










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
塔里木盆地苏盖特布拉克地区下寒武统肖尔布拉克组碳酸盐岩微生物建造特征*
[宋金民1  , 罗平
, 罗平1 , 杨式升1 , 翟秀芬1 , 周刚2 , 陆朋朋3 ]
, 罗平|
|

第一作者简介 宋金民,男,1983年生,2006年毕业于中国石油大学(华东),现为中国石油勘探开发研究院博士研究生,从事碳酸盐岩沉积和储集层研究工作。电话:010-83592185;E-mail:songjinmin@sohu.com。
塔里木盆地苏盖特布拉克地区下寒武统肖尔布拉克组发育 3种类型碳酸盐岩微生物(蓝细菌)建造,即下部微生物丘状和层状建造、中上部微生物礁和顶部叠层石建造。微生物丘状和层状建造的特点是似层状孔洞和纹层结构发育,形成于潮下高能带,与微生物席粘结有关,分布相对稳定。微生物礁是由枝状或丛状微生物骨架生长和微生物席粘结两种方式形成,分布较广,可分为两大期:第 1大期发育在海退背景下,呈宏观块状建造特征;第 2大期则发育在较大的海侵背景下,呈现两期点礁特征。叠层石建造发育在更大范围的海侵背景下,超覆于点状建造之上,要求的水动力条件相对较强,由微生物粘结作用形成,其中叠层石和核形石是其重要标志,仅分布在研究区北部的苏 Ⅱ、苏 Ⅲ和苏 Ⅳ剖面近顶部。
About the first author Song Jinmin,born in 1983,graduated from China University of Petroleum(East China)in 2006.Now he is a Ph.D.candidate and is engaged in researches of carbonate sedimentology and reservoir.Phone: 010-83592185;E-mail: songjinmin@sohu.com.
There are three kinds of carbonate rock microbial(cyanobacteria)constructions in the Lower Cambrian Xiaoerblak Formation in Sugaitblak area,Tarim Basin,including microbial mounds and laminated constructions at the lower part,microbial reefs in the middle and upper part and stromatolite constructions at the top.The microbial mounds or laminated constructions are characterized by the stromatactoid cavities and laminated texture formed in the subtidal high energy zone.The constructions with a wide and stable occurrence were related with sticky microbial mats.The microbial dendritic or shrub structure and its cohesive action are developed in reefs,formed by microbial growth and sticky microbial mats.They are widely spread in the study area.There are two stages of microbial reefs.At