
{kind=link}
{kind=link}
{kind=link}
白云岩有机成因模式:机制,进展与意义*
[李波 , 颜佳新, 刘喜停, 薛武强]
, 颜佳新, 刘喜停, 薛武强]
, 颜佳新, 刘喜停, 薛武强]
|
|

第一作者简介:李波,男,1986年生,现为中国地质大学(武汉)古生物学与地层学专业硕士研究生,从事碳酸盐岩沉积学研究.E-mail:libo_cug@163.com.
通讯作者简介:颜佳新,男,1962年生,教授,博士生导师,主要从事沉积地质学研究工作.通讯地址:湖北武汉中国地质大学(武汉)地球科学学院;邮编:430074;联系电话:027-67883504.E-mail:jxyan@cug.edu.cn.
白云岩是一种常见的碳酸盐岩,它广泛分布在古代碳酸盐岩台地中,却很少见于全新世沉积物中,这构成了一个未解之谜.近 20多年的研究成果表明,早期成岩过程中微生物参与的硫酸盐还原反应,甲烷生成和厌氧氧化反应以及有氧呼吸作用能够促进白云石的沉淀:细菌细胞和胞外聚合物( EPS)带有负电荷,能够聚集溶液中 Mg2+和 Ca2+;同时上述氧化还原反应产生 HC,提高了孔隙水中反应物的浓度;这样在细菌细胞周围形成一个对白云石超饱和的微环境,有利于白云石的沉淀.这是一种新的白云石成因模式,微生物活动和有机质是影响白云石形成最重要的因素,特殊的球状形态和碳同位素特征是鉴别有机成因白云石的重要标志.白云岩有机成因模式为认识地质历史时期大套白云岩的成因,探索"白云岩之谜"提供了新的思路.
About the first author Li bo,born in 1986,is a master candidate of palaeontology and stratigraphy in China University of Geosciences(Wuhan),and is engaged in carbonate sedimentology.E-mail: libo_cug@163.com.
About the corresponding author Yan Jiaxin,born in 1962,is a professor of China University of Geosciences(Wuhan),and is engaged in carbonate sedimentology.E-mail: jxyan@cug.edu.cn.
Dolomite is common in stratigraphic record.It is widely distributed in the carbonate platform in ancient times,however,rarely found in the Holocene sediments,which constitutes a mystery.The research achievements in last two decades indicate that sulfate reduction,methanogenesis, anaerobic methane oxidation and aerobic respiration in the early diagenetic process induce dolomite precipitation.Bacterial cells and extracellular polymeric substances(EPS)with negative charges could concentrate Mg and Ca ions in solution.At the same time,these three oxidation-reduction reactions generate bicarbonate and raise the concentration of reactants of the pore water.All of the above favor the formation of a micro-environment that is supersaturated respect to dolomite around cells,resulting in dolomite precipitation.This is a new dolomitization model.Microbial activity and organic matter are the most important factors for the precipitation of dolomite.The special spherical shape and carbon isotope characteristics of dolomite can be used to identify an organogenic origin.This organogenic dolomite model has significant implications for the origin of the massive dolomite in geological history and "Dolomite Problem" research.
白云石是岩石中一种常见的碳酸盐矿物, 它广泛分布在古代碳酸盐岩台地中, 却很少分布在全新世沉积物中, 这对"将今论古"原理是一个极大的挑战.所谓的"白云岩问题"指的是在常温常压条件下, 即使在对白云石过饱和的溶液中白云石也不能沉淀(Land, 1998).现代海水对白云石过饱和, 但是却很少有白云石沉淀, 因此白云岩成因问题被称为沉积学中的一个谜.
自从白云岩这个概念提出以来, 人们对其成因进行了超过一个半世纪的探索.然而, 迄今为止, 还未能全面了解它的形成机制.通过对现代白云岩和地质历史中白云岩的研究, 得出了多种白云岩成因模式, 如萨布哈模式, 卤水回流模式, 混合水模式, 毛细管浓缩模式, 热液白云岩模式等.虽然这些模式能解决局部地区白云岩的成因, 但是它们的应用存在很大的环境局限性, 无法用来解释古代岩石中所有白云岩的成因, 同时这些模式也没有给出令人满意的白云石沉淀的动力学机制.
