








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
华南泥盆纪遗迹化石及遗迹相*
[张立军1, 2  , 龚一鸣
, 龚一鸣1 , 马会珍3 ]
, 龚一鸣, 马会珍|
|

通过对华南泥盆系 6条剖面(四川北川甘溪、广西横县六景、广西桂林杨堤、贵州独山大河口—白虎坡、贵州贵阳乌当、四川广元后高坪)的遗迹沉积学系统研究,鉴定和描述遗迹化石 15属 24种,包括Arenicolites carbonaria,Arenicolites isp.,Chondrites cf.intricatus,Chondrites fenxiangensis,Chondrites filifalx, Chondrites isp., Chondrites maqianensis,Circulichnis isp., Dushanichnus dahekouensis, Diplocraterion parallelum, Rusophycus lungmenshanensis,?Helminthopsis isp., Palaeophycus tubularis,Palaeophycus curvatus,Phycodes palmatus, Planolites beverleyensis, Planolites isp., Planolites kwangsiensis, Rosselia socialis, Rhizocorallium jenense, Rhizocorallium isp.,Skolithos linearis, Thalassinoides isp.,Zoophycos isp.。根据遗迹化石之间的共生组合关系、实体化石特征和其他相标志,识别出 4种遗迹相: Skolithos,Rhizocorallium,Cruziana 和Zoophycos遗迹相。Skolithos遗迹相主要发育于浪控型海岸体系、潮控型海岸体系和障壁岛—潟湖沉积体系的高能沉积区;Rhizocorallium遗迹相主要发育于潮控型海岸体系和障壁岛—潟湖沉积体系的低能沉积区;Cruziana遗迹相发育在浅海陆棚、具有丰富的生物及食物的砂泥岩和灰岩为主的低能沉积区;Zoophycos遗迹相沿泥岩和泥灰岩发育的滨外沉积区分布。华南泥盆纪遗迹相从早泥盆世到晚泥盆世的演替规律是从Skolithos遗迹相→Rhizocorallium遗迹相→Cruziana遗迹相→Zoophycos遗迹相→Rhizocorallium遗迹相。早泥盆世晚期以后华南泥盆纪同时期的遗迹相自西南向东北的空间变化规律则表现为Zoophycos遗迹相→Cruziana遗迹相→Rhizocorallium遗迹相→Skolithos遗迹相。
About the first author:Zhang Lijun,born in 1982,is a Ph.D. candidate of China University of Geosciences(Wuhan).He majors in paleontology and stratigraphy and is mainly engaged in ichnology and palaeoenvironment.E-mail: ljzhanghpu@gmail.com.
About the corresponding author:Gong Yiming,born in 1958,professor,now is engaged in ichnology,Devonian and geobiology.E-mail: ymgong@cug.edu.cn.
第一作者简介:张立军,男, 1982年生,中国地质大学(武汉)博士研究生,专业为古生物与地层学,研究方向为遗迹学与古环境。E-mail: ljzhanghpu@gmail.com。
通讯作者简介:龚一鸣,男, 1958年生,教授,主要从事遗迹化石、泥盆系和地球生物学研究。 E-mail: ymgong@cug.edu.cn。
Fifteeen ichnogenera and twenty-four ichnospecies were recognized on the basis of detailed ichnosedimentology studies of six Devonian sections from South China.They included Arenicolites carbonaria, Arenicolites isp., Chondries cf. intricatus, Chondrites fenxiangensis, Chondrites filifalx, Chondrites isp., Chondrites maqianensis, Circulichnis isp., Dushanichnus dahekouensis, Diplocraterion parallelum, Rusophycus lungmenshanensis,? Helminthopsis isp., Palaeophycus tubularis,Palaeophycus curvatus,Phycodes palmatus, Planolites beverleyensis, Planolites isp., Planolites kwangsiensis, Rosselia socialis, Rhizocorallium jenense, Rhizocorallium isp., Skolithos linearis, Thalassinoides isp.,Zoophycus isp.. These trace fossils can be grouped into four ichnofacies of Skolithos, Rhizocorallium, Cruziana, Zoophycos ichnofacies in terms of body fossils and other facies indicators. Skolithos, Rhizocorallium, Cruziana and Zoophycos ichnofacies indicated tidal flat and barrier island-lagoon environments,intertidal-subtidal zones,upper-middle neritic shelf and lower neritic-bathyal environments,respectively.From the Early to Late Devonian,the replacement of ichnofacies was from Skolithos ichnofacies→ Rhizocorallium ichnofacies → Cruziana ichnofacies → Zoophycos ichnofacies → Rhizocorallium ichnofacies.
遗迹化石是古生物学与沉积学研究的重要信息载体, 对重建古环境、古生态和古地理, 诠释生物与环境之间的相互作用具有重要意义。正如岩相一样, 遗迹相也遵从瓦尔特相律, 在一定条件下, 遗迹相在纵向上的叠置特征能反映其在横向(空间)上的相变规律。像其他相分析一样, 对遗迹相的深入研究将大大提高寄主地层的古环境和古生态研究水平及解译精度。
