


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黔北务川-正安-道真地区铝土矿系中生物标志物及其地质意义*
[余文超1  , 杜远生
, 杜远生1 , 周琦2 , 金中国3 , 汪小妹4 , 覃永军2 ]
, 杜远生, 周琦|
|

第一作者简介 余文超,男,1988年生,中国地质大学(武汉)在读博士生,主要从事沉积地质学方面的研究。E-mail:evilchaos@sina.com。
通讯作者简介 杜远生,男,1958年生,中国地质大学(武汉)教授,博士生导师,主要从事造山带沉积地质学研究。E-mail:duyuansheng126@126.com。
黔北务川 -正安 -道真地区铝土矿位于石炭系黄龙组和二叠系梁山组之间,主要属于沉积型铝土矿。通过气相色谱( GC)与气相色谱 -质谱( GC-MS)手段分析铝土矿系钻孔岩心中的生物标志物,发现了含量丰富的正构烷烃、类异戊二烯烷烃、萜类化合物与甾类化合物。对 10个样品饱和烃组分 GC分析表明,正构烷烃主要呈双峰式分布特征,碳数分布 n-C14~n-C35,CPI值范围为 0.90~3.45,表现出奇偶优势。姥鲛烷 /植烷( Pr/Ph)值为 0.38~077,表现出明显植烷优势。对 3个样品饱和烃组分 GC-MS分析表明,萜烷类化合物以 C30藿烷占优势,相对丰度由大到小依次为五环三萜烷、三环萜烷、四环萜烷,并检测出少量的 γ-蜡烷。规则甾烷 C27-C28-C29呈近 L型或 V型分布,并含少量 4-甲基甾烷。根据生物标志物特征参数,结合岩心样品岩相学特征,确定了铝土矿系中有机质的陆上植物与低等菌藻类双重来源,铝土质沉积时为偏酸性较还原环境,明确了铝土矿系的形成过程受到来自陆上与沉积水体内两个古生态系统的双重作用。
About the first author Yu Wenchao,born in 1988,is a Ph.D.candidate of China University of Geosciences(Wuhan)and is mainly engaged in research of sedimentology. E-mail:evilchaos@sina.com.
About the corresponding author Du Yuansheng,born in 1958,is a professor and Ph.D.supervisor of China University of Geosciences(Wuhan)and is mainly engaged in the researches of sedimentology and geology of orogenic belt. E-mail:duyuansheng126@126.com.
The bauxite deposits between the Carboniferous Huanglong Formation and Permian Liangshan Formation in Wuchuan-Zheng'an-Daozhen area,northern Guizhou Province mostly belong to sedimentary bauxite.Abundant biomarkers including n-alkanes,isoprenoid,terpenoids and steroids in drilling core samples are detected by GC and GC-MS.The results of GC reveal that most of n-alkane in ten samples are composed of n-C14~n-C35 and characteristic of double-peak distribution. CPI ranges from 0.90 to 3.45,indicates obvious odd-even carbon number predominance.Pristane to phytane ratios(Pr/Ph)vary from 0.38 to 0.77.According to the results of GC-MS,the relative abundance from higher to lower is pentacyclic triterpanoids,tricyclic terpane,tetracyclic terpane,and C30 hopane is most abundant in the terpanes,meanwhile,small quantities of gammacerane are detected.Regular steroids distributed in “L” or “V” shape includes a small amount of 4-methyl sterane.Combined with petrographical characters of samples,the biomarker parameters indicate a reductive sedimentary environment which shows slant acidity,the organic matter in bauxite deposit originate from both terrestrial plant and bacteria/algae.It clearly shows double influences from terrestrial and aquatic palaeoecosystems during bauxite forming process.
