
{kind=link}
{kind=link}
关于 “超微细菌” 的争论:灰岩成因研究的重要驱动力*
[梅冥相 ]
]
]
|
|

作者简介:梅冥相,男,1965年生,中国地质大学(北京)教授、博士生导师,主要从事沉积学和地层学研究工作。E-mail: meimingxiang@263.net。
在高倍( >50000×)扫描电子显微镜( SEM)、原子显微镜( AFM)或透射电子显微镜( TEM)下,那些在岩石和矿物中所见到的纳米级别的物体常常吸引了地质学家们的注意。这些物体大小在 25~300 nm之间,多呈球状、棒状或卵圆状;对这些物体的成因,存在不同的看法而导致了剧烈的争论。因为这些物体与真菌中的球菌和杆菌总体相似,而且它们总是以链状或束状的形式产出,所以被解释为细菌的“侏儒形式”或“超微细菌”,或者是它们的化石对等物即“超微化石”。对于超微细菌的存在,来自于地质学角度的反对聚焦在这个级别的特征物可能存在多重成因,如微矿物包裹体或结晶学的边缘效应;从生物学角度看,对于独立生命所必需的所有构成来讲,直径小于 140 nm的球状体实在是太小了。因此,产生于世纪之交的这场争论,迄今还在持续着。尽管这场争论还没有一个肯定的结论,但是,它却成为研究灰岩成因的重要驱动力。
Those nanometer-scale spherical-,rod-,or ovoid-shaped objects in the 25~300 nm range,which are visible in rocks and minerals by high-magnification(>50000×) scanning electron microscope(SEM),atomic force microscope(AFM),or transmission electron microscope(TEM)analyses,attract the geologists’ attention.For the origin of these nanometer-scale objects,different views result in a strong disputation.Because of the general resemblance between these objects and eubacterial cocci or bacilli,and because of their tendency to occur in chains or clusters,it was initially proposed that the objects were “dwarf forms” of bacteria,“nannobacteria”,or their fossilized equivalents,“nannofossils”.The focus of geologic objections to the actuality of nannobacteria is that features of this scale may have many possible origins, e.g.,micromineral inclusions or crystallographic edge effects;further,a sphere <140 nm diameter is biologically thought to be too small to contain all the components necessary for an independent life.Therefore,this disputation that occurred at the turn of centuries and had continued thus far became an important impetus promoting the studies of the origin of limestones although an affirmative conclusion remains uncertain.
在地质学和生物学主题中, “ 超微细菌(nannobacteria)” 是否存在, 是世纪之交最有争议的科学命题之一。首先在热泉的碳酸盐矿物中, Folk(1993a)运用高倍电子显微镜(SEM)所发现的那些大小为0.025~0.3 μ m(25~300 nm)的球形、棒形或卵圆形的物体, 在被解释为“ 超微细菌” 及其化石的对等物— — “ 超微化石” (nannofossils)之后, 一直吸引了地质学家和生物学家的关注, 产生了较为激烈的争论(Kirkland et al., 1999)。随着Folk和Lynch(2001)将这些假定的“ 超微细菌” 与鲕粒及硬底的形成联系起来, 关于“ 超微细菌” 的争论大大地促进了碳酸盐沉积学的研究, 尤其是关于在微生物细胞物质上的钙碳酸盐成核作用的研究。所以说, 神话般的关于“ 超微细菌” 的争论, 成为研究灰岩成因的重要驱动力。
