和水平层理, 含大量的浅海生物化石, 如珊瑚, 海百合, 腕足类, 藻类和苔藓虫等.生物化石与碎屑混杂堆积, 分选差, 所以为原地埋藏.其中, 遗迹化石十分丰富, 尤其是在深灰色泥晶灰岩及钙质泥岩中, Zoophycos遗迹化石大量出现, 并且潜穴充填物为黑色沉积物, 伴生的其他遗迹化石还有Nereites, Chondrites, Teichichnus 和Planolites等.据前人研究(晋慧娟, 1999), 这些遗迹化石常出现在海水相对较深而且水体较平静的海底沉积物中.有时还出现斑状生物扰动层(如L5灰岩中部), 说明当时在较平静的海底有大量生物在底层中活动.故推测其形成在浪基面以下, 水体相对较深的浅海沉积环境.此外, 在该台地相带的碳酸盐岩中还出现多层风暴沉积(吴贤涛等, 1987).
近岸浅海相带发育在台地相区浪基面之上, 水体相对较浅的区域.在该相带内, 主要岩性为灰色中厚--厚层状亮晶生物屑灰岩, 鲕粒灰岩及泥晶生物屑灰岩, 局部发育不规则的交错层理.生物碎屑丰富, 由多种生物介壳组成, 以腕足类和珊瑚碎屑最多, 其次有海百合茎, 苔藓虫和类生物碎屑.遗迹化石大多为Zoophycos, 伴生有U形具蹼状构造的潜穴Rhizocorallium, 近水平具回填纹构造的Taenidium和沿层面分布的管状潜穴Planolites等.这些遗迹化石常出现在近岸浅海并经常受多种水流影响的沉积环境中, 大量介壳碎屑和鲕粒灰岩的出现也证明了这一点.
岩性为灰色, 深灰色, 灰黑色泥岩, 粉砂质泥岩, 夹薄层粉, 细砂岩.泥岩中发育水平层理和缓波状层理.沿层理面分布有大量的菱铁质结核及黄铁矿晶粒.常见植物碎屑, 半咸水的动物化石, 偶见海绵骨针, 含有Zoophycos遗迹化石.这些沉积特征显示其为一种较闭塞, 水体较平静且还原程度较高的沉积环境.
由深灰色, 灰黑色粉砂岩, 深灰色, 灰黑色泥岩, 碳质泥岩和煤组成.含有较多的植物叶片和根化石, 泥岩中黄铁矿结核常见.典型的沉积构造有波状层理, 透镜状层理和砂, 泥互层层理等.生物扰动构造及生物潜穴在互层的沉积物中发育, 生物潜穴以高角度倾斜或近乎直立的潜穴为主, 如Thalassinoides, Ophiomorpha, Rhizocorallium和Gordia等.这些沉积特征和遗迹化石组合特征反映水体较浅, 为经常有潮汐水流作用, 且周期性暴露水面的潮坪与泥炭沼泽环境.
前述浅海相碳酸盐岩主要发育在研究区太原组下部和上部, 而障壁岛--潟湖--潮坪体系则主要在中部碎屑岩段中发育.障壁岛相主要岩性以成熟度较高的中--细粒石英砂岩为主, 石英含量达95%以上, 分选性和磨圆度都十分好, 具硅质胶结, 这种岩性特征反映了滨岸地带的强水动力条件以及沉积物经过了较长距离的搬运.常见楔状, 波状, 板状交错层理, 并与海湾--潟湖相共生.一般来讲, 高能水动力条件不利于生物活动和化石的保存, 所以, 未发现完整的实体化石, 遗迹化石也相当稀少.
综上所述, 从岩性的纵向变化特征来看, 太原组下部和上部以浅海相碳酸盐岩为主, 夹滨岸潮坪--沼泽相的泥岩和煤, 遗迹化石丰富; 中部则以障壁岛, 海湾--潟湖, 潮坪--沼泽相陆缘碎屑沉积为主, 含薄煤层, 遗迹化石较少.总体上看, 焦作地区早二叠世太原组沉积自下而上反映了一个完整的海进--海退--海进的沉积旋回(图 2).
