
{kind=link}
{kind=link}
{kind=link}
{kind=link}
河北省石家庄地区晚更新世晚期古菱齿象-披毛犀动物群及其古气候*
[董明星1  , 张祥信
, 张祥信1 , 牛树银1 , 庞其清1 , 边鸿浩2 , 张倩1 ]
, 张祥信|
|

河北省石家庄市郊区地下 7~24m的古滹沱河沉积物中产出丰富的大型哺乳类化石:诺氏古菱齿象(Palaeoloxodon naumanni)、原始牛(Bos primigenius)、杨氏水牛(Bubalus youngi)、披毛犀(Coelodonta antiquitatis)、鹿(Cervus sp.)、四不像鹿(Elaphurus sp.)等,其中以诺氏古菱齿象和杨氏水牛数量最为丰富。在该化石层之上约 30cm处的针叶树茎杆保存完整,直径可达 20~50cm,年轮均匀清晰, 14C年龄为 8150±120 a BP。冲积层及其中的古哺乳类化石记录了晚更新世最晚期向全新世过渡期间石家庄地区由冷湿向干热转变的过程。
About the first author:Dong Mingxing,born in 1966,is an associate professor of Shijiazhuang University of Economics.He is mainly engaged in physical geography.E-mail: dmy27@hotmail.com.
第一作者简介:董明星,男, 1966年生,石家庄经济学院副教授,自然地理专业。通讯地址:石家庄槐安东路 136号;邮编: 050031。E-mail: dmy27@hotmail.com。
Occurring at 7~24m below the surface of current deposits of the ancient Hutuo River,the excavated fossils in the suburb of Shijiazhuang include Palaeoloxodon naumanni, Bubalus youngi, Bos primigenius, Coelodonta antiquitatis, Cervus sp., Elaphurus sp. etc.,among which the most abundant forms are Palaeoloxodon naumanni and Bubalus youngi,which coexist with fossil conifers in the same sequence in Hutuo River deposits.A δ14C age date of 8150±120 a BP is obtained from a fossil conifer,whose diameter varying between 20 cm and 50cm,containing sensitive tree-ring.The macro-mammalian fauna records the climate change from temperate cold to warm,humid to arid in Shijiazhuang area from the later Late Pleistocene to the Holocene.
