








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
山东诸城早白垩世中期超大规模恐龙足迹群及其古地理与古生态*
[许欢1  , 柳永清
, 柳永清1 , 旷红伟1 , 王克柏2 , 陈树清2 , 张艳霞2 , 彭楠1 , 陈军1, 3 , 汪明伟1, 4 , 王宝红1, 4 ]
, 柳永清, 旷红伟, 王克柏|
|

通讯作者简介 柳永清,男,1960年生,研究员,主要从事沉积地质学—沉积古地理和古生态学专业研究。E-mail:liuyongqing@cags.ac.cn。旷红伟,女,1969年生,教授,主要从事沉积地质学研究。
第一作者简介: 许欢,男,1987年生,硕士研究生,主要从事沉积地质与盆地分析研究。E-mail:xuhuan0816@126.com。
山东诸城黄龙沟恐龙足迹产于下白垩统中部莱阳群杨家庄组,其中以兽脚类为主,同时有蜥脚类和疑似鸟脚类足迹;兽脚类足迹又可进一步区分为大型兽脚类、似跷脚龙类(Paragrallator)和东方强壮百合龙类(Corpulentapus lilasia)。足迹共组成至少63条行迹,作者系统实测了其中50条,并据足迹大小指数—复步长和 GAE(跷脚龙—安琪龙—实雷龙)图解给出了识别 3类兽脚类足迹的经验性数值。足迹点沉积特征显示,浅湖相粉砂质泥岩和粉砂岩逐渐过渡为滨湖相砂岩。足迹层及其邻近层位波痕、泥裂发育,根据波脊线恢复的古岸线方向为东西向,波浪运动方向呈现出北至北西周期性变化,表明可能存在季节性风向变化。泥裂以及恐龙分布特征显示足迹点以 S/WS为湖心方向,以 N/EN为湖岸方向。似跷脚龙类和东方强壮百合龙类足迹分布区域的差异性表明东方强壮百合龙类生活区域更靠近湖岸。恐龙足迹中存在的明显叠盖现象表明足迹不是同时,而是多次活动遗迹。小型兽脚类行迹中较多近于平行的行迹体现了集群性生活习性。行进行为研究又显示兽脚类的绝大多数为快速奔跑状态,但蜥脚类等为慢行状态。兽脚类与蜥脚类、疑似鸟脚类足迹数量之比远大于正常捕食者和被捕食者之比也表明,早白垩世中期的胶莱盆地生存着以兽脚类为主、蜥脚类和鸟脚类为辅的恐龙动物群。早白垩世晚期,胶莱盆地以及沂沭断裂带内蜥脚类、鸟脚类恐龙以及鸟类逐渐繁盛,这与华北北部恐龙足迹所恢复的造迹恐龙动物群特征较为一致。中国北方、韩国、日本白垩纪恐龙足迹以及骨骼化石发育情况暗示,晚中生代中国北方以及东亚地区发育一个以恐龙等脊椎动物为主的大型陆地动物群。
, Liu Yongqing, Kuang Hongwei, Wang KebaiAbout the corresponding authors Liu Yongqing,born in 1960,is a research professor at the Institute of Geology,Chinese Academy of Geological Sciences,and is mainly engaged in sedimentary geology,palaeogeography and palaeoecology.E-mail:liuyongqing@cags.ac.cn.Kuang Hongwei,born in 1969,is a professor at Institute of Geology,Chinese Academy of Geological Sciences,and is mainly engaged in sedimentary geology.E-mail:kuanghw@126.com.
About the first author: Xu Huan,born in 1987,is a postgraduate candidate of Institute of Geology,Chinese Academy of Geological Sciences. He is mainly engaged in sedimentary geology and basin analysis.E-mail:xuhuan0816@126.com.
Theropod,sauropod and possible ornithopod dinosaur tracks were mainly developed in the middle Early Cretaceous Yangjiazhuang Formation,Laiyang Group,in Zhucheng area,Shandong Province.Most of them are theropod tracks,and belong to three types: Large sized theropod, Paragrallator and Corpulentapus lilasia.Among the 63 trackways identified,50 well-preserved trackways were measured systematically.Based on the foot size index-stride length diagram and GAE( Grallator-Anchisauripus-Eubrontes)plexus,we obtained the differential criteria for the theropod tracks.Large scale sections in the track site showed that shallow lake silty mudstone and siltstone gradually changed to shore-lacustrine sandstone.Track beds and the upper and lower beds developed abundant wavemarks and mud cracks.According to the wave crest,we reconstructed the palaeoshoreline which was EW in direction.The direction of the wave flow showed N-NW periodical change,which may be correlated with shift in the direction of the seasonal wind.The mud crack and the feature of dinosaur track distribution indicated that the center of the lake should be S/WS in direction,while lakeshore should be N/NE in direction.The differences in distribution area between Paragrallator and Corpulentapus lilasia revealed that the Corpulentapus lilasia track makers were living much closer to the lake shore.The phenomenon of superposition is common among dinosaur tracks,which indicated that the tracks were not left at the same time but in several times.Lots of parallel small-medium sized trackways represent gregarious habitats.Speed study showed that most of theropods were running fast,in contrast,sauropods and suspected ornithopods were walking slowly.The proportion between theropod tracks and sauropod and suspected ornithopod tracks indicated that in the middle Early Cretaceous,small-medium sized theropods were dominant in Jiaolai Basin,while sauropods and suspected ornithopods were less;in contrast,sauropods,ornithopods and avians gradually flourished in Jiaolai Basin and Yishu Fault Belt,which was consistent with the dinosaur tracks in northern North China.A comparison of the Cretaceous dinosaur tracks and bones has been made among northern China,Korea and Japan,proposing that a large-scale terrestrial biota represented by dinosaur existed in northern China and eastern Asia during late Mesozoic.
