


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
山东平邑盆地古近系湖相微生物碳酸盐岩形成机理*
[唐鑫萍1, 2  , 黄文辉
, 黄文辉1, 2 , 邓宏文1, 2 , 王文勇1, 2 , 穆娜娜1, 2 ]
, 黄文辉, 邓宏文|
|

第一作者简介 唐鑫萍,男,1987年生,中国地质大学(北京)矿产普查与勘探专业硕士研究生,研究方向为应用沉积学。E-mail:cugbtxp@126.com。
通讯作者简介 黄文辉,男,1961年生,理学博士,中国地质大学(北京)教授,博士生导师,主要从事沉积学教学与研究。E-mail:huangwh@cugb.edu.cn。
微生物碳酸盐岩是当前沉积学的研究热点之一,但关于古近系湖相微生物碳酸盐岩的报道还非常少。为探讨山东平邑盆地古近系官庄组中段湖相微生物碳酸盐岩的形成机理,笔者进行了野外实测以及室内偏光显微镜和扫描电镜观察。研究结果认为:( 1)研究区微生物碳酸盐岩主要有核形石、叠层石和凝块石;( 2)在核形石、叠层石和凝块石内部均发现了球状微生物化石、席状体和凝块状泥晶等微生物成因的显微组构,表明了它们形成过程中共同的生长、粘结和钙化等微生物作用;( 3)微生物作用对环境变化比较敏感,水体较小、变化频繁的湖泊环境,对微生物碳酸盐岩结构和形态的发育有很大影响;核形石、叠层石和凝块石的形成环境大致呈现出水动力依次减弱、陆源碎屑物质依次减少、水深依次加大的规律;( 4)提出了研究区微生物碳酸盐岩的成因模式,认为这 3种微生物碳酸盐岩正是以微生物活动为共同基础,在不同的环境条件下形成的。
About the first author Tang Xinping,born in 1987,is a candidate for his master degree in China University of Geosciences(Beijing),and is mainly engaged in researches of sedimentology.E-mail:cugbtxp@126.com.
About the corresponding author Huang Wenhui,born in 1961,is a professor of China University of Geosciences(Beijing),and is mainly engaged in researches of sedimentology.E-mail:huangwh@cugb.edu.cn.
The origin of microbial carbonate rocks has become a hot issue of sedimentology,the Paleogene lacustrine carbonate rocks,however,are still rarely noticed and studied.In order to study the origin of the Paleogene lacustrine microbial carbonate rocks in the Pingyi Basin,field inspection as well as observations of plain microscope and scanning electron microscope,were carried out.The following results were obtained:(1)Microbial carbonate rocks in the studied section mainly consist of oncolites,thrombolites and stromatolites.(2)In each of these carbonate rocks,coccoid cells,sheet structures and clotted fabric were found,which illuminate the common microbial actions during their formation,including secretion,agglutination and calcification.(3)These 3 microbial carbonate rocks were formed in different environments,which play a great role in the development of textures and shapes of microbial carbonates,because microbial fabrics and microbial actions are sensitive to the instable lacustrine environment.(4)Based on the analyses of the common microbial actions and different forming environments,the genetic model of lacustrine microbial carbonate rocks was presented.
