

{kind=link}
{kind=link}
{kind=link}
南天山洋古地理及构造演化:西南天山放射虫硅质岩与地层记录的再认识*
[吴浩若 , 李忠]
, 李忠]
, 李忠]
|
|

第一作者简介:吴浩若,男,1940年生,研究员,从事地层古生物学与古地理学研究。E-mail:wuhaoruo@mail.iggcas.ac.cn。
泥盆纪—石炭纪放射虫硅质岩在西南天山广泛分布,从东部的独库公路沿线到西部的阿合奇中—吉边境构成一条深水沉积带。独库公路沿线已发现中泥盆世晚期至早石炭世维宪早期的放射虫硅质岩,可用“库勒湖组”统称。从志留系顶统科克铁克达坂组经下泥盆统阿尔腾柯斯组到库勒湖组的生物地层和沉积相研究表明了南天山洋从浅海到深水洋盆的演化过程。南天山洋是塔里木北缘浅海陆架裂解产生的小洋盆。构造古地理和生物古地理研究表明,南天山洋是古特提斯的分支洋盆,不属古亚洲洋范围。塔里木以南的古特提斯分支洋盆,在早石炭世及之后的继续扩张,使塔里木北移,导致南天山洋和准噶尔—北天山区的古亚洲洋在早石炭世晚期和晚石炭世相继消亡。
About the first author: Wu Haoruo,born in 1940,is a professor researcher and engaged in palaeontology,stratigraphy and palaeogeography.E-mail:wuhaoruo@mail.iggcas.ac.cn.
Devonian-Carboniferous radiolarian cherts are widely distributed in southwestern Tianshan,and formed a deep-water sedimentary belt from the Dushanzi-Kuqa highway on the east to the Aheqi area on the west.The radiolarian cherts ranging from the late Middle Devonian to early Visean of Early Carboniferous were found along the Dushanzi-Kuqa highway,and may be used the name “Kulehu Formation”.Studies of the biostratigraphy and sedimentary facies on the succession from the Pridolian Keketiekedaban Formation through the Lower Devonian Artengkesi Formation to the Kulehu Formation indicate the evolution of South Tianshan Ocean from shallow marine to deep oceanic basin.The South Tianshan Ocean was a small oceanic basin caused by rifting of the continental shelf on north Tarim margin.Tectono-palaeogeographic and palaeobiogeographic studies indicate that the South Tianshan Ocean was a branch of the Palaeo-Tethys and not belongs to the scope of the Palaeo-Asian Ocean.The spreading of branches of the Palaeo-Tethys on the south to Tarim in the Early Carboniferous and afterwards led the drifting northward of Tarim and the closing of the South Tianshan Ocean and the Palaeo-Asian Ocean in Junggar and north Tianshan region.