铝土矿, 包括广义的铝土岩, 其形成与炎热潮湿的气候条件紧密相连(冯增昭, 1994), 因为这种特殊的气候条件有利于风化作用, 从而产生红土化物质, 为铝土矿的形成提供成矿母质(Price et al., 1997; Bogatyrev et al., 2009)。现今中国大部分铝土矿系(指含铝土矿的地层层系)沉积中都可见植物化石或其碎片, 少数情况下甚至可见到动物化石(廖士范和梁同荣, 1991), 这与铝土矿形成时期气候炎热、雨水充沛、生物繁盛的特征一致。铝土矿以其不同的形成过程可概括为原地风化残留与风化后经过搬运两种基本成因模式, 因此无论铝土矿系的种类如何, 都会受到陆上风化搬运过程及盆地内沉积过程的双重影响, 而铝土矿系所保留的地质信息, 正是这两个过程的综合反映。漫长的风化过程使得铝土矿中普遍存在含量较高的黏土矿物, 这有利于有机质的保存, 因为黏土矿物与有机质之间可以通过氢键、静电力、范德华力与离子键偶极力等键合方式结合形成有机黏土复合体(Cai et al., 2007)。保存在黏土矿物中的有机质非常稳定, 所以之前的研究发现铝土矿中普遍富集有机质(刘长龄和覃志安, 1999; Laskou and Economou-Eliopoulos, 2007; Hao et al., 2010)。近年来, 生物有机质对铝土矿形成具有重要影响这一认识逐步被人们接受(刘长龄, 1999), 但这种重要性, 不仅体现在有机质在搬运过程中对于胶体溶液的护胶作用方面, 更反映在微生物活动与铝土矿成矿的协同作用方面, 目前已在铝土矿系中发现很多同沉积时期微生物活动活跃的证据(Laskou and Economou-Eliopoulos, 2007; Hao et al., 2010)。文中通过对黔北务川-正安-道真地区铝土矿岩心中生物标志物的分析, 探讨了铝土矿系所记载的古生态系统。
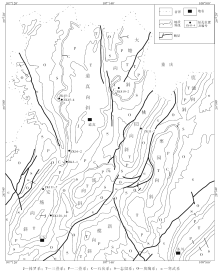
务川-正安-道真地区(简称务正道地区)铝土矿成矿带主体分布于贵州北部的务川、正安和道真地区, 向北直至重庆南川、武隆等地, 呈NNE向展布, 绝大部分矿床点集中分布在正安褶皱带的向斜扬起端(图 1)。含铝土矿地层为大竹园组, 介于上石炭统黄龙组和中二叠统梁山组之间, 上下均为平行不整合接触。志留纪末期至早泥盆世的广西运动的远程效应促使黔北地区抬升为陆、遭受经长期的古风化剥蚀, 使得区内大部分地区的志留系残缺不全(刘平, 2007; 殷科华, 2009)。而在中晚石炭世时, 研究区经历了一次海侵过程, 在部分地区形成了黄龙组灰岩; 晚石炭世的紫云运动使该区再次发生隆升, 直至中二叠世栖霞期才有海水进入(刘平, 1993)。在这段漫长的风化剥蚀过程中, 黄龙组灰岩与韩家店组页岩一同暴露地表, 为铝土矿的形成提供了物源与成矿所需的准平原化沉积间断期。此外, 古地磁研究表明, 石炭纪贵州处于南纬8° ~14° 的热带地区(王俊达和李华梅, 1998), 而且贵州北部地区石炭纪靠近东西向的狭长形扬子海海湾, 故而气候湿润(廖士范, 1992; 陈旭等, 2001)。据此推断, 当时古气候总体特点为炎热潮湿, 非常有利于铝土矿的形成。