遗迹相包括了在时间与空间上重现的遗迹化石生态组合, 反映了特定条件下生物对环境的响应。遗迹相最先由Seilacher在20世纪50、60年代提出并加以发展(Seilacher, 1953a, 1953b, 1967), Seilacher(1967)定义了初始的6种遗迹相, 并以特征的遗迹属命名。这6种遗迹相包括软底质的4种遗迹相(Skolithos, Cruziana, Zoophycos, Nereites), 1种底质控制的类型(Glossifungites)和1种单一的陆相软底质类型(Scoyenia)。Frey 和Seilacher(1980)增加了在硬底质上具有钻孔特征的Trypanites遗迹相, Bromley等(1984)增加了在木质底质上环绕的钻孔Teredolites遗迹相, Frey和Pemberton(1987)提出了海陆过渡环境或海岸带受海、陆双重条件控制的以永久性垂直到倾斜居住潜穴为主的Psilonichnus 遗迹相。上述9种遗迹相已经被证明在时间和空间尺度上都可以重复出现。近来, 又提出了Gnathichnus 和Entobia两种新的硬底质遗迹相(Bromley and Asgaard, 1993)。此外陆相地层遗迹相也有了新的扩展, 增加了古土壤的Termitichnus遗迹相(Smith et al., 1993)以及湖泊环境的Mermia遗迹相(Buatois and Mangano, 1995), 在此之后, Termitichnus遗迹相被丢弃, 取而代之的是Corprinisphaera遗迹相。控制遗迹相分布的因素最先被认为是水深(Seilacher, 1967), 随着研究的深入, 发现众多因素都会直接或间接地影响遗迹相的分布, 主要包括:底质的固结程度、底层水动力条件、物源、水体的混浊程度、盐度、沉积速率、氧化还原条件和温度等(MacEachern et al., 2007)。此外越来越多的研究表明, 一些遗迹相的组成及分布范围是随着地质时期的演变而变化的, 例如, 寒武纪浅海相的Cruziana遗迹相包括了一些Nereites遗迹相的遗迹属种(Crimes, 1987); 随着时间的推移, Zoophycos遗迹相的分布范围由滨浅海向深海逐渐过渡(Bottjer et al., 1987; Bromley, 1990)。
泥盆系在中国南方广泛出露, 相关遗迹化石的研究也取得了一定的成果, 主要集中在对单一遗迹属种的描述以及应用遗迹组构的功能来进行古环境分析(龚一鸣, 1985, 1987, 2004; 杨式溥等, 1987; 周志澄, 1996; 王约等, 1997)方面, 有37个遗迹属被先后报道, 包括Ancorichnus, Arenicolites, Arsterichnus, Baihuoichnus, Bifungites, Bimammiplichnus, Chondrites, Circulichnis, Cosmorphaphe, Cochilichnus, Cylindricum, Cruziana, Dimorphichnus, Daedalus, Dushanichnus, Diplocraterion, Diplichnites, Gordia, Helminthopsis, Lumbricaria, Megagrapton, Monocraterion, Ophiomorpha, Palaeophycus, Pelecypodichnus, Phycodes, Planolites, Rosselia, Rhizocorallium, Rusophycus, Scalaritaba, Scoyenia, Skolithos, Teichichnus, Thalassinoides, Treptichnus 和 Zoophycos(杨式溥, 1983, 1989; 何远壁等, 1985; 林文球等, 1986; 张欣平等, 1986; 杨式溥等, 1987; 王大任等, 1988; 张欣平和卿上康, 1988; 王约, 1992, 2003, 2004; 周志澄, 1996; 王约和龚一鸣, 1997; 王约等, 1998, 2006; 齐永安等, 2005; 张立军和龚一鸣, 2009)。此外部分学者对于泥盆系弗拉阶— 法门阶(F-F)灭绝事件前后遗迹群落形态和功能的变化作了相关的报道(王约, 2004; 王约等, 2006), 但是尚未有基于整个华南地区泥盆纪遗迹相变化及分布的相关研究工作。文中利用华南泥盆系6条剖面遗迹化石的详细材料, 结合前人大量的相关遗迹化石研究成果, 将遗迹化石的时间演替与空间分布相结合来探讨华南泥盆系遗迹相的组成、结构和分布及其与古环境之间的关系。
华南板块在泥盆纪位于古特提斯洋东侧, 靠近澳大利亚板块, 大约在赤道附近, 气候炎热。志留纪末的加里东运动使得海水从华南地区撤去, 仅在钦防地区留有一隅。加里东运动之后, 华南板块大部分隆起为陆, 从东到西分别是康滇、扬子、武陵、江南、武夷、云开、华夏等隆起, 构成向南伸出的钳形陆地, 包围南部的钦防海槽(陈代钊和陈其英, 1994)。泥盆纪初, 随古特提斯洋的向东扩张, 海水自南向北侵入, 海域从南往北逐步推进。早泥盆世早期以滨岸碎屑岩沉积为主, 晚期碎屑岩沉积和碳酸盐沉积交替出现。中、晚泥盆世以碳酸盐沉积为主, 反映了海侵逐渐扩大的过程; 其中在中泥盆世早期有一次明显的海退。晚泥盆世早期海侵范围最大, 晚泥盆世晚期又开始海退。自早泥盆世埃姆斯期开始至晚泥盆世, 在陆地内部出现了NW向拉张和NE向拉张走滑盆地, 海盆中出现浅水碳酸盐岩台地(象州型)和条带状较深水硅、泥和泥灰质海槽(南丹型)的岩相及生物相分异。在川西北龙门山地区, 泥盆系仅分布在受断裂带控制的地区, 早泥盆世以滨浅海相的碎屑岩沉积为主, 早泥盆世晚期至中泥盆世晚期以碎屑岩和碳酸盐岩混积为特征, 晚泥盆世主要以碳酸盐沉积为主。
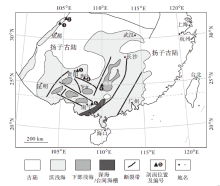
实测的6条剖面主要分布在桂中北(广西横县六景剖面、广西桂林杨堤剖面)、黔东南(贵州独山大河口— 白虎坡剖面、贵州贵阳乌当剖面)、川西北(四川北川甘溪剖面、四川广元上寺后高坪剖面)地区(图 1), 具有较强的代表性。上述剖面地层划分对比及遗迹化石分布见图2及图3。
 | 图1 华南晚泥盆世古地理和古构造(据赵锡文, 1985; 吴诒等, 1997; 有修改)( 剖面名称:❶— 四川北川甘溪(甘溪剖面), ❷— 四川广元上寺后高坪(后高坪剖面), ❸— 贵州贵阳乌当(乌当剖面), ❹— 贵州独山大河口— 白虎坡(独山剖面), ❺— 广西 桂林杨堤(杨堤剖面), ❻— 广西横县六景(六景剖面))Fig.1 Sketch showing the Late Devonian palaeogeography and tectonics of South China(modified from Zhao, 1985; Wu et al., 1997) |
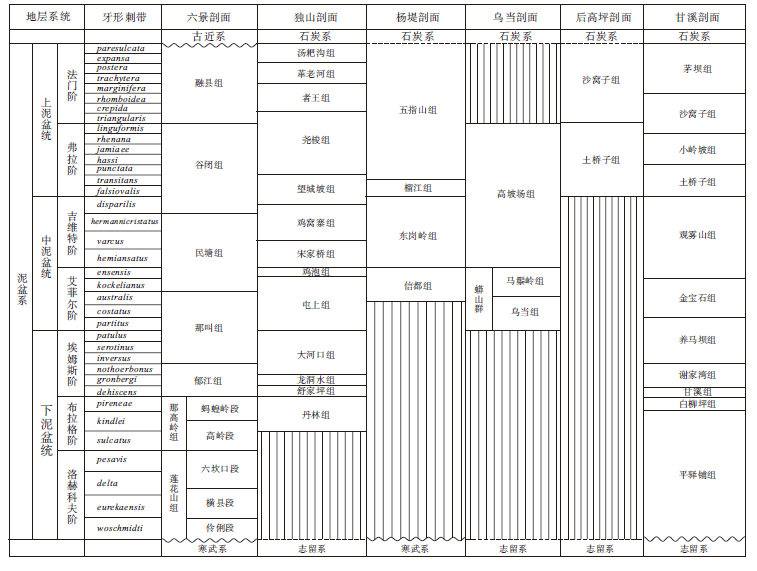 | 图2 华南泥盆系实测剖面地层划分对比Fig.2 Stratigraphic subdivision and correlation of the studied Devonian sections in South China |
甘溪剖面位于四川省北川县桂溪乡甘溪村— 沙窝子一带, 即北纬31° 54.302', 东经104° 41.643'。 剖面地处成都西北九寨沟旅游环线东段200km, 东距绵阳市江油市约35 km(图 1)。剖面起点始于北川县桂溪乡汽车站附近, 终点止于沙窝子石灰窑。泥盆系沿涪江支流平通河两岸分布, 全长约13km, 构造简单, 呈单斜连续出露。其底界与志留系呈角度不整合接触, 顶界与石炭系呈整合接触关系。遗迹化石主要分布在下泥盆统平驿铺组— 甘溪组的碎屑岩地层中, 中、上泥盆统碳酸盐岩地层中遗迹化石分布零星。在遗迹化石发育层段识别出4类遗迹化石组合和遗迹相(图 3-A)。
Skolithos遗迹相
平驿铺组下部第1~6和第8~13层砂质粉砂岩中, 共发育遗迹化石8属11种, 分别是Arenicolites isp.(图 4-C), Chondrites filifalx, Chondrites antiquus, Cylindrichnus concentricus, Diplocraterion parallelum(图 4-C), Palaeophycus tubularis(图 4-G), Palaeophycus alternatus, Planolites kwangsiensis, Rhizocorallium kueichouensis, Rhizocorallium jenense, Skolithos verticalis。Palaeophycus, Chondrites, Planolites和Skolithos是主要的优势属,
