

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
珠江口黄茅海河口湾表层沉积物中有孔虫和介形虫的分布及其影响因素*
[吴洁1  , 刘春莲
, 刘春莲1, 2 , 贾良文3 , 杨婷婷1 , 张素青1 , 殷鉴1 ]
, 刘春莲, 贾良文|
|

通讯作者简介: 刘春莲,女,1956年生,毕业于南京大学,在德国Würzburg大学获得博士学位。现为中山大学地球科学系教授,博士生导师。目前主要从事新生代古环境学研究。E-mail:eeslcl@mail.sysu.edu.cn。
第一作者简介:吴洁,女,1985年生,中德(中山大学—埃朗根纽伦堡大学)联合培养博士研究生,专业为第四纪古环境学。E-mail:wujie_109@126.com。
珠江口黄茅海河口湾的表层沉积物100个站位样品中共发现有孔虫21属36种、介形虫16属20种。优势种组合分别为Ammonia beccarii-Quinqueloculina akneriana rotunda-Cavarotalia annectens-Elphidium advenum和Neomonoceratina delicata-Sinocytheridea impressa-Bicornucythere bisanensis。有孔虫及介形虫的丰度和分异度均表现出“北低南高”,即口门低、向海高的特点。样品中有孔虫和介形虫属种的去趋势对应分析( DCA)表明,盐度是影响该区有孔虫和介形虫分布的最重要因素,其次是水动力条件,水深对有孔虫和介形虫的分布也有一定影响,但并非主要控制性因素。由于受径流影响较大,河口湾北部的水体盐度较低、径流动力较强,不利于有孔虫和介形虫的生存,导致其丰度及分异度均较低。河口湾南部水体盐度升高,潮流动力较强,同时受众多岛屿屏障作用,环境较为稳定,有孔虫和介形虫的丰度和分异度均迅速上升。
About the corresponding author: Liu Chunlian,born in 1956,is a professor of the Department of Earth Sciences of Sun Yat-sen University,with a Ph.D.degree obtained from the University Würzburg,Germany.She is currently engaged in Cenozoic palaeoenvironment.E-mail:eeslcl@mail.sysu.edu.cn.
About the first author: Wu Jie,born in 1985,is a joint Ph.D.candidate of the Department of Earth Sciences of Sun Yat-sen University and Universität Erlangen-Nürnberg. She is mainly engaged in Quaternary palaeoenvironment. E-mail:wujie_109@126.com.
Foraminiferal and ostracod assemblages in 100 surface sediment samples collected from the Huangmao Bay of the Pearl River Estuary were analyzed,leading to the identification of a total of 36 foraminiferal species belonging to 21 genera and 20 ostracod species belonging to 16 genera.The dominant species of the two faunas with a relative abundance higher than 10% are Ammonia beccarii-Quinqueloculina akneriana rotunda-Cavarotalia annectens-Elphidium advenum and Neomonoceratina delicata-Sinocytheridea impressa-Bicornucythere bisanensis, respectively.The abundance and diversity of foraminifers and ostracods increase gradually seawards across the estuary.The Detrended Correspondence Analysis of foraminiferal and ostracod species suggested that the most important factor affecting the distribution of foraminifers and ostracods in the surface sediments of the Huangmao Bay is water salinity,followed by hydrodynamics.Besides,water depth has also a certain effect on the faunal distribution but not the main controlling factor.As a result of freshwater influencing,the environment of northern estuary is of low salinity and high water energy,which is not conducive to survival of foraminifers and ostracods,leading to a low abundance and diversity of the two faunas.On the other hand,because of the salinity increasing and the islands acting as the barrier to turbulent sedimentary environment,the abundance and diversity of foraminifers and ostracods increase rapidly in the southern estuary.