在焦作太原组中观察和采集到大量遗迹化石, 共鉴定出10属11种.按生态或造迹生物的行为习性分类, 这些遗迹化石以进食迹为主, 常见有Zoophycos brianeus, Zoophycos villae, Teichichnus rectus, Rhizocorallium isp., Chondrites isp., Planolites isp.和Taenidium satanassi等.其他还有居住迹Thalassinoides suevicus和Skolithos linearis以及觅食迹Gordia marina等.根据个体遗迹化石分析, 推测该遗迹群落的造迹生物主要为蠕虫类, 环节动物和节肢动物等.现将常见遗迹种的形态, 构造, 保存及产状特征分别描述如下:
墙迹Ichnogenus Teichichnus(Seilacher, 1955)
直立墙迹Teichichnus rectus(Seilacher, 1955)(图 4-A)
描述:在纵向剖面上呈现为与层面近乎垂直的直形管状潜穴, 具U形蹼状构造, 潜穴宽1.2, cm, 可见长6, cm, 潜穴内蹼间距为0.3, cm, 填充物为灰白色, 明显浅于围岩.
讨论:遗迹属Teichichnus常被发现于开阔浅海--近滨沉积环境中, 被认为是由于食泥动物连续不断向上位移形成的一系列水平长潜穴并叠覆的蹼状遗迹构造(Seilacher, 1955), 也是Cruziana遗迹相的典型分子(Seilacher, 1967).它大多产生于滨外沉积环境(Frey and Howard, 1985), 是潮下带或浪基面之下浅海沉积环境中的常见分子(Corner and Fjalstadt, 1993).它的造迹生物主要是环节动物和节肢动物门类(Chisholm, 1970; Chamberlain, 1977).
层位:焦作下二叠统太原组L5灰岩.
蛇形迹Ichnogenus Ophiomorpha(Lundgren, 1891)
瘤状蛇形迹 Ophiomorpha nodosa(Lundgren, 1891)(图 4-B)
描述:圆柱形单个潜穴, 直径为3.2, cm, 可见长度为11, cm, 表面具突起的瘤状物, 瘤粒大小不一致, 一般为4 mm× 3 mm× 3, mm, 瘤体呈尖棱状, 顶端具尖刺, 排列较整齐, 剥蚀后外表光滑.潜穴内充填物为深灰色粉砂岩, 与围岩成分近同.
讨论:潜穴瘤状壁发育, 较厚, 说明沉积基底松软, 含水分且需要加固.Ophiomorpha造迹生物喜欢在这种较松软的基底上营造潜穴.从目前已发现的标本来看, 多数Ophiomorpha都出现在近岸砂岩(包括潟湖, 河口湾, 潮坪, 三角洲相)中, 少数见于近岸灰岩, 浅海风暴岩和深海浊积岩中.这些环境都容易发生快速的沉积作用, 沉积常具周期性, 沉积物松软, 含水分和有机质, 且含氧量较高.现代海岸带沉积物中挖潜的十足甲壳动物如Callianassids可营造类似Ophiomorpha 的潜穴(Pickett et al., 1971).
层位:焦作下二叠统太原组中部碎屑岩段.
海生迹Ichnogenus Thalassinoides(Ehrenberg, 1944)
苏爱维海生迹Thalassinoides suevicus(Kennedy, 1966)(图 4-C)
描述:在泥质灰岩层面上呈Y形叉枝状潜穴, 分枝末端较为圆滑, 呈鹿角形弯曲.潜穴宽4~8, mm.充填物为浅灰色, 明显浅于围岩, 部分潜穴充填物风化为黄褐色灰质泥岩.较为密集, 潜穴壁光滑无饰.
讨论:一般认为Thalassinoides是甲壳动物十足目虾类或类虾的居住兼觅食潜穴(Wetzel, 1991), 多见于滨海潮间带沉积环境, 也常见于滨岸地区半咸水环境, 如潮道, 潟湖, 河口湾和三角洲环境, 现代海岸地带玉虾类常形成类似的穴道(杨式溥等, 2004).
层位:焦作下二叠统太原组中段L6灰岩.