上世纪80年代对半深海--深海沉积物的研究发现, 在远洋富含有机质的沉积物中存在较多的白云石颗粒(质量分数远大于1%)(Baker and Burns, 1985).基于对白云石碳同位素以及孔隙水剖面的研究, Compton(1988)提出了有机成因白云岩模式(organogenic dolomite model).90年代, 在对现代巴西 Lagoa Vermehla超盐度潟湖沉积物孔隙水进行研究之后, Vasconcelos 等(1995)提出了微生物白云岩模式(microbial dolomite model).该模式虽然与Compton(1988)提出的有机成因白云岩模式原理相同, 但因为强调了微生物新陈代谢活动在白云石沉淀过程中发挥的重要作用, 故开始受到了人们的广泛关注.最近几年, 利用厌氧, 需氧微生物培养实验探索白云石的沉淀机制成为一个研究热点.细菌培养实验和现场观察已证实了有微生物参与的有氧呼吸, 硫酸盐还原, 甲烷生成和甲烷厌氧氧化均可促进白云石的形成(Vasconcelos and McKenzie, 1997; Wright, 1999; Warthmann et al., 2000; Moore et al., 2004; Roberts et al., 2004; S’ nchez-Rom’ n et al., 2008; Kenward et al., 2009).由于有机质驱动的氧化还原反应普遍存在于碳酸盐沉积物的早期成岩过程中, 所以白云岩有机成因模式是碳酸盐岩沉积, 成岩作用研究中必须考虑的重要因素.关于白云岩的成因问题, 国内已有较多的综述文献, 主要总结了白云岩形成的多种模式以及取得的一些研究进展等(王勇, 2006; 张学丰等, 2006), 但涉及到白云岩有机成因模式的介绍并不系统.文中着重于有机成因白云岩的形成机制介绍, 以反映这一领域的最新研究进展.
要解决"白云岩成因"问题, 首先必须了解阻止白云石形成的障碍, 即到底是哪些因素抑制了地表温压条件下白云石的形成.白云石的形成方式主要有原生沉淀和次生交代, 化学反应式分别如下:
C(aq)+M(aq)+2C(aq)=CaMg(CO3)2(s) (1)
2CaCO3(s)+M(aq)=CaMg(CO3)2(s)+C(aq) (2)
式(1)为原生沉淀白云石的化学反应式, 平衡系数K=a[C]* a[M]* (a[C])2/a[CaMg(CO3)2]. 其中a[x]代表溶液中离子的活度(固体或纯液体为1).Moore等(2004)根据孔隙水中离子活度计算的K值大约为10-16.1~10-16.4.现代海水中C, M和C的离子活度积为10-15.01(Warren, 2000), 比Moore等(2004)计算的K值大一个数量级, 所以理论上现代海水对白云石是过饱和的.然而, 白云石却很少沉淀, 说明现代海水中白云石的形成主要受化学反应动力学的控制, 而不是受热力学的控制.
式(2)为交代成因白云石的化学反应式, 平衡系数K=a[M]/a[C]=0.67.现代海水的Mg/Ca摩尔比为5.2, 远大于0.67, 因此理论上现代碳酸盐沉积中的白云岩化现象应广泛存在, 但实际上并非如此.同样说明反应的动力学阻止了白云岩化的进行.
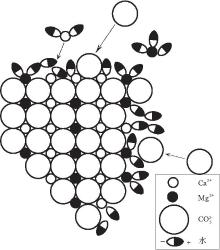
众所周知, 反应动力学问题主要是反应速率问题, 影响反应速率的因素主要是反应物的浓度, 反应的温度和"催化剂"的存在.在地表温压条件下, 影响白云石形成的浓度因素包括以下3个方面:(1)镁离子.海水中部分Mg2+, Ca2+是以水化合物或络合物的形式存在(Pytkowicz and Hawley, 1974), 而Mg2+与水之间的静电引力比Ca2+与水之间和Mg2+与C之间的静电引力大, 低温条件下Mg2+不易进入到白云石晶格中(图 1)(Lippmann, 1973, 1982), 所以在常温常压环境中, Mg2+ 的水合作用阻止了过饱和溶液中白云石的沉淀; (2)碳酸根离子.溶液中C的浓度和活度都非常低(Garrels and Thompson, 1962), 如现代海水中HC与C 的浓度加起来才2.27× 10-3 mol/L, 沉淀白云石所能利用的C十分少, 所以白云石难以沉淀.(3)硫酸根离子.溶液中存在的S是限制低温白云石形成的主要动力学因素, 即使是非常低浓度的S的存在, 也会抑制白云石的形成(Baker and Kastner, 1981; Kastner, 1984).因为在溶液中, Mg2+常与S结合形成中性强离子对MgS, 使溶液中可利用的Mg2+较少; 同时MgS会被吸附到正在生长的晶体表面, 减缓白云石的生长速率(Slaughter and Hill, 1991).因此在地表温压条件下, 若没有"催化剂"的存在, 白云石只能在高温下形成, 如大多数人工合成白云石实验所证明的那样(Land, 1998; Arvidson and Mackenzie, 1999).