其中Skolithos丰度(统计属种数量的百分比)为28.7%, Palaeophycus丰度为37.4%, Chondrites丰度为16.5%, Planolites 丰度为10.2%, 这些属种占了整个平驿铺组下部遗迹化石统计样品的92.8%。就遗迹属种习性类别而言, 居住迹和觅食迹占据了整个遗迹群体的大部分, 如Skolithos、Palaeophycus、Diplocraterion, 这些属种占据了整个统计样品的67%, 其中觅食迹占33.6%。
平驿铺组第9层灰色中— 薄层石英砂岩中发现的Skolithos verticalis, 垂直层面分布, 横切面呈圆形, 直径为6~8mm左右, 保存内生迹, 潜穴内部无构造为被动充填, 常被认为产在近岸高能的潮间带和近海滨不稳定的砂质底质沉积物中(Seilacher, 1967; 杨式溥等, 2004)。在第8~13层的浅灰色及深灰色中薄层石英砂岩中, 与Skolithos相伴生的遗迹化石主要有Planolites和Palaeophycus。Planolites主要表现为柱形潜穴, 一般略弯曲, 潜穴不分枝, 表面光滑, 直径为3~5mm, 充填物质颜色不同于母岩, 保存为内生迹、表生迹凹槽及底生迹凸起, 常被认为是食沉积物动物的觅食构造, 是一种广泛分布于各种深度沉积环境的分子(Fillion and Pickerill, 1984; 杨式溥等, 2004); Palaeophycus表现为圆柱形潜穴, 略弯曲, 表面光滑, 偶有不规则的分枝, 充填物质致密, 成分同母岩, 保存为内生迹、底生迹凸起或表生迹凹槽, 与层面平行分布, 直径为4~6mm, 长度不等, 最长可达10cm, 常见于浅海到深海环境, 一般认为是由捕食性生物或滤食性动物形成的开放居住潜穴, 是由重力作用形成的被动充填(Pemberton and Frey, 1982; 杨式溥等, 2004)。
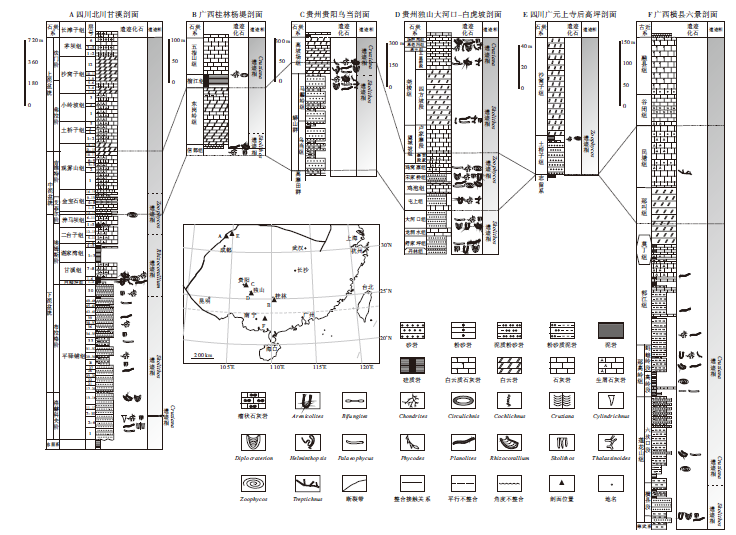 | 图3华南泥盆系实测剖面遗迹化石及遗迹相分布和对比Fig.3 Distribution and correlation of both trace fossils and ichnofacies from the studied Devonian sections in South China |
 | 图4 四川北川甘溪泥盆系剖面遗迹化石Fig.4 The Devonian trace fossils from Ganxi section, Sichuan of South China |
A— Rhizocorallium jenense, 保存在泥质粉砂岩中, 平行于上层面分布, 两翼管近平行, 末端较弯曲, 两翼管之间为同心状蹼状构造; B— Diplocraterion parallelum, 保存在砂岩中, 垂直层面排列, 仅保存U形潜穴的1个栖管, 栖管光滑无饰, 圆柱形末端呈圆桶状延伸到层面之上, 在层面上常见成对的圆形管口; C— 复式遗迹组合Diplocraterion parallelum和Arenicolites isp., 保存在砂岩中, 垂直层面分布, Diplocraterion parallelum为具蹼状构造的U形潜穴, U形潜穴的栖管较粗糙, 蹼状构造一般为前进式, 潜穴底部为半圆形, Arenicolites isp.为简单的无蹼层/板的U形潜穴, U形管垂直于层面, 管壁较粗糙; D— Rusophycus lungmenshanensis, 保存在砂岩中, 平行底层面分布。小型二叶形停息迹, 整体呈椭圆形, 前部宽度为6cm, 后部宽度为4cm, 长度约12cm, 高出层面约3mm, 二叶上具有前后两个方向的斜脊, 斜脊方向有变化, 前端斜向前方, 后部斜向侧方, 在中部偏后的部位形成无斜脊的三角形空白区, 中沟较清楚; E— Skolithos linearis, 与砂岩层面垂直的针形潜穴, 潜穴外表光滑, 颜色较围岩深, 潜穴内部无构造为被动充填, 横切面呈圆形; F— Zoophycos isp., 化石保存在砂岩中, 沿底层面呈全浮雕产出, 与层面平行的椭圆形蹼层。大蹼纹粗大, 小蹼纹不太明显, 部分小蹼纹充填物与围岩不同; G— Palaeophycus tubularis, 保存在泥质粉砂岩中, 平行上层面, 弯曲成弧形柱形潜穴, 半凸起于灰色砂岩层面上, 不分枝, 充填物同围岩; H— Chondrites fenxiangensis, 保存在泥质粉砂岩中, 分枝潜穴系统, 沿层面分布, 末端呈树枝状, 分枝粗细 基本一致, 至少见3次分枝。A为下泥盆统甘溪组, B至E和G至H为下泥盆统平驿铺组, F为中泥盆统养马坝组
与上述遗迹组构相伴生的主要沉积构造有楔状交错层理、水平层理及沙纹层理等。根据上述遗迹化石的古环境指相意义及相关的沉积构造, 将平驿铺组下段第1~6层和第8~13层划分为Skolithos遗迹相(图 3-A), 反映了滨海潮上带— 潮间带水动能较强的沉积环境。
平驿铺组中部第25~29层浅灰色中薄层石英砂岩及粉砂岩地层中, 共发育遗迹化石3属4种, 分别是Palaeophycus tubularis, Chondrites fenxiangensis(图 4-H), Skolithos verticalis, Skolithos linearis(图 4-E)。 Skolithos占据了整个平驿铺组中部遗迹群落丰度(属种统计数量的百分比)的58.7%, Palaeophycus占据了21.3%。从习性上看居住迹和觅食迹占据了统计数量的80%。从遗迹化石的种类及数量来看, 该段都较平驿铺组下部(第8~13层)少, 主要遗迹属潜穴直径的平均值都较下部略小, 如中部Skolithos潜穴的直径平均为3~4mm, 而下部平均值为8mm; Palaeophycus潜穴直径中部平均为3~4mm, 而下部平均为6 mm。主要遗迹属Skolithos, Palaeophycus表面均较平驿铺组下部光滑, 归入Skolithos遗迹相, 说明当时沉积环境逐渐稳定, 水动能减弱, 从侧面反映了由平驿铺组底部到中部海水逐渐加深的过程。由于本段主要的遗迹化石组合与平驿铺组下部一致, 因此不再进行遗迹属种的描述及分析。
平驿铺组顶部第48~50层中薄层泥质粉砂岩中, 发育Diplocraterion parallelum(图 4-B)和Chondrites filifalx, 下部石英砂岩及粉砂岩中常见水平层理及沙纹层理。Skolithos是该段地层中主要的遗迹属, 占据了96.2%。Diplocraterion的直径为1~2mm, 较中部小1~2mm, 反映了海水进一步加深、水动力条件进一步减弱的沉积环境, 将其归为Skolithos 遗迹相(图 3-A), 反映了滨海潮下带水动能较弱的沉积环境。
Cruziana遗迹相
平驿铺组下部第7层粉砂岩地层中, 共发育遗迹化石3属3种, 分别是Rusophycus lungmenshanensis (图 4-D), Chondrites cf. intricatus和 Planolites isp.。在第7层的深灰色炭质粉砂岩中发现由三叶虫停息形成的Rusophycus(图 4-D), 其保存为底生迹凸起, 外形呈椭圆形, 前部宽度为6mm, 后部宽度为4mm, 长度12cm, 高出层面约3mm, 二叶上具有前后两个方向的斜脊, 前端斜向前方, 后部斜向后侧方, 在中部偏后的部位形成无斜脊的三角形空白区, 中沟明显, 常见于潮坪及三角洲等环境(杨式溥等, 2004)。将第7层归入Cruziana遗迹相, 反映了滨海潮下带水动能较弱的沉积环境。
Rhizocorallium 遗迹相
甘溪组底部第1层薄层泥质粉砂岩中, 共发育遗迹化石3属4种, 分别是Chondrites fenxiangensis, Zoophycos cirinnatus, Rhizocorallium jenense(图 4-A)和 Rhizocorallium irregulare。Rhizocorallium是主要遗迹属, 占整个遗迹化石统计样品的63.4%, 其保存为内生迹, 表现为具有清楚蹼状构造的U形管, 顶部呈半圆形, 管壁光滑, 管直径为3~4mm, 宽约8~9cm, 两翼管与层面从斜交30° ~45° 至近平行于层面, 一般被认为是由甲壳动物所形成的觅食潜穴(Fü rsich, 1974)。