根珊瑚迹Ichnogenus Rhizocorallium(Zenker, 1836)
根珊瑚迹(未定种)Rhizocorallium isp.(图 4-D)
描述:平行于层面分布的U形管状潜穴.两翼管近平行排列.翼管长为18~23, cm, 管径1~1.3 cm.两翼间距为1.3~1.8, cm, U形管顶端呈半圆形.U形管间具明显弧形蹼状纹理构造, 蹼纹宽约2~4 mm.产于深褐色薄层细砂岩中, 充填物为红褐色铁质砂岩, 明显区别于围岩.
讨论:对于Rhizocorallium形态构造特征, Basan 和Scott(1979)认为两翼近平行的翼管之间为同心状前进式的蹼状构造.而大部分根珊瑚迹呈近水平或略有倾斜分布, 证明为采掘觅食迹, 但Veevers(1962)却认为是居住潜穴.Sellowood(1970)认为造迹动物同现代美人虾相似, 在筑穴时食沉积物, 建成潜穴之后即为滤食性, Fü rsich(1974)提出遗迹种Rhizocorallium jenense的造迹生物为原始的滤食动物.在海相遗迹相中, Rhizocorallium是Glossifungites遗迹相的典型遗迹化石, 常见于滨, 浅海沉积环境, 特别是潮间带和潮上带沉积环境, 其沉积底层往往是半固结的碳酸盐底层或稳定的, 凝结的和部分脱水的泥质底层.许多学者认为Rhizocorallium可形成于多种沉积环境, 如前滨高能环境(Fü rsich, 1975), 临滨带(Worsley and Mø rk, 2001), 斜坡环境(Knaust, 1998), 深水环境(Uchmann, 1992)和正常天气浪基面之下的滨外或潟湖环境(Fü rsich, 1981, 1998)等.还有的认为Rhizocorallium可出现在风暴沉积间歇期的较低能环境(Rodri'guez-Tovar and Pe'rez-Valera, 2008).
层位:焦作下二叠统太原组中部碎屑岩段.
丛藻迹 Chondrites Von(Sternberg, 1833)
丛藻迹(未定种)Chondrites isp.(图 4-E)
描述:该遗迹标本是一种小型分枝潜穴, 在灰岩层的垂直断面上呈现为小而长的椭圆形斑点状集合体, 直径0.5~1.2 mm.潜穴壁光滑, 局部可见极薄的深灰色泥质衬壁.潜穴内的充填物为灰白色钙质泥岩, 明显区别于围岩, 呈内迹保存在中厚层状灰色灰岩中, 并且常常与Nereites伴生.
讨论:一般认为, Chondrites是特殊的贫氧环境的遗迹化石标志, 晋慧娟等(2003)对中国近10个深海沉积区遗迹化石研究发现, 它是一种穿相遗迹化石.龚一鸣(2004)详细讨论了遗迹化石Chondrites的指相意义和阶层分布特点, 列举了国内外学者的研究成果, 其中许多学者认为Chondrites系缺氧和贫氧环境的指相遗迹化石, 并认为Chondrites是以缺氧环境中化学合成细菌为食的蠕虫动物所营造的化学共栖(chemosymbiotic)构造.
层位:焦作下二叠统太原组L4和L5灰岩.
类沙蚕迹 Nereites (Macleay, 1839)
类沙蚕迹(未定种)Nereites isp.(图 4-F)
描述:弯曲或回转式弯曲的蛇曲形潜穴.潜穴直径2~4, mm, 可见长度大于6, cm, 横切面为椭圆形, 有明显的衬壁, 衬壁厚1, mm左右, 由微小瘤状的黑色泥岩组成.潜穴内的充填物为浅灰色灰泥岩, 呈内迹保存在中厚层状灰色灰岩中.
讨论:Seilacher(1983)把 Nereites解释为食沉积物的内生动物所营造的底层内觅食及运动拖迹, 多产生于深海薄层浊积岩的岩层表面, 是深海遗迹相的典型代表, 常见于深海--半深海沉积环境.但是, Crimes和Anderson(1985)认为Nereites也可出现在浅水环境, 并且是一种广深性遗迹.有关Nereites的造迹生物有多种解释, 有的推测为蠕虫类(Richter, 1928)和腹足类(Raymond, 1931), 还有的认为是甲壳类动物(Fraipont, 1915).