随着Pangea大陆的裂解, 全球晚中生代(185~100 Ma)古地理格局发生了重大变化(Hay et al., 1999; Veevers, 2004)。白垩纪, 以恐龙为代表的脊椎动物群逐渐辐射开来, 尤其是早白垩世, 被认为是现生动物群起源和辐射的重要时期(Lee et al., 2001)。东亚白垩纪陆生生物群的研究近年来受到了广泛关注, 如早白垩世热河生物群, 其中包括带毛恐龙、哺乳动物、翼龙类、原始鸟类、鱼、昆虫以及早期被子植物(Sun et al., 1998, 2002; Xu et al., 1999; Zhang and Zhou, 2000; Zhou and Zhang, 2002, 2003; Luo et al., 2003; Wang et al., 2005; Ren et al., 2009)等。这些动植物化石的发现都是以骨骼等实体化石为依据, 而相应的足迹化石却较为少见, 如冀北— 辽西义县组仅发现两例(You and Azuma, 1995; Xing et al., 2009)。相比之下, 中国东部山东地区早白垩世恐龙骨骼化石较少, 仅见有少量的鹦鹉嘴龙(赵喜进, 1962), 而恐龙、翼龙、鸟类足迹却较多, 在即墨、诸城地区的莱阳群和莒南、东海地区的大盛群中发现了大量兽脚类、蜥脚类、鸟脚类足迹以及部分鸟类、翼龙类和龟类的足迹(李日辉和张光威, 2000; 李日辉等, 2005a, 2005b, 2008; Li et al., 2005, 2011; Lockley, 2007, 2008; Xing et al., 2010, 2012a; 邢立达等, 2010; 柳永清等, 2011)。此外, 经过本研究近3年野外地质调查和研究工作, 作者在沂沭断裂带中相继发现了多处早白垩世足迹化石点, 为研究中国东部地区早白垩世恐龙动物群生存演化、古生态和古环境提供了条件和重要新材料。
恐龙足迹是恐龙生存时留下的印记, 在某些方面能够提供骨骼化石无法提供的信息, 如行为学等, 具有重要的研究价值。目前, 全球各大洲均发现有恐龙足迹化石, 时代从晚三叠世— 晚白垩世, 其中以东亚(尤其是中国和韩国)、北美地区最为人所知(Lucas, 2007; Lockley et al., 2012)。中国足迹化石研究最早可追溯到20世纪20年代, 德日进和杨钟健对陕西神木恐龙足迹化石做了首次报道(Teilhard de Chardin and Young, 1929), 随后, 在云南、四川、甘肃、内蒙、河北、辽宁、山东等地相继发现了大量恐龙足迹(杨钟健, 1979; 甄朔南等, 1983, 1986; You and Azuma, 1995; Lockley et al., 2002; 张永忠等, 2004; Li et al., 2005, 2011; 李建军等, 2006, 2010; Azuma et al., 2006; Zhang et al., 2006; 舒柯文等, 2009; Xing et al., 2012a), 并呈现出晚三叠世局限于西南、侏罗纪遍及中东部地区、晚白垩世全面辐射开来的规律(Chen et al., 2006)。
近年来, 山东地区早白垩世恐龙足迹化石的研究引起了广泛关注。山东作为中国白垩系发育最为齐全的地区之一, 保存有大量恐龙骨骼和恐龙蛋化石, 其时代多集中在晚白垩世(杨钟健, 1958; 甄朔南和王存义, 1959; 胡承志, 1973; 胡承志等, 2001; 刘金远和赵资奎, 2004; 赵喜进等, 2007; Xu et al., 2010; Hone et al., 2011), 而早白垩世恐龙足迹的发现则弥补了骨骼化石缺乏所带来的不足。山东恐龙足迹化石的正式研究始于2000年, 由李日辉和张光威(2000)首先报道了莱阳群龙旺庄组足迹化石, 并命名为Paragrallator。随后, 在沂沭断裂带及以东地区发现了大量早白垩世足迹(Li et al., 2005, 2011; 李日辉等, 2005a, 2005b, 2008; 柳永清等, 2011; Xing et al., 2012a)。
诸城位于胶莱盆地东部, 因发现大规模恐龙骨骼化石而闻名于世(柳永清等, 2010), 而在其北部张祝河湾(Xing et al., 2010)、棠棣戈庄地区的大盛群和南部黄龙沟地区的莱阳群中均发现了恐龙足迹化石(柳永清等, 2011), 其中以黄龙沟地区规模最大, 然而, 该地区恐龙足迹形态学、分类学、埋藏学等方面的研究程度较低, 仅对其中少数兽脚类做了分类学研究(李日辉和张光威, 2000; Li et al., 2011), 柳永清等(2011)对足迹做了简要的埋藏学分析, 李素等(2011)对少数足迹做了变形特征和运动学方面的研究, 何碧竹等(2012)探讨了古地震与恐龙足迹的关系。作者在前人区域地层、盆地发育和演化、古生物学研究的基础上, 对整个黄龙沟地区恐龙足迹进行了较为全面系统的数据测量和分析, 获得了大量形态学和运动学方面的信息, 探讨了胶莱盆地早白垩世早期恐龙动物群古地理、古环境和古生态。
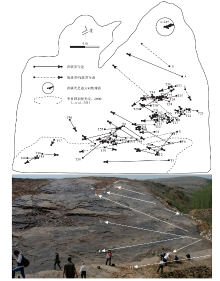