 | 图1 黔北务正道地区地质略图及钻孔位置分布Fig.1 Distribution of drilling cores and simplified geological map of Wuzhengdao area, northern Guizhou Province |
本次实验所涉及到的10个样品来自于研究区内8口不同的钻孔岩心(图 2), 样品岩性特征与矿物组成见表1。样品中或可见植物化石碎片(图 3-B, 3-C), 或可见碳质碎屑(图 3-D)。也有样品(ZK11-2-1)因绿泥石含量较高而呈现灰绿色(图 3-A)。样品在粉碎前, 均进行预处理:首先去除掉可能遭受污染的岩心表面, 再用去离子水与超声波清洗机清洗样品, 最后用二氯甲烷淋洗样品表面, 放入干燥箱中于40, ℃恒温烘干。将样品研磨至100目以下, 称取样品粉末约100 g, 用滤纸包裹后放入索氏抽提器中, 向与索氏抽提器连接的平底烧瓶中注入适量的90%二氯甲烷与10%甲醇的混合液作为溶剂并加入铜片以除硫, 每个样品中加入氘代与胆烷各30 μ L(浓度为0.05910 μ g/μ L)
| 表1 黔北务正道地区铝土岩系样品描述 Table1 Description of samples of bauxite-bearing strata deposit in Wuzhengdao area, northern Guizhou Province |
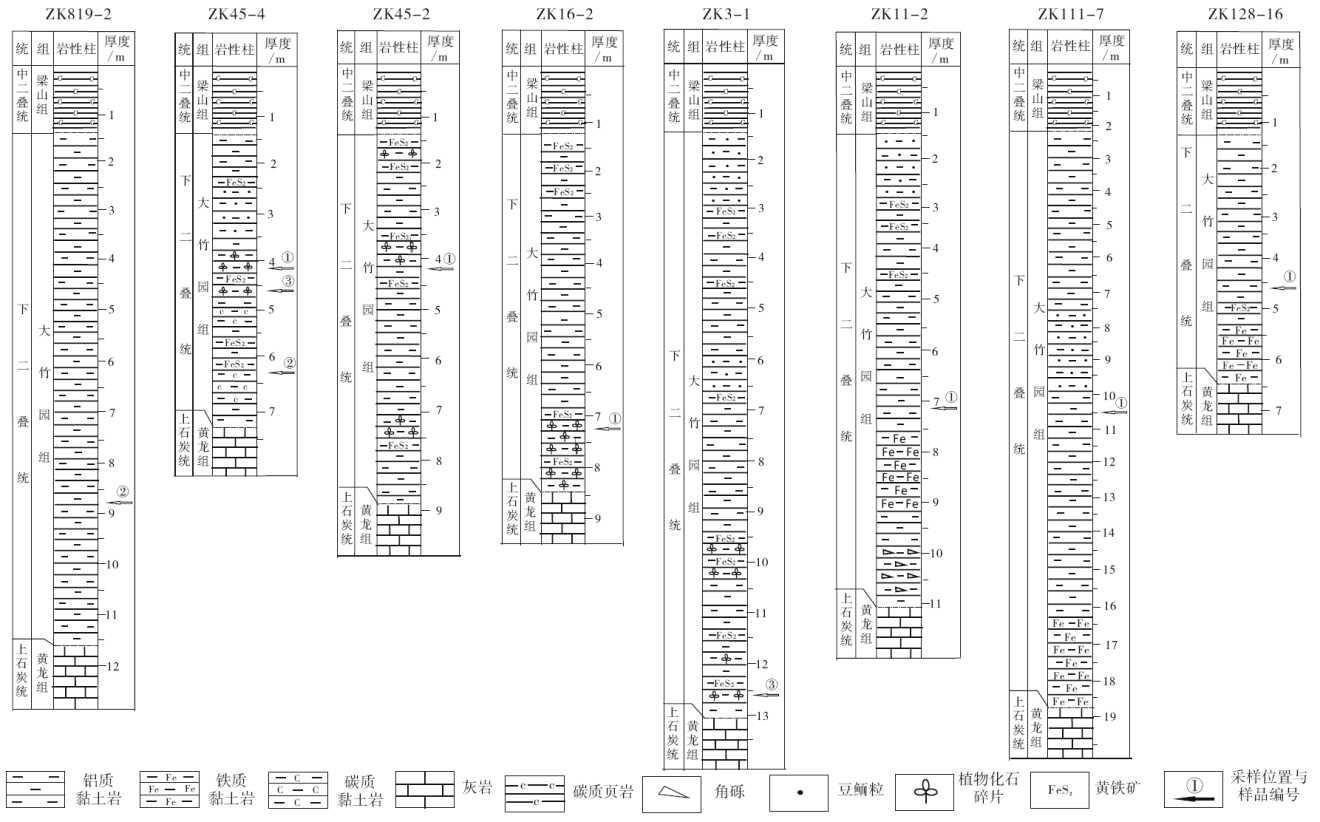 | 图2 黔北务正道地区取样钻孔含矿岩系柱状图Fig 2 Stratigraphic column of bauxite-bearing strata deposits in Wuzhengdao area, northern Guizhou Province |
 | 图3 黔北务正道地区部分岩心样品照片Fig.3 Photos showing drilling core samples in Wuzhengdao area, northern Guizhou Province |
作为标样。在50 ℃恒温水浴锅中抽提72 h。抽提完毕后的样品在40 ℃恒温下旋转蒸发, 残留物转移至细胞瓶中, 蒸干溶剂后得到样品氯仿A。族组分分析用硅胶柱层析法, 正己烷、二氯甲烷与甲醇作为洗脱剂, 获得饱和烃、芳烃与非烃组分。实验所使用的棉花、滤纸和硅胶均先由二氯甲烷抽提72 h, 玻璃器皿每次使用前均经马弗炉灼烧8 h以杜绝污染。样品饱和烃组分气相色谱(GC)分析用Shimadzu GC 2010气相色谱仪, FID检测器和ZB-1石英毛细管柱(60 m× 0.25 mm× 0.25, μ m); 载气为氦气, 升温程序:70 ℃恒温3 min, 以3 ℃/min升温至300 ℃, 恒温30 min。3个样品饱和烃组分采用HP6890 GC/HP5973 MSD气相色谱— 质谱(GC-MS)联用仪进行生物标志化合物分析。实验条件:离子源温度180, ℃, 离子源电离能量70, eV。DB-5MS石英毛细管柱(60 m× 0.25 mm× 0.25, μ m)。升温程度:70, ℃恒温1 min, 以10 ℃/min的速度升温至210 ℃, 然后以3 ℃/min的速度升温至300 ℃, 恒温30 min。
通过GC与GC-MS手段, 在样品饱和烃馏分中检测出了正构烷烃、异戊二烯烃、萜类化合物和甾类化合物。