与Rhizocorallium伴生的生物主要为石燕类。该组遗迹化石的分异度高, 且遗迹大都沿层面或低角度斜交层面展布, 以觅食潜穴为主, 将其划分为Rhizocorallium 遗迹相(图 3-A), 反映食物丰富、光照充足、盐度正常、中至低能的潮下带沉积环境。
Zoophycos 遗迹相
养马坝组第10层的灰黄色中薄层粉砂岩中发育丰富的Zoophycos isp.(图 4-F), 其沿底层面展布, 表现为圆弧形细长管道潜穴, 见浅色及深色细长的蹼板, 宽度约15cm, 长度约20 cm。层面上部及下部岩层可见水平层理及小型交错层理。Seilacher(1967)在建立Zoophycos遗迹相时, 认为其反映的环境为浪基面以下的大陆斜坡半深海环境, 随着研究工作的深入, 越来越多的资料表明, Zoophycos在不同的地史时期, 分布的深度发生过很大变化(Bischoff, 1968; Wetzel and Werner, 1981; Ekdale and Bromely, 1984; Ekdale and Lewis, 1991; Kotake, 1991), 在晚古生代, 常常出现在滨浅海的陆架上部环境(杨式溥等, 2004)。因此, 养马坝组中上部可划分为Zoophycos遗迹相(图 3-A), 反映了浅海陆棚的沉积环境。
对甘溪剖面遗迹相的系统分析表明, 该地区早、中泥盆世的环境变迁是:平驿铺组的遗迹相变化为Skolithos 遗迹相→ Cruziana遗迹相→ Skolithos 遗迹相, 反映了海侵由滨海潮间带开始, 随着时间的推移海水逐渐加深, 至甘溪组的Rhizocorallium遗迹相, 海侵达到最大; 尔后从甘溪组到谢家湾组出现了一个海退序列, 谢家湾组代表了较浅水的近岸潮坪环境; 从谢家湾组至养马坝组代表了一个新的海进序列, 至养马坝组的Zoophycos遗迹相, 海侵达到最大, 至养马坝组顶部海侵结束。
广西横县六景剖面是华南泥盆系著名的标准剖面之一, 位于南宁市东约60km处的湘桂铁路六景火车站附近, 即北纬22° 53.133', 东经108° 52.739'(图 1)。剖面出露良好, 总长度4600m, 总厚度1247.3 m(图 3-F)。其底界与寒武系呈不整合接触, 顶部为古近系覆盖而出露不全。广西横县六景泥盆系剖面遗迹化石主要分布在下、中泥盆统, 其中下泥盆统遗迹化石集中在莲花山组伶俐段、横县段、六坎口段和那高岭组高岭段、蚂蝗岭段, 郁江组六景段仅在灰色灰质泥岩中发现大量的Chondrites isp.(图5-C), 中上泥盆统少见遗迹化石, 仅在中泥盆统民塘组薄层石灰岩及灰质泥岩中有Treptichnus pollardi的报道(周志澄, 1996)。
Skolithos 遗迹相
莲花山组伶俐段顶部和横县段下部的紫红色厚层砂岩及粉砂岩中发育大量垂直层面的潜穴Skolithos lianhuashanensis和Diplocraterion parallelum(图 5-B)。遗迹化石单调(只有两属), 但是丰度很高, 布满整个层面。Skolithos lianhuashanensis垂直层面分布, 横切面呈圆形, 直径约7~9mm, 保存内生迹, 潜穴内部无构造为被动充填。Skolithos遗迹属被认为是Skolithos遗迹相的指相化石, 常形成于高能环境下的滨海潮间、潮下带的浅水环境, 但偶尔在深海扇及半深海斜坡环境也曾发现(Seilacher, 1967; 杨式溥, 1983; 杨式溥等, 2004)。Diplocraterion parallelum垂直层面分布, 表现为具蹼状构造的U形潜穴, U形潜穴的栖管横切面呈圆柱形, 直径为4mm, 蹼状构造呈前移式, 整个潜穴宽2cm, 深4~5 cm(图 5-B), 常被认为是滤食性动物(多毛类和甲壳动物)的居住构造, 其居住在高能量波浪地带, 常见于滨海地区潮间、潮坪相, 属于Skolithos遗迹相(杨式溥等, 2004)(图 3-F)。
 | 图5 广西横县六景泥盆系剖面遗迹化石Fig.5 The Devonian trace fossils from Liujing section, Guangxi of South China |
A— Arenicolites carbonaria, 保存在粉砂质泥岩中, 简单的无蹼层/板的U形潜穴, U形管垂直于层面, 管壁较粗糙; B— Diplocraterion parallelum, 保存在粉砂质泥岩中, 具蹼状构造的U形潜穴, 垂直层面排列, U形潜穴的栖管较粗糙, 圆柱形末端呈圆桶状延伸到层面之上, 在层面上常见成对的圆形管口, 蹼状构造一般为前进式, 潜穴底部为半圆形; C— Chondrites isp., 保存在粉砂岩中, 丛状分枝潜穴, 分枝密集, 至少见2次分枝, 分枝夹角30° ; D— Palaeophycus curvatus, 弯曲成弧形的柱状潜穴, 长6cm, 直径5mm, 半凸起于红色粉砂岩层面上; E— Planolites kwangsiensis, 水平柱状潜穴, 平行层面, 保存于灰黄色薄层泥质粉砂岩中, 潜穴直径3mm, 长约5 cm, 外表经风化后光滑; F— Skolithos linearis, 保存在粉砂质泥岩中, 与层面垂直的针形潜穴, 潜穴外表光滑, 颜色较围岩深, 潜穴内部无构造为被动充填, 横切面呈圆形; G— Chondrites cf.intricatus, 保存在粉砂质泥岩中, 树枝状分枝潜穴系统, 分枝细密, 直径约1mm, 长5~7mm, 单个潜穴成簇状, 分枝角度约30° , 常见1~2次分枝, 整体呈放射状。 A, D至G为下泥盆统那高岭组, B为下泥盆统莲花山组, C为下泥盆统郁江组
通过对上述遗迹化石古环境指相意义的分析, 结合莲花山组伶俐段和横县段紫红色粉砂岩及砂岩的岩性特征及保存的鱼类、双壳类、介形类、腕足类Lingula等化石, 将该段地层划分为Skolithos 遗迹相, 反映了干热气候条件下的高能河一湖及滨海的沉积环境。
Cruziana 遗迹相
莲花山组六坎口段细砂岩和粉砂岩中, 偶见波痕及交错层理, 上部出现大量的遗迹化石Cruziana, Chondrites, Zoophycos, Palaeophycus, Lumbricaria, 其中主要的遗迹属为Cruziana。根据Selaicher(1967)提出的遗迹相划分标准, 可将六坎口段归为Cruziana遗迹相。这个遗迹相的古海洋深度相当于Seilacher(1967)所划分的滨浅海区, 证明当时随着海水的加深, 水内所含的有机质不断地沉积在海底泥砂岩中, 造迹生物由滤食性逐渐转变为以食泥动物为主(杨式溥, 1983)。Cruziana为三叶虫类利用内肢挖掘形成的觅食潜穴(Seilacher, 1970), Chondrites一般认为是食泥造迹生物形成的觅食潜穴系统(Osgood, 1970), 被认为是广相遗迹。通过上述遗迹化石组合的古环境意义及相伴生的波痕、交错层理等沉积构造, 将本段划分为Cruziana遗迹相(图 3-F), 其指示了滨浅海潮下带水体盐度大体正常、水动力减弱、底部含有丰富有机质的沉积环境。
整个那高岭组沉积时期, 高岭段下部为灰绿色砂质泥岩、泥质粉砂岩, 在高岭段发现的4个遗迹属Chondrites, Planolites, Skolithos(图5-F), Bifungites(杨式溥, 1983)均产在不含实体化石的夹层中, 但同一层内个体数量多、密集成层、潜穴同层面平行或斜交。Bifungites的埋藏环境一般为细碎屑岩, 常见交错层理、波痕和底流痕, 产出的环境为海进或海退作用下的地台陆架浅海、海湾和海岸线附近, 常与Cruziana遗迹相的特征分子共存。Bifungites可能为软躯体滤食性和食细小沉积物动物的居住潜穴, 并非由活动型造迹生物所为(杨式溥等, 2004)。结合那高岭组高岭段泥质增多、粒度变细以及层理不发育、缺乏交错层理和化石大多为原地埋藏、生物分异度增加等特征, 认为自那高岭组底部的海侵开始, 遗迹化石组合属于Cruziana遗迹相(杨式溥, 1983)。