近年来, 西南天山地质研究备受关注, 该地区被认为是研究中亚造山带构造演化的关键区域, 但研究内容主要集中于岩浆岩和变质岩的地球化学特征、同位素年代学及构造地质学方面, 由此得出的南天山洋的时代和性质也不大一致, 其或被视为晚志留世— 泥盆纪的弧后盆地(Wang et al., 2011), 或被推测为425± 8, Ma古亚洲洋南缘的小洋盆(尤灵利等, 2006), 或被认为至少在早志留世就已经打开并存在(马中平等, 2006), 也有认为南天山洋发育时间是从震旦纪到泥盆纪、早石炭世进入残留海阶段(左国朝和李绍雄, 2011)等。相比之下, 从沉积地层方面探讨南天山洋的工作就少得多。而沉积地层是洋盆历史的直接地质记录, 放射虫硅质岩又是重要的深水远洋沉积物。文中除论述西南天山放射虫化石的时代分布外, 也从放射虫硅质岩的沉积序列、地层关系及相关的生物古地理变化等来探讨南天山洋的演化。
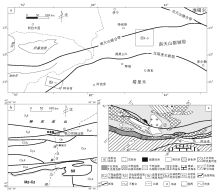
放射虫硅质岩在西南天山广泛分布, 近20年来, 在多处硅质岩中发现放射虫化石。主要分布于3个区域, 自东而西为:库车北的独库公路沿线, 拜城黑英山地区和阿合奇县托什罕河上游地区(图 1-a)。
西南天山地势险要, 交通不便。独库公路是穿越天山南北的交通要道, 也成为地质研究的重要路线。从铁力买提达坂往南, 经库勒湖、库尔干到公路里程976 km的区段沿线, 已有多处硅质岩中发现放射虫化石(刘羽等, 1994; 汤耀庆等, 1995; 王宝瑜等, 1997, 2001; Liu, 2001; Liu and Hao, 2006; 李玮等, 2007)(图1-b)。由于岩层强烈变形, 剖面露头不甚连续, 硅质岩的地层序列并不清楚, 有些地点放射虫组合的时代也有待进一步厘定。
在拜城县北东60 km的黑英山地区(图 1-a), 梅斯布拉克蛇绿混杂岩中含硅质岩和放射虫化石, 尤其是与玄武岩互层的硅质岩对确定蛇绿岩年龄有重要意义, 但其中放射虫化石的时代值得进一步分析。
在南天山西段阿合奇西北的中国— 吉尔吉斯边境地区, 近年来在多处地点发现的放射虫硅质岩, 提供了这一带南天山洋的沉积证据(图 1-c)。但由于位置偏僻、交通不便、地质研究程度低, 该地区放射虫硅质岩的地层归属比较混乱, 放射虫化石的时代分布也有争议。
以下由北而南对独库公路有关地点(图 1-b)的资料逐一评述。
 | 图1 西南天山放射虫硅质岩分布区及地质略图a— 独库公路沿线和阿合奇地区位置; b— 独库公路沿线地质略图(据李玮等, 2007); c— 阿合奇地区地质略图(据Li等, 2002; 李曰俊等, 2005; 李建兵等, 2009资料修编)Fig.1 Distribution of radiolarian cherts in southwestern Tianshan and their geological map |
2.1.1 铁力买提达坂
Liu 和Hao(2006)描述了铁力买提达坂剖面的部分放射虫化石, 主要讨论化石内部结构与分类, 地层叙述比较简略。该剖面底部为厚层灰岩— 礁灰岩, 沉积序列始于灰色— 褐色薄层硅质页岩与薄层灰岩及透镜状灰岩的互层, 其上是红色— 褐绿色硅质页岩夹纯的薄硅质层。放射虫化石产于薄硅质层中, 文中未列出全部化石名单, 仅说根据放射虫化石组合, 含化石层时代大约是晚泥盆世。
2.1.2 库勒湖
库勒湖位于铁力买提达坂西南。库勒湖蛇绿岩是库勒湖— 铁力买提达坂蛇绿岩带的组成部分, 位于库勒湖及其以南的独库公路南段(马中平等, 2006)。自1994年起, 相继报道了库勒湖一带硅质岩中的放射虫化石, 并常被作为库勒湖蛇绿岩的时代依据之一(刘羽等, 1994; 汤耀庆等, 1995; 高俊等, 1995, 1996; 马中平等, 2006)。汤耀庆等(1995)和马中平等(2006)都述及, 在库勒湖蛇绿混杂岩中, 紫红色硅质岩一部分直接覆盖在基性火山熔岩之上, 一部分呈独立岩块存在于混杂带中。但刘羽等(1994)文中并未述及蛇绿岩, 却说观测剖面出露有厚度不等的中— 厚层灰岩, 硅质岩空间上与灰岩相邻, 二者为断层接触。
刘羽等(1994)在9个样品中获得37个放射虫种, 划分出5个放射虫组合带, 时代从晚泥盆世弗拉斯期至早石炭世维宪期。最下部的Astroentactinia crassata-Tetrentactinia braysphaera带(Ⅰ ), 有3个优势种Astroentactinia crassata(Nazarov), A.stellata(Nazarov)和Tetrentactinia braysphaera (Foreman), 据Nazarov和Ormiston(1993)对欧亚和北美古生代放射虫生物地层学的研究, 前2种的延续时代包括整个晚泥盆世, 而第3种仅限于晚泥盆世法门期, 是该时期放射虫组合的2个命名种之一。刘羽等(1994)带Ⅱ 的特征种Entactinia palimbola (Foreman)也是该组合的常见分子。因此, 这两个带可以合并, 时代为晚泥盆世法门期。刘羽等(1994)带Ⅲ 、Ⅳ 和V的放射虫化石呈现早石炭世面貌, 但没有Albaillella, 缺少分带的典型种, 时代大致为早石炭世杜内期— 维宪早期。因此, 刘羽等(1994)发表的库勒湖地区放射虫组合的时代可进一步限定为晚泥盆世法门期至早石炭世维宪早期。