the first stage macroscopic massive buildups are developed during the regression setting,while at the second stage,two stages of point reefs are developed during the transgression setting.The stromatolite constructions are formed during a more broad transgression setting,overlapped on the point microbial reefs,when the hydrodynamic force was stronger.This type of microbial buildups is formed by microbial bonding and characterized by stromatolite and oncolite.They are only developed at the top of Su Ⅱ,Su Ⅲ and Su IV sections.
20世纪50年代, Twenhofel(1950)提出了碳酸盐岩微生物建造作用, 强调微生物活动对碳酸盐岩建造形成与发育的控制作用。在宏观造礁生物种类减少之后(Pickard, 1996; Riding, 2002), 如泥盆纪F/F界线(金善燏等, 1996; Pickard, 1996)和P/T界线(金善燏等, 1996; 童金南等, 1997; 王永标等, 2005; 吴亚生等, 2006, 2007; 杨浩等, 2006; 刘建波等, 2007; 罗茂等, 2007; 何磊等, 2010), 往往是微生物建造的发育时期。寒武纪是海生无脊椎动物开始繁盛的时代, 微生物建造普遍发育, 主要为蓝细菌形成的地貌隆起, 包括灰泥丘和生物礁, 主要结构类型为叠层石、凝块石、枝状石和均匀石(Copper, 1974; Heckel, 1974; Pratt, 1982, 1995; 李书舜和刘大成, 1986; Riding, 1991, 2000, 2002; Webb, 1996; 高建平和朱士兴, 1998; Scholl and Ulmer-Scholle, 2003; Flü gel, 2004; 梅冥相, 2007a)。
在中国, 寒武纪微生物主要以蓝细菌为主, 还有古杯海绵类, 蓝细菌形态类型有葛万菌(Girvanella)、附枝菌(Epiphyton)和肾形菌(Renalcis), 其形成的微生物建造经前人研究已经取得了很多成果。总结起来, 扬子地台微生物建造多见于中下寒武统(张俊明和袁克兴, 1994; 杨友运和叶俭, 1996; 叶俭等, 1996; 郑荣才, 1996; 张廷山等, 2005; 党皓文等, 2009; Hicks and Rowland, 2009), 华北地台微生物建造则发育于中上寒武统(巩恩普等, 1995; 彭阳和乔秀夫, 1995; 高建平和朱士兴, 1998; 穆西南等, 2003; Woo et al., 2008; 韩作振等, 2009; Lee et al., 2010; Woo and Chough, 2010)。在塔里木盆地也有上寒武统微生物建造的报道, 但仅限于地球物理资料(丁勇等, 2006)。
近期, 在塔里木盆地西北缘的苏盖特布拉克露头区下寒武统肖尔布拉克组发现了微生物建造。笔者以该露头区肖尔布拉克组微生物建造为研究对象, 在剖面实测和薄片鉴定的基础上, 揭示3种不同类型微生物建造的特征与演化, 厘清不同类型微生物建造在研究区的分布特征和展布规模, 为塔里木盆地寒武系盐下礁滩白云岩油气勘探提供依据。