蚂蝗岭段中部灰绿色薄层— 中层状泥质粉砂岩属于高水位体系域的海退期沉积, 在这一时期水体能量相对较大, 海水总体变浅。该段含Arenicolites (图 5-A), Chondrites(图 5-G), Palaeophycus(图 5-D), Planolites (图 5-E), Phycodes, Rhizocorallium, Teichichnus 共7个属的遗迹化石, 含遗迹化石的层位伴生有舌形贝和双壳类化石以及成层排列的磷结核。在分异度方面, Phycodes, Rhizocorallium和Teichichnus为Cruziana遗迹相主要成员, 在丰度方面以广相的Palaeophycus和Planolites等遗迹属最为丰富, 属于交切遗迹组构(龚一鸣等, 1997)。Palaeophycus, Planolites及Chondrites等遗迹属, 个体较小, 管径一般为1~3 mm。 根据上述遗迹化石的古环境指相意义及相关的沉积构造, 将本段划分为Cruziana遗迹相(图 3-F), 反映了海水盐度正常、海水加深、生物分异度增高的潮下带沉积环境。
通过对六景剖面系统的遗迹相分析表明, 该剖面早、中泥盆世的环境变迁是:莲花山组底部伶俐段和横县段的Skolithos遗迹相, 反映了海侵由滨海潮间带开始; 随着时间的推移海水逐渐加深, 至莲花山组六坎口段、那高岭组及郁江组的Cruziana遗迹相, 海侵达到最大; 尔后从郁江组到那叫组出现了一个海退的序列, 那叫组代表了较浅水的近岸潮坪环境。
贵州独山泥盆系剖面属于独山泥盆系— 石炭系标准地质剖面的下段, 位于贵州省独山县一带, 即北纬25° 49.849', 东经107° 30.864'(图 1)。剖面起点始于独山大河口附近, 终点止于独山白虎坡。泥盆系出露完整, 全长约12km, 构造简单, 呈单斜连续出露。在区域地质构造上, 该剖面地处独山箱状背斜的核部和西翼。遗迹化石主要分布在半坡、大河口、白虎坡3个剖面。早泥盆世遗迹化石主要分布在半坡剖面丹林组和舒家坪组、大河口剖面龙洞水组和大河口组。中泥盆世遗迹化石主要分布在大河口剖面屯上组、鸡泡组、宋家桥组和鸡窝寨组。晚泥盆世遗迹化石主要分布在白虎坡剖面尧梭组四方坡段中部和汤耙沟组。独山地区下、中泥盆统遗迹化石十分丰富, 根据遗迹化石的特征和产出层位, 划分为3种反映不同沉积环境的遗迹相(图 3-D), 分述如下:
 | 图6 贵州独山泥盆系剖面遗迹化石Fig.6 The Devonian trace fossils from Dushan section, Guizhou of South China |
A— Rhizocorallium jenense, 保存在砂岩中, 平行于层面分布, 两翼管近平行, 末端较弯曲, 两翼管之间为同心状蹼状构造, 蹼状构造之间的间距变化不定; B— Rosselia socialis, 保存在砂岩中, 垂直或接近垂直的纺锤状或圆锥状潜穴, 与层面呈30° 左右交角, 通道直径和居住潜穴的直径约为1cm, 潜穴深约6cm, 表面呈圆纹状; C— Arenicolites carbonaria, 保存在泥页岩中, 简单的无蹼层/板的U形潜穴, U形管垂直于层面, 管径宽约4mm, 潜穴深度约100mm, 管壁较粗糙; D— Zoophycos isp., 化石保存在粉砂质泥岩/砂岩中, 呈全浮雕产出, 与层面平行椭圆形蹼层, 大蹼纹粗且分叉, 部分小蹼纹充填物与围岩不同; E和F— Dushanichnus dahekouensis, 潜穴管以5° ~15° 的角度与层面斜交后在层面上呈近平行层面伸展, 线状遗迹不分叉或3次分叉, 构成扇形或帚形, 线状遗迹长1.2~5.6 cm, 潜穴管表面光滑, 呈圆柱形或因压扁呈椭圆形, 直径0.4~1.0 cm; G— Planolites beverleyensis, 产在泥质粉砂岩/砂岩层面上, 平行层面分布, 微弯曲, 潜穴长5cm, 直径6mm, 表面光滑/粗糙, 潜穴颜色较母岩深, 无衬里。A和B为下泥盆统舒家坪组, C为下泥盆统丹林组, D为中泥 盆统宋家桥组, E和F为下泥盆统大河口组, G为中泥盆统鸡泡组
Skolithos遗迹相
在下泥盆统丹林组顶部深灰色泥质粉砂岩、舒家坪组底部浅灰色石英砂岩、大河口组底部浅灰色石英砂岩及尧梭组四方坡段中上部浅灰色白云岩中, 共发育9个属的遗迹化石, 分别是:Arenicolites, Diplocraterion, Dushanichnus(图 6-E, 6-F), Monocraterion, Skolithos, Planolites(图 6-G), Palaeophycus, Rhizocorallium(图 6-A)和Rosselia(图 6-B)。以简单垂直层面的Skolithos最为常见, 其次为Arenicolites, Diplocraterion和Rhizocorallium。Arenicolites carbonaria (图 6-C)主要表现为U形潜穴, 垂直层面, 直径约4mm, 深度约10cm, U形管间距约1cm, 被认为是蠕虫类环节动物和小型甲壳动物的居住构造, 常见于潮坪及滨浅海潮间带环境。Rhizocorallium jenense的 U形管管壁光滑, 管直径为8mm, 宽约12cm, 其他保存特征与甘溪剖面甘溪组第1层的相同, 一般被认为是甲壳动物所形成的觅食潜穴(Fü rsich, 1974)。
与上述遗迹组构相伴生的主要沉积构造有楔状交错层理和水平层理等。通过上述遗迹化石的古环境指相意义及相关的沉积构造, 将下泥盆统丹林组、舒家坪组、大河口组划分为Skolithos遗迹相(图 3-D), 反映了滨海潮间带— 潮下带的水动能较强的沉积环境。
在F-F集群灭绝事件之后, 在尧梭组四方坡段白云岩中发育一些潜穴管细小、分异度低、属于浅阶层的觅食潜穴。尧梭组四方坡段主要为一套潮间带沉积, 因此将该组划分为Skolithos遗迹相(图 3-D)。这一丰度高而分异度低的遗迹化石组合通常发育于海平面快速变化形成的海进体系域和高位体系域中(王约等, 2006)。
Cruziana遗迹相
在上泥盆统者王组灰色薄层至中层状灰岩、革老河组灰黑色中厚层状灰岩、瘤状灰岩以及层间泥灰岩夹层(上部为灰色中厚层状灰岩)和汤耙沟组灰色厚层状钙质波状条带灰岩中, 主要发育大量爬行迹、表层觅食潜穴、停息迹等10个属种的遗迹化石, 分别是:Rhizocorallium, Teichichnus, Thalassinoides, Palaeophycus, Planolites, Chondrites, Ophiomorpha, Diplocraterion, Phycodes和Cochlichnus(王约, 2004; 王约等, 2006)。垂直层面方向上下移动的进食迹Teichichnus和层面上呈网孔状的分叉垂直潜穴系统Thalassinoides为主要的遗迹属, 此外, 近平行于层面的进食潜穴Phycodes和系统潜穴Chondrites也占有优势。因此这一段划分为Cruziana遗迹相, 反映了食物丰富、光照充足、盐度正常、中至低能的开阔陆棚浅海沉积环境(王约, 2004)。
Zoophycos 遗迹相
在中泥盆统鸡泡组生物碎屑灰岩或泥灰岩、宋家桥组下部浅灰色石英砂岩以及鸡窝寨组泥灰岩或黑色钙质页岩中, Zoophycos及Rhizocorallium最为常见。如宋家桥组下部浅灰色石英砂岩中的Zoophycos isp.(图 6-D)沿层面平行展布, 表现为圆弧形边缘细长管道潜穴, 具有大型蹼状构造, 显示为同心弧线, 无明显的顶弧与侧弧之别, 宽度约10cm, 长度约16 cm。
独山地区根据围岩及相伴生的沉积构造的不同, 从鸡泡组→ 宋家桥组→ 鸡窝寨组沉积时期, Zoophycos遗迹相反映了由滨浅海潮下带向陆棚过渡的沉积环境, 也指示了海水的加深, 并在鸡窝寨组沉积时期达到最大, 因此, 含有Zoophycos的深水页岩层位常被视为海平面上升至最高转折点的饥饿期沉积或“ 凝缩段” (龚一鸣等, 1997; 王约和龚一鸣, 1997)。
从对独山剖面系统的遗迹相分析看, 该地区泥盆纪的环境变迁是:从丹林组底部至大河口组的Skolithos遗迹相, 反映了海侵由滨海潮间带开始, 随着时间的推移海水逐渐加深, 在鸡窝寨组(Zoophycos遗迹相)海侵达到最大; 尔后从望城坡组到尧梭组出现了一个海退的序列, 尧梭组代表了较浅水的近岸潮坪环境; 而从尧梭组四方坡段至革老河组又出现了新的海进序列, 由四方坡段的Skolithos遗迹相过渡到革老河组的Cruziana遗迹相, 表明海水逐渐加深的过程。