2.1.3 库尔干南
王宝瑜等(1997)的独库公路库尔干道班以南的剖面(从公路里程955 km向南)分为4段。第1段为上志留统科克铁克达坂组灰色薄— 中层灰岩, 构成一个背斜, 厚61 m, 未见底。背斜北翼的灰岩曾发现早泥盆世洛霍考夫期delta带的牙形类化石(王成源和张守安, 1988)。第2段至第4段为下泥盆统阿尔腾柯斯组, 整合覆盖于志留系之上。第2段为灰绿色和紫红色硅质岩夹薄层状灰岩, 厚257 m, 含牙形刺、珊瑚和放射虫等化石。据牙形类化石号码(XI-15-1), 含早泥盆世牙形刺化石的层位在剖面第2段的南部, 靠近第1段地层。放射虫化石号码未标明, 但剖面图上其他的化石号码都在第2段上部, 放射虫化石应来自第2段上部。所列的放射虫化石属种, 有晚泥盆世常见的Entactinia palimbola等, 也有早石炭世常见的Entactinia vulgaris, Triaenosphaera hebes等, 但缺乏化石带的标准分子。
2001年, 王宝瑜等再次论述了该剖面。根据灰岩中分析出的早泥盆世布拉格期sulcata带的牙形刺重要分子Eognathodus linearis, 将阿尔腾柯斯组的时代厘定为早泥盆世洛霍考夫期— 布拉格期, 岩性为灰色薄— 中层状灰岩、硅质灰岩、灰绿色杂砂岩、细砂岩, 并指出在硅质岩中分析出石炭系牙形类化石, 但仍沿用了1997年文中的剖面图和分层, 未将下泥盆统和石炭系分开。
更往南, 在独库公路里程975 km和976 km之间, 李玮等(2007)在阿尔腾柯斯组上段的两块硅质岩样品中, 获得了保存较好的放射虫化石。李玮等(2007)将一块样品(D202— H1)的化石组合时代定为中— 晚泥盆世, 按文中所述, 组合的已知属种以Trilonche属的种最多。而这些种的时限为中泥盆世吉维特期— 晚泥盆世法门期(王玉净等, 2000; Wang et al., 2003)。另一方面, 化石名单中又缺乏法门期放射虫Tetrentactinia braysphaera-Ceratoikiscum famenium组合的特有分子。因此, 该样品的时代范围可缩小为中泥盆世吉维特期— 晚泥盆世弗拉斯期。李玮等(2007)将另一块样品(D203— H1)的化石组合对比为早石炭世的Albaillella indensis带, 但并未发现该带的带化石。而在其化石图版上有Eostylodictya rota, 这是早石炭世维宪初期放射虫Eostylodictya rota带的带化石, 该带位于Albaillella indensis带之上(Braun and Schmidt-Effing, 1993; 王玉净和杨群, 2007)。
综上所述, 独库公路沿线从铁力买提达坂至库尔干南, 由于构造原因, 放射虫硅质岩层多处反复出现, 其时代范围从中泥盆世晚期至早石炭世维宪早期。
黑英山地区梅斯布拉克蛇绿混杂岩中的硅质岩和放射虫化石已经得到比较详细的研究。舒良树等(2007)报道的放射虫化石来自4块样品, 鉴定结果可能相似, 因此都归入一个组合, 时代结论为晚泥盆世法门期— 早石炭世杜内期— 维宪期。按文中鉴定结果所述, 所得到的放射虫主要有6个属, 包括Stigmosphaerostylus, Archocyrtium, Pylentonema, Astroentactinia, Triaenosphaera和Trilonche。其中 Stigmosphaerostylus variospina, Archocyrtium ludicrum, A.venustum, A.delicatum分布于世界各地的晚泥盆世— 早石炭世地层中。Stigmosphaerostylus vulgaris, S.wuppertalensis, S.typical, Pylentonema antigua 等种在欧洲和中国常见于早石炭世杜内期— 维宪期地层中。并含有较多早石炭世杜内期— 维宪期Albaillella paradoxa带的放射虫, 如广泛分布在北美、德国、法国、土耳其、中国、泰国、马来西亚的Astroentactinia cf.mirousi Gourmelon, 广泛分布在法国、中国、泰国的Pylentonema cf.racheboeufi Gourmelon, 广泛分布在德国、法国、中国的Stigmosphaerostylus wuppertalensis (Won)以及广泛分布在德国、中国的Stigmosphaerostylus cf.typical (Won)和Trilonche almae (Won)。