河口区是全新世最大海侵以来快速沉积的海陆过渡地带(Nash et al., 2010; 刘春莲等, 2011), 对气候和环境变化非常敏感。由于受到径流及潮流的共同作用, 河口区的理化环境复杂多变。而广泛生活其中的微体生物— — 有孔虫及介形虫, 因其对环境变化非常敏感, 且壳体能够在沉积物中大量保存, 是河口海洋环境变化的良好载体(李涛等, 2011)。对现代河口区表层沉积物中有孔虫及介形虫分布的研究, 不仅可以了解微体动物种群分布与沉积环境之间的关系(Elliott et al., 1966; Debenay and Luan, 2006; Frezza and Carboni, 2009), 而且可为利用微体化石类群重建河口三角洲地区第四纪古环境演化提供可靠依据(Tibert and Scott, 1999; Mendes et al., 2004; 刘春莲等, 2008)。
对珠江口表层沉积物中有孔虫和介形虫的研究始于20世纪80年代, 李淑鸾(1985a, 1985b)和汪品先等(1986)较早对整个珠江口水域中有孔虫和介形虫的分布展开了研究, 随后其他学者也陆续在珠江口地区进行了一些有孔虫种群分析(陈木宏, 1989; 罗宪林等, 2001; 李涛等, 2011)。但由于珠江口环境复杂多变, 且大部分研究的采样密度较小、研究范围跨度较大, 无法对小尺度范围内的微体生物分布影响因素做出精准判别。作者对珠江口西南部黄茅海河口湾约500 km2水域内的100个站位表层沉积物样品中的有孔虫和介形虫进行了分析, 并且尝试利用去趋势对应分析方法(DCA)对研究区有孔虫和介形虫埋藏群与3个主要环境影响因子(盐度、水深和沉积物的砂含量)之间的相关性进行了定量分析, 其成果可为利用微体动物群重建珠江三角洲地区第四纪古环境所借鉴。
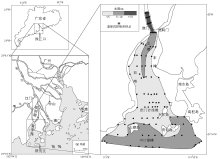
 | 图1 珠江口位置及黄茅海河口湾表层沉积物样品采样点和水深分布Fig.1 Maps of the Pearl River Estuary, sampling locations and water depth across the Huangmao Bay |
黄茅海位于珠江口西南侧的河口湾(图 1), 其北部湾顶为崖门, 入注河口湾的主要河流有两条, 一为珠江8个大分流水道之一的崖门水道, 接纳支流潭江汇入后的银洲湖段, 其水深较大, 是典型的溺谷河口(韦惺和吴晓星, 2011)。二为另一珠江8个大分流水道之一的虎跳门水道。黄茅海作为崖门水道和虎跳门水道的共同出海口, 是个喇叭形的河口湾, 外连南海, 水域面积约543 km2(吴超羽, 1995)。该水域岛屿众多, 滩与槽交错。总体地形格局是“ 三滩两槽” , 三滩指西滩、东滩和拦门沙浅滩, 水深5, m以下; 两槽为北部主槽和大襟岛与荷包岛之间的中口深槽, 水深一般大于5 m, 北部主槽的流水动力作用较强, 泥沙不易落淤(吴超羽, 1995; 贾良文等, 2012)。
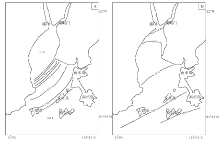