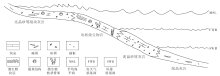
苏盖特布拉克位于新疆阿克苏地区乌什县境内, 东北距阿克苏市约80, km, 构造分区属于柯坪断隆东段。该区主要出露震旦系奇格布拉克组至中寒武统阿瓦塔格组, 地层出露良好, 剖面完整。实测的5条剖面自南向北依次为:苏0部面、苏南剖面、苏Ⅱ 剖面、苏Ⅲ 剖面和苏Ⅳ 剖面(图1)。
 | 图1 塔里木盆地苏盖特布拉克露头区剖面位置Fig.1 Location of studied sections in Sugaitblak area, Tarim Basin |
震旦系奇格布拉克组白云岩与上覆下寒武统玉尔吐斯组呈似平行不整合接触, 不整合面之上发育灰黑色硅质岩和灰色磷块岩。下寒武统从下到上依次为玉尔吐斯组、肖尔布拉克组和吾松格尔组, 中寒武统包括沙依里克组和阿瓦塔格组(表 1)。
| 表1 塔里木盆地苏盖特布拉克露头区寒武系地层系统 Table1 Stratigraphic framework of the Cambrian in Sugaitblak area, Tarim Basin |
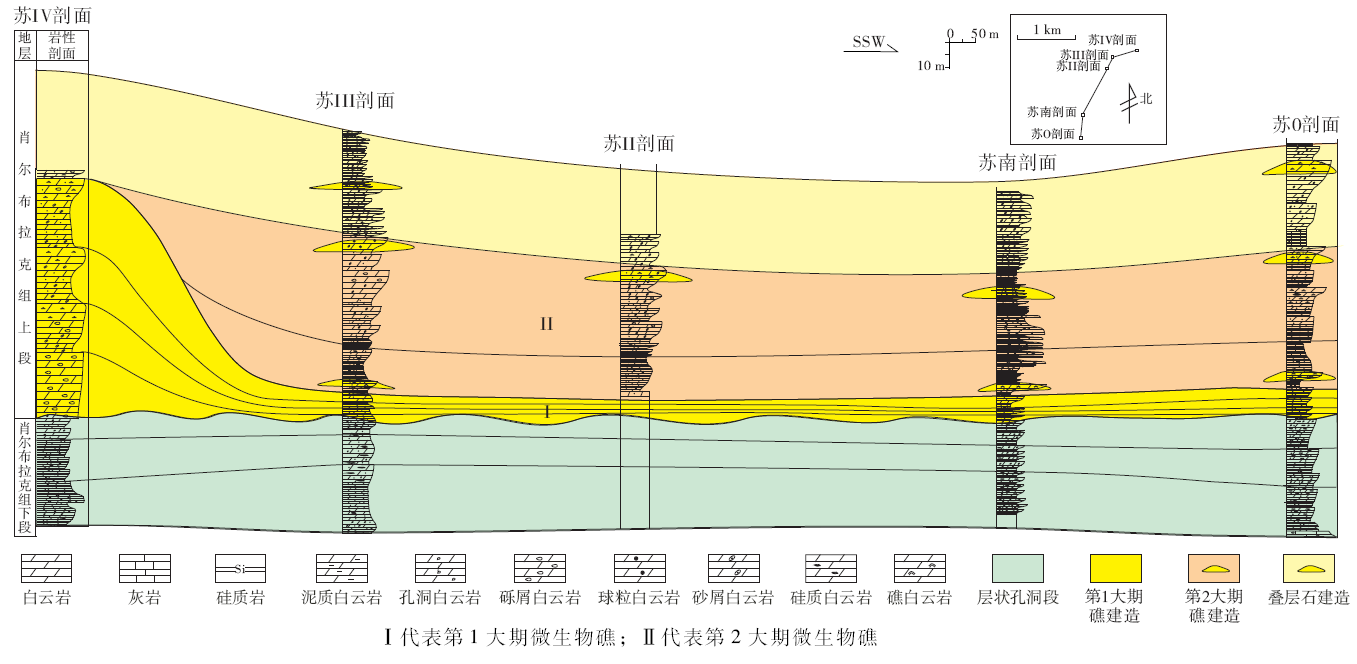
下寒武统肖尔布拉克组的微生物(蓝细菌)碳酸盐岩建造厚150.7, m, 可分为上、下“ 黑白” 两段(图 2), 两段之间呈假整合接触关系, 有1个小型沉积间断, 界面凹凸不平。下段岩性为深灰色纹层— 薄层状残余球粒粉晶白云岩, 发育硅质结核, 形成层状和丘状建造特征, 在研究区分布稳定。上段为微生物礁发育段, 南北沉积结构明显不同:研究区北部的剖面(苏Ⅳ 剖面)为第1大期微生物礁, 岩性主要为残余表附藻粉晶白云岩、残余凝块粉晶白云岩, 局部见有疑似古杯类; 在同时期的南部(苏0剖面)则相变为浅灰色极薄层— 纹层状残余球粒泥晶灰岩, 并见有2~3, cm长的直角石化石; 而第2大期微生物礁则主要发育在南部, 以多个点礁形式呈不规则块状展布, 在其上部发育叠层石建造。顶部为1层厚20, cm、横向分布稳定的深灰色硅质白云岩。
 | 图2 塔里木盆地苏盖特布拉克地区下寒武统肖尔布拉克组综合柱状图Fig.2 Comprehensive stratigraphic column of the Lower Cam-brian Xiaoerblak Formation in Sugaitblak area, Tarim Basin |
塔里木盆地早寒武世早期发生大规模海侵, 沉积了一套大范围分布的黑色页岩、含磷硅质岩或磷块岩, 随后发生持续性海退①(①张宝民, 李洪辉, 刘伟, 等.2008.塔里木盆地中部寒武系— 奥陶系碳酸盐岩成藏地质特征及勘探领域.中国石油勘探开发研究院内部报告.)。研究区属于碳酸盐岩台地相, 早寒武世早期为远离陆地、水体较深的台地相, 早寒武世中晚期则演变为浅水碳酸盐岩台地相(冯增昭等, 2002, 2005, 2006)。通过对野外沉积特征的总结和室内的薄片鉴定, 笔者认为苏盖特布拉克地区早寒武世处于潮下带沉积环境, 为潮下高能带与潮下低能带交替变化, 共发育4个潮下高能带— 潮下低能带的沉积旋回。微生物建造多发育在潮下高能带, 而潮下低能带则为深灰色、灰黑色泥晶白云岩和泥晶灰岩沉积。
在剖面实测和岩石薄片鉴定基础上, 认为苏盖特布拉克露头区下寒武统肖尔布拉克组发育3种类型的微生物建造:微生物丘状和层状建造, 高突起微生物礁和浅水高能叠层石建造。
主要发育于肖尔布拉克组下段, 宏观上呈现出连续性好的黑色块状体(图 3-1), 形成于潮下高能带。在研究区主要出露在苏0、苏Ⅲ 和苏Ⅳ 剖面, 分布相对稳定。这种微生物建造的主要特点为似层状孔洞构造和纹层构造发育。