从以上论述看, 这个放射虫化石组合既有出现于晚泥盆世— 早石炭世地层的种, 也有常见于早石炭世地层的种, 而无仅常见于晚泥盆世地层的种。因此, 其时代定为早石炭世更为合适。Albaillella paradoxa 带是早石炭世杜内早期的一个放射虫带, 该地区虽未发现其带化石, 但有多种该带常见化石。舒良树等(2007)文后图版中的放射虫Triaenosphaera cf .hebes Won也是早石炭世早期常见的种。所以, 该组合的时代可能为早石炭世杜内早期, 这也为梅斯布拉克蛇绿岩的时代上限提供了可靠证据。
2002年, Li等首次在本区托什罕河南发现早石炭世放射虫硅质岩(图 1-c, 化石点③)。2005年, 李曰俊等在托什罕河以北又发现早石炭世放射虫硅质岩(图 1-c, 化石点①②)。2009年, 李建兵等在托什罕河上游1:5万区域地质调查中, 在Li等(2002)采样区的西侧, 也发现早石炭世放射虫硅质岩(图 1-c, 化石点④⑤)。Li等(2002)在托什罕河南发现的放射虫化石保存较好, 在托什罕河以北发现的放射虫化石保存较差(李曰俊等, 2005), 但总体面貌相似, 都是以Albaillella为主的化石组合, 包括早石炭世杜内期各放射虫带的重要分子A.paradoxa, A.deflandrei, A.undulata, A.indensis, Stigmosphaerostylus variospina等。前者还有早石炭世维宪初期的重要化石Eostylodictya rota和Belowea valiabilis等(图 2)。因此, 放射虫硅质岩的时代至少涵盖了早石炭世杜内期至维宪早期。李建兵等(2009)称“ 图尤克阿秀组” 第3段由底到顶的硅质岩均富含放射虫化石, 但鉴定名单仅有Entactinia和Entactinisphaera等泥盆纪— 石炭纪常见属名, 不能提供确切时代。而以Entactinia和Entactinisphaera为主, 无石炭纪常见的Albaillella的放射虫化石组合, 则常见于晚泥盆世地层。
Li等(2002)、李曰俊等(2005)文中还报道有晚二叠世的放射虫化石Albaillella excelsa (?)和A.sp.cf.A.excelsa, 但只有个别个体, 且保存不好, 难以作为硅质岩系持续到晚二叠世的确凿证据。值得注意的是, 在阿图什西北约35 km的温古尔地区, 有一套灰岩和硅质岩交互的地层, 硅质岩中含放射虫化石, 灰岩中发现中二叠世栖霞期牙形类化石Neostreptognathodus spp., 为盆地相沉积(姚建新等, 2002)。所以, 在西南天山西段二叠系的某些层位, 可能仍有深水沉积。
前述生物地层学资料表明, 独库公路这段南北宽度近16 km的地层, 包括了从早泥盆世到早石炭世的沉积, 含深灰色粉砂岩、粉砂质泥岩、岩屑长石砂岩、紫红色及深灰色硅质岩夹薄层灰岩、火山角砾岩及凝灰岩等多种岩石类型(李玮等, 2007)。在李玮等(2007)的中泥盆世晚期放射虫化石点以北, 独库公路里程975 km和973 km之间, 陈刚等(2010)还发现硅质岩中有具洋岛型地球化学特征的碱性玄武岩夹层。
由于岩层遭受强烈构造变形, 因此多为被断层分隔的岩片, 缺乏完整的连续剖面, 地层划分比较困难。2004年完成的1:25万喀赞其幅地质报告, 仍将这套地层都划入下泥盆统阿尔腾柯斯组(图 1-b), 这显然不合适。王宝瑜等(2001)曾将阿尔腾柯斯组厘定为下泥盆统的灰岩、硅质灰岩、杂砂岩和细砂岩, 但对剖面中的晚泥盆世— 早石炭世放射虫硅质岩未作进一步论述。1994年, 刘羽等就探讨了独库公路硅质岩与灰岩的关系, 指出灰岩的时代不晚于中泥盆世, 而硅质岩的时代为晚泥盆世— 早石炭世, 二者为断层接触。1995年, 汤耀庆等曾建议用“ 库勒湖组” 之名(引用1993年刘羽硕士论文成果)代表黑英山— 库勒湖一带晚泥盆世— 早石炭世的碳酸盐岩、硅质岩、枕状熔岩和变质橄榄岩的蛇绿岩组合地层。但是, “ 组” 作为最基本的岩石地层单位, 代表着有正常沉积序列一套地层, 而“ 蛇绿岩” 属于特殊岩石地层单位(全国地层委员会, 2001)。因此, “ 库勒湖组” 应仅代表该地区中泥盆世— 早石炭世的硅质岩。