层位:焦作下二叠统太原组L4和L5灰岩.
线形迹 Gordia (Emmons, 1844)
海生线形迹Gordia marina (Emmons, 1844)(图 4-G)
描述:光滑细长的蠕形遗迹, 潜穴宽度3, mm, 宽度较一致, 不分枝, 表面光滑, 于层面上任意弯曲, 相互穿插并在局部形成环圈, 呈表迹沟痕保存在泥灰岩层面上.
 | 图4 河南焦作地区下二叠统太原组中的遗迹化石(A--垂直剖面上的Teichichnus rectus, 产于L5; B--垂直剖上的Ophiomorpha nodosa, 产于太原组中段; C--层面上的Thalassinoides suevicus, 产于L6; D--层面上的Rhizocorallium isp., 产于太原组中段; E--垂直剖面上的Chondrites isp., 产于L4和L5; F--垂直剖面上的Nereites isp., 产于L4和L5; G--层面上的Gordia marina, 产于L9灰岩顶面)Fig.4 Ichnofossils of the Lower Permian Taiyuan Formation in Jiaozuo area, Henan Province |
讨论:Hantzschel(1962, 1975)认为Gordia是任意弯曲但并非蛇曲形遗迹.Ksiazkiewicz(1977)认为Gordia是觅食迹.该属为广相性化石, 在各种较平静的浅水和深水底层上都可出现(龚一鸣, 1994; Gong and Yuan, 2002), 一般认为, 其造迹生物是蠕虫类动物(Aceñ olaza and Buatois, 1993; McCann, 1993)或腹足类动物(杨式溥, 1984; 龚一鸣, 1987).
层位:焦作下二叠统太原组L8灰岩顶面.
条带迹 Ichnogenus Taenidium(Heer, 1877)
萨塔纳斯条带迹Taenidium satanassi(D’ Ales-sandro et Bromley, 1987)(图 5-B① )
描述:直--微弯曲型柱状进食潜穴, 沿层面分布, 不分枝, 可见长度4, cm左右, 宽0.3~1, cm, 无衬壁.具清晰的新月型回填纹构造, 宽度1, mm左右, 回填纹间距约0.1 cm.同一潜穴内新月形充填物宽度相等, 但比潜穴宽度小得多.潜穴回填纹为深灰色与浅灰色交替出现.
 | 图5 河南焦作地区下二叠统太原组中Zoophycos和Taenidium遗迹化石的形态和产状特征(A--层面上的Zoophycos villae, 产于L7灰岩中; B--层面上的Zoophycos brianeus, 产于L4灰岩中; C--层面上的Taenidium satanassi(① )与伴生的Zoophycos brianeus(② ), 产于L5灰岩的燧石层面上; D--垂直剖面上具有红色铁质填充物的Zoophycos villae, 产于L7灰岩中; E--垂直剖面上具有黑色泥质和硅质填充物的Zoophycos brianeus, 产于L4灰岩中)Fig.5 Morphology and occurrence of Zoophycos and Taenidium ichnofossils from the Lower Permian Taiyuan Formation in Jiaozuo area, Henan Province |
讨论:本遗迹种以其具有宽度相等的新月形回填充填物, 并且以宽度远远小于潜穴宽度为特征而与其他遗迹种相区别.以往的研究表明, 该遗迹属可出现在浅海沉积中(Buckman, 2001), 也可产生在河, 湖沉积中(Savrda, 2000; Melchor et al., 2006).
层位:焦作下二叠统太原组L5灰岩.
动藻迹 Zoophycos(Massalongo, 1855)
强健动藻迹Zoophycos brianeus(Villa, 1844)(图 5-B② , 5-C, 5-D)
描述:鸡尾形或掌形螺旋状潜穴, 具有密集连续的蹼状构造并由一中心轴向外辐射伸出.整体宽15, cm, 长大于17 cm.具有清晰边缘管, 边缘管直径为3, mm, 蹼宽1~2 mm.纵向剖面上可见清晰的新月形回填纹且平行层面或微倾斜分布.呈内迹保存在薄--中厚层灰岩或燧石层中, 风化的层面上螺旋形蹼状构造因潜穴充填物为深色灰泥岩或硅质泥岩而凸显在灰色灰岩或燧石层面上.