通常喇叭型河口湾的潮差较大, 潮流动力较强。黄茅海河口湾的潮汐为不规则半日潮, 潮差分布由湾口向湾顶递减, 平均潮差由荷包岛的1.34 m逐渐减小到湾顶崖南的1.25 m(吴创收等, 2010)。径流是黄茅海河口最重要的动力之一(吴超羽, 1995)。河流带来的泥沙是塑造河口的最活跃物质。崖门水道年平均径流量为196× 104 m3, 年平均输沙量为363× 104 m3。虎跳门水道年平均径流量及输沙量分别为202× 104 m3和509× 104 m3(陈耀泰, 1992)。由于受到热带海洋性季风气候影响, 珠江口地区的降雨量和径流量呈现明显的季节性(Ji et al., 2011)。受此影响, 河口湾水域盐度值也呈现明显的季节性变化(Dong et al., 2004)。从图2(黄方等, 1994)可见, 黄茅海河口湾水体盐度平面分布趋势大致呈现出自湾顶向湾口逐渐增大的趋势。河口湾表层水体温度夏高冬略低, 据大万山岛海洋站(113° 43'E, 21° 56'N)记录, 月平均温度7月最高, 为28.9, ℃, 2月最低, 为16.5, ℃(赵焕庭, 1990)。
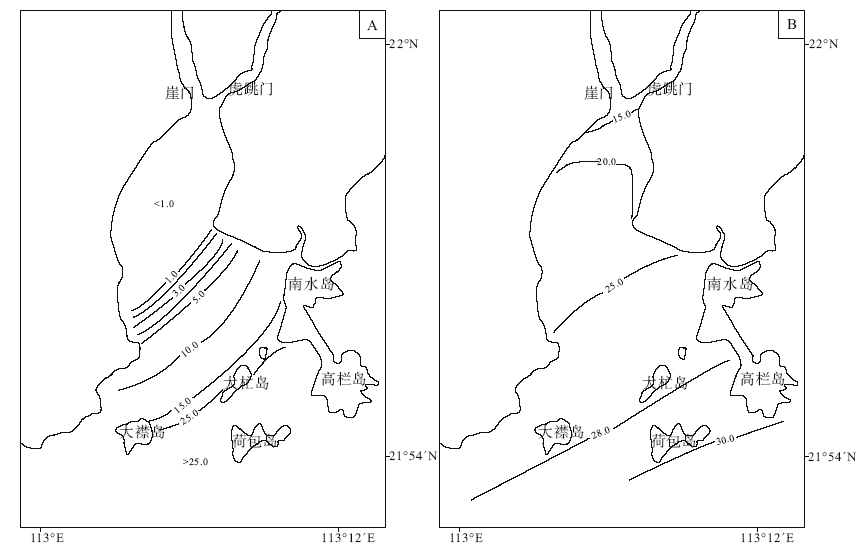 | 图2 黄茅海河口湾洪季(A)及枯季(B)底层盐度(‰ )平面分布(据黄芳等, 1994)Fig.2 Bottom water salinity(‰ )distribution in the Huangmao Bay during wet season(A) and during dry season(B)(after Huang et al., 1994) |
研究样品采集于2011年4月, 使用抓斗取样器在100个站位采集表层沉积物样品, 同时测得采集点水深数据并绘制成图(图 1)。沉积物样品在60, ℃左右的温度下烘干, 称取50 g左右干样在烧杯中用水浸泡24~48 h, 随后在沙浴中适当加热, 使其充分散开。然后用0.063 mm孔径过筛后再次烘干并统计重量。在实体显微镜下进行挑样, 鉴定分析。定量统计(以100 g干样计算)出每个样品中有孔虫和介形虫的总丰度(个体总数)、简单分异度(种数)以及复合分异度H(S)。其中复合分异度H(S)的计算方法如下:
采用Surfer 8.0软件对黄芳等(1994)测得的黄茅海河口湾洪季底层水体平均盐度值进行投影, 得到每个采样点的夏季平均底层盐度, 用于随后的定量分析。采用CANOCO 4.5软件对所有样品中的有孔虫和介形虫优势种(占全群比例高于2%的种)进行DCA排序。
沉积物粒径分布是物质来源、沉积区水动力条件等的综合反映。细粒沉积物较粗粒沉积物移动速度更快, 在水动力较强处沉积物较粗, 水动力条件较弱处沉积物较细(彭晓彤等, 2004)。因此可以通过沉积物粒径特征判别沉积环境的水动力条件。对每个表层沉积物的砂含量(粒径不小于0.063 mm的干样占全部干样的重量百分比)进行了统计, 得出研究区粒径分布统计图(图 3)。表层沉积物中砂含量最高的区域位于河口湾北部主槽, 其次为中口深槽和拦门沙浅滩, 河口湾东西两侧的浅滩砂含量最低。以上砂含量的区域性分布表明研究区不同水域水动力条件具有明显的差异。崖门水道及北部主槽水动力作用较强, 泥沙不易落淤, 此外由于河口湾“ 东进西出” 的近岸水流格局(杨雪舞等, 1997), 研究区东南部潮流动力较强, 表层沉积物的砂含量也较高; 由于河口湾边界及岛屿对水流的缓冲作用, 河口湾东西两侧浅滩和大襟岛— 大杧岛— 荷包岛水域水流动力作用较弱, 砂含量相对偏小; 拦门沙浅滩和中口深槽区砂含量居中, 这是由于这2个区域虽然分别受到来自北部的径流和来自南部的潮流的冲击, 但总体处于众岛屿的屏障下。