乌当情人谷剖面位于距贵阳市中心东约7km的乌当区旅游景点情人谷内, 即北纬26° 36.814', 东经106° 48.206'(图 1)。剖面出露良好, 总长度1000m, 总厚度296.61 m。其底界与志留系高寨田组呈平行不整合接触, 顶部与石炭系祥摆组呈平行不整合接触。遗迹化石主要分布在中泥盆统, 即蟒山群马鬃岭组顶部及高坡场组底部地层中(陈云明等, 2005)(图 3-C)。
Skolithos遗迹相
贵阳乌当蟒山群马鬃岭组产出的遗迹化石有简单垂直潜穴Skolithos, Conichnus wudangensis; 具蹼状构造的垂直U形潜穴Diplocraterion; 与层面微斜交的、具蹼状构造的U形潜穴Rhizocorallium(图 7-B); 与层面微斜交的觅食潜穴Teichichnus; 系统潜穴Chondrites(图 7-F), Thalassinoides, Phycodes palmatus(图7-A); 水平觅食潜穴 ?He-lminthopsis isp.(图7-D), Palaeophycus tubularis(图 7-C), Planolites(图 7-E)以及觅食迹Cruziana。贵阳乌当马鬃岭组的遗迹化石以垂直潜穴占优势, 系统潜穴和觅食潜穴也有相当的数量, 属于Seilacher(1967)划分的Skolithos遗迹相, 反映了水动力较强的砂质移动底质的滨岸沉积环境。
Cruziana遗迹相
贵阳乌当关山高坡场组产出的遗迹化石有水平觅食潜穴Planolites, Palaeophycus; 三叶虫/节肢动物觅食爬痕Cruziana; 与层面微斜交的、具蹼状构造的U形潜穴Rhizocorallium, 系统潜穴Chondrites, 其中Chondrites和Planolites遗迹属最为丰富。该遗迹组构的特征为分异度高、且遗迹大都沿层面或低角度斜交层面展布、中深和深阶层的遗迹Chondrites交切Palaeophycus。贵阳乌当高坡场组的遗迹化石以系统潜穴和觅食潜穴占有优势, 属于Cruziana遗迹相, 反映食物丰富、光照充足、盐度正常、中至低能的潮下带沉积环境。
杨堤剖面位于广西桂林至阳朔公路60km处的杨堤路口, 即北纬 24° 58.265', 东经110° 22.872'(图 1)。整个杨堤剖面缺失下泥盆统以及上泥盆统法门阶上部地层, 实测总厚度255.4 m。遗迹化石主要分布在中上泥盆统中, 即信都组、东岗岭组中部和五指山组底部(图 3-B)。
Skolithos遗迹相
中泥盆统信都组浅灰色细粒砂岩夹粉砂岩具交错层理和干涉波痕, 含遗迹化石Chondrites, Planolites(图 8-F), Palaeophycus, Rhizocorallium(图 8-E)和 Skolithos。Palaeophycus和Planolites为主要的遗迹属, 而垂直层面的居住潜穴Skolithos和系统觅食潜穴Chondrites少见。Planolites和Palaeophycus属广相型的常见分子, 根据遗迹化石的指相意义及相伴生的沉积构造, 将本段地层划分为Skolithos遗迹相, 反映了滨海潮间带的沉积环境。
Cruziana遗迹相
中泥盆统东岗岭组底部生物化石种类丰富, 包括珊瑚、枝状层孔虫、块状层孔虫等, 向上生物化石变得单调, 但含量仍较多。东岗岭组中部深灰色中薄层泥晶石灰岩夹藻纹层白云岩中含层孔虫、腹足类、腕足类和遗迹化石Planolites。该遗迹相以小型的Planolites montanus的大量出现为特征, Planolites montanus 是一种层内进食潜穴, 个体直径1mm 左右, 延伸长度约5~10mm, 平行层面分布, 不分枝; 在横剖面上表现为短小的蠕虫状潜穴、少数为短杆状, 在纵剖面上则呈蠕虫状弯曲; 潜穴表面光滑, 充填物颜色明显深于围岩。Planolites montanus常密集成群分布, 对沉积物造成强烈的扰动, 扰动指数一般可达3~5。
上泥盆统五指山组底部薄层石灰岩中发育遗迹化石Circulichnis(图 8-B), Thalassinoides isp.(图 8-A, 8-C, 8-D)和Chondrites, 遗迹化石丰度很高, 布满整个层面, 但遗迹属种单调。Chondrites为主要的遗迹属, 保存为表生迹, 呈分枝潜穴, 至少可见3次分枝, 潜穴直径为5 mm。 Chondrites被众多学者认为是缺氧环境的指示剂(Ekdale and Bromley, 1984; Seilacher, 1990; Fu, 1991; 龚一鸣, 2004), 常见于Cruziana遗迹相(杨式溥等, 2004), 而遗迹化石Circulichnis montanus也在Cruziana遗迹相中出现过。根据上述遗迹化石的指相意义及相伴生的沉积构造, 将中上泥盆统东岗岭组及五指山组划分为Cruziana遗迹相, 反映了由局限台地相向台地边缘斜坡相过渡的沉积环境。
 | 图7 贵州乌当泥盆系剖面遗迹化石Fig.7 The Devonian trace fossils from Wudang section, Guizhou of South China |
A— Phycodes palmatus, 潜穴粗而圆, 由同一点向末端方向不规则分叉, 标本有4个像手掌状的分枝潜穴, 主潜穴直径1.5cm, 分枝潜穴直径为1cm左右, 潜穴系统长10cm, 单个潜穴长3.5~4cm, 分枝潜穴内有蹼状回填构造, 在主潜穴顶端出现S形弯曲, 砂岩, 上层面; B— Rhizocorallium jenense, 保存在砂岩中, 平行于层面分布, 两翼管近平行, 末端较弯曲, 翼管长16cm, 直径10mm, 两翼管之间为同心状蹼状构造, 蹼状构造之间的间距变化不定; C— Palaeophycus tubularis, 弯曲成弧形柱形潜穴, 半凸起于灰色砂岩层面上, 不分枝, 充填物同围岩, 直径3mm, 潜穴长约7 cm; D— ?Helminthopsis isp., 保存在砂岩中, 不规则蛇曲形潜穴, 不分枝, 粗细一致, 潜穴直径约5mm, 平行于层面分布, 遗迹长约6 cm; E— Planolites isp., 保存在砂岩中, 短柱形潜穴, 充填物与围岩不同, 潜穴直径约8 mm; F— Chondrites filifalx, 保存在砂岩中, 小型细枝状潜穴系统, 分枝呈放射状排列, 潜穴呈细枝, 直径0.5mm, 长10mm, 分枝密集, 分枝角度30° , 遗迹平行层面分布。A至F为中泥盆统马鬃岭组
 | 图8 广西桂林杨堤和四川广元后高坪泥盆系剖面遗迹化石Fig.8 The Devonian trace fossils from Yangdi section, Guangxi and Hougaoping section, Sichuan of South China |
A, C, D— Thalassinoides isp., 保存在石灰岩中, Y字形分枝潜穴, 表面光滑或偶具小瘤, 在水平面上呈多枝网格状互相连接或垂直; B— Circulichnis isp., 保存在石灰岩中, 沿层面或底面分布的圆圈形或椭圆形潜穴, 构成完全闭合的圆形遗迹, 圆环直径约6cm, 此标本仅留圆环的一部分; E— Rhizocorallium isp., 保存在砂岩中, 平行于层面分布, 两翼管近平行, 翼管遭剥蚀, 蹼状构造清晰, 潜穴长约16cm, 两翼管间距为5 cm; F— Planolites beverleyensis, 保存在砂岩层面上, 平行层面分布, 微弯曲, 潜穴长10cm, 直径4mm, 表面粗糙, 潜穴颜色较母岩深, 无衬里; G— Zoophycos isp., 保存在粉砂质泥岩中, 呈全浮雕产出, 与层面平行、具有蹼状构造的椭圆形同心状潜穴构造。蹼状构造粗而分叉, 充填物与围岩不同; H— Chondrites maqianensis, 保存在粉砂质泥岩中, 呈全浮雕产出, 与层面平行、至少可见2次分枝的树冠状潜穴系统, 潜穴管较为细小, 每一分枝的潜穴粗细近于一致, 分枝外的潜穴较粗。A至D为上泥盆统五指山组, E和F为中泥盆统信都组, 广西桂林杨堤剖面; G和H为上泥盆统土桥子组, 四川广元后高坪剖面