 | 图1 阳离子的水合作用阻止白云石形成示意图(据Lippman, 1973)Fig.1 Sketch map of strong hydration of cation preventing formation of dolomite(after Lippman, 1973) |
近20年的研究发现, 微生物参与的氧化还原反应, 在早期成岩阶段的白云石形成过程中, 起到了"催化剂"的作用:它增加了孔隙水中的反应物浓度, 从而增大了白云石沉淀的反应速率, 有利于白云石的沉淀.根据微生物参与的反应的不同, 可以将有机成因白云石的形成分为3种模式:硫酸盐还原模式, 甲烷生成和厌氧氧化模式以及有氧呼吸作用模式.
S的存在会抑制白云石的沉淀(Baker and Kastner, 1981; Kastner, 1984), 因此降低孔隙水中的S浓度, 显然有利于白云石的沉淀.如深海缺氧, 富含有机质的沉积物中, 普遍发生的硫酸盐还原反应就可以降低孔隙水中的S浓度(Baker and Kastner, 1981):
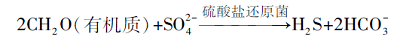 (3)
(3)
这个过程需要硫酸盐还原菌(SRB)的参与.如果孔隙水中存在可利用的Fe2+, 将会与式(3)中生成的H2S反应生成黄铁矿(Berner, 1980), 所以经常可以见到原生的草莓状黄铁矿与有机成因白云石共生:
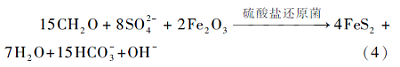
而蛋白质的脱氨基作用可产生氨气(Slaughter and Hill, 1991):
NHCO+H2O→ CO2+NH3(5)
形成的氨气与孔隙水反应形成氨水:
NH3+H2O→ N+OH-(6)
从以上反应式可以看出, 硫酸盐还原菌在对有机质降解过程中, 消耗了孔隙水中的S, 同时生成了HC, 提高了溶液的pH值和白云石的饱和度.这有利于克服白云石沉淀反应的动力学限制, 从而沉淀出白云石:
Mg2++Ca2++2HC+2OH-→ CaMg(CO3)2+2H2O (7)
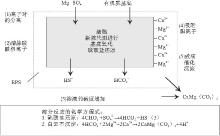
最近10多年, 硫酸盐还原菌培养实验有力地支持了上述白云石的沉淀机制.在培养实验中, 利用从Coorong潟湖和巴西Lagoa Vermelha潟湖中提取的硫酸盐还原菌进行培养, 沉淀出与潟湖沉积中形态完全一样的白云石颗粒.而在对照的实验中, 利用死亡的硫酸盐还原菌或不使用硫酸盐还原菌, 并没有发现白云石的沉淀(Wright, 1999; Warthmann et al., 2000; van Lith et al., 2003a; Wright and Wacey, 2005; Wacey et al., 2007).在碳酸盐还原菌培养实验结束后, 培养液中pH值明显增大, 而硫酸根离子浓度显著降低, 甚至为零.使用扫描电镜对实验过程进行观察发现, 白云石颗粒绝大多数形成于细菌聚集的地方.这说明硫酸盐还原菌在新陈代谢的过程中, 改变了细胞周围微环境的水化学条件, 营造出了一个有利于白云石沉淀的环境, 促进了白云石的沉淀.因为硫酸盐还原菌在新陈代谢过程中分泌出胞外聚合物(EPS), 消耗了S, 使得MgS强离子对中的Mg2+被释放, 溶液中Mg2+活度增大, 而这些阳离子(Ca2+和Mg2+)被带负电荷细菌的细胞壁吸附; 同时反应生成的HC增大了溶液的碱度; 这样在细菌细胞周围聚集了 Ca2+, Mg2+和HC, 形成了一个对白云石高度过饱和的微环境, 使得白云石在EPS内部的细菌细胞壁上成核(图 2)(van Lith et al., 2003a).