各个样品在气相色谱图上呈现不尽相同的分布特征(图 4; 表2), 碳数分布范围为n-C14~n-C35, 除1个样品(ZK45-4-3)呈现后峰式分布外, 其余样品均呈现双峰式分布, 但主峰位置有所区别, 可进一步划分为以前峰(n-C18或n-C20)为主峰的双峰式分布与以后峰(n-C31)为主峰的双峰式分布。n-C17/n-C31值范围为0.08~1.15。n-
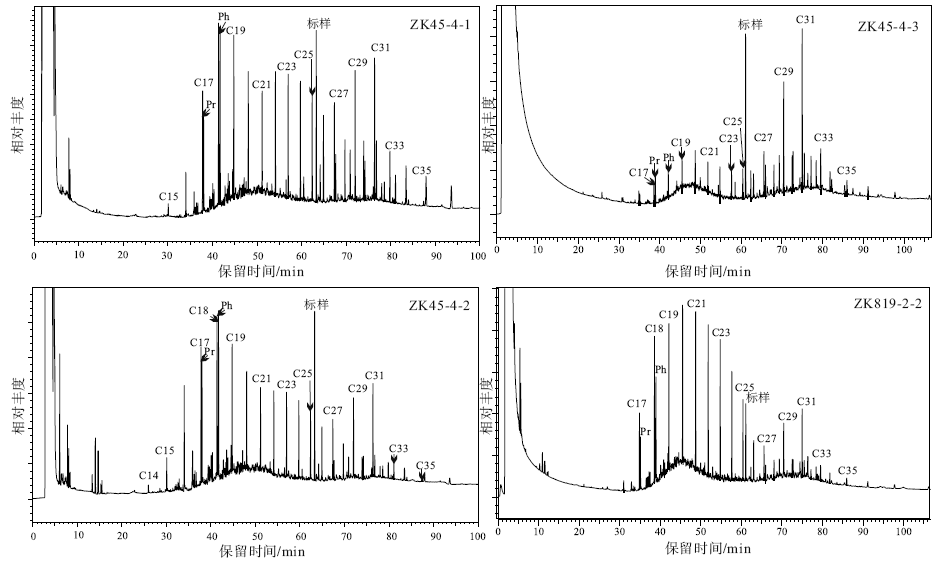 | 图4 黔北务正道地区部分样品饱和烃气相色谱图Fig.4 Saturated hydrocarbon fraction of partial samples gas chromatography in Wuzhengdao area, northern Guizhou Province |
| 表2 黔北务正道地区正构烷烃和类异戊二烯烷烃气相色谱分析数据 Table2 GC analysis data of n-alkanes and isoprenoids in Wuzhengdao area, northern Guizhou Province |
有的样品在n-C18~n-C24之间具有明显的偶碳数优势。CPI值范围为1.99~3.45(除1个样品为0.90外), 具较明显的奇偶优势分布, 且CPI值并未随深度发生改变, 这与区域上二叠系煤变质程度所指示的有机质成熟度存在一定差异, 推测这种奇偶优势可能主要与陆源有机质的大量输入有关(Hatch et al., 1987; Peters, 2005)。
样品中均检出含量较高的以姥鲛烷(Pr)和植烷(Ph)为代表的规则类异戊二烯烃(图 4; 表2)。Pr/Ph值范围为0.38~0.77, 均具有较明显的植烷优势。Pr/n-C17与Ph/n-C18两个比值的变化范围分别为0.50~1.28与0.59~1.67。(Pr+Ph)/(n-C17+n-C18)值范围为0.56~1.47。
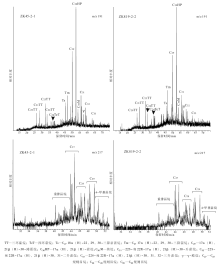
3个样品经过GC-MS测定, 在m/z 191质量色谱图上表现出类似的萜烷分布特征(图 5)。五环三萜烷中的藿烷系列最为明显, 其次为三环萜烷系列, 另见少量四环萜烷, 其相对含量由大到小依次为五环三萜烷、三环萜烷、四环萜烷, 其中以C30藿烷含量最高, C29降藿烷次之。五环三萜烷碳数分布范围为C28-C33, 并检测出了少量的γ -蜡烷, 没有检测到奥利烷或羽扇烷等其他非藿烷类五环三萜烷, 这与奥利烷等被认为是白垩纪或更年轻时代高等植物标志物的认识一致(Moldowan et al., 1994; Peters et al., 2005)。C27 18α (H)-22, 29, 30-三降新藿烷(Ts)与C27 17α (H)-22, 29, 30-三降藿烷(Tm)的比值(Ts/Tm)范围为0.51~0.66, γ -蜡烷与藿烷系列化合物的峰值— — C30藿烷的比值(γ /C30HP)范围为0.11~0.19, 17β (H), 21α (H)-莫烷与其相应的17α (H), 21β (H)-藿烷的比值(莫藿比)为0.13~0.15, 符合新生代以前的地层中该值低于0.15的特征(Peters et al., 2005)。三环萜烷相对丰度较低, 碳数分布范围为C20-C29, 各样品均以C23为主峰, 且C25之后各对峰的相对含量降低。此外, 四环萜烷碳数主要为C24。