晚更新世晚期, 全球气候剧烈变化, 甚至百年内气温波动达15 ℃以上(Hofreiter and Stewart, 2009)。气候的变化极大地影响了哺乳动物的种类(Lister and Stuart, 2008)。欧洲中北部、亚洲北部、北美北部向干冷的草原化方向发展。大型哺乳类如长鼻类、犀类等随气候波动而迁徙, 但最终在末次冰期结束前灭绝。河北省石家庄地区地处华北平原腹地, 洪、冲积地层中埋藏了丰富的哺乳类化石, 记录了这一特殊时期的环境变化。
石家庄市位于太行山前的洪、冲积平原上, 该平原主要由滹沱河洪(冲)积扇构成(图 1)。滹沱河具有明显的河谷形态, 系切割晚更新世冲积扇而形成。地势总体自西向东、自西北向东南倾斜, 地面平均坡降1.5‰ 左右。
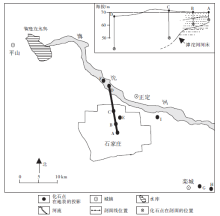
Fig.1 Distribution of Palaeoloxodon-Coelodonta fauna in Shijiazhuang area'>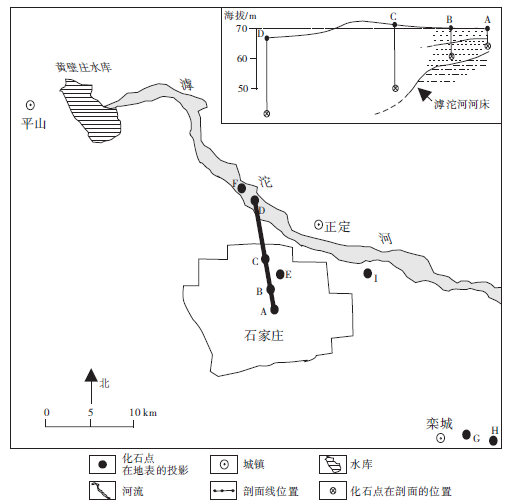 | 图1 石家庄地区的古菱齿象-披毛犀动物群化石分布(点A位于石家庄市裕华西路与中华南大街交口, 点B位于石家庄市维明南大街与自强路交口, 点C位于石家庄市翔翼路与友谊北大街交口, 点D位于滹沱河河床 (114° 55'96″E, 38° 11'84″N), 其他化石点位置见文中)Fig.1 Distribution of Palaeoloxodon-Coelodonta fauna in Shijiazhuang area |
近10 a来, 由于建筑施工和采砂, 在石家庄市周围约400 km2的冲积层中发现了一批很有价值的哺乳动物类化石, 这些化石包括诺氏古菱齿象(Palaeoloxodon naumanni)、原始牛(Bos primigenius)、杨氏水牛(Bubalus youngi)、披毛犀(Coelodonta antiquitatis)、鹿(Cervus sp.)、四不像鹿(Elaphurus sp.)等, 分布于地表之下7~24 m。
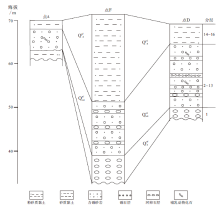
晚更新世至全新世地层以石家庄市区(图1中A)和滹沱河(图1中D、F)出露较好, 以上3个地点的地层对比见图2。A点可见古滹沱河基底, 为细砂质黏土、粉砂质黏土, 层理不清晰, 出露厚0.5m; 向上为滹沱河冲积层, 由透镜状细砂、中细砂组成, 斜层理和槽状交错层理清晰发育, 产古菱齿象门齿(图3), 厚约7m; 上部为洪积层, 由黄土状粉砂质亚黏土组成, 层理不清晰, 厚约2 m。D点(114° 55'96″E, 38° 11'84″N)下部(第1~13层)冲积层未见底, 以含砾砂为主, 夹有多层河卵石, 斜层理、交错层理发育, 古哺乳类化石产于第7层; 上部(第14~16层)为洪积层, 砂质黏土、黏土质粉砂、细砂, 纹层理不清晰, 含有古榆木化石, 不产哺乳类化石。地层由上而下为(分层号见图2):
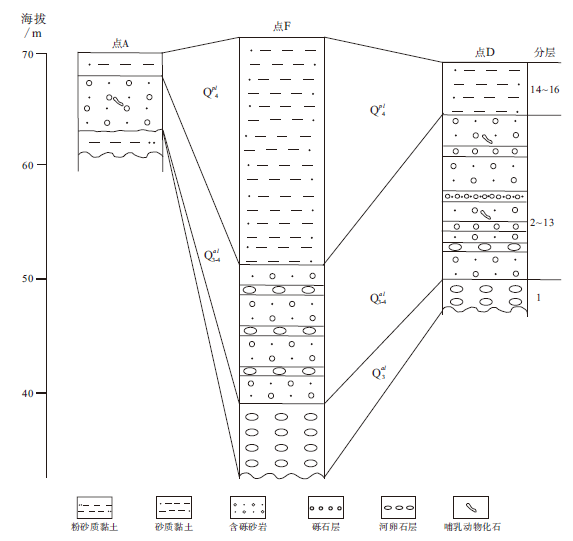 | 图2 石家庄地区A、F、D点洪(冲)积层划分对比Fig.2 Stratigraphic division and correlation of diluvium and alluvium at sites of A, F and D in Shijiazhuang area |
16)黄土状粉砂质亚砂土, 顶部海拔64m, 可见1996年大洪水剥蚀面 2.6 m
15)含砾细砂、粉砂, 呈透镜状, 产榆木化石。碎屑颗粒成分为长石、石英和云母, 分选好, 磨圆度差 3 m
14)砂质黏土、黏土质砂互层 3.2 m
13)细砂、中细砂, 底部有清晰剥蚀面, 剥蚀面上有细砾, 细砾成分为花岗岩岩屑和长石, 层理不发育, 洪积成因 4.1 m