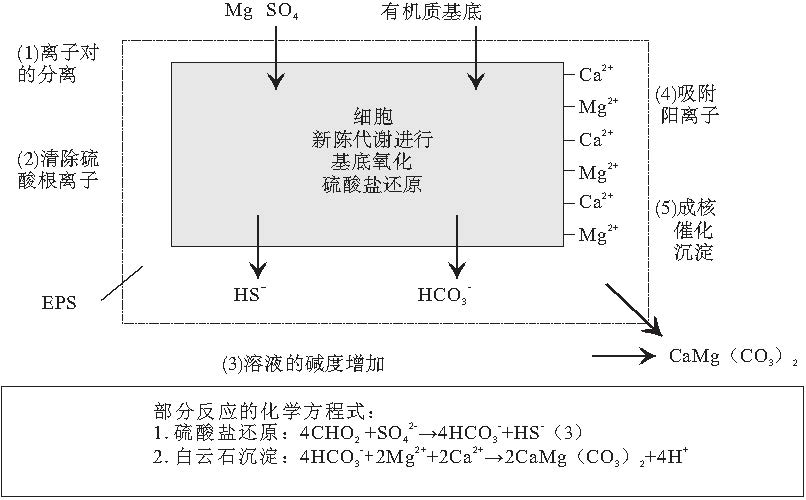 | 图2 硫酸盐还原菌促进白云石形成的模式图(据van Lith 等, 2003a)Fig.2 Schematic model showing role of sulphate-reducing bacteria in dolomite formation(after van Lith et al., 2003a) |
产甲烷细菌是一种严格厌氧古细菌, 广泛存在于各种极端的环境中.Roberts等(2004)在野外和实验室实验中均发现了异化型铁还原菌和/或产甲烷古菌在淡水中克服反应动力学条件以促进白云石沉淀的现象.Kenward等(2009)进一步证明了上述白云石沉淀发生在异化型铁还原菌停止活动而产甲烷古菌的新陈代谢活动繁盛以后, 证明产甲烷菌可以作为一类能够促进白云石在低温下沉淀的微生物.
甲烷的形成主要有两种方式:一种方式是自养产甲烷菌(autotrophic methanogens)还原CO2(Thauer, 1998):
另一种方式是食醋酸产甲烷菌(Acetoclastic Methanogens)利用醋酸发酵(Whiticar, 1999):
CH3COO-+H2O 
自养产甲烷菌消耗了溶液中的CO2, 有利于pH值的增大, 而食醋酸产甲烷生物在发酵时产生了HC, 提高了溶液的碱度, 两者均有利于白云石的沉淀.
甲烷形成后向上扩散, 在硫酸盐还原带和甲烷生成带之间形成了一个过渡带:硫酸盐--甲烷过渡带(SMT).在这个过渡带中, 甲烷的厌氧氧化过程(AOM)将CH4氧化为CO2, S被还原.该过程主要受古细菌和硫酸盐还原菌微生物群团的调节(Boetius et al., 2000).这些微生物群团各自以独立的方式进行反应, 整个过程的净反应如下:
CH4+S 
该反应消耗了硫酸盐, 使孔隙水中的S浓度降低, 同时生成了HC, 增大了孔隙水中反应物的浓度, 因而有利于白云石的形成.地质历史时期与甲烷厌氧氧化作用有关的自生白云石通常和海底甲烷水合物的释放有关, 如日本琉球群岛南部的Kuroshima Knoll地区中的烟囱状白云岩(Takeuchi et al., 2007).前人研究表明, 现代大洋沉积物剖面中大量存在甲烷厌氧氧化作用, 并且在许多取样站位发现了自生白云石颗粒(Middelburg et al., 1990; Moore et al., 2004; Meister et al., 2007).
在许多现代地质环境中, 比如巴西的Brejo do Espinho 潟湖, 沉积物顶部含氧微生物席中白云石颗粒的发现表明, 需氧微生物的新陈代谢活动在白云石的沉淀过程中可能同样发挥着重要的作用(S’ nchez-Rom’ n et al., 2009b).Rivadeneyra等(1993)利用实验证明, 喜盐性细菌可以调节白云石的沉淀, 同时伴随着方解石, 高镁方解石和一水方解石的沉淀.这表明异养微生物在进行有氧呼吸的过程中, 可以促进白云石在低温下沉淀.S’ nchez-Rom’ n等(2009b)将从巴西Brejo do Espinho潟湖中的含氧微生物席上部分离出来的喜盐性需氧细菌Virgibacillus marismortui和Marinobacter sp.在实验室含氧的条件下进行培养, 成功沉淀出白云石和水碳镁石, 证实了沉积物--水界面附近的有氧呼吸作用可以促进白云石沉淀.