 | 图3 塔里木盆地苏盖特布拉克地区下寒武统肖尔布拉克组潮下高能带微生物层状建造宏观结构及地层柱状图Fig.3 Laminated microbial construction and stratigraphic column in subtidal high energy zone of the Lower Cambrian Xiaoerblak Formation in Sugaitblak area, Tarim Basin |
3.1.1 似层状孔洞构造
似层状孔洞构造发育是此类微生物造的典型标志。在苏0剖面上, 似层状孔洞主要发育在5-2、5-3小层, 在5-4小层仅局部发育(图3-2), 岩石类型为深灰色纹层状残余球粒泥粉晶白云岩。似层状孔洞构造的孔洞、晶洞顺层分布, 孔洞总量为5%~15%, 局部密集发育, 可达40%。孔洞呈扁平状、椭圆状或不规则状, 大小一般为3 cm× 4 cm, 大的达4 cm× 10 cm, 孔洞内被亮晶白云石充填。野外剖面上, 似层状孔洞呈扁平状, 略平行于层面分布(图 4-1), 被微生物纹层包绕, 幅度在3~4, cm(图4-2), 孔洞大小一般为6, cm× 3, cm, 中粗晶白云石充填, 多发育在微生物浅色纹层内。
 | 图4 塔里木盆地苏盖特布拉克地区层状微生物建造中的似层状孔洞构造Fig.4 Photos showing stromatactoid cavities of laminated constructions in Sugaitblak area, Tarim Basin |
在偏光显微镜下观察, 这种类型的白云岩纹层结构发育, 纹层由深灰— 浅灰色互层的球粒白云岩组成, 浅色纹层晶粒较大, 为粉晶白云石, 深色纹层则为泥晶白云石, 纹层内可见灰黑色毫米级的暗色泥晶球粒结构, 深色纹层与浅色纹层之间的界线呈凹凸不平的特征(图 4-3, 4-4)。似层状孔洞呈斑块状分布, 多发育在微生物浅色纹层内, 大小一般为2 cm× 3 cm, 亮晶白云石充填, 孔洞与浅色纹层之间界线为高低起伏的微剥蚀面(图 4-3, 黄色箭头所指), 孔洞内中粗晶白云石与浅色纹层的泥粉晶白云石之间呈突变接触关系。
关于似层状孔洞结构的成因, 目前尚存在争议。Pickard(1996)认为层状孔洞是微生物席降解和腐烂产生的有机酸溶解早期碳酸盐岩形成的。张宝民等(2004)在研究塔里木盆地肖尔布拉克剖面的层状孔洞构造时, 认为其为宏观藻类的生殖器官化石。在研究区这种纹层结构和似层状孔洞结构中, 纹层内的泥晶球粒和纹层— 孔洞之间凹凸不平的突变接触特征与Pickard(1996)研究的欧洲早石炭世狄南统Waulsortian建造内的微生物结构相似, 且浅色纹层与孔洞相邻的边缘呈暗色发散状, 而非溶蚀残余状, 表明这可能是由微生物稳定作用产生的构造(梅冥相等, 2007b)。故笔者认为这种纹层— 似层状孔洞结构与微生物作用有关。
3.1.2 纹层构造
苏0剖面5-4小层上部波状纹层结构发育, 似层状孔洞仅局部发育。这种纹层结构在野外剖面上主要表现为低缓的波状, 横向连续性较好, 单个纹层厚约2~4, cm, 波动幅度为3~10, cm, 波长大约为80~90 cm。如图5-1所示, 波状纹层的隆起发育在微生物丘状建造之上, 每个丘状建造的隆起幅度也略有差异, 有的隆起幅度较大, 为8~10, cm, 有的则较为平缓, 为3~4 cm。微生物丘状建造内部纹层不发育, 而微生物丘状建造体的上部和侧翼则为波状纹层(图 5-2)。
 | 图5 塔里木盆地苏盖特布拉克地区微生物建造作用形成的纹层构造Fig.5 Photos showing laminated structure of laminated microbial constructions in Sugaitblak area, Tarim Basin |