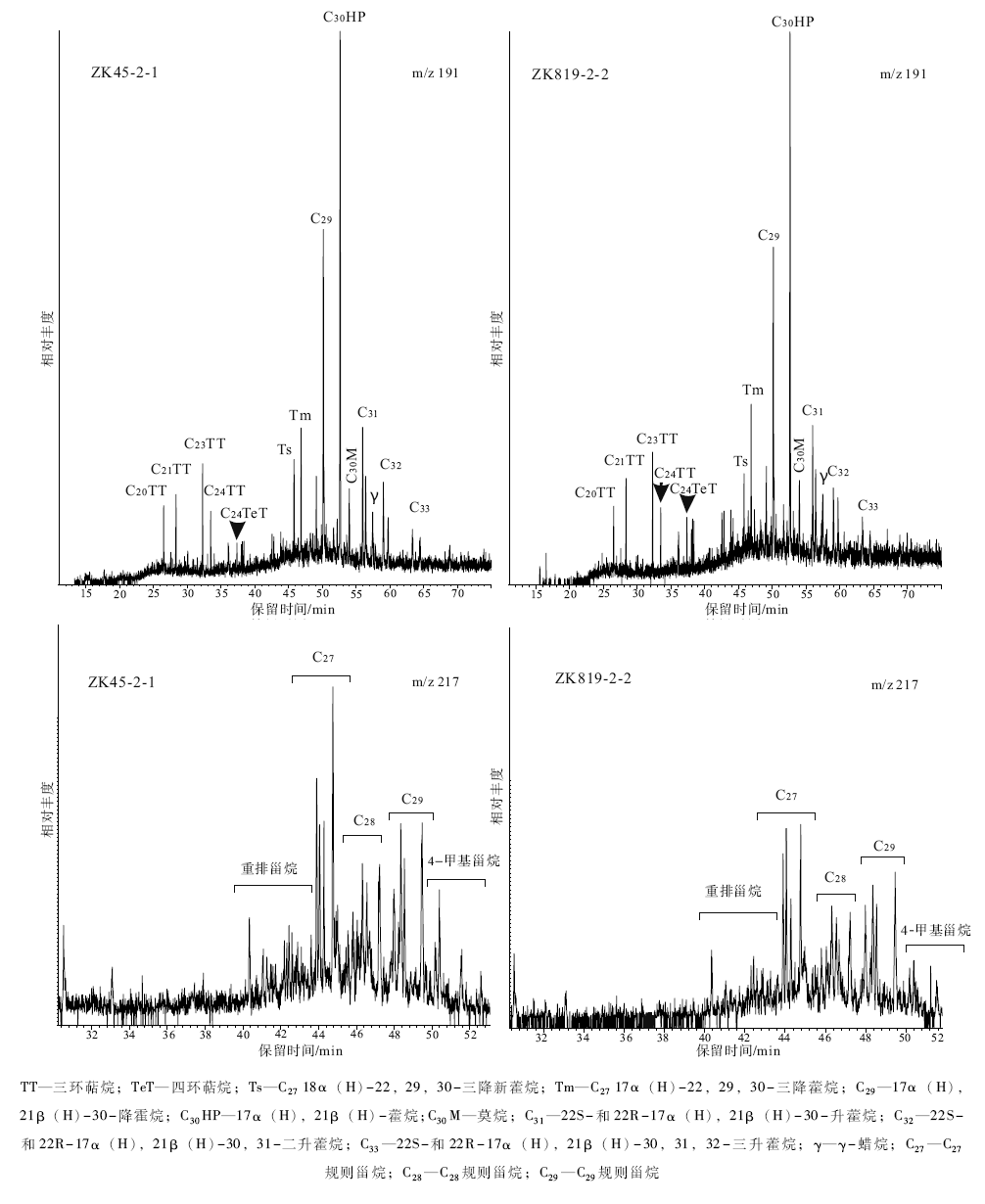 | 图5 黔北务正道地区部分样品饱和烃质量色谱图(m/z 191, m/z 217)Fig.5 Saturated hydrocarbon fractions of partial samples mass spectrogram(m/z 191, m/z 217)in Wuzhengdao area, northern Guizhou Province |
所检测的样品中甾类化合物保存情况较好, 在m/z 217质量色谱图上可见完整的甾类化合物系列(图 5), 其主要成分为碳数分布为C27-C29的规则甾烷与重排甾烷系列和少量C30的4-甲基甾烷。C27-C28-C29规则甾烷呈近L型或V型分布, 其中C27或C29规则甾烷具有最高的相对含量。C27重排甾烷/C27规则甾烷的比值范围为0.31~0.37(表 3)。
| 表3 黔北务正道地区样品藿烷与甾烷系列化合物参数 Table3 Geochemical parameters of hopene compounds and sterane compounds in Wuzhengdao area, northern Guizhou Province |
正构烷烃的分布特征, 对于确定母质输入具有重要的指示意义。低碳数的正构烷烃(碳数小于21的正构烷烃), 代表低等的菌藻类生源输入, 如蓝绿藻类来源的正构烷烃以n-C14~n-C19为优势(Han and Calvin, 1969); 而高分子量(碳数大于25的正构烷烃)奇碳数正构烷烃, 既可以直接存在于植物体内(谢树成等, 2007), 也可以来自于高等植物中的蜡, 蜡水解为含偶数碳的高分子量酸和醇, 在还原环境下通过脱羧基和羟基转化为长链奇数碳正构烷烃(Eglington and Hamilton, 1963)。特别值得注意的是, 碳数为n-C23的正构烷烃来源可能较为复杂, 在大多数样品中, n-C23的含量与相邻奇碳数正构烷烃(n-C21、n-C25)的含量相近(图 4)。前人认为n-C23与n-C21、n-C25可能具有相同的来源, 其均来源于一种高度分异的藻类(van Kaam-Peters et al., 1997), 但对于现代植物或泥炭中正构烷烃碳数分布的研究发现, n-C23主要来自于以Sphagnum(泥炭藓)为代表的陆生苔藓植物(Nott et al., 2000)。
黔北地区在晚石炭世到早二叠世发生喀斯特化, 并且其古气候特征非常有利于植物的生长, 廖士范和梁同荣(1991)根据区内铝土矿层中具较多植物根茎化石碎片但缺少高大乔木遗迹而认为其陆上植被系统可能类似现代热带草原。而对于现代喀斯特地区的植被研究表明, 在岩溶风化过程中, 灌木与苔藓植物广泛发育于适宜气候条件下的风化岩石和土壤表面, 并且会显著提高岩石与土层表面的含水量从而促进风化过程的发生(曹建华等, 2001; Lenton et al., 2012)。研究区样品正构烷烃多为双峰式分布, 并且在钻孔岩心中观察到陆上植物化石碎片(表 1; 图3-B, 3-C), 这说明其有机质来源不仅包括低等浮游生物、藻类及细菌生物, 还可能包括来自陆上维管束植物(如蕨类与裸子植物)与非维管束植物(如苔藓植物)的贡献, 并且, 气相色谱图上存在后峰大于前峰的现象表明, 有时这种来自陆上植物的有机质输入甚至占有机质输入总量的大部分。