有机质在海水中沉积下来后, 在有溶解氧存在的情况下, 会发生有氧氧化作用.这一过程需要需氧微生物的参与.需氧微生物在分解有机质的过程中, 释放出CO2和N, 提高了溶液的碱度和pH值, 反应可以简单表示如下:
C3O3H7N+2.5O2 
微生物通过有氧呼吸降解10× 10-3mol/L的氨基酸会导致溶液的pH值从7.0降到6.3.考虑到溶液中的重碳酸盐离子对pH值的缓冲作用, 理论上pH值不会有大幅度的降低.但在微生物培养实验中, 溶液的pH值反而升高到7.9, 这可能与反应产生的CO2的脱气作用有关(大于70%的CO2发生脱气)(S’ nchez-Rom’ n et al., 2009b).有氧呼吸作用产生了大量的HC, 使得在细菌细胞周围的微环境中, 形成白云石的反应物浓度大幅度地提高, 有利于克服白云石沉淀的反应动力学条件.
沉积物中富含有机物是形成有机成因白云石最重要的因素之一, 因为白云石中部分碳来源于有机质的降解.有机质经过氧化还原反应降解形成的HC极大地增加了孔隙水中沉淀白云石所需要的反应物浓度, 有利于白云石的沉淀.所以在现代沉积中, 有机成因白云石颗粒通常出现在有机质富集的层位(Kelts and McKenzie, 1982; Vasconcelos and McKenzie, 1997).当然, 微生物对有机质的降解可能会使原先富含有机质的沉积物在成岩之后并不富有机质.另外, 沉积物中来源于植物, 浮游动物和藻类的有机质的降解, 特别是蛋白质酶的破坏, 提高了孔隙水的pH值(反应式(3), (5), (6), (11)), 而pH值的升高有利于HC向C的转换, 增大了孔隙水中的C的活度, 有利于白云石的沉淀(Slaughter and Hill, 1991).
无论是有氧呼吸, 硫酸盐还原还是甲烷生成和甲烷厌氧氧化过程, 有机质的降解都需要微生物的参与.目前培养实验已经证实的, 可以促进白云石沉淀的微生物有喜盐性需氧微生物, 硫酸盐还原菌和产甲烷菌(表 1), 而在所有加入死亡菌种或不加菌种的对照实验中并未发现白云石的沉淀.但这并不表示上述3种类型微生物的所有属种都能促进白云石的形成, 在相似的条件下, 有些硫酸盐还原菌在培养实验中可以促进白云石的沉淀, 但另外一些硫酸盐还原菌只能促进方解石的沉淀(van Lith et al., 2003a).细菌培养实验证实:在产甲烷菌繁盛之后, 培养液对白云石的的饱和度由最初的19.40倍增加到2330.77倍(Kenward et al., 2009).这是因为微生物降解有机质产生了大量的HC离子, 而细胞壁和胞外聚合物质对金属阳离子的吸附作用使得Mg2+, Ca2+离子聚集, 所以在细菌周围产生了一个对白云石高度过饱和的微环境.另外, 利用扫描电镜观察细菌培养沉淀白云石的实验过程发现, 白云石的成核通常位于微生物细胞表面或排泄到细胞外的聚合物质上, 说明微生物能为白云石提供成核地点, 这对于白云石的形成是至关重要的(van Lith et al., 2002; van Lith, 2003a, 2003b; Roberts et al., 2004; Wright and Wacey, 2005; Bontognali et al., 2008; S’ nchez-Rom’ n et al., 2008, 2009a, 2009b).
| 表1 促进白云石沉淀的微生物种类以及培养液的化学成分 Table 1 Microorganism types and chemical composition of solution that promote dolomite precipitation in culture experiments |
在微生物新陈代谢活动的调节下, 白云石不仅能在Lagoa Vermelha, Brejo do Espinho以及Coorong潟湖这类高盐度环境中形成(Vasconcelos and McKenzie, 1997; Wright, 1999; van Lith et al., 2002), 而且在低盐度湖水甚至淡水中, 同样可以沉淀(表 1).Roberts等(2004)报导了淡水中微生物新陈代谢活动直接导致白云石沉淀的实例.在受石油污染的缺氧地下水环境中, 异化型铁还原菌和产甲烷菌先风化玄武岩, 释放出Mg2+, Ca2+和Fe2+, 提供了形成白云石沉淀所需要的阳离子, 然后白云石在细菌细胞表面成核.于炳松等(2007)在青海湖底沉积物中首次发现了球状, 椭球状白云石集合体和伴生的草莓状黄铁矿, 其与近代近海地区高盐度潟湖和萨布哈中发现的微生物成因的白云石极为相似, 表明有机成因白云石的形成并不局限于高盐度环境中, 极大地拓展了人们对有机成因白云石形成环境的认识.