层位:焦作下二叠统太原组L2, L4, L5, L6, L7和L8灰岩.
维拉动藻迹Zoophycos villae(Massalongo, 1855)(图 5-A, 5-E)
描述:个体巨大的叶瓣形螺旋状潜穴, 具有清楚的蹼状构造, 螺旋状叶瓣(lobate spreite)呈长柱形或长椭圆形, 叶瓣宽5.5~8.5, cm, 长约21, cm, 厚度约1, cm; 具边缘管道, 边缘管宽3~5, mm, 边缘管间的弧形蹼状潜穴(新月形回填纹)宽3~4 mm.螺旋状叶瓣平行于层面分布, 纵向剖面上可见清晰的新月形回填纹.填充物为红褐色粉砂质泥岩或黑色泥岩, 与围岩(灰色灰岩)有明显区别.
讨论:遗迹种Zoophycos brianeus以鸡尾形或掌形的蹼状构造而与Zoophycos villae明显不同, 后者的蹼状构造呈叶瓣形螺旋状潜穴.遗迹种Zoophycos villae与Bromley和Hanken(2003)所建新种Zoophycos rhodensis的叶瓣蹼状构造相似, 但后者的整体形态为轮齿状而与之有别.动藻迹的造迹生物被认为是星虫类(Wetzel and Werner, 1981; Olivero, 2007), 螠虫动物类(Kotake, 1992), 多毛类, 肠鳃类和帚虫动物类(Ekdale and Lewis, 1991; Ekdale, 1992)等.该遗迹属既可出现在缺氧的半深海和深海环境, 也可出现在含氧的滨, 浅海和受限的边缘海(如半封闭海湾, 局限的潟湖和海湾等)环境, 尤其是在富含有机质的软底或固底, 含氧量低和水体循环性差的静水沉积底层环境中, Zoophycos往往大量出现(Wetzel and Werner, 1981; Ekdale et al., 1984; 杨式溥等, 2004).
层位:焦作下二叠统太原组各层灰岩中均有发育.
焦作地区太原组中发育的遗迹化石, 根据其组成与产状特征及其围岩的沉积特征, 可识别出3种组合:(1)Zoophycos-Nereites遗迹组合; (2)Zoophycos-Taenidium遗迹组合; (3)Ophiomorpha-Thalassinoides遗迹组合(图 6):
1)Zoophycos-Nereites遗迹组合:该遗迹组合以层内进食迹为主, 发育在L4和L5灰岩中, 主要由Zoophycos, Nereites, Chondrites, Teichichnus和较强生物扰动层组成, 遗迹化石丰度较高.这些遗迹化石所形成的底层往往与贫氧或缺氧环境及相对较深较平静的水体有关.有的学者认为, 具有细小潜穴管的Chondrites, Zoophycos和Nereites是贫氧环境的标志(Ekdale and Mason, 1988; 晋慧娟, 1999).与Zoophycos伴生的Teichichnus也是潮下带或浪基面之下的常见分子(Corner and Fjalstad, 1993; 王约, 2003).较强生物扰动层也往往产生和保存于潮下带或浪基面之下, 在浪基面之上易于被周期性的潮汐和波浪水流所侵蚀.该组合所处的L4深灰色灰岩中, 许多Zoophycos潜穴内均充填了黑色泥岩, 证明有可能与贫氧的底层环境有关.然而, Zoophycos潜穴的宿主岩中又含有很多类, 珊瑚, 海百合茎和腕足类实体化石, 说明当时底栖生物在含氧的浅海水体中生活.因此, 可以猜想, 此遗迹组合的造迹生物在贫氧或缺氧的底层内所开拓的掘穴系统与底层上含氧的水保持着联通, 造迹生物依靠海水中的氧气生活, 对孔隙水的缺氧条件具有较强的忍受力.因此, 该遗迹组合可解释为产生于海水相对较深且平静的潮下或浪基面之下的浅海沉积环境, 与Cruziana遗迹相的下部相带一致.由此可知, L4和L5灰岩应是焦作地区早二叠世海水达到最深时期的沉积记录.