在岩石和矿物中, 那些纳米级别大小(25~300 nm)的球状、棒状或卵圆状物体, 常见于扫描电子显微镜(SEM)、原子显微镜(AFM)或透射电子显微镜(TEM)分析之中。因为这些物体与真菌中的球菌和杆菌形态上总体相似, 而且它们总是呈链状或束状产出, 所以一开始就被解释为是细菌的“ 侏儒形式” 或超微细菌(Folk, 1992, 1993a), 或者是它们的化石对等物, 即“ 超微化石” (McKay et al., 1996)。
这些纳米级别的物体, 还曾经在陆生碳酸盐岩、硫化物、氧化物、黏土和其他硅酸盐矿物之中被发现, 并被解释为石化了的微生物(Folk, 1992, 1993a, 1993b; Pedone and Folk, 1996; Sillitoe et al., 1996; Folk and Lynch, 1997; Vasconcelos and McKenzie, 1997; Chafetz et al., 1998); 更为重要的是, 在星外岩石, 包括火星陨石ALH84001以及在碳质陨石之中, 也发现了类似的球状物体(McKay et al., 1996; Folk and Lynch, 1997, 1998; Folk et al., 1998)。相似的纳米级别的球状物还被发现在哺乳动物的泄液中, 而且也出现在生物体外和体内的细胞毒素之中(Å kerman et al., 1993; Ciftcioglu et al., 1997; Kajander et al., 1997; Ciftcioglu and Kajander, 1998)。这些物体, 可能在组织钙化作用中起着作用(Kajander and Ciftcioglu, 1998), 而且在人的牙垢以及动脉血液的血小板中已经检测到。这些发现, 激发了关于古代微生物生命形式识别的激烈争论, 因为纳米级别物体的生物学属性及其存在, 曾经受到地质学和微生物群落证据的挑战。
对于超微细菌的存在, 地质学家反对的焦点是这个级别的特征物可能存在许多成因, 如微矿物包裹体或结晶学的边缘效应(Bradley et al., 1997); 另外一个经常反对的声音是, 纳米级别的物体可能是在进行SEM分析时, 所制作的导电重金属涂层的人工复制品。后一种可能性已经得到排除, 因为随着试验的进一步进行, 使用的涂金的程序避免了这样的人工复制品的形成。所以说, 在沉积岩中那些纳米级别的球状特征物确实是客观存在的(Peters, 1985; Folk and Lynch, 1997)。
关键的问题还是微生物群落的生物学属性, 这主要集中在“ 超微细菌” 本身, 这些纳米级别物体的大小经常只有典型细菌的千分之一。在直径上小于200 nm的少量细菌样本细胞, 可以作为微生物中细菌大小的下限, 这一点已经在很多教科书中报道过, 也包括了超微细菌(Kajander and Ciftcioglu, 1998)和超显微细菌(Torella and Morita, 1981)。对于识别超显微细菌的有说服力的证据, 已经由Morita(1997)提出。土壤中的超显微细菌有些只有80 nm, 其内部结构已经用TEM观察到(Bae et al., 1972), 而且海洋中的超显微细菌也曾经被描述过(Button et al., 1993)。
更为重要的是, 研究和观察表明, 当受到剧烈的应急压力时, 细菌才减小它们的大小和细胞的体积(Torella and Morita, 1981; Morita, 1988; Cusack et al., 1992); 再者, 细菌培养基是从200 nm的过滤器的过滤液中产生的(Hood and MacDonell, 1987; Kajander and Ciftcioglu, 1998), 但是, 相似的技术不会运用50 nm的过滤器来产生培养基(Kajander et al., 1997)。所以说, 对于独立生命所必需的所有构成来讲, 直径小于140 nm的球状物实在是太小了(Maniloff, 1996, 1997; Nealson, 1997), 这一点应该是怀疑超微细菌及其化石对等物— — 超微化石客观存在的关键; 基于生物学家认为200~300 nm是微生物可生存的大小的下限(Maniloff, 1996, 1997; Nealson, 1997), 对于这种已经被地质学家描述为超微细菌的特征物, 以及那些被发现在沉积岩中的50~200 nm大小的球状物, 就需要另一种解释。