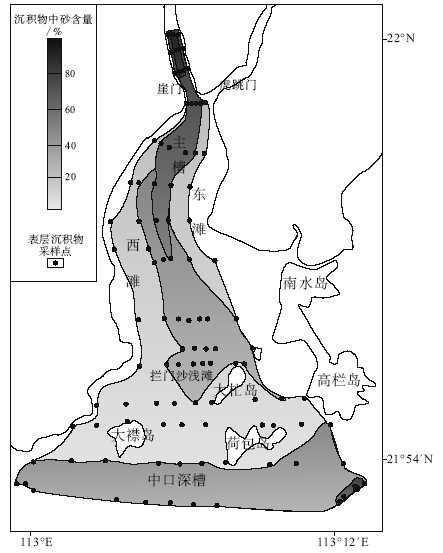 | 图3 黄茅海河口湾表层沉积样砂含量分布Fig.3 Range of sand content for each surface sediment sample in the Huangmao Bay |
除崖门口附近零星站位中发现的微体动物壳体较小且部分磨损外, 黄茅海河口湾大部分站位中的有孔虫及介形虫壳体大小不一, 磨损壳体极少, 介形虫左、右壳比例基本相当, 且优势度较高。以上特征表明研究区有孔虫和介形虫以原地埋藏为主, 同样的情况也出现在珠江三角洲第四系钻孔样品中(刘春莲等, 2008)。
黄茅海河口湾100个站位的表层沉积物样品中共鉴定统计出有孔虫21属36种、介形虫16属20种。其中有孔虫优势属种(占全群比例高于2%)主要有Ammonia beccarii(占全群比例22.6%)、Cavarotalia annectens(19.4%)、Quinqueloculina akneriana rotunda(19.3%)、Elphidium advenum(10.9%)、Elphidium hispidulum(8.5%)、Protelphidium granosum(3.7%)、Spiroloculina robusta(2.7%)、Rosalina bradyi(2.5%)、Quinqueloculina seminulangulata(2.3%)、Quinqueloculina contorta(2.1%)和Florilus decorus(2.1%)。以上优势种共占研究区有孔虫群落的96%, 其中Ammonia beccarii、Cavarotalia annectens、Quinqueloculina akneriana rotunda和Elphidium advenum 4个种所占比例总和高达72%。以上4个种在珠江三角洲第四系钻孔样品中所占比例总和也达到70%。Ammonia beccarii是世界上分布范围最广的广盐性沿岸浅水种, 常见于河口、潟湖等20 m水深以内的海陆过渡相环境(汪品先等, 1988), 在1‰ ~40‰ 的盐度范围内均能存活(Debenay et al., 1998)。Cavarotalia annectens、Quinqueloculina akneriana rotunda和Elphidium advenum较适宜的盐度范围为10‰ ~33‰ (汪品先等, 1988)。
研究区底栖有孔虫占绝对优势, 浮游有孔虫仅占全部有孔虫群的0.06%。有孔虫壳质分为玻璃质、瓷质和胶结质。本区玻璃质壳有孔虫无论丰度和分异度都占优(占全群比例为69.2%), 瓷质壳次之(27.1%), 胶结壳所占比例最小(3.7%)。