姥鲛烷(Pr)和植烷(Ph)一直以来都被认为是光合自养生物叶绿素侧链上植醇的成岩产物(Brooks et al., 1969; Rontani et al., 2003), 而由于类异戊二烯烃抵抗异养生物降解作用较正构烷烃更强, 所以正构烷烃降解作用的发生先于类异戊二烯烃(Peters et al., 2005)。因此, Pr、Ph之和与n-C17、n-C18之和的比值(Pr+Ph)/(n-C17+n-C18)可以作为评价自养生物与异养生物对于地质体中有机质贡献量相对比例的指标(Chen et al., 2011)。样品中(Pr+Ph)/(n-C17+n-C18)值为0.56~1.47, 且多数样品此项指标大于1, 表明自养生物有机质来源占优势, 这与正常生态系统中生产者数量大于分解者数量的认识相符。
萜烷化合物中, 三环萜烷与藿烷系列的分布特征和相对丰度同样受控于母质类型。样品在m/z 191质量色谱图上均表现出相对丰度小于藿烷的三环萜烷分布特征, 但其碳数分布(C20-C29)又较陆源植物为主要母质类型的窄碳数分布(C18-C24)宽, 代表了双重生源的输入特征。藿烷主要来源于原核生物, 细菌中广泛存在着合成包括C30里白烯和里白醇、C35细菌藿多醇等藿类的生化过程(谢树成等, 2011)。样品中C30藿烷为主峰, 且呈C31> C32-C35的分布特征, 而这正是细菌有机质输入的特征。
甾烷因其复杂的立体化学特征而具有重要的地球化学意义。甾烷指纹的复杂性主要是受到了来源的差异与成熟度差异引起的立体化学多样性的影响(Philp, 1985)。之前的研究中, 比较强调原油和烃源岩中C27/C29甾烷的相对含量, 并以此说明来源输入的不同, 对比的依据是C27甾烷主要是海相来源而C29主要是陆相植物来源(Huang and Meinschein, 1978)。但这是一种非常简化粗略的方法, 因为在前寒武纪及早古生代地层中海相来源的C29甾烷化合物的发现不断被报道(孟凡巍等, 2006)。总的看来, 类似于本次研究过程中规则甾烷C27-C28-C29化合物近似“ L” 与“ V” 式的分布规律一般被认为指示了一种陆上与水体内双重作用下的有机质来源。由于规则甾烷被认为来自真核生物(主要是藻类与陆源植物), 而17α (H)-藿烷主要来自细菌等原核生物, 因此它们之间的比值可以表征真核生物与原核生物对于有机质的贡献量, 从而可以反映原核生物与真核生物的相对变化(Cao et al., 2009; 谢树成等, 2011)。样品中规则甾烷/17α (H)-藿烷值为0.35~0.54, 指示陆源和(或)较强细菌活动所引起的大量原核生物生源有机质输入特征(Peters et al., 2005)。此外, 观测到的少量C30的4-甲基甾烷, 早先被解释为沟鞭藻来源(Peters, 2005), 但是沟鞭藻真正的化石记录始于晚三叠世, 而在早古生代地层中发现的4-甲基甾烷, 被认为来自于类似沟鞭藻祖先的物种(Moldowan et al., 1996; Moldowan and Talyzina, 1998)。