为了检测纳米级别的物体是很小的细菌或它们的化石对等物的假说, Kirkland等(1999)进行了40多次方解石沉淀作用试验, 他们的研究结果表明了以下几点:首先, 很多纳米级别的特征物是无机成因的, 或者是非超微细菌成因的; 其次, 在沉淀作用试验中, 产生了50~80 nm大小的自形方解石晶体, 以及更小的(< 50 nm)异型方解石晶体、圆形物或原始晶体, 这些较小的异型方解石晶体类似于以前所解释的“ 超微细菌” ; 第三, 在运用可溶解的有机组分进行沉淀作用试验的时候, 异型方解石晶体和圆形的方解石晶体的数量最多, 而且圆形的纳米级别的特征物在富有机质的环境中数量最多; 第四, 试验过程中产生的圆形或不规则形态的细菌碎片, 包括细胞壁的碎片、脱落了的细胞质和残余胞囊, 与以前所解释的“ 超微细菌” 极为相似; 第五, 对产生于沉淀作用试验中的大型的自形方解石晶体进行酸腐蚀, 也产生了类似于天然碳酸盐岩中的所谓的“ 超微细菌” 的特征物。所以说, 试验结果表明, 以前被假定为“ 超微细菌” 的纳米级别的特征物, 可能是细菌碎片, 这个解释还得到以下观察和研究的支持:
1)来自于犹他州大盐湖鲕粒中的纳米级别的特征物, 在靠近正在腐烂的鳃足虫卵的地方最为丰富, 远离之则变得稀少(Pedone and Folk, 1996)。
2)在二叠系Capitan组之中, 纳米级别的物体总是出现在微生物碳酸盐岩中, 在相同样品的淡水方解石亮晶中则很稀少(Kirkland et al., 1998)。
3)在试验中所观察到的类似于假定的超微细菌的特征物, 在形态上与现代环境中曾经观察到的细菌碎片或其他有机分子具有相似性, 这些有机分子包括类蛋白质或多糖类, 它们常常以较小的球形或卵圆形产出。
4)更为重要的是, 细菌将会通过产生富有机质的微环境来促进方解石的沉淀作用, 在其中, 方解石的可溶性产物实际上是过剩的(Simkiss and Wilbur, 1989); 尽管这些纳米级别的特征物, 是否是真正的超微细菌、或者是否是简单的原始晶体还不太清楚, 但是, 在细菌造成的方解石沉淀作用中, 所形成的类似于“ 超微细菌” 的纳米级别特征物是很普遍的(Chafetz and Buczynski, 1992)。
关于是否存在“ 超微细菌” , 争论还没有结束(Rasmussen et al., 2002), 主要表现在以下两个方面:首先, 培养基的试验结果还是被引证为“ 超微细菌” 存在的证据(生物学范畴中首选的说法; Kajander and Ciftcioglu, 1998); 其次, 所推测的与“ 超微细菌” 有关联的羟基磷灰石矿化作用(Kajander and Ciftcioglu, 1998), 似乎应该归因于自动传播的微晶中心的成核作用活动, 其成核作用、生长以及形态最有可能受到蛋白质调节, 从而还进一步解释为“ 超微细菌” 产生的生物矿化作用(Cisar et al., 2000), 间接地支持了“ 超微细菌” 的存在。
针对上述状况, 考虑到Kirkland等(1999)曾经将那些纳米级别的特征物解释为细菌碎片, 也考虑到生物学家认为200~300 nm是微生物可生存的大小下限, Schieber和Arnott(2003)在对沉积物中铁硫化物的微生物化石化作用进行相关实验时(Schieber, 2002), 对与“ 超微细菌” 相似的纳米级别的特征物进行了研究。实验结果(图 1)表明:在与微生物腐烂相关联的成岩矿化作用中, 产生了丰富的纳米级别的球状物, 与以前所解释的“ 超微细菌” (Folk, 1993, 1999; Pedone and Folk, 1996; Folk and Lynch, 1997)具有惊人的相似性。
 | 图1 被解释为“ 超微细菌” 的纳米级别的特征物的结构比较(转引自Schieber and Arnott, 2003)(左图引自网站(http://www.msstate.edu/dept/geosciences/4site/nannobacteria.htm), 该图像由Leo Lynch所提供, 而且被Folk和Lynch(1997)描述为牙买加的具有丰富超微细菌结构的微生物岩; 右图为正在腐烂的鱿鱼肌肉中的粒状纳米球结构(纳米球的大小 为50~120 nm; 引自Schieber and Arnott, 2003), 这些纳米球在形态上与Folk所称的超微细菌极为相似)Fig.1 Images showing comparison of nannobacterial textures(quoted from Schieber and Arnott, 2003) |