 | 图2 西南天山下石炭统艾克提克组的重要放射虫化石Fig.2 Some important radiolarian taxa from the Lower Carboniferous Aiketik Formation, southwestern Tianshan1, 2— Albaillella undulata Deflandre, × 200; 3~5— Albaillella paradoxa Deflandre, × 220; 6~8— Stigmosphaerostylus variospina (Won), × 150; 9, 10— Eostylodictya rota (Won), × 160; 11, 12— Belowea valiabilis (Ormiston et Lane), × 240 |
独库公路沿线这套地层北接志留系顶统科克铁克达坂组, 南界已接近塔里木北缘的中新生界分布区, 构成了南天山内部一条晚古生代深水沉积构造岩相带。根据上述生物地层资料, 可以识别一个大致的沉积序列:底部为志留系顶统科克铁克达坂组灰色薄— 中层灰岩, 该组在南天山广泛分布, 富含床板珊瑚和层孔虫等化石(王宝瑜等, 2001)。下泥盆统阿尔腾柯斯组灰岩覆盖其上, 与其呈整合接触, 含牙形类而无珊瑚、腕足类等浅水底栖生物化石, 可能为斜坡相沉积。中泥盆统下部地层还未能确定, 中泥盆统上部、上泥盆统和下石炭统下部(至维宪阶下部)都含有放射虫硅质岩, 还含有火山岩和浊积岩等。这是一个从早泥盆世开始逐渐加深的深水沉积序列, 是南天山洋洋盆演化的沉积记录。
Li等(2002)在托什罕河南发现的早石炭世放射虫硅质岩, 原属于艾克提克群(图 1-c, 化石点③)。艾克提克群由深灰色— 灰绿色砂岩、泥岩、灰岩和硅质岩组成, 硅质岩主要发育于下部(南部)(Li et al., 2002)。李曰俊等(2005)在托什罕河以北发现的早石炭世放射虫硅质岩属乌帕塔尔坎群(阿帕达尔康组的同物异名)(图 1-c, 化石点①和②)。李建兵等(2009)发现的早石炭世放射虫硅质岩(图 1-c, 化石点④和⑤)在Li等(2002)采样区的西侧, 他们将其作为新建的“ 图尤克阿秀组” 的第3段。
李建兵等(2009)的“ 图尤克阿秀组” 分为3段, 第1段为火山岩与碳酸盐岩岩石组合, 第2段为复理石建造, 第3段为深海硅质岩建造。但仅图尤克阿秀剖面(图 1-c, 化石点⑤)含这3段地层, 第1段和第2段之间还是断层接触关系。另外2个辅助剖面中, 廓加尔特剖面仅有第1段地层, 与中泥盆统托格买提组灰岩呈断层接触; 开勒特外克剖面(图 1-c, 化石点④)仅有第3段地层, 下不见底, 上覆上石炭统阿依里河组。因此, 这3段地层的关系不清楚, 而且岩性差别很大, 不宜归入同一岩石地层单位。
从地质略图和采样位置看, 李建兵等(2009)的图尤克阿秀剖面和开勒特外克剖面, 与Li等(2002)的采样剖面均位于托什罕河以南的一个近东西向带内(图 1-c, 化石点③、④和⑤)。3个剖面的岩性相似, 均为硅质岩与砂页岩互层。据Li等(2002)的研究区地质略图, 这一近东西向带正是艾克提克群的分布区。蔡士赐等(1999)认为艾克提克群是晚石炭世阿衣里河组的同物异名。艾克提克群上部灰岩曾发现晚石炭世化石, 岩性也与阿衣里河组相似, 可以并入阿衣里河组。但含早石炭世放射虫硅质岩的下部, 显然不应归入阿衣里河组, 或可改称为艾克提克组。
托什罕河以北的地层资料矛盾较多。据李建兵等(2009)的区域地质略图, 这一带出露的地层, 北部主要为下泥盆统阿帕达尔康组, 南部为中泥盆统托格买提组和新建的“ 图尤克阿秀组” 。阿帕达尔康组为一套细碎屑岩夹碳酸盐岩及少许火山岩, 其上的托格买提组由碳酸盐岩、硅质泥岩和千枚岩组成(蔡士赐等, 1999)。陈秀琴和廖卓庭(2006)认为, 东阿赖山西部的托格买提组岩石类型很复杂, 不仅有碳酸盐岩、硅质岩、砂岩, 还有片岩和基性火山岩等, 应另建新组。李建兵等(2009)的廓加尔特剖面仅出露小块 “ 图尤克阿秀组” 第1段地层, 为中基性火山岩与碳酸盐岩互层, 周围是大片的托格买提组地层。因此, “ 图尤克阿秀组” 第1段可能也属于托格买提组。在李建兵等(2009)的区域地质略图上, 李曰俊等(2005)的齐齐尔加纳克苏河采样剖面(图 1-c, 化石点①)位于“ 图尤克阿秀组” 的玄武岩分布区。李曰俊等(2005)提供了硅质岩中的放射虫化石鉴定结果, 无具体的剖面资料, 但认为乌帕塔尔坎群构造变形强烈, 岩性复杂, 主要包括碎屑岩、碳酸盐岩和中— 酸性火山岩, 并分布有超基性岩, 极可能是南天山的蛇绿混杂岩。而李建兵等(2009)认为齐齐尔加纳克苏河剖面的放射虫硅质岩与玄武岩互层。因此, 这里放射虫硅质岩的产出形式, 或是混杂岩中的硅质岩块, 或是火山岩中的硅质岩夹层, 有待进一步研究。
