







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黄河中下游焦作区段现代边滩沉积中的生物遗迹*
[胡斌1, 2  , 王媛媛
, 王媛媛1 , 张璐1 , 单婷婷1 ]
, 王媛媛|
|

第一作者简介 胡斌,男,1952年生,河南理工大学资源环境学院教授,博士生导师,主要从事古生物地层学、遗迹学和沉积学的教学研究工作。E-mail: hub@hpu.edu.cn。
沉积物中的生物遗迹记录是生物与环境相互作用的结果,是沉积环境、气候及生态环境的良好指示标志。文中主要研究了黄河中下游焦作区段现代边滩沉积物中的生物遗迹及其造迹者的组成与分布特点,即:( 1)近岸边滩:几乎无植被,以层面上的生物觅食迹为主,其造迹生物主要有鸟类、长泥甲虫(鞘翅目长泥甲科)和隐翅虫(鞘翅目隐翅虫科)等;( 2)远岸边滩:有大量植物根出现,大多数生物遗迹是层内 U形、 Y形和 W形潜穴以及层面上的 F形、 Y形爬行迹和觅食迹,其主要造迹生物为蝼蛄(直翅目蝼蛄科)、蟋蟀、狼蛛和蠕虫类等;( 3)泛滥平原:出现大量植物根,以层内 J形、漏斗形等的生物居住迹和进食迹为主,主要造迹生物为蝼蛄、蟋蟀、田鼠、蚁狮和狼蛛等。基于沉积物粒度分析、总有机碳测定、菌落平板计数法等手段分析得知,研究区现代生物遗迹的深度随沉积物中微生物群落数量的增加及植物根长度的增长而增加,而丰度和分异度随沉积物粒度的递减及总有机碳含量的增加而增大。该成果一方面能为研究区黄河边滩上生物成因的沉积构造提供新资料,另一方面也能为地史时期河流沉积环境的研究乃至古地理、古生态环境重建提供生物遗迹学方面的科学依据。
About the first author Hu Bin, born in 1952, is a professor and Ph.D.supervisor of Henan Polytechnic University and is mainly engaged in teaching and researches of biological,stratigraphy,ichnology and sedimentology.E-mail: hub@hpu.edu.cn.
Biogenic traces records are the results of the interaction of organisms and environment and the good indicator of sedimentary environment,climate and the ecological environment.The composition and distribution characters of biogenic traces and their makers in modern point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area,Henan Province, are as follows:(1)The main trace-makers on point bar near riverbed are birds and Coleptera.The main traces are the feeding traces of Coleptera and birds on the bedding plane.(2)The main trace-makers on distal point bar and temporal channels are insects as mole crickets,crickets and worms.The main traces are the U-shaped,Y-shaped,W-shaped and vertical feeding burrows of mole crickets,crickets and worms in intrastratal bedding as well as F-shaped and Y-shaped feeding and crawling traces developed on bedding plane.(3)The main trace-makers on flood plain are ant lion,cricket,field mouse and spiders.The biogenic traces are characterized by the J-shaped and funnel shaped feeding and dwelling burrows.By means of grain size analysis,admeasurements of total-organic-carbon,plant bacteria colony counting,the abundance and diversity of traces increase with decreasing grain size and increasing TOC in this study area.The depth of traces below the sediment surface is closely related to the food source and amount of bacteria in the sediments.This study is not only significant to identify the biogenic structures and can be applied to the study of fluvial sedimentary environment,ecology,palaeoichnology research,but also is available to the palaeoclimate,palaeoenviroment and palaeoecology reconstruction.