黄茅海河口湾介形虫优势属种主要有Neomonoceratina delicata(29.8%)、Sinocytheridea impressa(25.0%)、Bicornucythere bisanensis(17.0%)、Alocopocythere kendengensis(7.4%)、Sinocythere sinensis(4.7%)、Pistocythereis bradyformis(3.4%)、Stigmatocythere bona(3.0%)、Keijella hodgii(2.9%)和Neosinocythere elongata(2.1%), 共占研究区介形虫群落的95%。Neomonoceratina delicata、Sinocytheridea impressa和Bicornucythere bisanensis 3个种比例总和达77%。上述3个种在珠江三角洲第四系钻孔样中所占比例总和达到52%。Neomonoceratina delicata为典型的海陆过渡相滨岸浅海种, 是近岸河口的代表属种, 其适应的盐度为13‰ ~34‰ (侯祐堂等, 2002)。Sinocytheridea impressa为广温、广盐性浅水种, 在1‰ 至正常海水中均能生存, 是中国近岸河口环境中分布最为广泛的介形虫(赵泉鸿和汪品先, 1988)。Bicornucythere bisanensis是典型的广盐性喜半咸水种, 适宜其生存的盐度范围为27‰ ~34‰ (赵泉鸿等, 1986)。
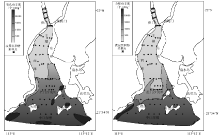
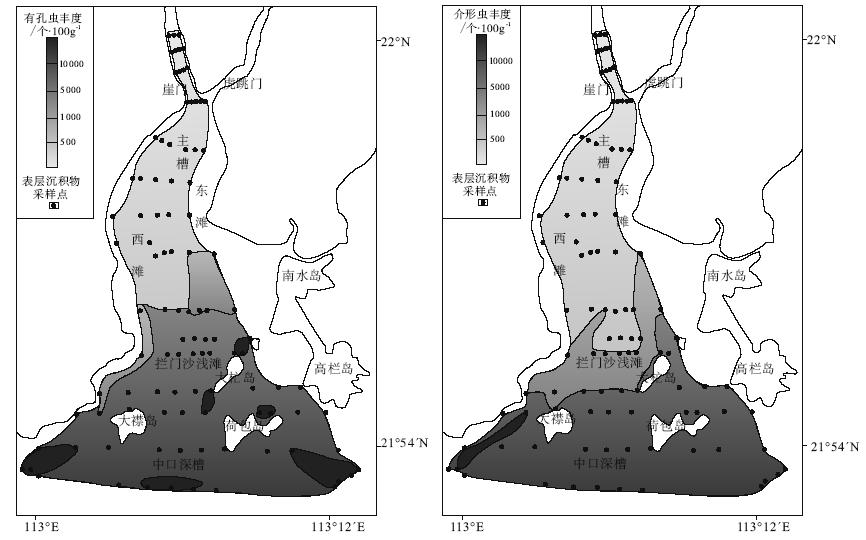 | 图4 黄茅海河口湾表层沉积物中有孔虫和介形虫丰度分布Fig.4 Distribution of foraminiferal and ostracod abundance in surface sediments of the Huangmao Bay |
研究区内有孔虫和介形虫的丰度变化很大(图 4)。在每100 g干样中, 河口湾北部的有孔虫及介形虫个体数量非常小, 一般在500枚以下, 且有孔虫及介形虫壳体个体较小, 说明该区域的环境不利于生物个体发育。河口湾顶部几乎没有发现任何微体动物壳体, 这可能与该区域受径流影响较大、径流动力较强有关。而在河口湾中部及南部, 有孔虫及介形虫数量明显增多, 在拦门沙浅滩以南的水域微体动物个体总数大都在1000枚以上。以上微体动物壳体数量的显著变化表明河口湾南部水域环境较北部更适合有孔虫及介形虫的生存。
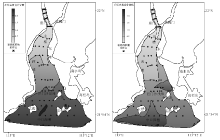
分异度方面, 河口湾内有孔虫简单分异度(种数)在0~14之间变化, 复合分异度在0~2.13之间变化。介形虫简单分异度及复合分异度变化范围分别为0~12和0~2.03。研究区有孔虫和介形虫的分异度变化趋势基本同丰度变化一致, 均表现为“ 北低南高” 的特点(图 5, 图6), 即南部湾口拦门沙浅滩及其以南水域微体动物种类明显多于其北部湾顶。这可能是由于湾口水体盐度环境较湾顶稳定, 能适应此环境的种类较多。
为了探究黄茅海河口湾各环境变量(盐度、水深和沉积物的砂含量)对微体动物分布的影响, 作者对表层沉积物样本中的有孔虫和介形虫进行了去趋势对应分析(DCA)。DCA是目前国际上应用较为广泛的一种排序方法, 主要用来研究生物与环境因子之间的相关性, 最早由Hill和Gauch(1980)将此方法应用到生态学领域, 近年来逐渐被部分第四纪地质与环境工作者用来定量分析近岸环境, 尤其是河口三角洲环境的生物分布与环境因子之间的相关性(Cearreta et al., 2008; Kemp et al., 2009; Zong et al., 2010), 以达到定量重建第四纪海平面变化的目的。DCA排序能够反映群落在主要排序轴上的梯度分布, 通过环境变量与排序轴的相关性系数就可以确定影响群落分布格局的主要环境因素。