12)含砾砂层, 底部与砾石层之间有剥蚀面, 砾石约5%~10%, 粒径多在2~6cm, 砂占90%以上, 分选差。砾石为次圆状— 次棱角状, 砂质颗粒成分主要为石英、长石和云母, 棱角状 4.1 m
11)河卵石层, 砾石成分为灰岩、花岗岩等, 磨圆度较好, 次圆状 0.1 m
10)中粗砂层, 分选好, 槽状交错层理发育, 磨圆差 0.25 m
9)含针叶树茎杆细砂层, 针叶树茎杆14C年龄8150± 120 a BP 0.3 m
8)细砂层, 底部为10cm厚河卵石层, 产古菱齿象、水牛、原始牛、四不像鹿等化石 0.3 m
7)中粗砂层, 斜层理发育 1.5 m
6)含砾砂层, 砾石含量约5%, 由下至上粒径由粗到细, 斜层理和槽状交错层理发育 3 m
5)含砂砾石层, 灰岩砾石粒径2~5cm, 磨圆度高, 为次圆状 2 m
4)中粗砂层, 含砾石, 斜层理发育 0.7 m
3)砾石层, 楔状或透镜状, 夹有砂粒, 斜层理发育 0.4 m
2)中细砂层, 发育大型槽状交错层理 0.6 m
1)河卵石层, 直径多在10~1cm, 磨圆度较好, 分选中等 > 1.5 m
动物群中以古菱齿象化石(图3)数量最多, 分别发现于石家庄市区洪冲积层地下7 m、23.5 m(图1中A、C)、石家庄市长安区财富大厦(图1中 E)地下12, m、栾城县龙化村(图1中H)地下9, m、栾城县胡家寨村(图1中G)地下16 m、滹沱河河床(图1中D)23.5m以及西塔口村(图1中Ⅰ )等地; 水牛、原始牛和四不像鹿化石(图4)发现于滹沱河河床(图1中D)23.5m处; 披毛犀化石(图3)发现于图1中B点地下10m处; 四不像鹿化石(图4)还发现于石家庄市区(图1中C)地下23.5m处; 在图1中D点之下23.2m处比化石稍高层位产出有23件直径20~50cm的针叶树茎杆化石(图4); 榆木则发现于地表下5.6m处。距离滹沱河越远, 产于河流相地层中的哺乳类化石埋深越浅(图 1)。
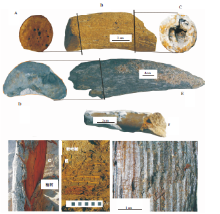
 | 图3 石家庄地区的古菱齿象-披毛犀动物群化石照片(一)(A~D为Palaeoloxodon naumanni, A为M3, B为m3, C为门齿断面, D为下颌骨; E为Bubalus youngi头骨; F为Coelodonta antiquitatis带m3和m2右下颌)Fig.3 Photos of Palaeoloxodon-Coelodonta fauna in Shijiazhuang area(Ⅰ ) |
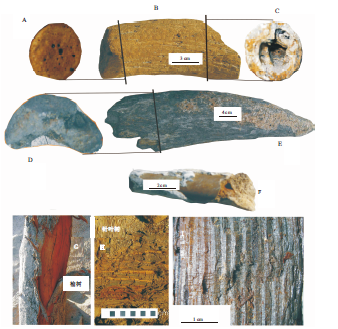 | 图4 石家庄地区的古菱齿象-披毛犀动物群化石照片(二)(A、B、C分别为Bos primigenius角的远端断面、角及角的近端断面; D、E为Bubalus youngi角的近端断面和右角; F为Elaphurus davidianus角; G为地层中的Ulmus pumila; H为地层中的针叶树茎杆, 灰白相间比例尺, 分别代表1 cm; Ⅰ 为针叶树茎杆断面, 年轮均匀清晰)Fig.4 Photos of Palaeoloxodon-Coelodonta fauna in Shijiazhuang area(Ⅱ ) |
石家庄地区的古菱齿象-披毛犀动物群含有古菱齿象、披毛犀、原始牛、王氏水牛和四不像鹿等化石。由于这些化石是晚更新世的常见分子, 且化石层上部约30cm处的细砂层中的针叶树茎杆的14C年龄为8150± 120 a前, 推测哺乳类化石层的时限约在10000 a前即晚更新世晚期。