Schieber和Arnott(2003)进一步论述到, 运用鱿鱼肌肉所进行的生物软体组织的逐渐腐烂试验, 总是会产生纳米球, 他们推测这是微生物酶与埋藏的样本相互作用的结果。酶攻击有机分子的特殊腱, 例如, 胰蛋白酶会劈开连接赖氨酸和精氨酸残余物的肽腱, 而胰凝乳蛋白酶裂开连接苯基丙氨酸、色氨酸和络氨酸的肽腱(Lodish et al., 1995); 当酶劈开肌肉纤维或细胞壁聚合物结构的时候, 似乎就会产生越来越小的碎片。实验表明, 当蛋白质有机组织被暴露在蛋白质降解酶之中, 或者通过细菌活动或者浸入纯净的酶溶液之中, 纳米球就是随后的降解作用过程的副产品:在开始腐烂试验之后的两天内, 就可以观察到纳米球, 而且越来越丰富; 初始的酶降解作用, 很明显把较大的组织单元(如细胞壁、肌肉纤维)分解成若干具有数百纳米大小的亚单元。进一步观察表明, 酶的独特形态决定了它们的功能、以及什么类型的腱被切断; 在一个给定的情况下, 亚单元狭窄的大小范围可能与初始阶段受到影响的腱的重复距离存在联系。所以说, 酶所驱动的生物组织降解作用是所观察到的纳米球的主要形成原因。
在适合于生物组织矿化作用存在的情况下, 纳米球所具有的矿化组织的普遍性, 以及关于化石中软体组织的保存作用的研究(Briggs and Kear, 1994; Hof and Briggs, 1997; Wilby and Briggs, 1997; Gabbott, 1998)已经表明了这一点, 例如, 研究者已经注意到, 磷酸盐化的软体组织经常表现为微球粒结构。这种微球粒结构表明了一个由致密填集的黏土矿物或磷灰石微球粒组成的底层微组构, 其大小范围从30 nm(Wilby and Briggs, 1997)到150 nm(Gabbott, 1998)。
多数微生物学家认为, 超微细菌是活着的生命实体(Nealson, 1997; Cisar et al., 2000), 所以说, 还不能完全抛弃将它们作为有活力的生命形式的可能性(Kajander and Ciftcioglu, 1998; Uwins et al., 1998; Madigan et al., 2002), 因为已经由化学分析和DNA检测所证明。然而, 甚至在接受这个主流的微生物学观点的情况下, 对于岩石记录中所描述的“ 超微细菌” 的成因(Folk, 1993; McKay et al., 1996; Pedone and Folk, 1996; Folk and Lynch, 1997), Schieber和Arnott(2003)的试验观察和研究提供的另一个可能的解释是:它们是生物有机组织细菌腐烂的矿化作用副产品。
本质上讲, 在足够高的放大倍数下进行检测, 与成岩早期矿物沉淀作用相关联的有机质腐烂的任何沉积物, 都应该显示出矿化纳米球的存在。其实, 这正是Folk和他的合作者, 在对土壤、沉积物、矿物和岩石的高倍放大研究时所发现的情况(Folk, 1993, 1999; Pedone and Folk, 1996; Folk and Lynch, 1997, 2001)。
如果超微细菌的支持者(如Kajander and Ciftcioglu, 1998)能够克服现有的疑问(Cisar et al., 2000), 并且肯定这种生命形式确实存在的话, 纳米球似乎需要一个特殊的而且营养物丰富的环境(如人和奶牛的血浆浆液), 在更加稀释的自然环境中它不应该如此丰富。
医疗研究者报道过与矿化纳米球相关的疾病, 如肾结石、动脉阻塞和白内障(Kajander and Cifticioglu, 1998; Rasmussen et al., 2002)。但是, 矿化纳米球解释的批评者曾经建议, 这些归因于超微细菌的生物矿化作用所形成的矿化纳米球, 可能是由非活着的生物分泌的有机高分子所产生(Cisar et al., 2000)。
与微生物腐烂相关联的成岩矿化作用的化石记录, 应该表现出丰富的纳米球的保存。如果在大多数沉积岩中, 与“ 超微细菌” 相似的结构可能是有机物质的细菌降解作用的副产品的话, 就不会存在被称作超微细菌的微小生命形式。Schieber和Arnott(2003)进一步指出, 矿化纳米球可能表明了有机组织上的细菌酶作用, 而且可以作为一个栩栩如生的微生物活动的典型代表(Schieber and Arnott, 2003)。
被有争议地描述为“ 超微细菌” (Folk, 1993, 1999; Vasconcelos and McKenzie, 1997; Dupraz et al., 2004)的纳米级别的特征物, 确实客观存在于碳酸盐岩和碳酸盐沉积物之中, 从而激发了沉积学家对碳酸盐沉淀作用机制的研究和思考, 尤其是微生物活动与灰岩的研究。
尽管鲕粒曾经被认为是真正的无机碳酸盐沉淀物的典型代表(Davies et al., 1978; Sandberg, 1983), 但是, 一个多世纪以来(从1892年Rothpletz的早期描述和研究, 一直到Reitner等(1997)和Neuweiler等(1999)的研究), 地质学家已经认识到, 有机过程及其产物与微生物本身在鲕粒的形成中可能起着很重要的作用, 证据包括鲕粒内部的藻类和细菌、鲕粒内部或鲕粒表面的微生物钻孔作用和黏液层、各种有机化合物(谷氨酸和谷氨酸盐等), 以及这些有机物在钙碳酸盐沉淀作用和成形作用之间的分子水平紧密关系的细节的观察和研究(详见Folk和Lynch(2001)的回顾)。