 | 图5 黄茅海河口湾表层沉积物中有孔虫和介形虫简单分异度分布Fig.5 Distribution of foraminiferal and ostracod species number in surface sediments of the Huangmao Bay |
 | 图6 黄茅海河口湾表层沉积物中有孔虫和介形虫复合分异度分布Fig.6 Distribution of foraminiferal and ostracod compound diversity H(S)in surface sediments of the Huangmao Bay |
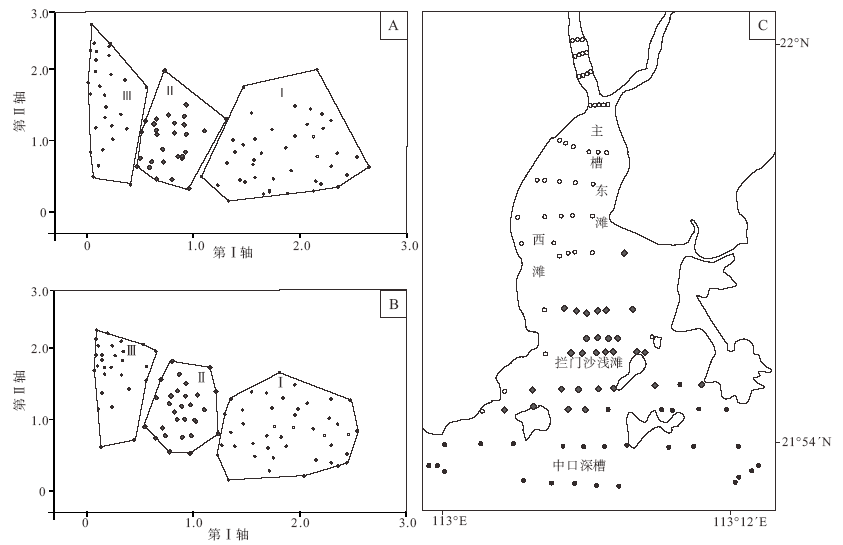 | 图7 黄茅海河口湾表层沉积物中有孔虫(A)及介形虫(B)数据去趋势对应分析(DCA)排序结果Fig.7 Ordination results of Detrended Correspondence Analysis(DCA) of foraminiferal(A) and ostracod(B) data in surface sediments of the Huangmao Bay |
黄茅海河口湾表层沉积物样品中有孔虫和介形虫的DCA排序结果(图7)显示, 所有样本在第Ⅰ 排序轴上得到了非常好的分化, 根据分析结果可将100个采样站位划分为3大区域。区域Ⅰ 主要为崖门水道及北部主槽; 区域Ⅱ 包括东西两侧浅滩及拦门沙部分浅滩; 区域Ⅲ 主要包括部分拦门沙浅滩及其以南水域。DCA同时提供了有孔虫和介形虫属种与代表各环境变量的排序轴之间的相关性系数(表 1)。由表1可知, 第Ⅰ 轴所代表的环境变量与有孔虫属种分布的相关性最高(0.657), 其他3轴依次为0.337、0.282和0.141; 与介形虫属种分布相关性最高的同样为第Ⅰ 轴所代表的环境变量(0.586)。说明第Ⅰ 轴所代表的环境变量是影响研究区有孔虫和介形虫分布的最主要因素。
| 表1 表层沉积物样品有孔虫和介形虫属种去趋势对应分析结果统计 Table1 Statistics summary of Detrended Correspondence Analysis of foraminifers and ostracods in surface sediments |
| 表2 环境因子与有孔虫属种排序轴之间的相关系数 Table2 Correlation coefficients between environmental factors and foraminiferal species ordination axes |
通过环境因子与排序轴的相关性分析(表 2)可以找出与排序轴显著相关的环境因子, 因为各相关系数表示排序轴与真实环境梯度之间的相关程度, 而这些环境因子就是影响群落分布格局的主要因素。影响有孔虫分布的3个环境因子中与第Ⅰ 轴相关性最高的是盐度, 呈显著负相关, 相关系数为-0.6498; 与第Ⅱ 轴相关性最高的是沉积物的砂含量, 成负相关, 相关系数为-0.2277; 与第Ⅲ 轴相关性最高的是水深, 成正相关, 相关系数为0.1995。影响介形虫分布的环境因子与前3轴相关性最强的依次为盐度(-0.4913)、沉积物的砂含量(-0.3198)和水深(0.1969)(表 3)。