3.3.1 新疆西南天山东段与西段的对比
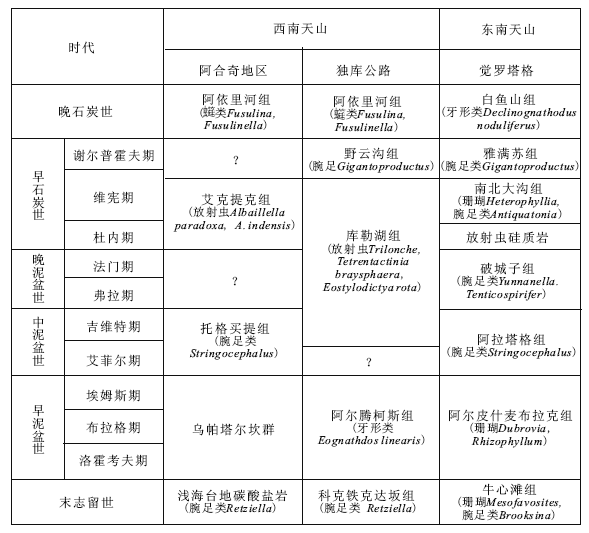
虽然西南天山东段的独库公路沿线地层划分尚较粗略, 西段阿合奇地区地层问题更多, 但出露的地层主要都为泥盆系— 石炭系。阿合奇地区北侧, 吉尔吉斯境内伊塞克湖南和纳伦河南(图 1-a), 志留系罗德洛阶— 普里道利阶含小莱采贝类腕足类化石组合的灰岩广泛分布(Rong et al., 1995), 可与独库公路铁力买提达坂以北的志留系上部地层(图 1-b)对比。独库公路沿线下泥盆统阿尔腾柯斯组含牙形类化石的灰岩在阿合奇地区尚未发现, 阿合奇地区原划作下泥盆统的乌帕塔尔坎群或阿帕达尔康组主要为一套细碎屑岩, 并无可靠的化石作为时代依据。但更往西, 在乌恰县萨瓦亚尔顿金矿区的灰岩和板岩互层中, 曾在下泥盆统洛霍考夫阶中发现牙形类化石 Ancyrodelloides等, 在埃姆斯阶中发现牙形类化石Polygnathus spp.(王成源等, 2000), 岩性和时代都可以和阿尔腾柯斯组对比。阿合奇地区的中泥盆统托格买提组为灰岩和玄武岩互层。虽然托格买提组仅分布于西南天山西段, 但独库公路沿线可能为中泥盆世的硅质岩中也有具洋岛型地球化学特征的碱性玄武岩夹层(陈刚等, 2010), 而典型的洋岛型沉积建造正是玄武岩和灰岩互层。阿合奇地区的艾克提克组硅质岩目前仅有可靠的早石炭世放射虫化石证据, 但不排除含晚泥盆世放射虫化石的可能, 可以和独库公路沿线的库勒湖组硅质岩对比(表 1)。
| 表1 南天山志留纪晚期— 石炭纪地层对比 Table1 Correlation of the Late Silurian to Carboniferous in south Tianshan |
在阿合奇地区, 李建兵等(2009)文中的开勒特外克剖面中, 上石炭统阿依里河组砂岩整合于下石炭统硅质岩系之上, 但未作文字说明。独库公路沿线未见硅质岩的上覆地层。但通常认为, 在哈尔克山南坡, 阿依里河组的下伏地层为野云沟组, 主要为含早石炭世维宪晚期至谢尔普霍夫期的、珊瑚和腕足类等化石的碳酸盐岩系(蔡士赐等, 1999)。阿合奇地区是否存在与野云沟组相当的地层, 有待研究。
3.3.2 与东南天山觉罗塔格地区对比
二者的沉积序列可以对比, 但也有重要差别。觉罗塔格南坡志留系上部牛心滩组主要为富含珊瑚的碳酸盐岩(王宝瑜等, 2001), 可与西南天山的科克铁克达坂组对比。整覆其上的阿尔皮什麦布拉克组为细碎屑岩和灰岩, 除产珊瑚、腕足类等化石外, 还发现早泥盆世牙形类化石(蔡士赐等, 1999; 廖卫华和弗朗西斯科, 2004)。其时代与西南天山的阿尔腾柯斯组相当, 但属于浅海台地沉积。中泥盆统阿拉塔格组为碎屑岩夹碳酸盐岩和火山岩, 产中泥盆世腕足类和珊瑚化石。上泥盆统破城子组中下部以细碎屑岩和灰岩为主, 上部为中酸性火山岩和火山碎屑岩, 含晚泥盆世腕足类化石, 仍是浅海沉积(蔡士赐等, 1999)。
石炭系在觉罗塔格东段广泛分布, 根据1:5万区调资料, 左国朝等(2006)分析了该区的构造地层演化。那里沿北纬42° 分布的康古尔塔格断裂带有洋壳断片出露, 包括硅质岩、凝灰岩、玄武岩和辉长岩等, 硅质岩中含石炭纪放射虫化石。洋壳断片向南逆掩至下石炭统南北大沟组之上, 该组为含杜内晚期或维宪早期珊瑚和腕足类化石的碳酸盐岩。逆掩其上的放射虫硅质岩时代应较其早, 即可能为杜内期, 可与西南天山的库勒湖组放射虫硅质岩对比。而平行不整合或低角度不整合于南北大沟组之上的雅满苏组, 由各种火山熔岩和火山碎屑岩及生物碎屑灰岩组成, 为火山岛弧带沉积, 含维宪期— 谢尔普霍夫期珊瑚和腕足类等化石。在西南天山, 与其时代相当的野云沟组为不含火山岩的浅海碳酸盐沉积, 属于稳定构造环境产物。
上述情况表明, 独库公路库勒湖— 库尔干一带、拜城黑英山和阿合奇西北的放射虫硅质岩的时代和构造位置都可对比, 构成西南天山一条泥盆纪— 石炭纪深水沉积带。空间上, 它的南面是现今的塔里木地块北缘, 北面都是一套志留纪晚期沉积。据王宝瑜等(2001), 这套志留系以碳酸盐岩为主, 包括下部的伊契克巴什组和上部的科克铁克达坂组, 在哈尔克山南坡广泛分布, 延展至吉尔吉斯境内, 即为上述伊塞克湖南的志留系。塔里木志留系以滨海相红层为主, 晚期甚至可能隆起成陆(赵文金等, 2009)。而北面的志留系碳酸盐岩富含床板珊瑚、层孔虫、腕足类等底栖动物化石, 前二者还多形成礁体, 其特征的小莱采贝类腕足动物是志留纪晚期生活在近岸、正常浅海(平均浪基面以上)的底栖动物群(戎嘉余等, 1994)。因此, 这套志留纪晚期沉积可能代表当时塔里木古陆边缘的浅海陆架环境, 西南天山地区在志留纪可能为大塔里木的一部分。