微生物岩(microbialite)由Burne和Moore(1987)首先提出, 指由底栖微生物群落捕获和粘结碎屑沉积物、或以它们为矿物沉淀中心而形成的生物沉积岩。微生物碳酸盐岩(microbial carbonate)则是以碳酸盐矿物为主要成分的、最常见的一种微生物岩(Riding, 2000)。近年来的研究表明, 微生物的活动与碳酸盐岩的形成有密切关系, 这极大地扩展了碳酸盐岩的成因研究(Barbara et al., 2007; Woo et al., 2008; 张杰和童金南, 2010)。由于微生物碳酸盐岩还可以成为理想的油气储集层和烃源岩(杨浩等, 2007; 史晓颖, 2008; Beatriz and Marc, 2010), 因此微生物碳酸盐岩已成为了当前沉积学的一个研究热点。目前国内外微生物碳酸盐岩的研究范围主要集中在二叠系— 三叠系、早古生界、中新元古界的海相地层(Sheehan and Harris, 2004; Riding and Liang, 2005; Riding, 2006), 对古近系湖相微生物碳酸盐岩研究还非常少(Arenas et al., 2007)。
前人对平邑盆地古近系官庄组核形石岩性特征、核心和纹层的特点开展了许多研究并对其进行了分类和沉积环境分析(管守锐和白光勇, 1985; 谢庆宾等, 1999; 李熙哲等, 2000; 唐鑫萍等, 2011)。但是, 对该区发育的核形石、叠层石和凝块石等一系列微生物碳酸盐岩, 目前还缺乏系统与综合的成因解释。文中在野外实测和偏光显微镜及电子显微镜观察的基础上, 初步分析了研究区湖相微生物碳酸盐岩的形成机理。
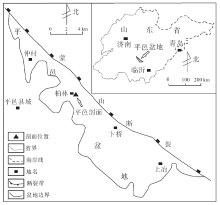
平邑剖面位于山东省平邑县柏林镇柏林村(图 1), 面积为540 km2, 是一个受蒙山断裂控制的中新生代断陷湖盆(李熙哲等, 2000)。盆地内侏罗系、白垩系和古近系发育, 其中古近系官庄组可进一步分为上、中、下3段。官庄组中段(后文简称“ 官中段” )的碳酸盐岩极为发育, 约占该段沉积地层总厚度的40%。前人根据古生物、地球化学、岩石学特征和构造背景等证实了官中段碳酸盐岩为内陆湖泊沉积(管守锐和白光勇, 1985; 谢庆宾等, 1999)。
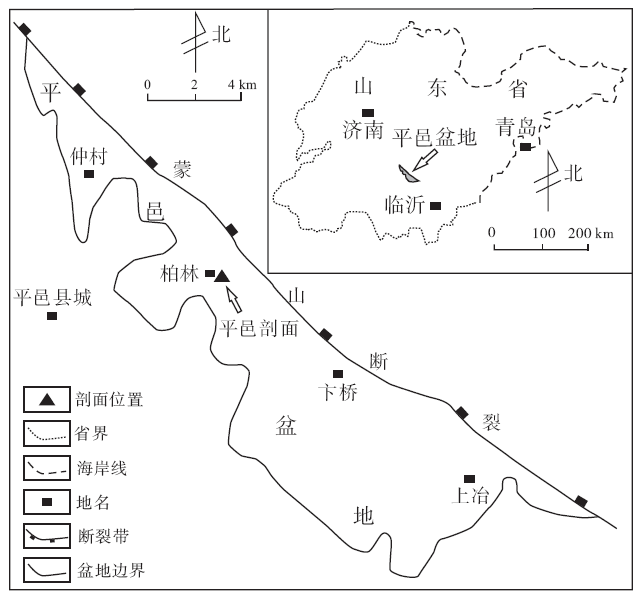 | 图1 平邑盆地及平邑剖面位置示意图Fig.1 Sketch map showing location of Pingyi Basin and Pingyi section |
文中实测的碳酸盐岩剖面厚26.03, m, 为官中段底部地层(图 2), 与下伏的官下段砾岩呈不整合接触。根据岩相的变化, 结合微生物碳酸盐岩的类型和构造特征, 将该剖面分为14层, 分别在剖面各层进行采样。所有样品均在中国地质大学(北京)海相储集层演化与油气富集机理教育部重点实验室进行测试:薄片观察采用德国Leica偏光显微镜; 各层具代表性的1 cm3左右的新鲜样品, 采用捷克TESCAN VEGA Ⅱ -LSH钨灯丝扫描电子显微镜进行观察, 电压为20 kV。
 | 图2 平邑剖面古近系官庄组中段微生物碳酸盐岩综合柱状图Fig.2 Comprehensive column of microbial carbonate rocks of the middle member of Paleogene Guanzhuang Formation at Pingyi section |