从镜下显微特征来看, 纹层由亮暗相间结构, 根据暗色层的产状和分布特征, 主要有两种类型:第1种为纹层波状起伏, 由厚度相当的暗层和碳酸盐岩微亮晶纹层形成相间结构, 纹层厚度为2~4, mm, 暗层为泥粉晶白云岩, 边缘凹凸不平, 为一微剥蚀面, 有机质充填, 显微镜反光下呈暗色, 亮层则为亮白色粉细晶白云岩(图 5-3), 微剥蚀面与亮晶白云石呈突变接触, 且伸向亮晶白云石部分呈发散状, 这与压溶缝聚敛收缩的特征明显不同, 因此可以排除压溶作用的影响。纹层呈幅度较大的波状起伏(图 5-3), 幅度为1~2, mm。第2种为纹层较平直, 浅色纹层厚度大, 暗层厚度小、断续分布于浅色纹层中, 仅在浅色纹层边缘为连续的波状微细暗层(图 5-4)。暗色层显微镜反光特征为暗色, 说明其为有机质; 纹层界线处通常颜色更深, 连续性变好, 略呈微波状起伏。浅色纹层厚度为2~3, mm, 暗色纹层厚度为0.5~1, mm。
纹层结构的形成与微生物的间歇性生长活动密切相关, 主要与以下两种过程相关:微生物胞外聚合物(EPS:extracellular polymeric substance)促进碳酸盐沉淀和微生物捕获、固结颗粒沉积物(Riding, 2000)。这是因为微生物既能营自养生活又能营异氧生活, 其分泌胞外聚合物, 粘合沉积物形成一个粘性的表面席。在有碳酸盐沉积物供给时, 捕获碳酸盐颗粒形成亮层; 在没有沉积物供给时, 微生物胞外聚合物形成暗层。纹层结构边缘通常呈波状起伏的特征(图 5-3, 5-4), 说明粘性微生物席相对较软, 且表面存在不规则的微地貌, 呈现出微剥蚀面的特征。
笔者建立了纹层状微生物丘状和层状建造模式(图 6)。在此背景下, 主要发育纹层状泥晶球粒灰岩沉积, 微生物主要形成层状和丘状微生物席, 通过胞外聚合物生长和粘结捕获作用, 形成碳酸盐岩建造。在埋藏过程中, 微生物席降解产生有机酸, 溶解早期灰岩沉积物, 形成似层状孔洞。微生物丘状建造主要发育在微生物席的局部高部位, 有些丘状建造发育结束后, 在其上发育波状纹层; 有些建造体则呈继承性发育。似层状孔洞与丘状建造共生, 其形成与微生物活动密切相关, 发育在纹层相对较厚的层段, 具有顺层分布的特点。
 | 图6 塔里木盆地苏盖特布拉克地区深水纹层状微生物丘状和层状建造模式Fig.6 Depositional model of microbial mounds and laminated constructions in deep water in Sugaitblak area, Tarim Basin |
肖尔布拉克组上段在研究区北部表现为一种具有高突起的生物礁, 代表剖面为苏Ⅳ 剖面。该剖面位于研究区东北部, 微生物礁主要发育在6~7层, 总厚度67, m, 横向延伸3 km, 是研究区微生物礁最高、规模最大的部位(图 7)。
 | 图7 塔里木盆地苏盖特布拉克地区苏Ⅳ 剖面微生物礁及地层柱状图Fig.7 Microbial reef and stratigraphic column in Su Ⅳ section in Sugaitblak area, Tarim Basin |
在野外剖面上, 肖尔布拉克组上段微生物礁共发育两大期, 北部苏Ⅳ 剖面只见有第1大期微生物礁, 其由北向南前积, 第2大期礁则不发育。第2大期微生物礁主要分布在南部, 呈现出由南向北超覆的特征。综合两大期微生物礁的沉积类型, 发现其发育4种岩石类型(图8):(1)深灰色微晶灰岩; (2)泥晶砂屑灰岩; (3)块状微生物岩; (4)亮晶砂屑凝块灰岩(蓝细菌凝块岩)。微晶灰岩一般发育在风暴浪基面以下, 泥晶砂屑灰岩则发育在正常浪基面和风暴浪基面之间, 块状微生物岩发育在正常浪基面之上, 而亮晶砂屑凝块灰岩则沉积在平均海平面以下的浅水区。在苏盖特布拉克地区, 北部的苏Ⅳ 剖面主要发育岩性组合②-③-④; 而在南部4种岩性全部发育。但微生物建造主要是指岩性组合③-④, 即块状微生物岩和蓝细菌凝块岩。