对巴哈马台地的Joulters Cay现代鲕粒的观察和研究(Folk and Lynch, 2001), 发现了以下明显的现象:(1)鲕粒内部富集有机黏液, 有机质呈分散聚集的环状物产生在鲕粒的皮层之中; (2)鲕粒皮层以及沉积作用的旋回性表现很明显, 当鲕粒被切穿到核心而且在岩石显微镜下观察时, 很明显鲕粒皮层是由旋回性的重复作用所形成:结晶的双折射的(文石)环状物, 为2~4μ m厚, 随后是一个大约1μ m的均质间隔; (3)鲕粒皮层是一个有机质黏液和文石的复合体, 即鲕粒皮层主要由文石的针状晶体所构成, 较薄的黏液环, 对鲕粒来讲呈同心状, 经常融入小型的文石球, 但是常常缺乏文石针, 而且较厚的文石环也含有一些黏液膜; (4)鲕粒外圈的微生物钻孔主要是空洞, 又是具有一个烘烤后变黑的边缘, 然而内部的微生物钻孔则由碳酸盐沉淀物所充填, 说明鲕粒生长与微生物活动存在一些联系; (5)末端浑圆的文石棒状物, 富集在鲕粒(皮层)的富有机质部分, 因为在很多种碳酸盐岩之中, 这些末端球与假定的“ 超微细菌” 的形态和大小相同(Folk, 1993), 就可以认为文石棒状物的末端球可能是超微细菌, 而且这种超微细菌促进了文石的沉淀作用, 再者, 假定的“ 超微细菌” 已经在富有机质的现代(Pedone and Folk, 1996)和古代碳酸盐岩(Gournay et al., 1999; Kirkland et al., 1999)之中被发现。
针对上述现象, 考虑到超微细菌是否能够作为一种微生物生命形式, Folk和Lynch(2001)对鲕粒的形成过程进行了以下设想和解释。
首先, 如果认为超微细菌存在的假说是正确的, 一个精美的、与其相关的鲕粒沉淀作用模式就可能存在。也就是说, 鲕粒皮层内的文石形成在那些覆盖在鲕粒上的、而且不混溶的有机质黏液内, 具有末端球的文石棒状物、以及无轴的超微颗粒则形成在鲕粒皮层内的黏液席中, 这可能是由超微细菌所产生的文石沉淀作用的实例。
其次, 如果假定的超微细菌作为一种微生物生命形式不存在的话, 类似的现象也可以得到解释(Folk and Lynch, 2001)。人们很早就知道, 水化学性质影响了碳酸盐多型晶状态以及沉淀作用能力, 在较大的级别上, 则影响着晶体形态。最近的研究已经表明, 纳米级别的、与传说中的超微细菌相似的他形方解石原始晶体, 在具有高度溶解的有机质的溶液之中发生沉淀作用(Kirkland et al., 1999)。Folk和Lynch(2001)还进一步论述到, 在鲕粒中以及在很多碳酸盐岩中, 衰变的细菌片段也与传说中的超微细菌极为相似(Kirkland et al., 1999), 如果这些颗粒保留一些潜在电荷, 或者以一些其他方式造成碳酸盐过饱和作用, 它们对鲕粒的形成还是很重要的。
所以说, 如果确实存在超微细菌这样一种生命形式, 那么巴哈马的鲕粒最有可能的就是直接的微生物成因; 但是, 换句话说, 纳米级别的圆形物可能是完全无机成因的, 初始种子晶体或者圆形物可能形成于文石晶体的“ 中毒式的” 生长。
最后, Folk和Lynch(2001)谦虚地指出:“ 在两种情况下, 亚球形的纳米级别的特征物肯定是了解鲕粒形成的关键。正确的问题被提出, 就像其他许多相应而生的问题一样, 答案只能少量施舍(In either case, the subspherical nanometre-scale features certainly are the key to understanding the ooids.The correct questions are being asked but, as with many consequential problems, the answers are doled out in small portions.)” 。他们这种令人尊敬的科学态度, 以及合理的推论和认识, 为碳酸盐沉积学的发展提供了宝贵的经验和进一步思考的途径。
在现代和古代地质环境中, 微生物广泛地卷入碳酸盐矿物的沉淀作用中(Riding, 2000; Krumbein et al., 2003), 其结果形成了各种各样的微生物碳酸盐岩(Riding, 2000; 梅冥相, 2007a, 2007b), 包括大量的泥晶或灰泥及其聚集体如鲕粒、球粒等, 这方面的观察和研究导致了对灰岩成因的进一步研究(Castanier et al., 1999)。微生物的卷入具有双重性:增强了水生环境的碱度, 而且形成许多新陈代谢的产物。微生物新陈代谢的产物, 已经得到了广泛的调查(Castanier et al., 1999)。知道得较少的是微生物如何影响碳酸盐的成核作用(Schultze-Lam et al., 1996), 因为地球化学模拟和实验室试验表明, 碳酸盐的成核作用过程控制了大部分地球历史中的微生物碳酸盐形成作用, 而不是直接的微生物新陈代谢作用。