Riding(2000)按照宏观组构特征将微生物碳酸盐岩分为叠层石、凝块石、树形石和均一石4类, 这与Shapiro(2000)的中型结构分类相近。在此基础上, 梅冥相(2007a)增加了核形石和纹理石, 将微生物碳酸盐岩分为6类。平邑剖面发育的微生物碳酸盐岩主要有核形石、叠层石和凝块石3类。
核形石也常被称为“ 藻灰结核” (冯增昭等, 1994), 主要特点是由核心和纹层组成包覆颗粒结构(Flü gel, 2004)。它是平邑剖面官中段中最常见的微生物碳酸盐岩, 主要发育在剖面下部。
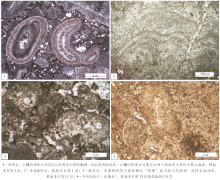
平邑剖面核形石主要为球状、椭球状, 颗粒直径为数毫米至数厘米(图 3-1, 3-2), 其核心物质主要包括生屑、砂屑和陆源碎屑, 有的发育两个或多个核心, 纹层多呈规则、连续的同心状, 一般由明暗相间的纹层交织叠覆而成。暗层主要由泥晶方解石组成, 有机质含量多; 亮层主要由微亮晶方解石组成, 有机质含量少(图 4-1)。
 | 图3 平邑剖面古近系官庄组中段微生物碳酸盐岩的野外照片Fig.3 Photos showing microbial carbonate rocks of the middle member of Paleogene Guanzhuang Formation at Pingyi section |
 | 图4 平邑剖面古近系官庄组中段微生物碳酸盐岩的显微照片Fig.4 Microscopic photos showing microbial carbonate rocks of the middle member of Paleogene Guanzhuang Formation at Pingyi section |
叠层石集中发育在剖面的顶部, 在第14层形成叠层石灰岩(图 3-3)。也有小规模的叠层石在剖面底部与核形石伴生(图 3-2)。
平邑剖面的叠层石主要为层状叠层石(包括水平状或波状), 未见柱状叠层石。叠层石的纹层可分为暗层和亮层(图 4-2), 肉眼下亮层厚3~5, mm, 暗层厚1, mm左右。显微镜下, 暗层主要由暗色泥晶方解石组成, 富含有机质; 亮层主要由微亮晶方解石组成。
凝块石是指富含有机质、 没有明显内部组构和外形的暗色泥晶碳酸盐聚合体 (Aitken, 1967; Aitken and Narbonne, 1989; Ezaki et al., 2003), 一般具有不规则的颗粒形态, 类似于前人所指的“ 凝聚颗粒” 中的“ 团块” (Tucker and Wright, 1990; 梅冥相, 2007b)。
平邑剖面官中段的凝块石肉眼观察多呈斑块状(图 3-4), 显微镜观察常呈泡沫状(图 4-3)、凝聚颗粒状, 具有明显的粘结凝聚特征。凝块直径为毫米或厘米级, 由暗色微生物泥晶和相互粘结凝聚的钙化颗粒组成; 凝块周围是后生的孔隙填充物或胶结物。这与Kennard和James(1986)描述的“ 中凝块” (mesoclots)组构相似。
通过扫描电子显微镜观察, 在平邑剖面官中段的核形石、凝块石、叠层石中均发现了球状微生物化石、席状体及凝块状泥晶等具有微生物成因特征的显微组构。
在对平邑剖面官中段样品的扫描电镜观察中, 发现了一些球状微生物化石(图 5-1, 5-3), 直径一般为5~10, μ m, 总体具有个体小、结构简单的特征, 并常与不具有晶体形态特征的、相互粘结的席状体伴生。