类异戊二烯烷烃中姥鲛烷(Pr)和植烷(Ph)的比值一直以来都作为判别沉积环境中氧化还原条件的经典指标。这个指标主要依据在假定Pr是由植醇经各种氧化和脱羧反应形成、Ph是由植醇经过加氢作用和脱水作用形成的前提下, 认为Pr形成于氧化环境而Ph形成于较还原环境。应当指出, 这种假说提出后, 已经有了一些变化。因为之后发现维生素E也可成为Pr的重要来源(Goossens et al., 1984), Ph也可来自于古菌类。只有肯定Pr与Ph多数来源于叶绿素的情况下, Pr/Ph< 1或Pr/Ph> 1的情况才可分别指示氧化与还原环境, 但在实际研究过程中, Pr与Ph的来源较难确定, 因此, 一个较为严格的标准是, Pr/Ph< 0.5 为强还原性膏盐沉积环境, 0.5~1.0为还原环境, 1~2为弱还原— 弱氧化环境, 大于2者见于偏氧化性环境, 如河湖及滨海沼泽或浅湖/海沉积(王春江等, 2005)。而对于本次研究中Pr/Ph值在0.8~2.5之间的样品, 用Pr/Ph指标判断氧化还原条件应该非常慎重(Volkman and Maxwell, 1986; Peters et al., 2005)。10个样品的Pr/Ph值范围为0.38~0.77, 其中7个分布在0.5~1.0的区间内, 也出现了3个小于0.5的低值。结合铝土矿系的岩相学特征— — 多为深灰色、深黑色等还原环境的指示色并伴有大量原生或次生黄铁矿, 说明铝土矿系的沉积环境应为低Eh值的还原环境。但是, 由于样品中并未发现伽马蜡烷的高值且未观察到石膏、石盐等标志性矿物, 结合当时的古气候条件看来, 并没有到达高盐度强还原性的膏盐沉积环境。
Ts/Tm的比值不仅和有机质成熟度有关, 也与源岩形成时的沉积环境有关(Moldowan et al., 1986; Chen et al., 2011), 黏土矿物的存在也会促使Tm向Ts转化的过程(Xie et al., 2007)。当沉积环境的Eh值较低, 即较为还原的条件下, Ts/Tm值增加(Peters et al., 2005)。研究样品Ts/Tm值范围为0.51~0.66, 指示较为还原的环境, 与Pr/Ph值所指示的沉积环境一致。
γ -蜡烷是一种C30三萜烷, 它可以通过还原四膜虫醇生成, 而四膜虫醇被认为是一种在某些原生生物、光合作用细菌和一些可能的其他生物的细胞膜中取代了甾类化合物的类脂化合物(王锐良等, 1990)。强还原超盐度条件受到这些能够产生四膜虫醇生物的偏爱, 因此大量γ -蜡烷(一般以γ -蜡烷/α , β -藿烷比值的形式表征)的出现, 指示了这种强还原超盐度的沉积环境。本次研究样品中γ -蜡烷普遍存在, 但相对含量很低, γ -蜡烷/C30藿烷值范围为0.11~0.19, 说明铝土矿系形成时水体并未出现异常的超高盐度, 这与前面讨论的Pr/Ph出现低值的解释相对应。此外, 类似样品中三环萜烷相对丰度小于藿烷, 且C25之后各对峰的相对含量降低的情况一般出现在淡— 微咸水沉积环境中(孟仟祥等, 2004), 这也符合之前利用较低γ -蜡烷含量对于沉积环境的判断。