在自然环境中, 微生物细胞和碳酸盐矿物之间紧密的空间组合, 被作为微生物参与成核作用的证据(van Lith et al., 2003a), 这个概念得到了实验室试验提供的间接证据的支持(Warthmann et al., 2000; Bosak and Newman, 2003)。微生物细胞以及细胞外聚合物质(EPS)总是带有负电荷而具有粘接二价钙离子的能力, 从而被经常引证为碳酸盐成核作用的场所(van Lith et al., 2003a; Dupraz et al., 2004)。但是, 缺乏成核作用过程的直接观察, 而且成核作用地点没有得到确定。如果钙碳酸盐成核作用发生在微生物细胞表面的话, 那它就会导致细胞变成坟墓、最终导致微生物的死亡(Morita, 1980; Southam and Donald, 1999)。因此, 碳酸盐成核作用的场所对于微生物细胞来讲是至关重要的; 再者, 成核作用过程可能控制了碳酸盐沉淀物的形态。在古代岩石以及在地外生命的探索中, 需要指示微生物卷入矿物形成形态的代表性标志(Allen et al., 2000), 对于微生物如何影响成核作用过程的理解, 是定义这个代表性标志的基本步骤。
针对上述研究现状, Aloisi等(2006)通过实验室试验, 调查了硫酸盐还原细菌(Desulfonatronum lacustre)的钙碳酸盐成核作用过程, 并进行了微生物碳酸盐沉淀物的地球化学研究和透射电子显微镜(TEM)观察, 从而提出了微生物碳酸盐成核作用在纳米级别上的一个新模式:大量的碳酸盐沉淀作用发生在形成于靠近微生物细胞壁的那些纳米微球(nanoglobule; 图2)上, 而且钙化明显只发生在被释放在微生物细胞周围含水的环境之中的这些微球上, 这些纳米级别的微球(简称为纳米微球(nanoglobule)), 也正是那些有争议地被描述为超微细菌的、纳米级别的钙化球状物(Folk, 1993, 1999; Pedone and Folk, 1996; Gournay et al., 1999; Kirkland et al., 1999; Schieber and Arnott, 2003; van Lith et al., 2003a; Dupraz et al., 2004)。
Aloisi等(2006)的试验表明:微球形成的初始步骤发生在一个与细菌细胞表面密切相关的EPS膜中, 几种生物物质可能组成这些微球的核心。实际上, 与D.lacustre微球相似的纳米级别的球状体, 已经在生物宏观大分子如磷脂和蛋白质上的羟基磷灰石的成核作用的实验室试验中生成过(Cisar et al., 2000; Vali et al., 2001; Benzerara et al., 2004); 换句话讲, 微球可能开始形成于EPS片段上、或形成于D.lacustre细胞壁的细胞膜胞囊上(Beveridge, 1999)。所有这些物质均产生在细胞表面, 而且具有使金属离子成核的能力, 从而可能作为微球核心的候选物。不管微球核心的属性如何, Aloisi等(2006)的试验描述的成核作用过程, 使一个存在争议的微生物钙化作用过程变得更加清楚。
在新陈代谢活跃的微生物细胞上的金属离子的粘结过程, 由于细胞的质子化作用而受到抑制(Urrutia et al., 1992)。在Aloisi等(2006)的试验中, 很局限的钙化作用发生在细胞表面以及靠近细胞表面的地方。尽管其他试验没有能够识别出碳酸盐成核作用的地点, 实验室的试验已经表明, 新陈代谢活跃的硫酸盐还原细菌(SRB)能够产生剧烈的碳酸盐成核作用(van Lith et al., 2003b); 然而, 在环境流体中, 微球上的钙碳酸盐沉淀作用, 远离细胞表面, 解决了这个明显的模糊性。沉淀作用发生在微球上而不是发生在细菌细胞壁上的事实, 将有益于D.lacustre所代表的硫酸盐还原细菌之类的微生物的新陈代谢活动, 因为它减少了产生细胞坟墓的机会。另外, 由于碳酸盐沉淀作用, 在SRB细胞周围含水的环境中, 还将造成钙饱和程度和碱度的降低, 对细菌生活产生了一个有利的化学条件(Hammes and Verstraete, 2002)。