 | 图1 山东诸城位置及区域地质— 构造— 沉积特征A— 据张岳桥等, 2008, 有修改; B— 据柳永清等, 2010, 有修改; C— 据山东省区域地质志; D— 为作者实测Fig.1 Location of Zhucheng in Shandong and regional geology-tectonic-sedimentary characteristics |
胶莱盆地位于华北克拉通东部, 沂沭断裂带东侧的鲁东地区, 为典型的晚中生代伸展断陷盆地(施炜等, 2003; 张岳桥等, 2008)。盆地东南部为胶南隆起, 西北部为胶北隆起(图 1-A)。西界NNE向郯庐断裂和东侧NNE向牟平— 即墨断裂右旋走滑拉分活动共同控制着盆地的形成和发展(戴俊生等, 1995; 张岳桥等, 2007, 2008; Zhu et al., 2012)。盆地基底为太古界胶东群、下元古界荆山群、粉子山群和上元古界蓬莱群, 沉积盖层为白垩系、古近系和第四系, 二者之间为角度不整合接触。白垩系由3套地层单元组成, 即下白垩统莱阳群、青山群和上白垩统王氏群。
莱阳群为一套河湖相碎屑沉积, 是盆地形成初期的产物, 分布广泛, 向东延入黄海, 目前在黄海近岸的灵山岛上就有良好出露(吕洪波等, 2011)。莱阳群自下而上可分为瓦屋夼组、林寺山组、止凤庄组、水南组和龙旺庄组(莱阳地区)或杨家庄组(诸城地区)、曲格庄组和法家茔组(刘明渭等, 1994), 诸城地区见到中间4个组(图1), 整体呈现出粗— 细— 粗的韵律结构。大量古生物资料显示莱阳群时代为早白垩世(陈丕基等, 1980; 沈炎彬, 1981; 马其鸿, 1983; 关绍曾, 1989), 同位素年代学研究指出莱阳群形成时间为135~120 Ma(李金良等, 2007; 张岳桥等, 2008)。
诸城地区位于胶莱盆地南部, 属于诸城凹陷, 白垩系发育齐全, 受控于百尺河断裂和五莲断裂(张岳桥等, 2008)(图 1-A, 1-B)。该区莱阳群沉积厚度较大, 最大厚度可超过2500 m, 为莱阳期胶莱盆地的沉降中心(陆克政和戴俊生, 1994; 张岳桥等, 2008)。皇华镇地区莱阳群整体为一套灰绿色砂岩、粉砂岩夹泥岩, 顶部为灰紫色砂砾岩, 表现为粗— 细— 粗的韵律结构, 与下伏太古界胶东群片麻岩和上覆下白垩统青山群安山岩皆呈角度不整合接触(图 1-C)。
诸城黄龙沟足迹点足迹化石产出于莱阳群杨家庄组上部, 大比例尺实测剖面表明(图 1-D), 足迹层整体厚4.5 m, 下部为灰褐色、灰绿色粉砂质泥岩、泥质粉砂岩、粉砂岩, 水平层理、波状交错层理发育, 为浅湖亚相沉积, 上部为浅灰绿色细砂岩、中砂岩及少量粗砂岩韵律层, 其底部发育楔状交错层理, 恐龙足迹出现于第3层砂岩之中, 波痕和泥裂构造极为发育, 砂体厚度横向上较为稳定, 为滨湖亚相沉积。垂向上沉积相的变化表明, 从杨家庄组沉积早期至晚期湖盆水体逐渐变浅。
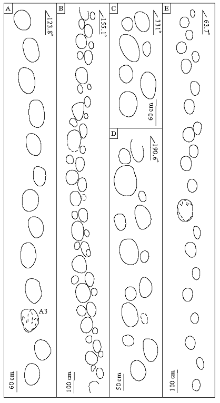
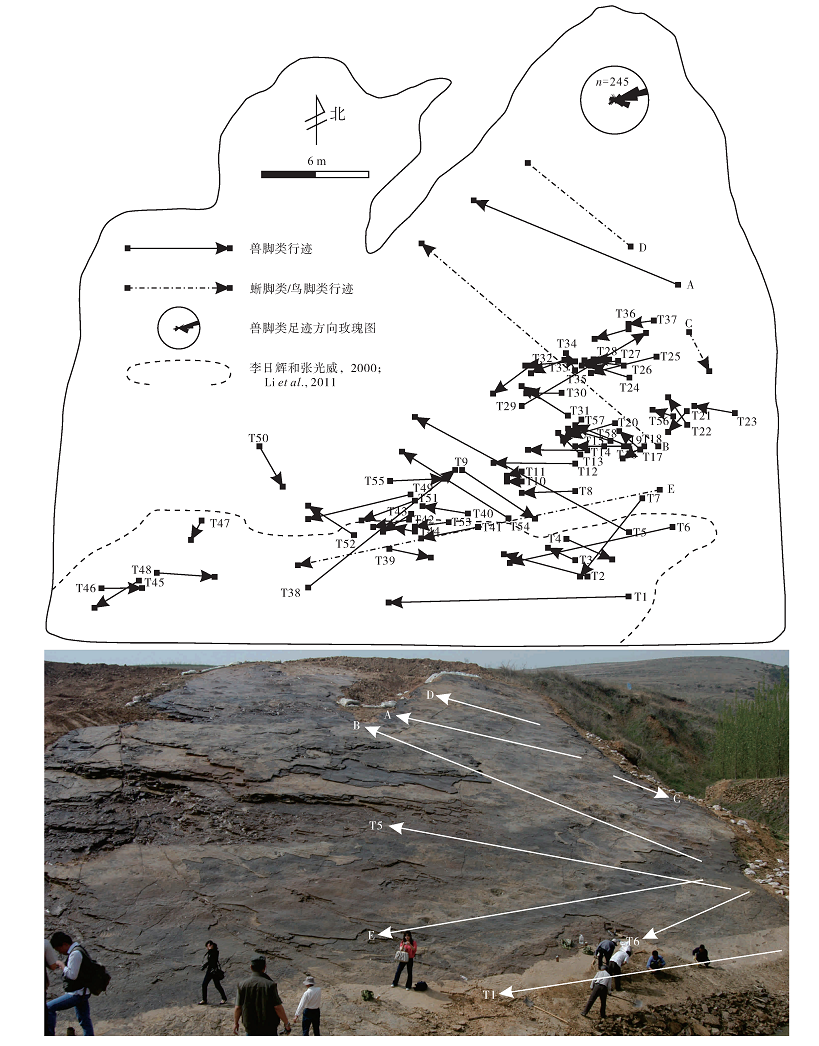
足迹化石点位于诸城市皇华镇黄龙沟(N35° 51'49.27″, E119° 27'32.87″), 是目前已发现规模较大的恐龙足迹动物群之一, 在近3000 m2的面积上出露了11000多个恐龙足迹(柳永清等, 2011)(图 2), 以兽脚类为主, 还含有少数蜥脚类、鸟脚类?足迹(图 3), 是目前山东地区已发现白垩系最低层位的恐龙足迹。足迹主要集中在同一层面上, 赋存于黄绿色细砂岩中, 与泥裂、波痕相伴生。兽脚类足迹印痕清晰, 爪和趾垫较为明显(图 3-A, 3-B, 3-C), 部分大型蜥脚类足迹中充填有极细砂岩, 表明足迹为恐龙真实印记。此次, 对足迹层面进行了全面分析, 共识别出63条行迹, 并对其中50条保存较好的行迹(共352个足迹)进行了相关数据的测量, 其中46条为兽脚类行迹, 3条为蜥脚类行迹, 1条为疑似鸟脚类行迹(图 2)。
 | 图2 诸城皇华镇黄龙沟恐龙足迹化石点行迹分布Fig.2 Trackway distribution in dinosaur track sites in Huanglonggou of Huanghua, Zhucheng |
作者采用Thulborn(1990)足迹测量方法, 分别测量了足迹方位、足长、足宽, Ⅱ 、Ⅲ 、Ⅳ 趾长、趾宽、趾间角(α 和β )、足迹深度、单步长、复步长、步幅角和行迹宽(图 4), 对少数趾部后端分离不清楚的足迹采用了Moratalla等(1988)足迹测量方法, 并对所有足迹进行了精细拍照、编录, 对部分行迹进行了素描。
 | 图3 诸城皇华镇黄龙沟足迹点足迹类型A— 大型兽脚类足迹; B— 似翘脚龙足迹(Paragrallator); C— 东方强壮百合龙足迹(Corpulentapus lilasia); D、E— 特殊大型兽脚类足迹, D为T50行迹中足迹, E为A行迹中足迹; F— 疑似鸟脚类足迹, 为E行迹中足迹; G— 蜥脚类足迹Fig.3 Footprint types in track sites in Huanglonggou of Huanghua, Zhucheng |