四川广元后高坪剖面位于四川广元长江沟佛三村后高坪, 即北纬32° 20.155', 东经 105° 26.535'(图 1), 整个剖面以上泥盆统沙窝子组碳酸盐岩和土桥子组中粒石英砂岩为主。土桥子组石英砂岩中见小型交错层理, 中部一层泥岩及粉砂岩(图 3)中发育丰富的Zoophycos(图 8-G)和Chondrites(图 8-H), 归为Zoophycos遗迹相。剖面上部主要以石灰岩及白云岩沉积为主。从整个剖面的沉积构造、遗迹相及岩石组合上来看, 沉积环境由早期滨海较粗碎屑岩和浅海中上部细碎屑岩相过渡到晚期碳酸盐岩台地相(张立军和龚一鸣, 2009)。
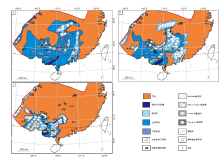
通过搜集和统计前人在华南泥盆纪相关剖面的遗迹化石研究成果, 结合作者的实际工作, 将华南泥盆纪遗迹化石属种进行分类(图 9, 图10), 按时代编制出中国南方早、中、晚泥盆世遗迹相分布图(图 11)。
 | 图9 华南泥盆纪剖面遗迹化石分布和遗迹相示意图(A— 华南各剖面遗迹化石产出位置及古环境示意图(据Seilarcher, 2007, 有修改), B— 华南中泥盆世古地理图及剖面分布位置(据赵锡文, 1985; 吴诒等, 1997; 有修改))Fig.9 Sketch showing distribution of trace fossils and ichnofacies in the Early, Middle and Late Devonian from South China |
从图9可以看出, 华南泥盆系中先后发现的37个遗迹属的遗迹化石可以划分为4种遗迹相, 包括3种原型遗迹相(Skolithos遗迹相、Cruziana遗迹相、Zoophycos遗迹相)及1种地区性遗迹相(Rhizocorallium遗迹相), 4种遗迹相特征及指相意义见表1。 Skolithos遗迹相主要以Skolithos, Arenicolites, Diplocraterion为主要遗迹属, 同时伴生少数Rosselia等, 主要发育在浪控型海岸体系、潮控型海岸体系和障壁岛— 潟湖沉积体系的高能区, 如前滨— 中上临滨、潮下砂坪— 潮道、障壁岛等区域; Cruziana遗迹相以Cruziana, Rusophycos和Phycodes为主, 伴生少数Chondrites和Rhizocorallium, 主要沿浅海陆棚具有丰富生物及食物的砂泥岩和灰岩为主的低能沉积区分布; 介于上述两个遗迹相之间, 根据泥盆纪沉积环境的特点, 提出了Rhizocorallium遗迹相, 主要以Rhizocorallium和Thalassinoides为特征属, 伴生Palaeophycus和Planolites等, 发育在潮控型海岸体系和障壁岛— 潟湖沉积体系的低能区, 如潮间带、潮下带浅水区、潟湖— 沼泽区; Zoophycos遗迹相主要以Zoophycos为主, 伴生Thalassinoides, Chondrites等, 主要沿泥岩和泥灰岩发育的滨外沉积区分布。
从图11可以看出, 华南泥盆纪遗迹化石组合和遗迹相从早泥盆世到晚泥盆世的演替规律是从Skolithos遗迹相→ Rhizocorallium遗迹相→ Cruziana遗迹相→ Zoophycos遗迹相→ Rhizocorallium遗迹相。早泥盆世晚期以后华南泥盆纪同时期的遗迹化石组合和遗迹相自西南向东北的空间变化规律则表现为Zoophycos遗迹相→ Cruziana遗迹相→ Rhizocorallium遗迹相→ Skolithos遗迹相。
在湘桂海域, 由于受北东向断裂的控制, 海侵自西南向东北方向推进, 在垂向上逐渐由碎屑岩沉积演变为碳酸盐岩沉积, 在横向上海侵范围扩大, 碳酸盐岩的沉积范围逐渐扩大, 硅质碎屑岩沉积逐渐向大陆内部推进; 在古陆边缘的滨岸碎屑岩沉积区主要发育Skolithos遗迹相, 以六景剖面为代表的桂中地区早泥盆世遗迹相由潮间高能带的Skolithos遗迹相过渡到浅海陆架的Cruziana遗迹相(图 3-F和图10-❶), 其中Skolithos遗迹相和Rhizocorallium遗迹相在空间上可以重叠。在黔东南海域, 由海平面变化引起的沉积相带的迁移和碎屑滨线的进退是以递进式方式向大陆内部推进的, 贵州独山早泥盆世遗迹相由滨海高能带的Skolithos遗迹相过渡到滨浅海相的Rhizocorallium遗迹相(图 10-❸)。在广大的滇黔桂地区, 由于遗迹化石工作开展较少, 根据深度及沉积相等推断这一地区主要以Rhizocorallium和Cruziana遗迹相为主。在川西北海域, 海侵自西向东推进, 遗迹相在甘溪剖面主要发现为早期的滨海高能带的Skolithos遗迹相向晚期滨浅海低能带变化的Rhizocorallium遗迹相(图 3-A, 图11-A)。
 | 图10 华南泥盆纪遗迹相时空分布断面图(❶— 广西横县六景, ❷— 贵州桑郎水库, ❸— 贵州独山白虎坡, ❹— 贵州贵阳乌当, ❺— 广西桂林杨堤, ❻— 广东花县赤泥, ❼— 广东韶关天子岭, ❽— 湖南新化炉观, ❾— 湖南大庸— 张家界, — 湖南醴县羊耳山水库)Fig.10 Transect map showing spatiotemporal variations of the Devonian ichnofacies in South China |
 | 图11 华南早、中、晚泥盆世遗迹相分布Fig 11 Sketch showing the Early, Middle and Late Devonian ichnofacies distributions in South China |
| 表1 华南泥盆纪遗迹相特征及古环境意义 Table1 Characters of the Devonian ichnofacies and their palaeoenvironmental significance in South China |
在湘桂海域, 至东岗岭组沉积中期, 发生了一次快速的沉积相带迁移, 碳酸盐沉积迅速扩展至广西全境及湖南大部分地区, 碎屑滨线也从桂东北— 湘南一带迅速后退至湘中益阳一带, 从东岗岭组沉积早期至东岗岭组沉积中期迁移了350~400km, 在滨线迅速迁移的同时, 广西海域几乎同时出现了数条以硅质岩— 黑色泥岩为主的台(间)沟深水沉积, 标志着碳酸盐岩台地进入到一个新的演化阶段; 六景剖面中泥盆统民塘组主要为Cruziana遗迹相(图 3-F, 图10-❶), 桂林杨堤剖面中泥盆统信都组反映滨岸沉积Skolithos遗迹相, 过渡到榴江组反映台间盆地沉积的Cruziana遗迹相下部和东岗岭组反映潮下带沉积的Cruziana遗迹相上部(图 3-B, 图10-❺)。湘西南宁远半山剖面, 早一中泥盆世的碎屑岩地层是上覆于湘桂加里东褶皱基底之上的一套海侵碎屑岩系, 通过岩石薄片粒度分布及沉积构造特征将其划分为由障壁海岸到无障壁海岸再到浅海陆棚的沉积环境, 结合遗迹化石组合划分为由早泥盆世早期的Rhizocorallium遗迹相变化为中泥盆世晚期的Cruziana遗迹相(龚一鸣, 1987)。在湖南大庸张家界的碎屑岩中, 发现了大量Cruziana遗迹相的化石组合(图 9)(张欣平等, 1986), 湘西北、鄂西南地区的陆源碎屑岩地区可划分为Skolithos遗迹相。与此同时, 海侵的加大使得在赣西粤北地区发育大量沿古陆分布的陆缘碎屑岩, 依据相关岩相及沉积构造可划分为Skolithos遗迹相。在黔东南海域, 从沉积岩的空间分布看, 陆源碎屑岩主要沿古陆边缘分布, 向海依次出现正常浅海灰岩— 瘤状灰岩、黑色页岩或硅质岩。贵州贵阳乌当蟒山群马鬃岭组Skolithos遗迹相(图 3-C, 图10-❹)
在空间上沿古陆边缘高能带分布, 贵州独山屯组上部— 鸡泡组的Cruziana遗迹相反映了开阔浅海陆棚的沉积环境, 鸡窝寨组Zoophycos遗迹相产于黑色页岩中, 反映了中泥盆世海侵达到最大的临界值(图 3-D, 图10-❸)。在川西北海域, 则由中泥盆世早期的Cruziana遗迹相, 变化为中泥盆世晚期保存在养马坝组水平层理发育的粉砂岩中的Zoophycos遗迹相(图 3-A), 反映出海侵进一步扩张的特征, 由水动力较强的滨岸沉积环境转变为浅海陆架的沉积, 并在Zoophycos遗迹相发育时海侵达到最大, 这与黔东南海域独山剖面中泥盆世遗迹相的变化(图 11-B)几乎是同时的。