但是, 还有待于证实的问题是, 这些纳米级别的微球的形成, 是否是一个涉及到细胞坟墓保存的生理机制、或者是否发生在不依赖于介质而相对于碳酸盐矿物饱和状态的条件响应。以前的研究表明, 纳米级别的碳酸盐球状物, 确实形成在微生物碳酸盐沉淀作用试验之中(Vasconcelos et al., 1995; Warthmann et al., 2000)。但是, 纳米微球的成因以及在碳酸盐成核作用中的卷入不能得到阐释。在Warthmann等(2000)所进行的实验之中, 第一阶段发育的是碳酸盐“ 哑铃” 体, 纳米级别的碳酸盐颗粒形成在SRB细胞的极性终端。附着在D.lacustre极性终端的大型微球的聚合物(图 2), 表明了Warthmann等(2000)试验中碳酸盐沉淀作用的初始阶段, 受到成核过程的控制。另外, 纳米级别的微球曾经在沉积环境中被广泛描述过(Folk, 1993, 1999; Vasconcelos and McKenzie, 1997; Dupraz et al., 2004), 而且有争议地被描述为超微细菌(Folk, 1993, 1999); 这个概念曾经经受过强烈的批评, 因为所描述的形式小于微生物生命的理论大小下限(200~300 nm)(Maniloff, 1996, 1997)。
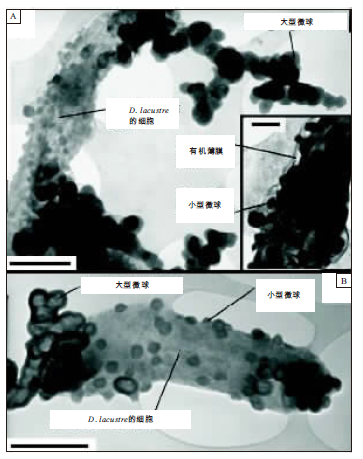 | 图2 运用硫酸盐还原细菌(Desulfonatronum lacustre)进行的沉淀试验中所形成的碳酸盐凝絮粒的透射电子显微镜(TEM)图像(引自Aloisi等, 2006)(A— 完全由小型微球所覆盖的D.lacustre细胞的细节, 这些小型的微球(small globules)嵌入在可能由细胞外聚合物质(EPS)组成的电子致密物质的薄层(< 20 nm厚)之中, EPS由D.lacustre所分泌; 大型微球(large globules)形成聚集物, 这些聚集物部分附着在D.lacustre细胞的极性边上(比例尺为500 nm); 嵌入的小图:小型微球附着在D.lacustre细胞上, 而且被EPS覆盖 (比例尺为20 nm); B— D.lacustre细胞的细节, 表明了大型微球的聚集物附着在细菌细胞的极性终端上(比例尺为500 nm))Fig.2 Transmission electron microscope(TEM)images of carbonate floccules formed in precipitation experiment with sulfate-reducing bacterium Desulfonatronum lacustre(quoted from Aloisi et al., 2006) |
如图2所示, Aloisi等(2006)的沉淀作用试验之中所产生的纳米级别的微球, 具有与沉积物中所描述的假定的超微细菌共同的化学和形态特征。由真正的细菌造成的微球, 还被解释为酶所驱动的生物组织腐烂过程的产物(Schieber and Arnott, 2003), 这个过程揭示了沉积环境中的那些像超微细菌的纳米级别的球状物体的成因。总之, 在天然环境中纳米级别的钙化球状物广泛存在, 而成核作用试验表明, 在微生物派生的纳米微球上的钙碳酸盐成核作用, 可能是微生物钙化作用的一个重要步骤。
Bontognali等(2008)的试验研究, 进一步支持了Aloisi等(2006)的沉淀作用试验的结果。在30℃下的缺氧环境之中, Bontognali等(2008)运用一种能够调节白云石沉淀作用的硫酸盐还原细菌, 即Desulfovibrio brasiliensis, 进行矿化作用试验, 得到的结果是:微生物所分泌的细胞外聚合物质(EPS)在矿化作用过程之中起到了关键作用, “ 超微细菌” 形状的颗粒代表了在EPS内部的碳酸盐成核作用的早期阶段, 而且会逐渐演变成具有颗粒结构的大型球状物。Desulfovibrio brasiliensis的细胞, 位于发生矿化作用的EPS聚合物的外表, 而且这些细胞是游动的, 从而避免了在矿化作用形成的矿物内部变成坟墓; 因此, 在EPS内的矿化作用还解释了为什么微生物残余物不一定都能够出现在生物成因的碳酸盐之中。
Pacton等(2010)的进一步研究也说明, 纳米级别的球状物可以作为沉积有机质中普遍的微生物产物, 并非是“ 超微细菌” , 这种球状物在以EPS和细菌为主的保存较好的有机质中产出最多; 它们呈束状产出, 与细菌的细胞壁和EPS紧密相关联, 而与藻类生物的细胞壁和其他的陆生有机碎片没有联系。所以说, 纳米级别的球状物是先成形成的有机质在早期成岩阶段的代表物, 表明了微生物对有机质保存作用的重要贡献。因此, 纳米级别的球状物揭示了微生物与其他有机碎片之间复杂的相互作用, 从而成为微生物在有机质早期成岩阶段的作用信号。