在没有微生物存在的低温条件下无法合成白云石(Land, 1998).将实验温度提高至150 ℃ , 白云石才可以从饱和溶液中沉淀(Arvidson and Mackenzie, 1999).可见在无微生物参与的情况下, 温度对白云石的成核起着至关重要的作用.但在微生物新陈代谢活动的促进下, 白云石成核温度可大幅降低(< 50 ℃ )(表 1).显然微生物在白云石的沉淀中充当了一个类似"催化剂"的角色.虽然在地表温压条件下微生物的活动可以促进白云石的沉淀, 但温度(< 50 ℃ )对白云石形成的影响仍然存在.相同条件下的细菌培养实验发现, 白云石晶体在 35 ℃ 时的生长速率比 25 ℃ 快(S’ nchez-Rom’ n et al., 2008, 2009a), 表明温度的升高能够增大白云石的沉淀速率.Vasconcelos等(2005)进行的厌氧微生物培养实验发现, 当培养温度为25 ℃ , 30 ℃ 和40 ℃ 时, 沉淀物为富钙白云石和方解石, 而将温度升高到45 ℃ 时, 仅沉淀白云石, 并且在40 ℃ 和45 ℃ 时, 形成的白云石是完全有序的(Mg/Ca摩尔比为50:50).这表明温度的升高有利于白云石的形成, 而白云石的有序度与培养温度存在正相关性, 其可能与细菌生存的最适宜温度有关(37~38 ℃ ).虽然较高的温度更加有利于白云石的形成, 但是温度太高会抑制微生物的活动, 导致微生物死亡, 反而会抑制有机成因白云石的形成.
传统的白云岩成因模式常需要提供额外的富 Mg2+流体(如蒙脱石的伊利石化释放的 Mg2+), 或者是溶液中Mg/Ca摩尔比的提高机制(如毛细管蒸发).但是在现代远洋环境中, 沉积物沉积下来之后, 往往很少有其他富 Mg2+流体的加入.上覆海水向沉积物的扩散以及早期成岩过程中高镁方解石向低镁方解石的转换才是最大的Mg2+来源(Mitchell and Horton, 1995; Dix, 1997; Yan et al., 2005).质量平衡计算表明, 只要沉积物埋藏在沉积物/水界面以下几十米之内, 上覆海水向沉积物的扩散作用就可以提供足够的 Mg2+(Compton and siever, 1986).在微生物作用下, 正常海水的Mg2+浓度以及Mg/Ca摩尔比足以使白云石沉淀(表 1).即使在Mg/Ca摩尔比小于1的地下水中(Roberts et al., 2004), 白云石也能沉淀.这得益于微生物的细胞和分泌的胞外聚合物吸附金属阳离子的能力, 使得 Mg2+, Ca2+等金属阳离子在微生物细胞周围的微环境中富集.
综上所述, 碳酸盐沉积中的有机质和微生物活动在白云石的沉淀中发挥着最为重要的作用, 控制了白云石的形成, 应该在今后的研究中加以重视.盐度, 镁离子对白云石沉淀的影响并不显著, 超高的盐度和富 Mg2+流体并不是必须的.温度的升高对白云石的形成影响巨大, 是一个关键性的因素, 但在"催化剂"的存在下, 其仅仅影响了白云石的成核速率和晶体有序度.
白云岩有机成因模式的现代实例研究较为成熟, 其在古代的应用也为大家所关注.目前, 有机成因白云石的识别特征主要包括两方面:形貌学特征和碳同位素特征.
形貌学(显微组构)的研究可以深入了解生物矿化过程, 同样适用于了解白云石沉淀的环境条件(Riding, 2000).无论是现代的超盐度潟湖中原地沉淀的有机成因白云石(Vasconcelos and McKenzie, 1997; Wright, 1999; Vasconcelos et al., 2005), 还是利用微生物培养实验沉淀出来的白云石(Warthmann et al., 2000, 2005; Wright and Wacey, 2005; S’ nchez-Rom’ n et al., 2008, 2009a, 2009b), 无一例外具有球粒状, 哑铃状或是花椰菜状的外形以及放射纤维状的内部构造(图 3), 与高温实验形成的菱形的白云石完全不同(Arvidson and Mackenzie, 1999).