足迹所反映的恐龙行进速度的计算方法分别采用了Alexander(1976)和Thulborn(1990)的计算公式。Alexander(1976)认为恐龙的运动速度(V)与复步长(SL)和个体大小(臀高(h)=4× 足长(FL))存在密切关系, 并提出了恐龙的速度计算公式V=0.25× g0.5× SL1.67/(4FL)1.17。Thulborn(1990)将恐龙行进速度划分为3种:行走(SL/h< 2)、慢跑(2< SL/h< 2.9)、快跑(SL/h> 2.9), 他认为Alexander(1976)的计算方法只适合于恐龙行走时的状态, 对于慢跑和快跑的运动状态均不适用, 于是提出了一套适合于恐龙跑动时的计算公式V=[gh(SL/1.8h)2.56]0.5。作者根据恐龙的SL/h比值, 当SL/h< 2时, 用Alexander(1976)提出的计算公式; 当SL/h> 2时, 用Thulborn(1990)提出的计算公式。
 | 图4 足迹测量方法示意图DLⅡ 、DLⅢ 、DLⅣ — Ⅱ 、Ⅲ 、Ⅳ 趾长; DWⅡ 、DWⅢ 、DWIV— Ⅱ 、Ⅲ 、Ⅳ 趾宽; FL— 足长; FW— 足宽; α — Ⅱ 与Ⅲ 趾间角; β — Ⅲ 与Ⅳ 趾间角Fig.4 Measurement method of footprints |
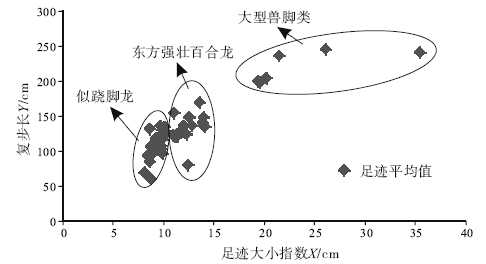 | 图5 足迹大小指数— 复步长图解Fig.5 Footprint size index-stride length diagram |
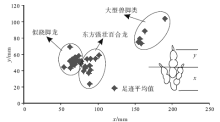
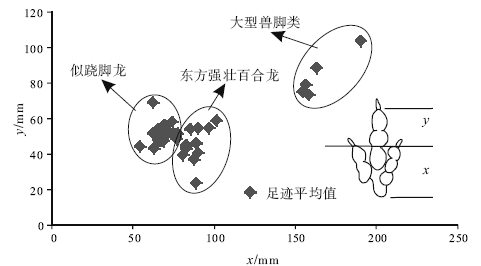 | 图6 GAE图解(测量方法据Olsen, 1980, 有修改)Fig.6 GAE plexus(Method of measurement is modified from Olsen, 1980) |
| 表1 诸城黄龙沟足迹点兽脚类足迹要素 Table1 Elements of theropod tracks in track site of Huanglonggou, Zhucheng |
按照足迹化石特征, 黄龙沟恐龙足迹化石点上万枚足迹可分为3大类:兽脚类、蜥脚类和鸟脚类, 其中绝大多数为兽脚类, 所有化石均呈下凹足迹保存, 主要集中分布在层面的中下部(图 2)。兽脚类足迹形态清晰, 而蜥脚类和疑似鸟脚类足迹多数较浅(足迹层上部个别足迹较深), 仅显示外部轮廓, 无明显趾痕。
Li等(2011)对足迹层下部区域(图 2)兽脚类足迹研究后将其划分为3类:大型兽脚类足迹、似跷脚龙足迹(Paragrallator)和东方强壮百合龙足迹(Corpulentapus lilasia)。作者选择了45条兽脚类行迹(共245个足迹)并对其进行了数据统计和分类学研究, 结果与Li 等(2011)较为一致, 并获得了识别这3类兽脚类足迹的经验性数值。
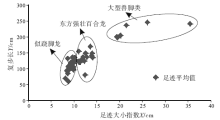
足迹研究与骨骼研究的最大区别在于无法给出准确的分类, 早在1836— 1865年, 一些学者就尝试着对美国康乃迪克州和马萨诸塞州早侏罗世兽脚类足迹进行分类, 如今一些类别名称仍然在广泛使用, 如Grallator、Eubrontes、Anomoepus、Otozoum和Batrachopus(Lull, 1953; Lucas, 2007)。Moratalla等(1988)根据大量足迹数据, 给出了区分兽脚类和鸟脚类足迹的经验性比值, Romilio和Slisbury(2011)用这一方法重新判定了澳大利亚昆士兰地区早白垩世恐龙足迹, 将原先所认为的大型兽脚类改为了鸟脚类。Thulborn和Wade(1989)在研究恐龙运动状态时使用了足迹大小指数— 复步长图解, 从而区分了不同类型的肉食性恐龙, Azuma等(2006)在研究内蒙古早白垩世恐龙足迹时也使用了这一方法, 取得了良好效果。Weems(1992)运用内三角形图解来区分不同个体的恐龙, 随后Olsen等(1998)和Lockley(2009)在其基础上进行了进一步推广, 运用GAE(跷脚龙— 安琪龙— 实雷龙)图解, 很好地限定了Grallator、Anchisauripus和Eubrontes的分布区域, 并提出了恐龙异速生长的概念。因此, 认为足迹大小指数— 复步长图解和GAE图解可以起到区分不同类别恐龙足迹的作用, 并将其运用到了研究区。
如图5和图6所示, 45条行迹数据主要集中在3个区域, 表明主要存在3类兽脚类足迹, 通过对这3个区域足迹形态的比较, 发现正好与Li 等(2011)所划分的3类兽脚类足迹吻合。因此, 研究区兽脚类足迹主要由以上3类组成。
此外, 图5和图6还给出了上述3类兽脚类足迹的特征值, 为今后鉴定这3种足迹提供了经验性数值。图5中横坐标足迹大小指数X为足长× 足宽的平方根, 不同类型足迹主要区别在足迹大小指数上, 似跷脚龙足迹8< X< 11, 东方强壮百合龙足迹11< X< 15, 大型兽脚类足迹15< X< 40, 大型兽脚类足迹的复步长均大于似跷脚龙足迹和东方强壮百合龙足迹。图6中的测量方法在Olsen(1980)的基础上有所改变, 将Y的上部移至Ⅲ 趾末端, 不包含爪的长度, 减小因爪痕不清所带来的误差, 且本图区分不同类型恐龙足迹的关键在于X长度, 图中所示似跷脚龙足迹50< X< 75, 东方强壮百合龙足迹75< X< 105, 大型兽脚类足迹150< X< 200。