现代沉积经历了一系列物理、化学及生物作用相互影响的复杂过程, 是多种地质作用的产物, 因而有关现代沉积中气候、土壤、生态系统、地理环境和生物多样性、微生物与宏体生物之间的关系研究对解读古气候、古土壤、古环境及其与生物遗迹丰度和分异度的关系具有一定的启发性(Villani et al., 1999; Vittum et al., 1999; Brake et al., 2002; Hasiotis, 2002, 2004; Hembree and Hasiotis, 2006; Kraus and Hasiotis, 2006; Counts and Hasiotis, 2009)。遗迹学研究的是现代和古代生物生活时在沉积物底层中(层面和层内)遗留下来的活动遗迹(吴贤涛, 1986; 胡斌等, 1997)。这种生物活动遗迹与其沉积底层、沉积环境、生态环境等有着密切的关系。现代生物遗迹研究称为新遗迹学, 它是通过对现代各类生物的造迹活动进行详细的观察和描述, 了解各类造迹生物的活动规律、造迹行为习性、遗迹特点和分布特征及其与沉积底层的关系, 进而分析沉积环境。
国外在上世纪已有大量针对陆相沉积物中现代生物遗迹的系统研究(Pryor, 1967; Ratcliffe and Fagerstrom, 1980; Hasiotis and Bown, 1992; Hasiotis and Mitchell, 1993; Gobetz, 2005; Hembree and Hasiotis, 2006, 2007; Lawfield and Pickerel, 2006; Pearson and Gingras, 2006), 而中国仅刚刚起步。黄河是长度居中国第二、世界第五的河流, 然而针对现代黄河边滩的生物成因沉积构造除鱼类游泳迹(王冠民等, 2003)外少有报道。文中将运用沉积物粒度分析、总有机碳测定、菌落平板计数法等手段分析现代生物遗迹的组成与分布特征及其受控的环境因素, 这一方面能为研究区黄河边滩上生物成因的沉积构造提供新资料, 另一方面也能为地史时期河流沉积环境的研究乃至古地理、古生态环境重建提供科学依据。
研究区位于河南省西北部焦作市武陟县黄河滩(N34° 57', E113° 28'), 隔河相望郑州桃花峪(图 1-A), 海拔约95, m(图 1-B), 为黄河中游与下游的分界点。三门峡至桃花峪区间的河段由小浪底而分为两部分:小浪底以上, 河道穿行于中条山、崤山之间, 为黄河干流上的最后一段峡谷; 小浪底以下, 河谷渐宽, 是黄河由山区进入平原的过渡地段。河道比降变缓, 河床比较稳定, 下切力减弱而侧蚀力增强, 因此河槽逐渐拓宽和曲折, 两岸有滩地出现。河水的pH值为8.08, 溶氧量为4.2 mL/L。
 | 图1 黄河中下游焦作区段现代边滩沉积剖面位置Fig.1 Location of modern point bar deposits section of the Middle-Lower Reaches of Yellow River in Jizuo area |
该地区为大陆季风性气候, 四季分明, 年均气温14.4 ℃, 年降雨量575.1, mm, 全年无霜期约200 d。冬季受西伯利亚— 蒙古高压控制、气候干冷, 春秋季受低压控制。该边滩沉积可划分为近岸边滩沉积、远岸边滩沉积、暂时性水渠沉积和泛滥平原沉积。由于该地区植被稀少, 沉积物主要由粉沙夹黏土质粉沙组成, 同时沙尘暴天气常见, 因此该地区边滩上风沙成因的沉积构造(图 2-A, 2-B, 2-C, 2-E)和椭球形的沙包泥粒在秋冬季较为常见(图 2-D)。这些沉积构造的方向受控于秋冬季风与河水水流。由于河水间歇性的涨落, 远岸边滩上泥裂(图3-B)较为常见。边滩沉积物表面波痕十分发育, 类型众多, 不仅有流水波痕, 还见浪成波痕。波痕以小型为主, 形态多变(图 3), 如有直脊波痕、曲脊波痕、对称波痕、不对称波痕和干涉波痕等。不同类型波痕形成多种类型的层理构造, 从剖面上(图 4)可见到明显的上攀层理、小型槽状交错层理、板状交错层理和楔状交错层理, 水平层理等也较常见, 还可以见到卷曲层理、各类同生变形构造等。沉积物表面生长有少量植被, 生物潜穴发育。