在湘桂海域, 海侵达到最大, 扩张到鄂西及鄂东, 湖南新化晚泥盆世佘田桥组和锡矿山组泥岩和泥灰岩中, 划分出反映古陆边缘潮上带沉积环境的Skolithos遗迹相、台地边缘及浅海陆棚相的Rhizocorallium和Cruziana遗迹相(张欣平等, 1986), 桂林杨堤剖面五指山组底部的Cruziana遗迹相, 则反映了台地边缘斜坡相的沉积环境。在黔东南海域, 晚泥盆世早期末时, 正常浅海碳酸盐岩曾一度扩展到都匀以北地区, 代表了泥盆纪的最高水位期。泥盆纪末, 碳酸盐岩分布范围再度扩展到都匀以北, 最后以碎屑滨线向海进积而告终, 代表了泥盆纪最后一次海水进退变化历史。贵阳乌当晚泥盆世高坡场组的Cruziana遗迹相(图 3-C和图10-❹)反映了潮下带水动力较弱的沉积环境, 而贵州独山剖面晚泥盆世可划分为由早期的反映潮下高能带的Skolithos遗迹相至晚期反映陆棚相的Cruziana遗迹相(图 3-D和图10-❸)。在川西北海域, 由于主要为碳酸盐岩与白云岩的沉积, 仅在广元后高坪剖面粉砂质岩层中见Zoophycos和Chondrites(张立军和龚一鸣, 2009), 依据岩性及沉积构造特征将其划分为潮坪— 碳酸盐岩台地— 潮间带的沉积环境(图 11-C)。
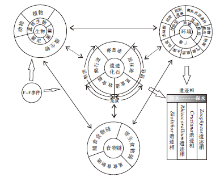
生物包含3个域, 即细菌、古菌和真核生物, 进一步可分为3个大类, 即动物、植物和微生物(顾德兴, 2000)。现代生物与环境之间的相互作用是通过食物链来体现的, 动物— 动物、植物— 微生物、动物— 微生物之间形成了3条普遍存在的食物链, 即捕食食物链、寄生食物链和觅食食物链(顾德兴, 2000; 李博等, 2000)。在地质历史时期中, 遗迹化石正是对上述生物生活习性的反映, 同时遗迹化石的保存形态及围岩又能反映当时的生态环境。按照造迹生物的生活习性, 遗迹化石可以划分为5种主要的习性类型, 即牧食迹、觅食迹、居住迹、爬行迹、停息迹, 其中牧食迹、觅食迹和爬行迹可认为是生物觅食食物链和捕食食物链在地质历史当中的反映。因此, 可以从两个方面来探讨遗迹化石及遗迹相分布的控制因素。
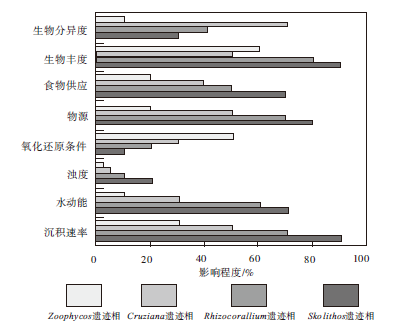
从图12可以看出, 控制遗迹化石分布的环境因素可分为两大类, 即广谱因素和局域因素。广谱因素主要包括底质、盐度、水深和气候, 局域因素可细分为浅阶层和浅— 深阶层两小类, 浅阶层因素主要包括水动能、浊度和沉积速率; 浅— 深阶层因素包括氧化还原条件、食物供应和物源。泥盆纪时期, 扬子板块位于赤道附近, 广谱因素如气候及盐度等条件较为一致, 而局域因素则因地而异, 如氧化还原条件对Zoophycos分布的影响最大, Zoophycos在含氧量低和软底质的滨外泥岩及泥灰岩沉积区分布最为广泛。由于无法将上述影响因素定量分析, 因此现仅从定性的角度对影响华南遗迹化石及遗迹相分布的环境因素进行分析(图 13)。从图13可以看出, 食物供应、物源、浊度、水动能及沉积速率等因素对于遗迹相的影响, 自Skolithos遗迹相→ Rhizocorallium遗迹相→ Cruziana遗迹相→ Zoophycos遗迹相依次减弱, 而氧化还原程度对Zoophycos遗迹相的影响最大。
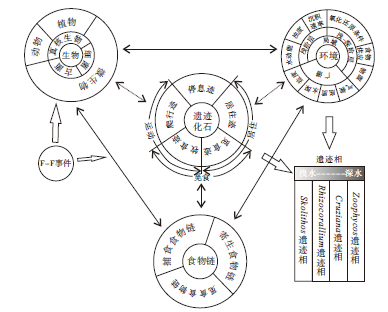 | 图12 华南泥盆纪遗迹化石所反映的生物与环境之间的相互作用Fig.12 Sketch showing interaction between organism and environments indicated by the Devonian trace fossils from South China |
1)生物多样性。早泥盆世— 晚泥盆世早期, 生物界异常繁盛, 尤以低纬度为最, 鱼类以及底栖生物腕足类、珊瑚等为此时期代表性生物, 而陆生植物如裸子植物、高大乔木和大型森林此时也相继出现。生物种类的繁盛导致生物之间的竞争加强, 食物链逐渐增多, 由此遗迹化石的种类及数量亦随之增多, 因此在早泥盆世— 晚泥盆世早期, 扬子板块遗迹化石及遗迹相广泛分布于滨浅海地区。通过图13可以发现, 生物的丰度及分异度也对遗迹相的分布存在一定的影响, 生物丰度对Skolithos遗迹相影响最大, 生物分异度则对Cruziana遗迹相影响相对较高。
2)F-F灭绝事件。发生于晚泥盆世弗拉期与法门期之交的生物集群灭绝事件造成了严重的后果, 例如泥盆纪生物礁和具泥盆纪特征的珊瑚、层孔虫和苔藓虫等基本灭绝, 全球151种珊瑚全遭劫难, 47个浅海相珊瑚也只有2~3个属残存(廖卫华, 2001)。此外, 生物的类型也发生了较大的变化, 如F-F事件之前珊瑚骨骼及其微细结构都属于泥盆纪类型, 而F-F事件之后则属于石炭纪或接近石炭纪类型(廖卫华, 2001)。低纬度地区礁生态系、浅水海相生物受到重创, 而高纬度生态系、深水生物和陆生动、植物受影响甚小(白顺良, 1998; Droser et al., 2000)。在 F-F 事件后, 台地相和礁相的珊瑚几乎全部灭绝(廖卫华, 2001); 又有研究指出, 菌藻类的繁盛是F-F事件的原因之一(龚一鸣等, 2002, 2005; 徐冉等, 2006)。上述研究表明, 扬子板块在晚泥盆世处于赤道附近, 浅海相的生物受到重创, 导致生物类型减少, 直接影响到食物链的存在。此外, 由于菌藻类繁盛及赤潮的出现, 滨浅海相浮游藻类繁盛, 致使海水底层缺氧, 影响到底栖造迹生物的正常生活。上述因素都影响到遗迹化石的分布, 但局部地区例如独山地区, 由于其特殊的地理位置, 所受影响较小, 先后发育了一些潜穴直径细小的Planolites, 其造迹生物主要是对于生态环境条件的要求较低的食泥性生物(王约等, 2006)。因此, F-F灭绝事件对于泥盆世生态系统的重创, 导致了生物类型及丰度的降低, 同时破坏了生态系统的食物链及营养级, 进而导致晚泥盆世晚期华南遗迹化石的发育及遗迹相的组成及分布。
1)通过对华南6条剖面精细的遗迹学研究, 鉴定并描述了遗迹化石15属24种, 同时根据遗迹化石之间的共生组合关系、实体化石特征和其他相标志, 识别出4种遗迹相(Skolithos遗迹相、Rhizocorallium遗迹相、Cruziana遗迹相和Zoophycos遗迹相)。Skolithos遗迹相主要发育于浪控型海岸体系、潮控型海岸体系和障壁岛— 潟湖沉积体系的高能区, 如前滨— 中上临滨、潮下砂坪— 潮道、障壁岛等区域; Rhizocorallium遗迹相主要发育于潮控型海岸体系和障壁岛— 潟湖沉积体系的低能区, 如潮间带、潮下带浅水区、潟湖— 沼泽区; Cruziana遗迹相分布于以具有丰富生物及食物的砂泥岩和灰岩为主的浅海陆棚低能沉积区; Zoophycos遗迹相分布于泥岩和泥灰岩发育的滨外沉积区。
2)华南泥盆纪遗迹相从早泥盆世到晚泥盆世的演替规律为Skolithos遗迹相→ Rhizocorallium遗迹相→ Cruziana遗迹相→ Zoophycos遗迹相→ Rhizocorallium遗迹相。早泥盆世晚期以后华南泥盆纪同时期的遗迹相自西南向东北的空间变化规律则表现为Zoophycos遗迹相→ Cruziana遗迹相→ Rhizocorallium遗迹相→ Skolithos遗迹相。
3)控制华南泥盆纪遗迹相分布的因素主要有生物及环境因素, 生物因素主要包括生物的多样性及F-F灭绝事件; 环境因素可分为两大类:广谱因素和局域因素, 前者包括底质、盐度、气候和水深, 后者又可细分为浅阶层和浅— 深阶层因素, 浅阶层因素包括水动能、浊度和沉积速率, 浅— 深阶层因素包括食物供应、物源及氧化还原条件。
致谢 感谢在野外工作中给予帮助的王约教授和徐冉博士以及曾剑威、冯启、曾雄伟、何磊、金晓波、陈慧等同学!感谢对文稿提出宝贵修改意见的齐永安教授!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|