3.2.1 兽脚类
兽脚类足迹是足迹化石点最多的一种类型, 根据Li 等(2011)的分类方案, 主要分为大型兽脚类足迹、似跷脚龙足迹(Paragrallator)和东方强壮百合龙足迹(Corpulentapus lilasia), 虽然其研究区域仅集中在足迹层面下部的一小块区域, 但已基本涵盖了所有兽脚类足迹的类型。根据46条兽脚类行迹相关数据(表 1), 对上述3种兽脚类足迹进行系统分析。
大型兽脚类足长大于25 cm, 3趾型, 两足行走。由于保存较完整, 印痕清晰, 皆具有行迹特征。已识别出的兽脚类行迹中有6条为大型兽脚类, 足迹普遍具明显的趾垫和爪痕, 趾节清晰。整体足长25.3~37.0 cm, 足宽14.1~34.0 cm, Ⅲ 趾最长, Ⅳ 趾略长于Ⅱ 趾, Ⅲ 、Ⅳ 趾间角(α )略大于Ⅱ 、Ⅲ 趾间角(β ), 步幅角较大, 173° ~175° , 近于直线运动, 单步与足长比不大于4.25:1, 4条为行走状态, 2条为快跑。其中行迹T50个体较大(图 3-D), 由3个足迹组成, 平均长37 cm, 宽34 cm, 趾间角分别为36.5° 和33.5° , 与鸟脚类相似, 通过运用Moratalla 等(1988)的形态比值判别方法, 各项数据比值符合兽脚类特征(表 2)。因此, 将T50归为大型兽脚类行迹。现对其中2条最长、形态最为清晰的行迹做进一步描述, 叙述如下。
| 表2 诸城黄龙沟足迹点特殊兽脚类足迹要素 Table2 Elements of special theropod tracks in track sites of Huanglonggou, Zhucheng |
T1行迹位于足迹层面最下端(图 2, 图7-T1), Li 等(2011)曾对其进行过形态学分析, 但没有做行迹等方面的研究。该行迹由11个足迹组成, 中部缺少2个足迹, 行迹方位84.3° , 足长26~31 cm, 足宽15.0~18.5 cm, Ⅲ 趾最长, Ⅳ 趾长于Ⅱ 趾, 3趾宽度相近, Ⅲ 趾略向内侧偏转, 单步长111~122 cm, 复步长222.5~247 cm, 步幅角178° ~174° , 速度33.8~9.4 km/h, 且随着复步长的缩短, 步幅角逐渐变大, 速度逐渐减小, 表征着恐龙由快速奔跑向行走状态的转变(图 8-T1)。
T29行迹位于足迹层面中部密集区(图 2), 由11个足迹组成, 趾节、趾垫和爪痕清晰, 行迹方向230° , 足长23~30 cm, 足宽13~17 cm, 趾部特征与T1相似, Ⅱ 与Ⅲ 趾间角为17° ~23° , Ⅲ 与Ⅳ 趾间角为18° ~23° , 单步长90~107 cm, 复步长182~206.5 cm, 步幅角163° ~178° , 速度7.7~9 km/h, 表明恐龙处于行走状态(图 8-T29)。
似跷脚龙足迹(Paragrallator)为李日辉和张光威(2000)命名的一新的足迹类型, 该足迹特征为两足行走, 3趾型, 足长13cm左右, 趾端具尖爪, 中趾最长, 足迹长大于宽, 趾间角较大, Ⅱ 、Ⅳ 趾间角40° 左右, 趾垫不清, 单步与足长比值小于4:1。该类足迹在兽脚类中所占比例最大, 且集中分布在足迹层面的中下部, 中部该类足迹密集度较高。共识别出行迹26条, 总体行迹方向NEE, 少数为W, 足长10~15.5 cm, 足宽6.1~12.7 cm, 且绝大多数集中在6.5~7.5 cm, 足迹较为纤细, Ⅲ 、Ⅳ 趾间角(约20° )略大于Ⅱ 、Ⅲ 趾间角(约18° ), 单步长53~76.5 cm, 单步与足长比值多大于4, 步幅角主要集中在170° ~177° , 复步长为100~130 cm, 大多数处于快速奔跑状态, 这可能是促成单步与足长比值大于4的原因。现选择4条行迹进行详细描述。
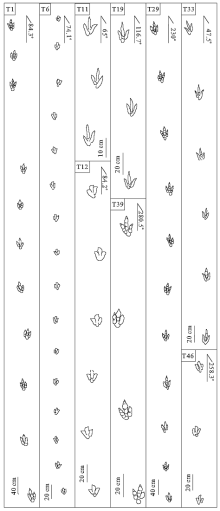
 | 图7 诸城黄龙沟足迹点部分兽脚类行迹特征(T1中只显示了部分足迹)Fig.7 Characteristics of some theropod trackways in the track sites in Huanglonggou, Zhucheng (T1 just shows parts of footprints) |
T11行迹位于足迹层中部(图 2), 由3个足迹组成, 3趾纤细, 形态清晰。行迹方向65° , 足长11~11.5 cm, 足宽6~7.5 cm, Ⅱ 与Ⅲ 趾间角(α )为17° ~19° , Ⅲ 与Ⅳ 趾间角(β )为15° ~24° , 且β > α , 单步长30~32 cm, 单步与足长比值为2.8, 复步长为60 cm, 步幅角为165° , 速度为3.1 km/h, 处于行走状态(图 8-T11)。
T19行迹位于足迹层中部密集区(图 2), 由3个足迹组成, 3趾纤细, Ⅲ 趾向内侧偏转, 部分Ⅲ 趾略显趾垫。行迹方向116.7° , 足长13~14 cm, 足宽7~8 cm, Ⅱ 与Ⅲ 趾间角(α )为19° ~20° , Ⅲ 与Ⅳ 趾间角(β )为20° ~21° , 且β > α , 单步长59 cm, 单步与足长比值为4.4, 复步长118 cm, 步幅角为174° , 速度为22.8 km/h, 处于快速奔跑状态(图 8-T19)。