Bontognali等(2008)以及Pacton等(2010)的研究, 均强调了EPS的作用, 进一步否定了“ 超微细菌” 作为一种生命形式的存在。Garcí a-Ruiz等(2009)关于锶和钡碳酸盐、二氧化硅的纳米级别的晶体形态学的研究, 也对“ 超微细菌” 作为一种生命形式的存在持否定态度; 更为重要的是, Martel和Young(2008)对人体血液中纳米级别的球状物的研究, 也对其形成提出了非生物成因的解释而否定了“ 超微细菌” 作为一种生命形式的存在。这些研究, 更加肯定了Chafetz和Buczynski(1992)关于细菌调节的碳酸盐成核作用的研究, 也肯定了Castanier等(1999)关于灰岩成因的观点:“ 除了纯粹的蒸发盐和自养生物成因, 绝大多数灰岩可以被认为是异养细菌成因的, 而且它们所包含的碳主要来自于原始有机质” 。
就像戴永定(1994)在《生物矿物学》一书中, 对生物矿物体的结构、构造、类型、形态等进行的全面介绍和论述那样, 作为碳酸盐岩的主要组成部分, 除了显生宙以来伴随着生物骨骼化作用而产生的生物矿化作用形成的生物碎屑以外, 泥晶和灰泥本身及其聚集体— — 鲕粒、核形石和叠层石等, 更多是微生物复杂的新陈代谢过程之中的有机矿化作用的产物(Trichet and Defarge, 1995; Mann, 2001; Skinner and Jahren, 2003; Reitner, 2004; Perry et al., 2007)。虽然超微细菌作为一种生命形式, 随着研究的深入逐渐被否定, 但是, 围绕超微细菌的争论, 促进了灰岩成因研究的巨大进展。同时, Castanier等(1999)关于绝大多数灰岩是异养细菌成因的观点, 又意味着立足于宏观表象特征的微生物碳酸盐岩分类体系(纹层状的叠层石、紊乱状的凝块石、树叶状的树形石和贫乏组构的均一石, 以及球状的核形石和纹理状的纹理石; Riding, 2000; 梅冥相, 2007a, 2007b)还欠全面, 也就是“ 微生物碳酸盐岩” 的定义本身的科学依据也欠充分。
因此, 图1和图2所示的那些曾经被假定为“ 超微细菌” 的纳米微球, 有可能是酶所驱动的生物组织腐烂过程的产物(Schieber and Arnott, 2003), 而且可能是碳酸盐成核作用的地点(Aloisi et al., 2006), 从而代表了复杂的微生物形成的有机矿化作用过程。这些复杂的微生物形成的有机矿化作用过程, 常常发生在微生物席内, 尤其是微生物席内部的EPS上(Decho et al., 2005; Bontognali et al., 2008; Pacton et al., 2010), 为研究叠层石、鲕粒乃至灰泥的形成机制这些谜一样的碳酸盐沉积学问题, 提供了重要的思考途径和坚实的研究基础(Brehm et al., 2006; Perry et al., 2007; Dupraz et al., 2009; Daniel and Chin, 2010)。所以说, “ 超微细菌” 的争论, 是促进碳酸盐沉积学发展的动力源泉。如果说, 在1994年出版的里程碑式的宏篇巨著— — 《中国沉积学》中, 冯增昭、刘宝珺、王英华、沙庆安、钱凯、赵震等学者对碳酸盐沉积学当时的研究进展和现状进行了精辟的总结和论述的话, 大致开始于1992年的“ 超微细菌” 的提出以及后来的持续性争论与研究, 从另一个侧面反映了碳酸盐沉积学近20年来的飞跃发展。
超微细菌及其化石对等物— — 超微化石, 从1992年被Folk提出以来, 就引起了生物学家和沉积学家的巨大关注和持续性研究, 产生了激烈的争论, 而且争论还在继续着。与其相关的持续性研究, 如将这些曾经被解释为超微细菌的纳米级别的球状物, 解释为细菌碎片、生物组织腐烂过程的副产品, 乃至解释为在EPS内部的复杂的有机矿化作用的产物, 导致了许多新概念的诞生和老概念的修订, 尤其是更加肯定了灰泥或泥晶及其聚集体(鲕粒、核形石、叠层石等)的微生物成因, 从而大大地促进了碳酸盐沉积学的发展。这些认识, 对碳酸盐沉积学中的那些谜一样的问题, 如叠层石、鲕粒、灰泥(或泥晶)的成因机制等, 提供了重要的思考途径并为进一步研究奠定了坚实的基础, 所以说, 发生在世纪之交的这场争论成为碳酸盐沉积学发展的动力源泉。因为在运用实验室的试验结果去解释复杂的岩石记录的时候, 要穿越成岩作用“ 过滤器” , 成岩作用“ 过滤器” 常常会造成许多原生特征的丢失; 更为重要的是, 复杂的自然条件还难以运用实验室的试验来进行全面的模拟。正是这些原因, 根据宏观形态特征提出的微生物碳酸盐岩的概念及其相关的分类体系, 将会面临着巨大的概念转变和重新修订。以简单的化学和矿物组分为特征的碳酸盐岩, 代表着地球表层重要的钙化作用和固碳作用, 影响了地球大气圈和水圈构成的演变, 而且由地表环境中不稳定的矿物所组成, 由其所蕴含的许多科学命题将有待于进一步研究和阐释。
致谢 北京大学刘建波副教授和中国科学院地质与地球物理研究所吴亚生研究员精心审阅了论文, 并提出了诸多极为宝贵的修改意见; 成文过程中得到了《古地理学报》主编冯增昭教授的悉心指导。作者对上述学者的无私帮助致以衷心感谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|