 | 图6 河南焦作地区下二叠统太原组遗迹组合的沉积环境模式(Zoophycos; Te--Teichicnus; O--Ophiomorpha; Th--Thalassinoides; B--Bioturbational bed; Ch--Chondrites)1--Zoophycos--Nereites遗迹化石组合; 2--Zoophycos--Taenidium遗迹化石组合; 3--Ophiomorpha--Thalassinoides遗迹化石组合)Fig.6 Sedimentary environmental model of ichnoassemblages in the Lower Permian Taiyuan Formation in Jiaozuo area, Henan Province(N--Nereites; R--Rhizocorallium; Pa--Palaeophycus; P--Planolites; G--Gordia; Ta--Taenidium; Z-- |
2)Zoophycos-Taenidium遗迹组合:以大量Zoophycos成层出现为特色, 伴生近水平管状进食潜穴Planolites, 具有回填纹构造的进食管状潜穴Taenidium和与层面斜交的U形且具蹼状构造的Rhizocorallium等遗迹化石.该遗迹组合产生于除L4和L5灰岩以外的其他各层灰岩中, 遗迹化石丰度较高.前已述及, 这些遗迹化石几乎都是滨岸带低能和浅海沉积环境(Cruziana遗迹相)中的常见分子.与前一组合相比, 产生在浅海较深水沉积中的Nereites 和Teichichnus 以及反映贫氧条件的Chondrites 都没有出现, 也未发现成层的斑点状生物扰动构造, 并且其围岩都是浅灰色的碳酸盐岩.尤其是Rhizocorallium在其他地区也见于滨浅海及潟湖环境(杨式溥等, 2004).这些特征表明, 该遗迹组合形成于比前一组合水体更浅的浅海沉积环境.因而可解释其形成于近岸浅海沉积环境.
3)Ophiomorpha-Thalassinoides遗迹组合:该遗迹组合由螺旋状潜穴Zoophycos, 具蹼状构造的U形潜穴Rhizocorallium, 具分枝潜穴系统的Thalassinoides, 具瘤粒状垂直管状潜穴Ophiomorpha 和层面拖迹Gordia组成.主要出现在L5灰岩之上的碎屑岩段和L9灰岩顶部, 常见于泥质灰岩或钙质粉砂岩和薄层细砂岩中.该组合中产于L7灰岩的Zoophycos潜穴内常见有红色铁质充填物, 使潜穴内的新月形回填纹呈现红色, 明显区别于其他组合中的Zoophycos遗迹化石, 并且化石个体巨大, 说明其具有氧化环境的特征, 与本组其他灰岩层中的Zoophycos是不同的.该组合中的Thalassinoides, Ophiomorpha, Gordia和Rhizocorallium都是海岸附近潮间带和潮上带最常见的分子.生物挖穴活动的底层有软底也有半固结的泥质或钙质沉积物底层.反映该组合形成于周期性暴露的潮坪和浅水海湾--潟湖沉积环境.
1)焦作地区下二叠统太原组为一套海陆交互相沉积, 已识别出的遗迹化石有10属11种, 包括Zoophycos brianeus, Zoophycos villae, Teichichnus rectus, Nereites isp., Chondrites isp., Rhizocorallium isp., Thalassinoides suevicus, Ophiomorpha nodosa, Gordia marina, Taenidium satanassi和Planolites isp.等.这些遗迹化石多为全浮痕保存, 少数为半浮痕表迹保存, 其丰度较高.
2)根据遗迹化石的组成与产状特征及其围岩的沉积特征, 焦作地区下二叠统太原组的遗迹化石可识别出3种组合:(1)Zoophycos-Nereites遗迹组合, 出现在水体相对较深, 浪基面之下且沉积底层内氧气贫乏的远岸浅海沉积环境; (2)Zoophycos-Taenidium 遗迹组合, 产生于水体相对较浅, 浪基面之上的近岸浅海沉积环境; (3)Ophiomorpha-Thalassinoides遗迹组合, 发育于海岸带潮坪或海湾潟湖沉积环境.基于遗迹化石组合的分布特点, 提出了焦作地区太原组遗迹化石的沉积环境模式.
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|