 | 图5 平邑剖面古近系官庄组中段微生物碳酸盐岩扫描电子显微镜照片Fig.5 Scanning electron micrographs showing microbial carbonate rocks of the middle member of Paleogene Guanzhuang Formation at Pingyi section |
研究表明, 钙化的微生物大多数为蓝细菌(cyanobactera)(Riding, 2000), 但因为大部分微生物不具钙质骨骼, 只有极少部分可以被钙化并保存为化石(Scholle and Ulmer-Scholle, 2003), 加之它们在形态和大小方面非常相似(特别是球状微生物化石), 所以对其属种的鉴定非常困难(Flü gel, 2004; 韩作振等, 2009)。文中根据小球状化石的形态、结构和大小, 认为它是一种钙化的微生物化石。
扫描电镜下所观察到的席状体为非自形泥晶结构, 其直径一般为10, μ m左右, 席状体之间多为锯齿状连接(图 5-1, 5-2), 呈相互粘结的方式出现, 常与球状微生物化石伴生(图 5-3), 覆盖在方解石颗粒的表面。其席状体的外形、非自形泥晶结构、相互连接的展布方式是它的主要识别标志。
平邑剖面官中段的席状体在结构和形态上都非常类似于Defarge等(1996)报道的现代多边形EPS(extracellular polymeric substances, 即胞外聚合物质)组织和Camoin等(1999)报道的全新统钙化的EPS组织。温志峰等(2010)在新近系叠层石中也发现了类似的席状体, 并认为其是钙化的EPS组织。文中初步认为这些席状体是钙化的EPS组织, 其是由微生物分泌的一种具粘结性的物质, 一般被认为是形成微生物岩的基础(Decho, 1990; Riding, 2000)。
凝块状泥晶组构的特点是1~3, μ m大小的泥晶颗粒“ 凝聚” 而成凝块状、海绵状的泥晶网。凝块状泥晶组构常常发育在叠层石和凝块石中, 与凝块石是不同的概念, 前者是一种显微组构, 而后者是宏观组构(Riding, 2000; 梅冥相, 2007a)。
Riding(2000)认为凝块状泥晶的成分主要为钙化的EPS和捕获的泥晶。平邑剖面官中段微生物碳酸盐岩多以泥晶组构为特征(图 5-4), 虽然均一的泥晶组构包含了一系列可能的成因(梅冥相, 2007a), 但凝聚的泥晶颗粒一般可代表一种有取向的微生物聚居作用(Southam and Donald, 1999)。
平邑剖面官中段微生物碳酸盐岩分布的变化比较快, 这与湖相碳酸盐岩通常具有的单层薄、变化快、韵律明显等特点一致(Cohen and Thouin, 1987; Bolhar, 2007)。相对于海洋水体, 湖泊水体体积比较小, 湖相沉积的碳酸盐对陆源注入、水体涨落等环境变化非常敏感(Freytet and Plaziat, 1982; Arenas et al., 2007), 因此环境对于湖相微生物碳酸盐岩发育的影响可能比海相的要显著。
平邑剖面微生物碳酸盐岩发育层段总的环境背景为:温暖的气候, 浅湖至半深湖的水体, 较少的陆源注入(李熙喆等, 2000; 唐鑫萍等, 2011)。在这一背景下, 文中3类微生物碳酸盐岩的形成环境依然存在一定的区别。
1)核形石。与平邑剖面官中段其他微生物碳酸盐岩相比, 核形石的明显特点是具有包覆颗粒结构。许多学者认为, 包覆颗粒结构与生长过程中的悬浮、翻滚有关, 一般形成于间歇性的水动力环境中(Vé drine et al., 2007; Schlagintweit and Gawlick, 2009)。