科克铁克达坂组之上, 即为下泥盆统阿尔腾柯斯组, 在西南天山多处报道二者为整合接触关系, 包括上述独库公路库勒湖北剖面和库尔干南剖面(王宝瑜等, 2001)。含早泥盆世牙形类化石的阿尔腾柯斯组仍以灰岩为主, 但已缺少珊瑚和腕足类浅海生物, 可能为斜坡相沉积。中泥盆统托格买提组含大量火山熔岩, 显示地壳的张裂。中泥盆世晚期出现放射虫硅质岩, 晚泥盆世— 早石炭世早期放射虫硅质岩发育, 显示了深海盆地的特征。晚泥盆世晚期— 早石炭世早期的蛇绿岩, 则是南天山洋壳的标志。因此, 西南天山志留纪至早石炭世的沉积序列, 指示了南天山洋从浅海逐渐加深直至出现洋壳的演化过程, 表明南天山洋是塔里木边缘浅海陆架裂解产生的小洋盆。从上述东南天山的资料看来, 康古尔塔格缝合带代表的康古尔小洋盆(左国朝等, 2006)早石炭世才出现。因此, 南天山洋可能是从西向东发展。
从大地构造位置看, 西南天山之北为中天山伊宁地块, 伊宁地块早、中奥陶世的笔石动物群与澳大利亚关系密切, 迄今为止澳大利亚典型的Oncograptus upsilon等在中国只见于伊宁地块, 伊宁地块的Isograptus动物群也与澳大利亚一致, 是典型的太平洋大区的笔石动物群(陈旭等, 1998)。晚奥陶世地层在伊宁地块西延的哈萨克斯坦南部Chu-Ili地区研究较详, 称Dulankara 组, 分为3段, 下部 Otar段的腕足动物群与华南关系最密切。其上是Degeres段和Akkol段, 前者以细砂岩和粉砂岩为主, 富含腕足类化石, 后者下部为瘤状灰岩, 含藻类和床板珊瑚化石, 上部泥岩, 含笔石化石。岩性和化石组合表明该组形成于浅海陆架沉积环境(Popov et al., 2000; Popov and Cocks, 2006)。陈旭等(2006)指出, 哈萨克斯坦南部Chu-Ili地区上奥陶统赫南特阶地层发育良好, 其中的笔石和壳相动物群序列和湖北宜昌王家湾北剖面(赫南特阶全球层型剖面)以及扬子区其他剖面均为一致。
据Rong等(1995)对志留纪罗德洛世— 普里道利世亚洲— 澳洲腕足动物生物地理区的研究, 北方的蒙古— 鄂霍茨克区以图瓦贝为特征, 南方的中— 澳区以小莱采贝为特征, 这是2个浅水底栖动物群落, 前者分布于西伯利亚板块南缘, 后者分布于扬子台地西缘、塔里木西北缘(即上述西南天山地区)、华北地块北缘和澳洲东缘。
上述生物古地理证据表明, 西南天山和伊宁地块在奥陶纪— 志留纪均和华南及澳大利亚关系密切, 塔里木和华南地块的志留系地层序列和岩性特征非常相似, 并有共同的鱼类化石组合。因而, 赵文金等(2009)在其志留纪古地理图中, 将二者连接为塔里木— 华南联合板块。朱日祥等(1998)根据古地磁资料指出, 扬子与塔里木在早古生代所处纬度相同, 两地块记录的磁偏角变化相似, 说明两地块在早古生代是相连的。早泥盆世晚期华南地块出现多条裂陷带, 昌宁— 孟连带、哀牢山带和南盘江地区的深水沉积都从此时开始发育, 到晚泥盆世— 早石炭世时形成一系列古特提斯分支洋盆, 放射虫硅质岩广泛分布(吴浩若, 2003)。这与上述南天山洋的发生、发展过程一致。古生物地理和构造演化的一致性, 表明南天山洋也是古特提斯洋的一个分支, 不属古亚洲洋范围。
早石炭世生物古地理证据也支持这个论断。廖卓庭等(2011)指出, 艾比湖— 依连哈比尔尕— 星星峡断裂以北的“ 准噶尔— 北天山区” 与南邻的整个中、南天山构造单元, 由于大地构造属性和沉积体系各不相同, 历来两者之间的沉积组合与生物组合存在截然不同的清晰界线。中、南天山地区, 广泛分布有以大长身贝为代表的早石炭世腕足类化石群; 北天山的依连哈比尔尕山、博格达山、哈尔力克山及其以北的广大准噶尔地区, 则至今从未真正发现过这类腕足动物类化石。而大长身贝类腕足动物化石正是特提斯生物地理区系的专属性分子。
华南的古特提斯洋在早石炭世后持续发展, 可延续至中晚三叠世, 但南天山洋在晚石炭世即已消失。舒良树等(2007)在黑英山梅什布拉克牧场西侧沟谷中, 发现中— 下石炭统砂岩— 粉砂岩(无时代依据)连同中石炭统卡拉苏组生物碎屑灰岩均角度不整合于蛇绿混杂岩之上。卡拉苏组为上石炭统(石炭系二分)阿衣里河组的同物异名(蔡士赐等, 1999)。按上述李建兵等(2009)的资料, 则阿依里河组整合于下石炭统放射虫硅质岩之上。不论其间接触关系如何, 西南天山上石炭统, 包括阿依里河组和喀拉治尔加组, 都是浅海沉积。之后即有局部的二叠纪深水相沉积发育(姚建新等, 2002), 属台内盆地沉积, 而非洋盆沉积。