 | 图8 塔里木盆地苏盖特布拉克地区微生物礁沉积模式Fig.8 Depositional model of microbial reef in Sugaitblak area, Tarim Basin |
北部苏Ⅳ 剖面第1大期微生物礁由4个成礁旋回组成, 每个旋回厚约10~20, m。旋回结构相似, 下部为残余砂屑或残余球粒白云岩, 为颗粒滩沉积; 上部为残余表附藻或残余凝块白云岩。第2大期礁在苏南剖面第6层和苏0剖面第7层可见枝状的微生物骨架, 岩性为残余凝块白云岩, 厚度约2, m。暗色斑块为微生物蓝细菌叶状体结构, 大小为1 cm× 2 cm; 叶状体之间的浅色结构为蓝细菌结构之间的孔洞, 为晶粒白云石充填。纵向上下部的斑块较大, 向上斑块变小。而在北部, 则对应为从南部超覆的亮晶凝块岩, 位于剖面的顶部。
从镜下显微特征来看, 苏Ⅳ 剖面的微生物礁体中发育两种类型的蓝细菌结构:原地生长的附枝菌(Epiphyton)和凝块结构。
附枝菌为一类钙质微生物, 属蓝细菌生物, 主要发育于寒武纪到泥盆纪, 尤以寒武纪最盛。前人在描述附枝菌的产状时, 总结出了两种生长形态:树枝状和簇球状(张俊明等, 1985)。在研究区附枝菌的生长形态目前仅见树枝状, 主要发育于苏Ⅳ 剖面的第3个旋回, 白云石化作用改造后呈泥粉晶结构, 宏观上呈块体出现, 厚度在5~8, m之间, 横向连续较好(图 9-1)。在偏光显微镜下为具分叉的叶状体, 单个叶状体长0.3~1.5, mm, 宽0.05~0.1, mm, 颜色暗, 内部不显示微细结构, 由隐晶白云石组成, 叶状体之间的空隙被亮晶泥粉晶白云石充填(图 9-2, 红色箭头); 叶状体常群集出现, 呈树枝状向上、向外分叉丛生, 形似小灌木丛, 附枝菌丛体长度为3~5, mm, 宽度的变化呈喇叭状, 底部宽度约1, mm, 上部最宽的部分为5, mm, 丛体与丛体之间可见大的孔洞(图 9-2, Vug), 亮晶粉晶— 粗晶白云石充填。这种产出形态与前人报道的华北地台东部中上寒武统的附枝菌(张俊明等, 1985; Riding, 2002; 韩作振等, 2009; Lee et al., 2010; Woo et al., 2010)和俄罗斯西伯利亚台地早寒武世的附枝菌(Scholle and Ulmer-Scholle, 2003)树枝状结构很相似。
 | 图9 塔里木盆地苏盖特布拉克地区微生物礁和苏Ⅲ 剖面叠层石建造Fig.9 Photos showing microbial reef and stromatolite constructions at Su Ⅲ section in Sugaitblak area, Tarim Basin |
研究区第2种蓝细菌结构为凝块结构, 前人进一步将其分为钙化微生物凝块石、粗粒粘聚状凝块石、树枝状凝块石、钙华凝块石等(Riding, 2000)。在中国, 上扬子区依迪卡拉系灯影组白云岩和奥陶系红花园组灰岩为凝块石发育的典型代表(梅冥相, 2007)。在研究区, 凝块结构呈斑块状、暗色、内部为泥晶球粒结构, 原岩主要为蓝细菌凝块灰岩, 主要见于苏Ⅳ 剖面的第2个旋回和第4个旋回, 多发育在树枝状附枝菌结构之上。偏光显微镜下, 凝块大小为(1~3)mm× (0.5~4)mm, 含量为40%~50%, 边缘呈不规则起伏状, 向内收敛, 产状不规则, 有时是几个呈团簇状产出, 有时是单个出现, 凝块的周围为正常泥粉晶沉积物, 但其内部常见不规则状的孔洞, 为亮晶粉晶白云石充填(图 9-3)。单个凝块和团簇状凝块(图 9-3)均具有发散的边缘, 这可能与微生物胞外聚合物作用有关。蓝细菌凝块结构的成因目前仍有争议, 一些学者(Kennard and James, 1986; Pentecost and Riding, 1986; Riding, 1991, 2000)认为凝块石是由球菌钙化形成的, 但也有学者(Riding, 2000)认为是微生物粘结作用形成的。