核形石纹层的规则程度历来被认为是反映形成环境水动力条件的有力证据(Flü gel, 2004; Gradziń ski et al., 2004)。平邑剖面官中段核形石的纹层多呈规则、连续的同心状(图 3-1), 而这种类型的纹层一般形成于较强水动力条件下; 核形石的核心物质常由陆源石英组成(李熙喆等, 2000; 唐鑫萍等, 2011), 显示其形成环境中含有一定量的陆源碎屑注入。故推测平邑剖面核形石形成于离湖岸较近的浅湖环境, 其水动力条件较强。
李熙喆等(2000)认为, 平邑盆地官中段核形石形成所需的水体动荡环境, 起因于河流入湖所引起的湖流作用, 而且认为, 河流作用的影响是间接的, 而湖流作用影响是直接的, 核形石仍生长于水体相对清澈的环境。
2)叠层石。平邑剖面官中段叠层石与核形石的共同特点是两者均具有明暗相间的纹层结构, 这种纹层一般形成于规律性变化的环境, 如日夜更替、水平面升降、气候变迁等(Monty, 1967; Reid, 2000; 温志峰等, 2010)。现代实例的研究也表明, 现代叠层石主要生活在水体循环、温暖、透光性好的潮间及浅潮下带环境(朱士兴, 1993; Andres and Reid, 2006)。
叠层石与核形石的不同点在于核形石的纹层形成了同心圈状, 并形成了一个相对独立的球体颗粒, 而叠层石没有形成独立的颗粒形态。叠层石一般被认为是原地叠置生长形成的(Andres and Reid, 2006; 韩作振等, 2009), 故与经历翻滚运动而形成的核形石相比, 叠层石形成环境的水动力条件要弱些。
平邑剖面官中段叠层石的亮层比暗层要宽(图 4-2, 4-3), 故在纹层中亮层所占的比例明显要大; 在亮层内常含少量细小的石英碎屑颗粒, 说明存在少量的陆源注入。故综合推测, 平邑剖面官中段叠层石可能形成于离湖岸较近、水深较浅、中等强度水动力的环境。
3)凝块石。常被认为与底栖微生物群落的原地钙化作用有关(Ferris et al., 1997; 刘建波等, 2007)。平邑剖面凝块石多具有不规则的外形, 小凝块体之间常呈互相粘接的特点, 主要成分为细粒的暗色泥晶, 几乎没有发现陆源碎屑颗粒。与核形石和叠层石相比, 它的暗色有机质含量最多、颜色最深, 所反映的水动力条件最弱。故平邑剖面凝块石可能是在较深水、较弱水动力条件下原地钙化形成的, 与巴哈马Lee Stocking 岛的现代凝块形成环境类似, Feldmann 和McKenize(1998)认为该处凝块石发育的水深通常大于叠层石。
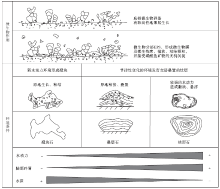
综合以上分析表明, 平邑剖面官中段核形石、叠层石、凝块石的形成环境大致呈现出水动力依次减弱、陆源碎屑物质依次减少、水深依次加大的规律(图6)。
1)微生物碳酸盐岩中的生物。微生物是所有形体微小的单细胞或个体结构较为简单的多细胞、甚至无细胞结构的低等生物的总称(Riding, 2000; 韩作振等, 2009)。底栖微生物群落的发育是形成微生物碳酸盐岩的基础(Baud et al., 2007)。对现代海洋叠层石的研究表明, 微生物碳酸盐岩中的生物主要包括光合原核生物(如蓝细菌等)、真核微体藻类(如褐藻、红藻、硅藻等)、化学自养或异养微生物(如硫细菌等), 以及一些后生动物(如介形虫等)(Konishi et al., 2001)。
除上文提到的球状微生物化石之外, 在平邑剖面官中段还发现了少量介形虫化石(图 4-4)。因此, 在研究区底栖生物群落中, 蓝细菌常作为生产者, 介形虫作为消费者, 组成底栖生态系统。