在古代岩石的记录中, 同样发现了许多类似的球状白云石, 如科威特始新统到第四系岩石中的原生球状白云石胶结物(Gunatilaka et al., 1987; Gunatilaka, 1989), 印度西北部晚更新世碳酸盐岩台地中的球状白云石(Rao et al., 2003), 以及在地表潜水层中形成的球状白云石胶结物(Khalaf, 1990; El-Sayed et al., 1991; Spö tl and Wright, 1992).另外, Nielsen 等(1997)将比利时东部喀斯特化的Dinantian白云岩序列顶部的球粒状白云石解释为微生物成因.Lee和Golubic(1999)把中国华北中元古界高于庄组中的球粒状白云石解释为微生物成因.Mastandrea等(2006)在意大利南部上三叠统微生物白云岩的表面发现了钙化的球状硫酸盐还原菌.Buczynski和Chafetz(1991)认为哑铃状形态是细菌诱导沉淀的碳酸盐矿物的独有特征, 可以用来鉴别岩石记录中的生物成因的碳酸盐矿物.
位于巴西Brejo do Espinho 潟湖沉积物柱5, cm处的白云石为球状无序的富钙白云石, 但往下至21, cm深处, 白云石有序度增加, 为自形的菱形白云石(S’ nchez-Rom’ n et al., 2009b).在Lagoa Vermelha潟湖同样存在这种随沉积物埋深增加, 球状无序白云石向菱形有序白云石转变的现象.用稀盐酸对这种菱形的白云石颗粒表面进行刻蚀处理后, 内部仍然为球状形态(Vasconcelos and McKenzie, 1997).Vasconcelos和McKenzie(1997)认为球状白云石成核之后, 随埋深的增加, 会经历一个成熟化(ageing)的过程:富钙球状白云石颗粒表面以无机的形式继续生长, 球状白云石周围的空间被无机的白云石覆盖充填, 原先圆滑的表面变平整, 球状形态转变成菱形, 同时有序度增加, 转变为菱形有序白云石.
海相碳酸盐岩中的碳主要来源于海水中溶解的无机碳(δ 13C值接近于0‰ PDB), 先期形成的碳酸盐溶解后形成的碳以及大气水中溶解的碳.除了以上3个碳源之外, 有机成因白云石还应包括有机质在埋藏时经受不同氧化还原反应所形成的碳, 因此也具有与同时期其他碳酸盐岩不同的碳同位素特征.不同碳源进入白云石晶格中的比例决定了有机成因白云石的碳同位素值, 其分布在-61‰ ~+26‰ PDB之间(Compton, 1988; Jø rgensen, 1989; Sample and Reid, 1998; Mazzullo, 2000; Aloisi et al., 2002; Meister et al., 2007; Takeuchi et al., 2007).
 | 图3 有机成因白云石的形态特征(A--细菌和球状白云石紧密共生(S’ nchez-Rom’ n et al., 2008); B--白云石球体内部的放射状构造(S’ nchez-Rom’ n et al., 2009a); C--哑铃状白云石球体聚合形成花椰菜状构造(Warthmann et al., 2000); D--正在生长的哑铃状的白云石球体, 箭头指示胞外聚合物粘附在白云石表面(S’ nchez-Rom’ n et al., 2009b); A--D均为扫描电镜照片)Fig.3 Morphological characteristics of organogenic dolomite |
与有氧呼吸, 硫酸盐还原作用有关的白云石中的部分碳来源于细菌降解有机质形成的HC, 其δ 13C值为-25‰ PDB左右(Claypool and Kaplan, 1974).这部分碳的加入可能导致白云石的碳同位素值出现较大负偏(δ 13C值为-25‰ ~0‰ PDB; Hein et al., 1979; Kelts and McKenzie, 1982; Hennessy and Knauth, 1985; Burns and Baker, 1987; Burns et al., 1988; Dix, 1997; Vasconcelos and McKenzie, 1997; van Lith et al., 2003a; Pierre and Rouchy, 2004; S’ nchez-Rom’ n et al., 2009b), 其可作为有机质降解形成的碳进入到白云石中的证据.生物成因的甲烷具有极低的碳同位素值负偏特征(δ 13C值平均约为-60‰ PDB; Whiticar, 1999), 而甲烷厌氧氧化作用形成的白云石中的部分碳来自于甲烷, 因此与甲烷厌氧氧化作用有关的白云石的碳同位素值可能会强烈负偏(δ 13C在-61‰ ~-25‰ PDB; Jø rgensen, 1989; Aloisi et al., 2000, 2002; Meister et al., 2007; Takeuchi et al., 2007), 这可作为鉴别与甲烷厌氧氧化作用有关的白云石的标志.甲烷生成时, 碳同位素分馏, 重碳在HC富集(Whiticar, 1999).部分含重碳的HC进入白云石中, 导致白云石碳同位素值可能呈现出较大正偏(δ 13C在0~+26‰ PDB; Friedman and Murata, 1979; Pisciotto and Mahoney, 1981; Kelts and McKenzie, 1982; Mazzullo et al., 1995; Sample and Reid, 1998; Pufahl and Wefer, 2001; Pierre and Rouchy, 2004), 这成为识别甲烷生成作用参与白云石形成的标志.