重排甾烷/规则甾烷值同样也受热成熟度与源岩无机特征的影响, 在成熟早期到成熟期范围内主要取决于沉积环境中的黏土含量与酸性物质(Peters et al., 2005); 因为这两种物质的催化作用被认为是沉积物中重排甾烷形成的机理(Rubinstein et al., 1975)。铝土矿系中普遍含有黏土矿物(廖士范和梁同荣, 1991), 其C27重排甾烷/C27规则甾烷值为0.31~0.37, 可能表征了一种酸性(低pH值)、还原(低Eh值)的环境。
黔北地区铝土矿形成于特殊的气候环境下, 伴随着这种温暖潮湿气候, 陆上植被繁盛, 土壤间、沉积水体内微生物活跃, 这也正是现今发生典型红土化地区(如热带雨林气候带与热带草原气候带)的环境特征(王建, 2001)。红土化为铝土矿的形成提供了成矿母质。如前所述, 与典型新生代红土型铝土矿不同, 黔北铝土矿并非单纯红土物质在地表原地堆积风化富集而形成的铝土矿类型, 其形成过程还包括搬运与再次沉积。因此, 黔北铝土矿中的正构烷烃呈现类似于有陆源物质输入的水体沉积环境的双峰式分布特征。黔北铝土矿中的生物标志物实际上来自于两个生态系统的双重作用, 即风化与搬运过程时的陆上生态系统与沉积过程中的水域生态系统, 其中, 前者的生产者主要是陆上植物, 后者的生产者主要是水体中的藻类与光合细菌。在陆上风化阶段, 特别是在石炭纪风化阶段发生喀斯特化的黔北地区, 不仅有着陆上植物的生源输入, 微生物的活动同样也会带来较大影响(曹建华等, 2001), 因为无论是碳酸盐岩还是硅酸盐岩, 其风化过程都伴随着强烈的微生物活动(连宾等, 2005, 2008; 连宾, 2010)与陆生植物的影响(潘根兴等, 2000; 曹建华等, 2001)。而在搬运过程及之后铝离子胶体溶液发生沉降的过程中, 同样伴随着微生物活动及有机质, 例如腐殖酸在铝质迁移中的护胶作用(陈履安, 1996)及微生物絮凝剂对于铝质的絮凝作用(杨慧芬和张强, 2004; 盛艳玲等, 2006)。生物标志物所指示的酸性还原环境, 正是铝土物质就位之后的沉积环境。这与前人研究认为的铝土矿古风化壳或附近的湖沼及潟湖水域具有类似沼泽化的低pH、低Eh沉积环境特点的结论(刘长龄等, 1992)相吻合。而进入沉积水体之后, 水体内的生物再次作用于铝土物质, 在铝土矿内所含生物标志物方面能够得以体现的是其中作为生产者的光能自养型菌藻类与化能自养型细菌(如硫细菌、硝化细菌等)以及作为分解者的细菌与真菌等, 这也说明了为何生物标志物中会有大量原核生物生源有机质输入的特征。
1)黔北务川-正安-道真地区铝土矿系富含正构烷烃、类异戊二烯烃、萜类化合物与甾类化合物。正构烷烃呈双峰式分布, Pr/Ph值显示明显的植烷优势。萜类化合物以藿烷类五环三萜烷为主, 三环萜烷次之, 并有少量四环萜烷。甾类化合物主要为规则甾烷、重排甾烷及少量4-甲基甾烷。
2)生物标志物参数显示, 铝土矿系中有机质来源主要是陆生植物与菌藻类低等生物共同贡献。
3)沉积环境参数显示, 铝土矿系在沉积时应该处于偏酸、还原、缺氧的沉积环境。
4)铝土矿中的有机质来源于当时的陆上生态系统与水域生态系统的双重作用。生物标志物中大量原核生物生源有机质输入特征显示, 在铝土矿形成过程中伴随着强烈的微生物活动。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|