 | 图2 黄河中下游焦作区段边滩上的风沙成因沉积构造Fig.2 Aeolian sedimentary structures in point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area |
 | 图3 黄河中下游焦作区段边滩上的各种形态波痕Fig.3 Various ripple marks in point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area |
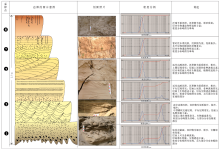
 | 图4 黄河中下游焦作区段边滩剖面上的物理沉积特征及粒度分布特征Fig.4 Physical sedimentary characters and grain size distribution of point bar deposits in the Middle-Lower Reaches of Yellow River in Jiaozuo area |
通过对典型边滩连续剖面(图4)的观察, 自下而上可见底层冲刷面, 沉积物为细沙和粉沙, 并混合有沙泥粒, 向上为小型槽状交错层理、变形层理; 其上为远岸边滩沉积, 沉积物为粉沙、泥质粉沙, 发育上攀层理、缓波状层理、水平层理, 其中层内和层面生物潜穴及植物根迹发育; 最上层为泛滥平原沉积, 沉积物为泥、粉砂质泥, 层内生物潜穴和植物根迹丰富。
通过对剖面上沉积物样品的粒度分析(图 4)发现, 该剖面沉积物粒度自下而上由双峰转变为单峰, 粒径由粗变细, 分选性由差变为极好, 说明水动力强度由强变弱。
为便于对比研究, 本次工作在采样点①、③、⑤、⑦、⑧和②、④、⑥(图 1-B)分别采集了两组分析样品。样品采集时, 用土钻分别采集离层面0~5 cm、5~10 cm、10~20, cm深的样品, 每个样品由采样点中心及其周围共5个点的土样混合而成, 重1, kg左右。将样品分别装入袋中密封并做好标记, 带回实验室置于4 ℃条件下保存, 以供测试分析。
微生物数量测定采用混合平板计数法, 细菌采用牛肉膏蛋白胨培养基。当培养皿中均匀的长满菌落时, 即可裸眼观察并计数。结果按下式计算:
每克土样的菌数=同一稀释度几次重复的菌落平均数× 稀释倍数。
粒度分析选用采样点①、③、⑤、⑦、⑧处的表层沉积物, 通过激光粒度仪Rise-2006测定。总有机碳(TOC)含量的测定选用与粒度分析相同的样品, 使用Apollo 9000型仪器测定。
近岸边滩是指靠近河床的边滩(图 1-B, ①②③④), 几乎没有植被发育, 生物遗迹以层面分布为主, 常见泥甲虫(鞘翅目长泥甲科)、隐翅虫(鞘翅目隐翅虫科)的觅食迹和居住迹及鸟类的足迹。其中鸟类的足迹(图 5-A)沿水陆交界线分布, 与各种波痕相伴生。泥甲虫喜湿, 钻入浅层泥土中觅食腐殖质, 形成层面上呈圆柱状隆起、具分支、且外壁有纹饰的蜿蜒延伸的觅食迹(图 5-B, 5-C), 宽约2~3, mm, 深约2~4, mm, 长约2~30 cm。另外, 该觅食迹同时也是长泥甲科的居住迹, 它们在里面产卵, 1年可繁殖4代(Clark and Ratcliffe, 1989; Lawrence and Newton, 1995)。而隐翅虫经常寄居在长泥甲科的潜穴中(图 5-A, 5-D), 觅食微生物。隐翅虫是喜湿无脊椎动物中很重要的一科, 因为它的生活习性及在近岸边滩的丰度较大, 不仅为鸟类提供了大量的食物来源, 而且为近岸边滩土壤微生物及有机物的分解转化、养分的能量转化和供应起到了基础作用(Obrtel, 1968; Ausden, 2001; Jorge et al., 2009)。因此, 其遗迹经常与长泥甲虫潜穴和鸟类足迹共生(图 5-A)。底层沉积物相对含水量大, 在0.113%~0.132%之间。在夏季雨后, 气温约为33~37, ℃之间, 这些生物遗迹组合常与泥裂、气坑构造(钟建华和周瑶琪, 1999)、微生物席及苔藓共生。