叠层石建造作用主要发育苏Ⅲ 剖面顶部第2层的上部(图 10), 位于研究区的北部, 发育在下寒武统肖尔布拉克组上段微生物建造的近顶部, 向南部深水区(苏0、苏南剖面)相变为潮下高能滩沉积。这种叠层石建造作用在苏Ⅱ 剖面、苏Ⅲ 剖面及苏Ⅳ 剖面上都发育, 宏观上主要呈厚层— 块状产出, 总厚度为0.8~1, m, 横向连续性较好。沉积背景为肖尔布拉克组沉积晚期的大范围海侵。沉积位置相当于图8岩性组合④的分布区域。与其他两类建造不同的是, 叠层石建造的水体相对较浅, 微生物结构的抗浪性增强, 叠层石白云岩、核形石结构发育。
 | 图10 塔里木盆地苏盖特布拉克地区苏Ⅲ 剖面叠层石建造的发育位置及宏观展布特征Fig.10 Macroscopic occurrence and stratigraphic column of stromatolitic constructions at Su Ⅲ section in Sugaitblak area, Tarim Basin |
在苏Ⅲ 剖面上, 这种叠层石微生物建造主要呈丘状、微波状或平行纹层状产出, 每个纹层厚约2~3, mm, 相互叠置, 形成叠层结构。由下到上, 微生物叠层的厚度逐渐增加, 下部叠层体总厚度约10, cm, 至中部厚度增加至20~30, cm, 上部厚度为40~50, cm, 呈现出下细上粗的沉积韵律结构(图 9-4)。岩性为褐灰色中厚层状叠层石泥粉晶白云岩。
在偏光显微镜下, 叠层石建造的纹层厚度在0.5~1.5, mm之间, 呈亮暗相间的结构, 暗层为深灰色泥晶白云岩, 亮层则为浅灰色粉晶白云岩, 互相叠置呈丘状叠层结构; 纹层呈波状(图 9-5, 箭头所指), 起伏的幅度大小不等, 大的为1, mm, 小的为0.2~0.3, mm, 亮层内见到孔洞, 可能为鸟眼孔, 亮晶白云石充填或半充填(图 9-5, Vug), 局部可见有机质充填的粒间隙(图 9-5, 红色箭头所指)。叠层石结构是由微生物层纹与微生物捕获的沉积物而成的层纹交替而成。波状起伏的纹层表明微生物席表面不规则的微地貌及其微生物活动的不均一性, 指示着其微生物成因。
核形石白云岩也是该类微生物建造的重要标志。核形石是由微生物生长形成的结核状构造, 一般发育于周期性波浪或潮汐作用的地区, 多形成于浅水沉积环境(Scholle and Ulmer-Scholle, 2003)。在研究区苏Ⅲ 剖面, 核形石结构位于叠层结构的上部, 核形石层厚约20~40, cm, 横向连续性好。核形石的个体大小不一, 大的为(5~8)mm× (4~6)mm, 小的为2 mm× 3 mm, 形态多样, 呈圆形或次圆形、椭圆形, 核心部分大小为(0.1~0.3) mm× (0.1~0.2)mm, 各个核形石排列紧密, 密度为8~10个/cm2, 与前人报道的云南震旦系王家湾组核形石的特征相似(曹仁关等, 1991)。在偏光显微镜下, 见到的最大的核形石为8 mm× 6 mm(图 9-6), 其核心为砂屑、砾屑, 大小为(0.5~2)mm× (0.4~1)mm, 核心外部为亮暗相间的纹层, 共发育10层纹层, 亮层为泥粉晶白云石, 层厚较大, 在0.1~0.4, mm之间, 暗层成分为泥晶白云石, 层厚较小, 为0.1~0.2, mm, 第1~2层暗纹层厚度更小, 为0.03~0.05, mm。纹层直径由内到外不断增大, 内部纹层直径为3, mm, 外部纹层可达6~8, mm, 由于重结晶作用, 外部局部蓝细菌纹层结构不甚清晰。
这种类型的微生物建造形成于水动力相对动荡的水体中, 10层蓝细菌纹层直径不断增大的现象也说明核形石在形成过程中, 水动力逐渐增强, 水体扰动和振荡强度不断变大, 微生物生长圈层也随之增大, 微生物的抗浪性也逐渐增强。