T33行迹位于足迹层中部偏上位置(图 2, 图7-T33), 由6个足迹组成, 3趾纤细, 形态清晰, 部分足迹具有趾垫, Ⅲ 趾向内侧偏转。行迹方向47.5° , 足长13~16 cm, 足宽6.5~7 cm, Ⅱ 与Ⅲ 趾间角(α )为16° ~23° , Ⅲ 与Ⅳ 趾间角(β )为17° ~20° , 单步长多集中在62~63 cm, 单步与足长比值为4.3, 复步长113~124 cm, 步幅角为173° ~178° , 速度为22.6 km/h, 与T19运动状态相近(图 8-T33)。
 | 图8 诸城黄龙沟足迹点兽脚类行迹示意图Fig.8 Theropod trackways in track sites in Huanglonggou, Zhucheng |
T39行迹为该类型中较为特殊的一条行迹, 位于足迹层面下部 (图 2, 图7-T39), 由4个足迹组成, 与前3条行迹不同的是该足迹趾部粗壮, 具有明显的趾垫, 形态上与大型兽脚类足迹(Eubrontes)相似。行迹方向为280.5° , 足长16~19 cm, 足宽10~11 cm, Ⅱ 与Ⅲ 趾间角(α )为16° ~19° , Ⅲ 与Ⅳ 趾间角(β )为19° ~21° , 且β > α , 单步长82~89 cm, 单步与足长比值为5, 复步长为166~172 cm, 步幅角为173° ~175° , 速度为29.8 km/h, 处于快速奔跑状态(图 8-T39)。
东方强壮百合龙足迹(Corpulentapus lilasia)为Li 等(2011)命名的一新类型足迹, 其特征为外形似百合花, 长12~14 cm, 3趾型, 两足行走, 3趾粗壮, Ⅲ 趾较短, 足迹长宽比较小, 无明显趾垫, 具爪痕。该类足迹在兽脚类较多, 且主要分布于足迹层面下部。共识别出行迹13条, 行迹整体方向向东, 足长12.5~17 cm, 宽9.2~12.5 cm, Ⅲ 趾略长于Ⅱ 趾, 与Ⅳ 趾长度相近, Ⅱ 趾与Ⅲ 趾间角(α )为17.8° ~22.5° , Ⅲ 趾与Ⅳ (β )趾间角为19° ~23.5 ° , 且β > α , 单步长多集中在60~70 cm, 单步长与足长比值大于4, 复步长集中在120~150 cm, 行迹宽为3~7.5 cm, 整体处于快速奔跑状态。现选择3条行迹做详细描述。
T6行迹为该类足迹中最长行迹, 位于足迹层面下部(图 2), 由16个足迹组成, 3趾形态清晰, 无趾垫, 具有爪痕, 足迹方向74.1° , 足长12~13 cm, 足宽9~10 cm, Ⅱ 与Ⅲ 趾间角(α )为16° ~21° , Ⅲ 与Ⅳ 趾间角(β )为16° ~22° , 单步长集中在58° ~62° , 单步与足长比值为4.8, 复步长114~132 cm, 步幅角多为170° ~178° , 速度为24.3 km/h, 处于快速奔跑状态(图 8-T6)。
T12行迹位于足迹层面中下部(图 2, 图7-T12), 由6个足迹组成, 爪痕清晰, Ⅲ 趾向内侧偏转。足迹方向84.2° , 足长13 cm, 足宽10~12.5 cm, Ⅱ 与Ⅲ 趾间角(α )为17° ~22° , Ⅲ 与Ⅳ 趾间角(β )为21° ~24° , 且β > α , 单步长62~63.5 cm, 单步与足长比值为4.7, 复步长为125~128.5 cm, 步幅角在174° ~178° 之间, 速度25 km/h, 为快速奔跑状态(图 8 T12)。
T46行迹位于足迹层下部东侧(图 2), 由3个足迹组成, 形态清晰, 爪痕明显。足迹方向为258.3° , 足长12.5~13 cm, 足宽8.5~11 cm, Ⅱ 与Ⅲ 趾间角(α )为16° ~22° , Ⅲ 与Ⅳ 趾间角(β )为20° ~22° , 单步长72~84 cm, 单步与足长比值为6.1, 复步长为155 cm, 步幅角为177° , 速度为33.2 km/h, 表明恐龙处于快速奔跑状态, 为足迹点运动速度最快恐龙(图 8-T46)。
 | 图9 诸城黄龙沟足迹点特殊兽脚类、蜥脚类和疑似鸟脚类行迹特征A— 大型兽脚类行迹; B— 蜥脚类行迹; E— 疑似鸟脚类行迹Fig.9 Characteristics of special theropod, sauropod and suspencted ornithopod trackways in track sites in Huanglonggou, Zhucheng |
需要指出的是, 在所有兽脚类足迹中, 还存在1条极为特殊的大型兽脚类行迹A(图 9-A, 图10-A)。A行迹位于足迹层面上部, 由13个足迹组成, 外形皆呈椭圆形, 大多无明显趾部形态, 仅A3足迹轮廓较为清晰(图 3-E, ), 测量数据显示(表 2), A3足长71 cm, 宽63 cm, 是黄龙沟足迹点兽脚类中最大个体, 依据Moratalla等(1988)的判别标准, A3有62.5%几率属于兽脚类, 因此将其归为兽脚类足迹。A行迹总体方向123.8° , 足长60~70 cm, 宽50~65 cm, 单步长100~120 cm, 复步长200~220 cm, 步间角为145° ~165° , 足迹深度平均5.8 cm, 速度为3.3 km/h, 表明处于慢速行走状态。