在现代地表环境条件下, 白云石可以在多种环境中生成:从浅海到深海; 从低盐度内陆湖泊到高盐度潟湖.由于白云石的沉淀并不局限于强烈蒸发环境, 并不需要Mg/Ca摩尔比升高和淡水注入等额外条件, 关键是必须有微生物活动的参与, 因此白云岩的有机成因模式对认识白云岩的成因具有深远的意义.以前由于忽视了微生物活动在白云石形成中的作用, 过于强调蒸发, Mg/Ca摩尔比的提高或者是淡水的注入等因素的影响, 因而提出了相应的白云石成因模式.现在看来, 这些因素的作用被高估了.在应用这些模式的时候, 要特别关注微生物的活动.比如提出萨布哈白云岩成因模式的波斯湾Abu Dhabi潟湖, 这里的白云石通常是出现在有机质富集的层位, 大多数为直接沉淀, 并非交代文石形成(Wenk et al., 1993); 同时白云石的碳同位素值(0‰ , PDB)比同一位置的文石(3‰ PDB)低, 表明白云石中部分碳来源于有机质的降解(Baltzer et al., 1994), 指示微生物通过降解有机质而参与了白云石的形成.
富有机质的沉积物堆积形成以后, 势必在早期成岩阶段发生有氧呼吸, 硫酸盐还原, 甲烷生成以及厌氧氧化等.可想而知, 古代海相碳酸盐沉积中, 这种有机成因的白云石应该极为普遍.事实上, 古代海相沉积中确实普遍存在这种成因的白云石, 只不过常被研究者忽视了, 或者被错误地解释为深埋藏成因的白云石.
如意大利南部三叠系诺利克阶广泛发育厚层的叠层石, 矿物成分几乎全部是隐晶质的富钙白云石, 形成于干旱气候条件下的潮汐环境中.早先研究一般认为这类白云岩的形成与超盐度卤水在密度的驱动下循环有关(Iannace and Frisia, 1994; Haas and Demé ny, 2002).最近几年, 该成因模式受到了质疑.扫描电镜研究发现, 在叠层石的微晶纹层(特别是暗纹层)中, 半自形或自形白云石晶体的晶粒孔和白云石晶体内部保存着纳米级--微米级的球粒状细菌(矿物成分为白云石), 席状的矿化的胞外聚合物以及残余的有机质, 表明微生物参与了该白云岩的形成(Mastandrea et al., 2006; Perri and Tucker, 2007).
虽然经过百余年的探索, 仍未完全解决白云岩的成因之谜, 但是近20多年的研究发现, 早期成岩过程中有微生物新陈代谢活动参与的氧化还原反应能够促进白云石在地表温压条件下发生沉淀, 这为解决"白云岩成因问题"提供了新的思路.正如前面介绍的, 微生物新陈代谢活动的广泛分布, 可以在多种环境中促进白云石的沉淀.因此以后在研究地质历史时期的白云岩成因时, 必须考虑微生物这一因素.有机成因白云岩模式的提出势必会引起"白云岩成因问题"研究的新热潮, 并在以下几个方面的研究中可能会有所突破:(1)结合现代微生物培养实验, 在古代白云岩中寻找更多的微生物作用证据, 比如微量金属元素的富集或是特殊的同位素分馏作用; (2)进一步观察微生物沉淀白云石的过程, 特别是白云石成核的最初始阶段; (3)进一步确定有机成因白云石形成的影响因素; (4)继续开展青海湖等实地有机成因白云石的研究工作.随着研究工作的深入, 白云岩的有机成因模式必定会在解决"白云岩成因问题"中发挥出更加重要的作用.
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|