虽然古菱齿象和水牛共生, 被认为是间冰期暖湿多树林环境的动物(薛祥煦, 1999; 同号文, 2007), 但是古菱齿象也和披毛犀、原始牛共生, 发现于晚更新世晚期的陕西关中(薛祥煦, 1999)、河北迁安爪村(裴文中和黄万坡, 1958; 河北省地质矿产局, 1989; 许清海等, 2002)等地, 被认为是干冷草原环境动物(周本雄, 1978; 许清海等, 2002)。总之, 古菱齿象可能是一种生态适应比较广的哺乳类, 由于不与冻土苔原的猛犸象和湿热环境的剑齿象共生, 应该生活在温凉的半干旱半湿润环境。动物成员中的原始牛是中国华北地区乃至整个欧亚大陆晚更新世最常见的动物之一, 发现于安徽蒙城县(谢万明, 1988)、山西大同(周明镇, 1953)等地, 与其他大型动物一样生活在晚更新世暖热的乔木草地环境(Tsoukala, 1992)。水牛曾在更新世中国中东部地区多处发现, 在北方地区发现了大约 27 处水牛化石及亚化石地点(薛祥煦和李晓晨, 2000), 由于在黄土堆积中没有发现水牛化石, 所以认为水牛是一种生活于温暖、湿润环境下的喜水动物。四不像鹿生活在温暖湿润而有沼泽的平原环境, 喜欢采食鲜嫩的水生植物。四不像鹿、原始牛和水牛都具有大型的角, 不适合在密闭的森林中生活, 应该是在开阔的稀树草原环境中生活; 古菱齿象-披毛犀动物群产于河流冲积层:下部为10cm厚河卵石层, 上部为20cm厚细砂, 说明当时气候湿润。古菱齿象牙釉质标本碳同位素均值为δ 13CPDB为-5.375‰ , 显示其以C3/C4混合植物为食, C3植物稍占优势; 氧同位素均值δ 13OPDB为-7.77‰ , 对应冷湿气候。综合上面的分析, 石家庄地区的古菱齿象-披毛犀动物群生活在温凉湿润的针叶林草原环境。
石家庄的古菱齿象-披毛犀动物群为典型的古菱齿象-披毛犀动物群, 可以和相同化石组合的河北迁安爪村(裴文中和黄万坡, 1958; 河北省地质矿产局, 1989)、陕西关中(薛祥煦, 1999)、北京地区(黄万波, 1979, 1990; 尤玉柱, 1981; 李超荣等, 2000)、河北玉田县孟家泉(河北省文物研究所, 1991)、河南许昌灵井(李占扬和董为, 2007)、内蒙古萨拉乌苏(祁国琴, 1975)、新蔡(裴文中, 1956)等地的动物群进行对比, 这些动物群均生活在晚更新世晚期。
在30~22 ka BP期间, 中国北方从渭河谷地到河北平原至东北平原的某些地段曾分布有繁茂的云杉、冷杉、松林、疏林草原和草原(孔昭宸和杜乃秋, 1980; 周昆叔, 1984a, 1984b), 并伴有湖泊扩张和水位上升(薛滨等, 2001)。在22~11 ka BP期间, 气候非常寒冷干燥, 草原已推进至现代常绿阔叶林区的北部(中国第四纪孢粉数据库小组, 2000), 渤海和黄海已经退出成为陆地, 东海下降120~130 m(汪品先, 1990; Wang and Sun, 1994)。部分古菱齿象向东南迁移至台湾澎湖列岛(祁国琴和何传坤, 1999)一带度过末次盛冰期, 另一部分古菱齿象则在末次盛冰期中灭绝(Norton et al., 2010)。由于海平面下降, 石家庄地区受到强烈冲刷剥蚀, 沉积了晚更新世砾石层(
致谢 牙釉质碳氧稳定同位素由中国科学院地质与地球物理研究所稳定同位素地球化学实验室测试完成, 14C测年由中国地震局地质研究所地震动力学国家重点实验室完成, 参加野外工作的还有石家庄公交总公司刘秋军先生。作者在此深表谢意。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|