3.2.2 蜥脚类
黄龙沟恐龙足迹化石点共识别出3条保存较为完整的蜥脚类行迹, 并进行了数据实测(图 10-B, 10-C, 10-D; 表3)。该蜥脚类为4足行走, 后足较大, 趾端不明显, 后足呈椭圆形凹坑, 前足呈近圆形, 较后足浅, 部分足迹周围形成突起的脊状边缘, 后足步间角普遍大于前足, 整体行迹宽较窄, 行迹方向多样。
B行迹为足迹点最长、保存最为完好的1条蜥脚类行迹, 位于足迹层面中上部(图 2, 图9-B), 整体由22个前足、25个后足组成, 后足形态较前足清晰, 足迹方位155.1° 。前足长集中在40~55 cm, 宽43~54 cm, 平均复步长180.8 cm, 步幅角88.7° , 行迹宽88.6 cm, 足迹深度平均5.4 cm。后足长86~90 cm, 宽75~80 cm, 复步平均长192.2 cm, 步幅角108.7° , 行迹宽58.2 cm, 足迹深度平均8.7 cm。行进速度为2 km/h, 表明恐龙处于慢速行走状态(图 10-B)。
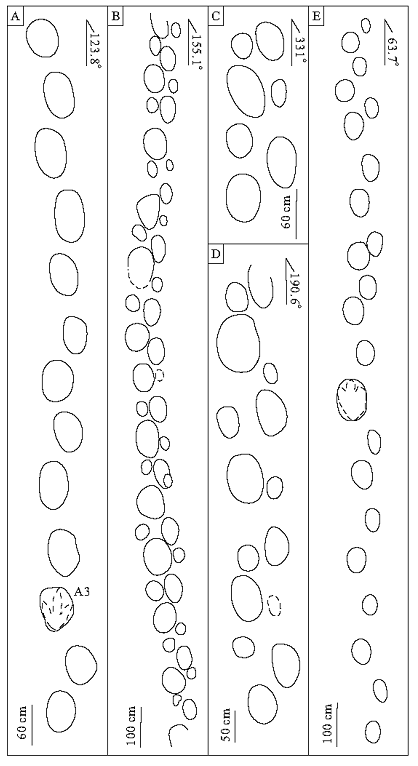 | 图10 诸城黄龙沟足迹点蜥脚类和鸟脚类行迹示意图A— 兽脚类行迹; B、C、D— 蜥脚类行迹; E— 疑似鸟脚类行迹Fig.10 Sauropod and ornithopod trackways in track sites in Huanglonggou, Zhucheng |
C行迹位于足迹层面中部, 行迹长度较短, 仅保存有4个后足和3个前足, 印记皆较浅, 呈椭圆形, 足迹方向为331° 。前足长35~36 cm, 后足长30~37 cm, 复步长146 cm, 步幅角为112° , 行迹宽56 cm, 深度为3.6 cm。后足长60~62 cm, 宽51~59 cm, 复步长154 cm, 步幅角为150° , 行迹宽43 cm, 深度为5.7 cm, 速度为2 km/h, 为慢速行走状态(图 10-C)。
D位于足迹层面最上部, 由6个前足、8个后足组成, 足迹较浅, 仅见椭圆形轮廓, 足迹方向为190.6° 。前足长37~38 cm, 宽30~45 cm, 复步长189.2 cm, 步间角为92.8° , 行迹宽74.5 cm, 深度为3.7 cm。后足长60~74 cm, 宽60~64 cm, 复步长204.8 cm, 步间角为118° , 行迹宽50.5 cm, 深度为7.1 cm。速度为1.4 km/h, 处于缓慢行走状态(图 10-D)。
3.2.3 疑似鸟脚类
根据形态和运动特征来看, 行迹E为两足行走, 部分足迹显示出明显的三趾型特征, 三趾短粗, 呈圆形轮廓, 足部两侧对称, 与鸭嘴龙类似(甄朔南等, 1996)(图 3-F)。一般情况下蜥脚类为4足行走, 且后足步幅角为120° ~140° , 行迹E显示出明显的2足行走特征, 且行迹较窄, 步幅角均超过140° , 与蜥脚类特征不符。因此, 将行迹E归为鸟脚类。
E行迹位于足迹层面下部(图 2), 由22个足迹组成, 部分足迹显示出明显3趾形态。行迹方向63.7° , 足长集中在50~60 cm, 宽40~55 cm, 单步长110~130° , 复步长200~230 cm, 步幅角140~155° , 足迹深度平均6.2 cm, 速度为3.7 km/h, 为慢速行走状态(图 10-E)。
胶莱盆地在早白垩世莱阳期经历了湖盆形成、扩张到萎缩的过程, 在杨家庄组沉积早期(水南期), 湖盆面积最大, 湖平面水位最高, 诸城在该时期为胶莱盆地沉积中心之一。黄华店地区浅湖、三角洲发育, 气候由干旱转为湿润。进入杨家庄组沉积晚期(龙旺庄期), 构造活动逐渐频繁, 气候向半湿热— 半干旱转化, 到曲格庄组时期, 盆地南部抬升(姜在兴等, 1993)。从诸城黄龙沟足迹点大比例尺实测剖面可以看出, 足迹层由浅湖相泥质粉砂岩、粉砂岩逐渐过渡为滨湖— 三角洲相砂岩, 足迹层位之上则被三角洲相砂岩所覆盖, 整体反映了水体逐渐变浅。
足迹层面及其上下层位皆发育波痕、泥裂构造, 作者对足迹点不同层位波痕进行了统计, 共分出了7层, 足迹层1层, 下部1层, 上部5层 (图 11-A~G)。足迹层波痕保存不好, 波峰、波谷形态不明显, 但波脊较为平直, 应属浪成波痕, 波脊线走向222° 。足迹层上下波痕形态清晰, 均为不对称型波痕, 部分波痕表现出波峰尖锐、波谷圆滑特征, 波脊较为平直, 波痕指数7~10, 为浪成波痕。通过测量波痕陡坡面倾向, 获得了波浪运动方向, 整体表现为北-北西的周期性变化, 同时也指示出湖岸线方向为近于东西向。由于足迹上下邻近层位处于滨湖环境, 湖水较浅, 因而季节性风力的作用很可能是形成这种周期性波浪运动的原因。此外, 从足迹层面波痕和泥裂分布来看, 波痕主要发育在南部和西南部, 泥裂集中在中下部位置(北部), 表明湖盆深水区应该位于南部和西南部, 而北部和东北部为湖岸、陆地区, 这与张岳桥等(2008)恢复的胶莱盆地莱阳期古地理较为一致。此外, 泥裂多以薄层夹于中层砂岩之中, 表明泥滩遭受洪泛时间较短, 而暴露的时间相对较长, 其气候环境应该为季节性的半干旱— 干旱(Paik et al., 2001)。与足迹层泥裂相比, 其上部层位泥裂大小和范围明显扩大(图 11-H, 11-Ⅰ ), 说明气候干旱程度在加剧。