2)微生物碳酸盐岩形成的生物学过程。在微生物碳酸盐岩形成过程中, 碳酸盐矿物的聚集大致可归纳为3种作用:(1)微生物对碎屑颗粒的捕获和粘结; (2)微生物本身的钙化作用(如蓝细菌的钙化形成球状化石); (3)碳酸盐矿物在生物或沉积物表面上的无机沉淀(Burne and Moore, 1987; Riding, 1991)。在这些作用过程中, 起关键作用的是EPS、微生物膜及微生物席, 它们是微生物碳酸盐岩形成的生物基础(Decho, 1990; 温志峰等, 2004)。EPS是由微生物分泌的一种具粘结性的物质, 可以使微生物附着于基底之上, 并为其提供物理和化学保护(Decho, 1990)。微生物膜是一层有机质薄膜, 由生存于EPS中的薄层细菌群落(几十至几百微米厚)组成, 附着于需水环境的基底(Riding, 2000)。微生物膜继续生长而形成较厚的层被称为微生物席(一般为毫米级), 微生物席可以覆盖较大的沉积面、捕捉沉积物(Riding, 2000; 梅冥相等, 2007)。
在平邑剖面, 前文提到的席状体可认为是钙化的EPS组织, 而凝块状泥晶的成分主要为钙化的EPS和捕获的泥晶, 可指示岩石形成时的微生物作用。
经过综合分析, 提出了研究区微生物碳酸盐岩的成因模式(图 6)。虽然形成微生物碳酸盐岩的因素可能很多, 但经过分析认为, 微生物活动、环境条件是微生物碳酸盐岩形成过程中的2个关键因素。平邑剖面微生物碳酸盐岩正是以微生物活动为共同基础, 在不同的环境条件下形成的。(1)共同的微生物基础, 包括微生物成因组构(EPS、微生物膜、微生物席等)和各种微生物作用(吸附、粘结和钙化作用等)。(2)不同的形成环境, 包括水动力条件、水深、陆源碎屑物注入等, 影响了其结构和形态的发育。微生物作用是形成微生物碳酸盐岩的物质基础, 而环境作用既为微生物作用创造条件, 也影响了微生物碳酸盐岩的发育, 加上平邑古湖盆具有水体体积较小、变化频繁的环境特点, 使环境因素在湖相微生物碳酸盐岩的形成过程中具有更明显的作用。
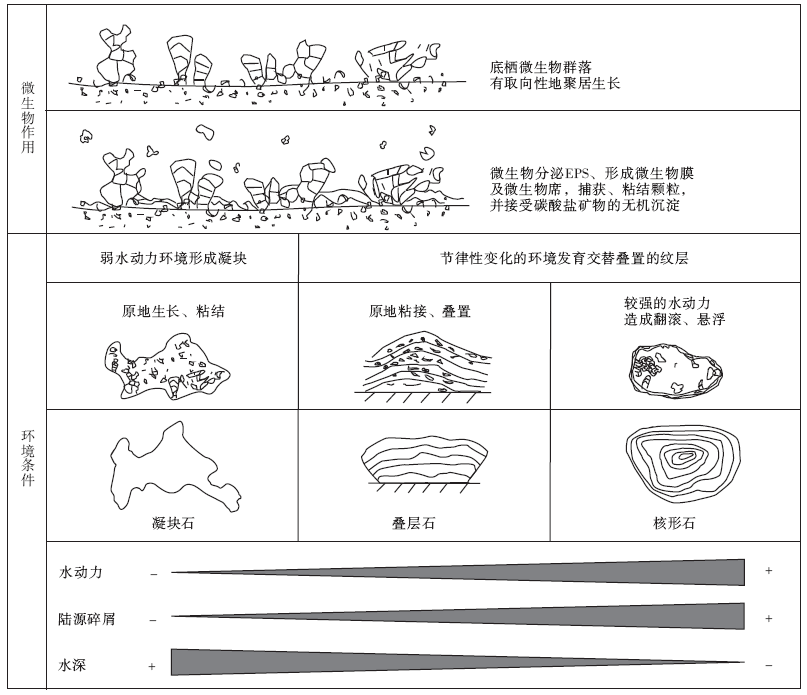 | 图6 平邑盆地湖相微生物碳酸盐岩成因模式Fig.6 Genetic model of lacustrine microbial carbonate rocks from Pingyi Basin |
1)在研究区的核形石、叠层石和凝块石这3类微生物碳酸盐岩内部, 均发现了球状微生物化石、席状体、凝块状泥晶等具有微生物成因特征的显微组构, 表明了岩石形成过程中共同的生长、粘结、钙化等微生物作用。
2)微生物作用对环境变化比较敏感。水体体积较小、变化频繁的湖泊环境, 对微生物碳酸盐岩的发育有较大影响。核形石、叠层石、凝块石的形成环境大致呈现出水动力依次减弱、陆源碎屑物质依次减少、水深依次加大的规律。
3)平邑微生物碳酸盐岩是以微生物活动为共同基础, 在不同的环境条件下形成的。微生物作用是形成微生物碳酸盐岩的物质基础, 环境作用既为微生物作用创造条件, 也影响了微生物碳酸盐岩形态和结构的发育。
致谢 承蒙冯增昭教授对原稿提出宝贵的修改意见和建议,并无私地为作者提供了一些珍贵的参考资料;幸得审稿专家管守锐老师、刘建波老师和谢庆宾老师对原稿进行了耐心细致的审阅,提出了许多宝贵的修改意见,谨此表示衷心感谢!
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|