| 表3 环境因子与介形虫属种排序轴之间的相关系数 Table3 Correlation coefficients between environmental factors and ostracod species ordination axes |
以上结果表明影响黄茅海河口湾表层沉积物样品中有孔虫和介形虫分布的最主要环境因素为水体盐度, 其次是沉积物的砂含量。由于沉积物的砂含量是沉积环境水动力条件的最直接反映, 故也可认为水动力条件是影响研究区微体动物分布的因素之一。此外水深对有孔虫和介形虫的分布也有一定影响, 但并非控制性因素, 这主要是由于研究区水深变化不大, 仅在0~23, m内变化, 而样品中大部分有孔虫和介形虫属种均能在0~20, m水深范围内生存。区域Ⅰ (崖门水道及北部主槽)受径流作用影响最大, 水体盐度接近淡水, 且环境比较动荡, 只有少数适应性非常强的广盐性属种可以在此存活, 故有孔虫和介形虫丰度及分异度均最低。区域Ⅱ (东西两侧浅滩及拦门沙部分浅滩)属径流与潮流共同作用区, 水体盐度居中且呈现由北至南逐渐升高的趋势, 有孔虫和介形虫丰度及分异度也大体呈现出与盐度相同的变化趋势。区域Ⅲ (部分拦门沙浅滩及其以南水域)尽管受潮流及沿岸流影响, 但由于此区域岛屿众多, 水体环境较为稳定, 且此区水体盐度普遍高于10‰ , 比较适合大多数种类生存, 有孔虫和介形虫无论是丰度还是分异度均达到最高值。
黄茅海河口湾表层沉积物样品中共发现有孔虫21属36种, 优势种组合为Ammonia beccarii-Cavarotalia annectens-Quinqueloculina akneriana rotunda-Elphidium advenum, 介形虫16属20种, 优势种组合为Neomonoceratina delicate-Sinocytheridea impressa-Bicornucythere bisanensis。有孔虫及介形虫的优势属种均为广盐— 半咸水性浅水种组合。有孔虫及介形虫的丰度及分异度变化较大, 但基本表现为“ 北低南高” 的趋势。对样品中有孔虫和介形虫属种的去趋势对应分析表明, 水体盐度是影响研究区有孔虫和介形虫分布的最主要因素, 其次是水动力条件, 水深对有孔虫和介形虫分布也有一定影响, 但并非控制性因素, 这主要是由于研究区水深变化不大, 大多数属种均能在此水深范围内生存。河口湾北部由于受径流影响较大、水体盐度较低、水动力条件较强, 不利于海相性强的生物生存, 有孔虫和介形虫的丰度及分异度均较低。而河口湾南部水域盐度明显高于北部, 且由于众多岛屿的天然屏障作用, 环境较为稳定, 有孔虫和介形虫无论是丰度还是分异度均达到最大值。
致谢 国家留学基金委(CSC)对中德联合培养博士生吴洁留德期间的学习提供了资助。感谢德国埃朗根— 纽伦堡大学(Universitä t Erlangen-Nü rnberg)古生物研究所Prof.Dr.Franz T.Fü rsich对微体动物鉴定工作提供的实验条件以及对生态学分析方面的指导。感谢中山大学地球科学系侯卫生老师在应用软件方面的帮助与指导。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|