 | 图5 黄河中下游焦作区段近岸边滩生物遗迹Fig.5 Biogenic traces in point par near river bed in the Middle-Lower Reaches of Yellow River in Jiaozuo area |
远岸边滩(图 1-B, ⑤、⑥)是指近岸边滩和泛滥平原之间的边滩。主要植被为狗牙根Cynodon dactylon (Linn.)Pers和苍耳属植物。主要造迹生物为蝼蛄(直翅目蝼蛄科)、蟋蟀等。蝼蛄钻入表层沉积物中觅食, 形成了Y形或F形分枝潜穴(图 6-A, 6-C), 直径约2~6, cm, 长约30~70 cm, 潜穴的分叉点一般多位于植物根处, 并且沿植物根潜入深度约15~37, cm的土壤中进食植物根, 形成与地表近垂直的进食潜穴(图 6-B)。其潜穴直径约2~5, cm, 内部有卵和粪球粒(图 6-D)。底层相对含水量约为0.123%~0.142%。成年蝼蛄选择在底层湿润、松散和轻度盐碱化的土壤中产卵, 两年产卵1次。它们潜入50~149, cm深的土壤深处越冬。到春天, 当地表以下10, cm以内的土壤温度升至10 ℃以上时, 蝼蛄开始爬向表层土壤中进食植物根, 并在地表形成纵横交错的觅食迹。成年蝼蛄在五月份的时候钻入地表以下10~15, cm产卵, 平均1个成年蝼蛄产卵102个, 历时15~22 d。到六月底幼虫继续向地表以下20~38, cm深度挖掘潜穴筑房室化蛹, 历时约22 d。待羽化后, 继续在潜穴中度过冬天(韩召军等, 2008)。
 | 图6 黄河中下游焦作区段远岸边滩上的蝼蛄潜穴及在不同相对含水量底层中的爬迹(箭头所示)Fig.6 Burrows of mole cricket and their crawling traces on different relative water content(as arrows showing)in distal point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area |
此外, 在远岸边滩常见的生物遗迹还有层面上呈近圆形、垂直于层面延伸的垂直潜穴, 直径1.5~2, mm(图 7-A, 7-B, 7-C), 长5~15, cm, 且与植物根迹伴生, 在夏季雨后会大量出现。
 | 图7 黄河中下游焦作区段远岸边滩上的垂直潜穴(箭头所示)Fig.7 Vertical burrows on distal point bar of the Middle-Lower Reaches of Yellow River in Jiaozuo area(as arrows showing) |
边滩上暂时性水渠边(图 1-B, ⑦)造迹生物数量多且沉积物粒度和含水量变化较大, 植被丰富, 生物遗迹丰度和分异度均较高。常见蝼蛄和蟋蟀潜穴等, 且具有一种造迹生物营造多种生物遗迹的特点(图6-E, 6-F, 6-G, 6-H)。
研究区段内泛滥平原沉积(图 1-B, ⑧)上常见各种农作物如花生、黄豆等植物根迹(8-A)。生物遗迹常见垂直潜穴、蜘蛛洞穴(8-E)、蚁狮捕食穴(8-D)、蟋蟀居住迹(8-C)和田鼠居住迹等。其中蚁狮在干燥、砂质的底层中营造漏斗状的觅食迹(图 8-D)。蟋蟀营造层内U形、F形或J形觅食迹和居住潜穴(图 8-C), 直径约8~17, mm, 深度约17~32, cm, 底层相对含水量约为0.125%~0.159%。蟋蟀在潜穴中进食, 产卵。内部可见植物碎屑和粪球粒充填。
 | 图8 黄河中下游焦作区段泛滥平原上的生物遗迹Fig.8 Biogenic traces on floodplain in the Middle-Lower Reaches of Yellow River in Jiaozuo area |