肖尔布拉克组下段为深灰色微生物纹层状球粒白云岩段, 以发育似层状孔洞和波状纹层为特征, 微生物通过形成粘性的微生物席粘结细粒沉积物, 形成丘状和层状建造, 共划分出3个亚段:第1亚段微生物波状纹层发育, 第2亚段似层状孔洞构造发育, 第3亚段发育小型丘状层纹构造, 并含有似层状孔洞构造。肖尔布拉克组上段微生物礁共发育两期, 第1大期微生物礁是在海退背景下形成, 生物礁规模较大, 以微生物骨架形式成礁, 主要发育在东北部地形高部位, 呈现出4个由北向南前积的结构; 而此时, 对应南部深水区的苏0剖面则为微粒泥质灰岩和微晶灰岩。第2大期微生物礁主要发育在南部, 规模相对较小, 蓝细菌凝块和骨架均发育, 但以凝块为主, 表现为3期向北部第1大期礁体上超的结构, 前2期呈现出点礁的特征, 沉积旋回以泥晶灰岩开始, 以变浅的微生物点礁复合体结束, 第3期上超规模更大, 微生物通过粘结作用形成叠层石和核形石建造。肖尔布拉克组上段微生物礁的沉积模式如图11所示。将5条剖面中相对稳定的肖尔布拉克组下段拉平对齐, 上段微生物礁按照其发育期次和沉积结构的变化进行对比, 肖尔布拉克组的微生物建造横向展布特征如图12所示。因此, 下寒武统肖尔布拉克组微生物建造从下而上呈现出丘状、层状建造→ 微生物礁→ 叠层石、核形石建造的演化规律。
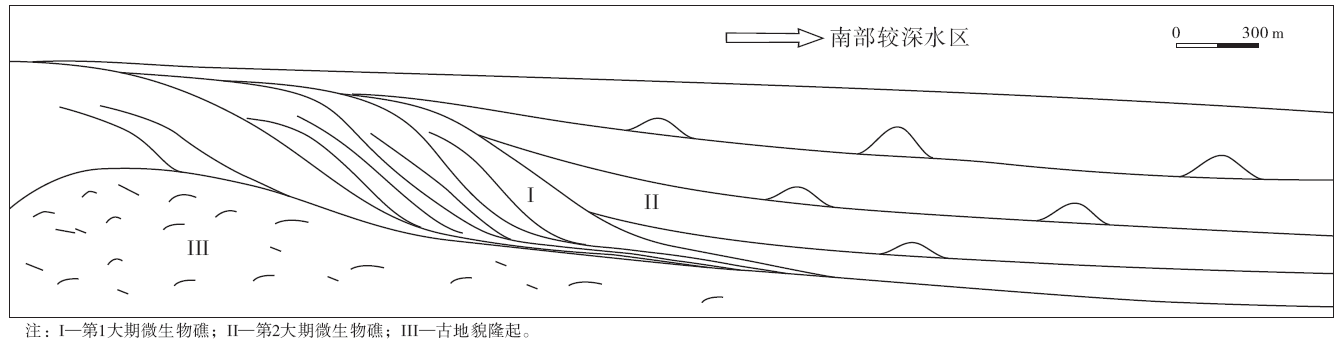 | 图11 塔里木盆地苏盖特布拉克地区肖尔布拉克组上段微生物礁沉积模式Fig.11 Depositional model of microbial reefs in the upper member of Xiaoerblak Formation in Sugaitblak area, Tarim Basin |
肖尔布拉克组3种不同类型的微生物建造在研究区各剖面上的发育和展布特征也有所不同。微生物丘状和层状建造主要发育于肖尔布拉克组下段, 分布较为稳定, 厚度为30~40, m, 分布在苏0、苏南、苏Ⅲ 和苏Ⅳ 剖面下部; 微生物礁主要发育于肖尔布拉克组中上部, 分布范围较广, 在5条剖面均发育, 厚度为40~60, m, 构成了研究区微生物建造的主体; 而浅水高能叠层石建造则发育在肖尔布拉克组顶部, 厚度为20~40, cm, 规模相对较小, 仅分布在苏Ⅱ 剖面、苏Ⅲ 剖面和苏Ⅳ 剖面。
1)肖尔布拉克组发育3种类型微生物建造, 即下部微生物丘状和层状建造、中上部微生物礁隆和顶部叠层石建造。微生物丘状和层状建造特点是层状孔洞和纹层结构发育, 其形成与粘性的微生物席有关; 微生物礁是由丛状微生物通过自身生长和微生物席粘结两种方式形成, 第1大期微生物礁形成于海退背景下, 分布在东北部的古地貌高部位, 为宏观块状建造体; 第2大期微生物礁则形成于海侵背景下, 主要分布在南部, 呈现点礁特征。叠层石建造则为更大范围海侵的产物, 超覆于微生物点礁之上, 分布在研究区北部高地貌处, 水动力条件相对较强, 由微生物席粘结作用形成, 叠层石和核形石结构发育。
2)微生物建造在横向上发育特征有所不同。微生物丘状和层状建造主要发育于肖尔布拉克组下段, 分布范围较大, 在苏0、苏南、苏Ⅲ 和苏Ⅳ 剖面都发育; 微生物礁主要发育于肖尔布拉克组中上部, 发育范围较广; 而叠层石微生物建造则发育在肖尔布拉克组顶部, 仅分布在北部的苏Ⅱ 、苏Ⅲ 和苏Ⅳ 剖面的近顶部, 向南相变为潮下高能滩沉积。
致谢 在文章的审阅和修改过程中,中国石油大学(北京)冯增昭教授和两位评审专家给予了中肯的指导和帮助,成都理工大学杨迪同志也提出了很多宝贵意见。野外剖面实测和样品采集工作得到了饶红娟、王鑫等同学的协助,在此一并表示衷心感谢!
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|