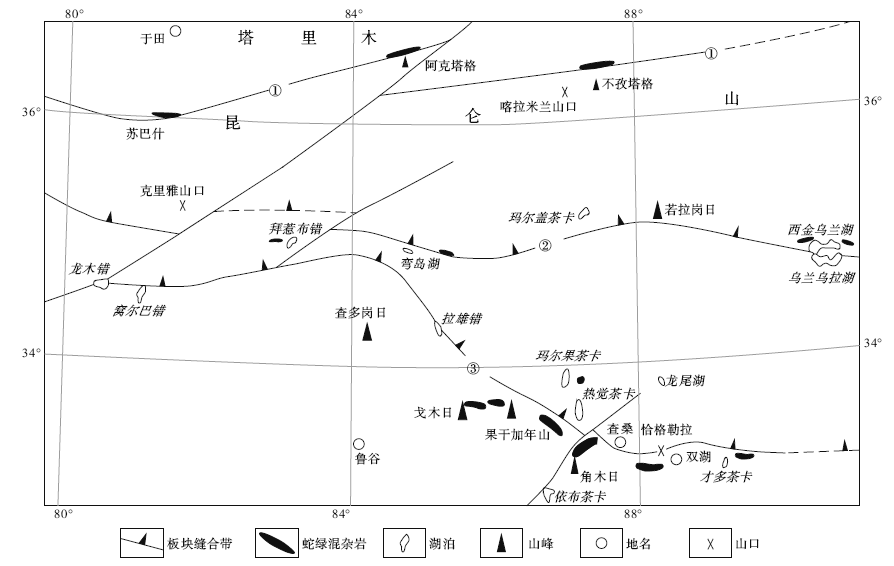 | 图3 塔里木南的古特提斯蛇绿岩分布Fig.3 Distribution of Palaeo-Tethyan ophiolite belts to south of Tarim①— 康西瓦— 木孜塔格缝合带; ②— 西金乌兰— 金沙江缝合带; ③— 龙目错— 双湖缝合带 |
南天山洋的消失远早于华南的古特提斯洋盆, 应与其位置有关。西南天山的蛇绿岩带为古特提斯最北的缝合带, 离古亚洲域的北天山和准噶尔最近。北天山巴音沟蛇绿混杂带与西准噶尔达拉布特蛇绿混杂带关系密切, 二者放射虫硅质岩时代均为晚泥盆世— 早石炭世, 同属于准噶尔边缘最年轻的缝合带, 其上的晚石炭世浊积岩则代表了深海盆地的充填消失过程(廖卓庭等, 1993, 2011; 吴浩若等, 1993)。因此, 南天山洋与准噶尔— 北天山区的古亚洲洋从早石炭世晚期至晚石炭世的消亡过程大致同步, 南面的南天山洋稍早于北面的北天山— 准噶尔洋。古地磁资料表明, 塔里木板块和西伯利亚板块在二叠纪前对接, 而泥盆纪至晚二叠世西伯利亚板块基本保持稳定(朱日祥等, 1998)。因而, 二者的对接主要由塔里木的北移完成, 塔里木的北移则与古特提斯的扩张相关。
中国的古特提斯蛇绿岩带众多, 在塔里木南面的青藏高原北部, 也有多条古特提斯蛇绿岩带(图 3)。在塔里木南侧的昆仑造山带, 有康西瓦— 木孜塔格蛇绿混杂岩带。兰朝利等(2002)在新疆东昆仑木孜塔格蛇绿混杂岩中找到早石炭世放射虫硅质岩; 尹福光等(2002)在西昆仑阿克塔格玄武岩之上的硅质岩中发现早石炭世放射虫化石。韩芳林等(2004)、计文化等(2004)在西昆仑于田南部的苏巴什蛇绿混杂岩中, 发现石炭纪和中二叠世的放射虫硅质岩, 认为其向西接康西瓦构造带, 向东通过木孜塔格蛇绿混杂岩带与阿尼玛卿蛇绿岩带相连。昆仑带南面, 青海可可西里的西金乌兰— 冈齐曲蛇绿混杂岩, 含早石炭世和早二叠世放射虫硅质岩, 一般认为是金沙江蛇绿岩带的西延(边千韬等, 1997)。近年的青藏高原地质调查中, 在可可西里以西的藏北羌塘腹地, 多个1:25万图幅都发现了西金乌兰— 金沙江带的踪迹。如杨子江和李咸阳(2006)在岗扎日幅、玉帽山幅和玛尔盖茶卡湖幅东西延伸的若拉岗日构造带(认为是西金乌兰— 金沙江带的西延)发现晚二叠世— 中三叠世的放射虫化石。陆济璞等(2006)在查多岗日幅若拉岗日构造带发现弯岛湖蛇绿混杂岩, 其中含中三叠世晚期— 晚三叠世早期放射虫硅质岩。张振福等(2004)在黑石北湖幅的碎石山、拜惹布错一带发现蛇绿混杂岩, 将西金乌兰— 金沙江带向西推至东经83° 附近。再往南, 羌塘中部双湖地区的构造混杂岩(含蛇绿混杂岩和岛弧火山岩)中发现晚泥盆世和晚二叠世放射虫硅质岩(朱同兴等, 2006)。上述情况表明, 塔里木南面的多个古特提斯分支洋盆依然持续至中二叠世甚至晚三叠世。有理由认为, 正是它们在早石炭世及其后的继续扩张, 使塔里木北移, 导致南天山洋和准噶尔— 北天山区的古亚洲洋在早石炭世晚期和晚石炭世相继消亡。
1)泥盆纪— 石炭纪放射虫硅质岩在西南天山广泛分布。从东部的独库公路沿线到西部的阿合奇中— 吉边境, 构成一条深水沉积带。独库公路沿线已发现的中泥盆世晚期至早石炭世维宪早期的放射虫硅质岩, 可用“ 库勒湖组” 统称。
2)生物地层和沉积相资料表明, 从志留系顶统科克铁克达坂组经下泥盆统阿尔腾柯斯组到库勒湖组, 记录了南天山从浅海逐渐加深直至出现洋壳的演化过程。
3)古生物地理证据表明, 西南天山和伊宁地块在奥陶纪— 志留纪与华南及塔里木关系密切, 西南天山志留纪晚期沉积可能代表当时塔里木古陆边缘的浅海陆架环境, 南天山洋是塔里木边缘浅海陆架从早泥盆世开始裂解产生的小洋盆。
4)东南天山泥盆系以浅海相沉积为主, 康古尔塔格缝合带仅有早石炭世放射虫硅质岩。因此, 南天山洋可能是自西向东打开。
5)构造古地理和生物古地理研究表明, 南天山洋是古特提斯洋的分支洋盆, 不属古亚洲洋范围。
6)塔里木以南的古特提斯洋分支洋盆, 在早石炭世及其后的继续扩张, 使塔里木北移, 导致南天山洋和准噶尔— 北天山区的古亚洲洋, 在早石炭世晚期和晚石炭世相继消亡。
致谢 研究生高剑、郭春涛清绘部分图件初稿, 谨此致谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|