近岸边滩、远岸边滩和泛滥平原沉积物中生物遗迹的组成和分布与沉积底层的性质有一定关系。通过对研究区段黄河边滩沉积物样品进行TOC测定、菌落平板计数法统计、粒度分析、相对含水量测定等分析, 得知沉积物粒度曲线自近岸边滩到泛滥平原由明显的双峰变为单峰, 沉积物粒度变细(图 4)。研究区段的TOC含量自近岸边滩到泛滥平原逐渐增大(图 9; 图10)。对采样点不同底层沉积物中微生物, 尤其是菌落数量的测定, 发现菌落在不同位置和不同深度数量相差很大(表 1)。近岸边滩河水流速变化快而且河水间歇性涨落, 水动力较强, 造成沉积物分选差。而近岸边滩0~5, cm深度位置菌落数量最多(表 1), 这可能是由于河水间歇性的涨落影响了沉积物的氧化还原环境, 从而抑制了深层沉积物中菌落的生长和繁殖。然而也是因为较强的水动力为近岸边滩带来了河水中的营养物质和底层沉积物较高的相对含水量, 从而为近岸边滩的造迹生物如鞘翅目等食泥动物提供了食物来源。这类生物遗迹的分布受两个因素的影响, 除了需具有充足的食物来源, 还受沉积底层的性质影响, 如粒度为粉沙、相对含水量高且沉积物粘结性强(Clark and Ratcliffe, 1989)。因此, 在近岸边滩生物遗迹以层面分布的觅食迹为主。
 | 图9 各采样点TOC含量曲线图Fig.9 TOC content of different sampling points |
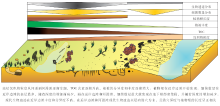
 | 图10 黄河中下游焦作区段现代边滩沉积中生物遗迹分布模式图Fig.10 Distribution model of biogenic traces in modern point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area |
| 表1 黄河中下游焦作区段边滩沉积物中的细菌数量 Table1 Microbios in point bar deposits of the Middle-Lower Reaches of Yellow River in Jiaozuo area |
在远岸边滩和泛滥平原上的沉积物主要受风力搬运, 由于风力变化小, 而且只能搬运有限粒度, 因此粒度小、分选好。底层相对含水量较低, 粘结性较低, 适于生物营造层内进食和居住潜穴。菌落数量最大的深度和多数植物根均集中在5~10, cm的深度, 这是由于植物根系可为微生物栖息生长提供适宜的场所, 其分泌物可使微生物具有更丰富的资源可以利用, 产生明显的根迹效应(项学敏等, 2004; 叶淑红等, 2006; Gilbertson, 2007), 而植物根本身也是多数昆虫的食物来源。因此在远岸边滩和泛滥平原的生物遗迹主要以层内分布的Y形、U形、W形进食潜穴和居住潜穴为主。
1)黄河中下游焦作区段边滩沉积特点为:沉积物粒度曲线自近岸边滩到泛滥平原由明显的双峰变为单峰, 沉积物粒度变细。TOC含量自近岸边滩到泛滥平原逐渐增大。在边滩近岸0~5, cm深度位置菌落数量最多, 远岸边滩和泛滥平原沉积物中的菌落数集中在5~10, cm的深度。
2)生物成因沉积构造分布有以下特点: ①近岸边滩, 几乎无植被, 以层面上的生物觅食迹为主, 其造迹生物主要有鸟类、长泥甲虫(鞘翅目长泥甲科)和隐翅虫(鞘翅目隐翅虫科)等; ② 远岸边滩, 开始有大量植物根出现, 以层内的生物觅食潜穴为主, 其主要造迹生物为蝼蛄(直翅目蝼蛄科)、蟋蟀、狼蛛和蠕虫类等; ③ 泛滥平原, 出现大量植物根, 以层内的生物居住迹和进食迹为主, 主要造迹生物为蝼蛄、蟋蟀、田鼠、蚁狮和狼蛛等。
3)现代生物遗迹的丰度和分异度随着沉积物粒度的变细和TOC的增高而增大。而生物遗迹的深度与植物根和沉积物中菌落数关系密切, 这些菌落可以改变土壤的物理、化学和微环境, 使有机物质转化为营养物质, 为营养物质的循环提供了基础。因此, 沉积底层性质如沉积物粒度分布、相对含水量及沉积物中的TOC含量、菌落数量等共同影响了黄河中下游焦作地区边滩上的现代生物遗迹的丰度和分异度。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|