 | 图11 诸城黄龙沟足迹点波痕和泥裂特征A~E— 足迹层上部层位波痕; F— 足迹层波痕; G— 足迹层下部层位波痕; H— 足迹层上部泥裂; I— 足迹层泥裂Fig.11 Wavemark and mud crack in track sites in Huanglonggou, Zhucheng |
 | 图12 诸城黄龙沟足迹点足迹与泥裂、波痕关系A— 蜥脚类足迹盖在波痕之上; B— 泥裂发育在兽脚类足迹之中; C— 泥裂发育在蜥脚类足迹之中; D、E— 波痕发育在蜥脚类足迹中; F— 泥裂层盖在兽脚类足迹之上Fig.12 Relationship between footprints and mud crack and wavemark in track sites in Huanglonggou, Zhucheng |
足印在形成之后需要在特殊的环境中才能被保存下来, 许多学者已对这一过程进行了大量研究(Tucker and Burchette, 1977; Avanzini and Frisia, 1997; Paik et al., 2001)。在黄龙沟足迹点足迹层面上, 部分足迹与泥裂、波痕的关系很好地反映了足迹的形成过程(图 12)。在一次洪泛过后, 恐龙在湖岸沉积物上留下足迹, 如果之前形成有波痕, 那么足迹就会覆于其上(图 12-A), 当湖水退去之后, 之前形成的足迹被暴露地表, 发生干化固结, 并在其表面形成泥裂(图 12-B, 12-C), 当再次涨水时, 干化的足迹就被新的沉积物覆盖、保存, 随后进入成岩阶段, 在足迹被覆盖过程中可能会遭受波浪作用, 在其表面形成后生波痕(图 12-D, 12-E)。
从黄龙沟足迹点足迹类型来看, 以小型兽脚类为主, 且主要分布在足迹层面中下部(北部), 上部仅发现少数大型兽脚类足迹, 而大型蜥脚类足迹则延伸到足迹层面最上部(南部), 表明南部水体较深, 北部水体较浅, 为近岸环境, 与沉积相研究结果一致。此外, 兽脚类中的似跷脚龙足迹(Paragrallator)和东方强壮百合龙足迹(Corpulentapus lilasia)也存在明显的分区性, 似跷脚龙足迹主要分布于足迹层面的中下部, 而东方强壮百合龙足迹则分布于足迹层面的下部, 表明似跷脚龙和东方强壮百合龙类恐龙生活习性上存在一定差异, 即东方强壮百合龙类恐龙更适应于生活在水体更浅的岸边, 而似跷脚龙能够在一定深度水体中活动。
恐龙足迹中存在明显的叠盖现象, 图13所示, 大量兽脚类叠盖在蜥脚类之上, 表明兽脚类足迹出现在蜥脚类之后。在兽脚类之间也存在这一现象, 说明黄龙沟足迹点足迹不是同时、而是几次留下。不同期次的兽脚类行迹中可以发现较多行进方向近于一致的行迹, 这种现象进一步表明小型兽脚类集群性的生活习性, 相比之下, 其他地区的一些小型兽脚类也存在同样习性, 如辽西朝阳、北票晚侏罗世— 早白垩世Grallator(Zhen et al., 1989; 张永忠等, 2004)、北美早侏罗世Grallator(Ostrom, 1972; Lockley, 1991)、澳大利亚中白垩世昆士兰地区的虚骨龙类(Thulborn and Wade, 1979, 1984)等, 并且这一习性在蜥脚类和鸟脚类中也广泛存在(Currie, 1983, 1995; Lockley et al., 1986; Lim et al., 1994; Lockley and Hunt, 1995; Matsucawa et al., 1997, 2001; Zhang et al., 2006)。
兽脚类行迹方向大多为近E方向(图 2), 与湖岸线近于平行, 而蜥脚类和鸟脚类行迹则与湖岸线平行或相交, 表明蜥脚类和鸟脚类适应水边的生活, 这可能与湖边植被茂盛有关, 世界上大多数蜥脚类和鸟脚类恐龙也生活在湖岸或海岸, 如西班牙北部早白垩世鸟脚类生活在滨湖环境中(Platt and Meyer, 1991), 加拿大晚白垩世鸟脚类恐龙随着季节性干旱, 会向海岸三角洲或湿地迁移(Nadon, 1993), 甘肃早白垩世河口群中发现了蜥脚类和鸟脚类游泳迹(Li et al., 2006)等都能够说明这一点。
恐龙运动速度研究表明, 兽脚类大部分处于高速奔跑状态, 而蜥脚类和鸟脚类则处于慢速行走状态, 且相对于行走状态的兽脚类要慢得多, 表明兽脚类行进速度一般较蜥脚类和鸟脚类快, 推测兽脚类沿着湖岸快速向东迁移, 可能在集群追踪迁徙过程中的蜥脚类或鸟脚类。值得指出的是, 在兽脚类行迹T1中发现了明显的减速现象, 随着速度的降低, 其复步长度也逐渐减小, 韩国白垩纪小型兽脚类运动特征的研究也得出相似的结论(Kim and Huh, 2010)。
值得指出的是, 从足迹点兽脚类、蜥脚类以及疑似鸟脚类足迹的比例来看, 完全超出了正常的捕食者与被捕食者比例范围, 如北美科罗拉多晚侏罗世足迹中捕食者和被捕食者比例为1:30(Lockley et al., 1986), 韩国Hai县白垩系津东组足迹中捕食者与被捕食者比例为1:25(Lockley et al., 2006a), 表明早白垩世中期胶莱盆地以小型兽脚类为主, 而蜥脚类和鸟脚类较少, 而早白垩世晚期蜥脚类和鸟脚类逐渐繁盛, 且出现了鸟类足迹, 如诸城北部张祝河湾、莒南后左山、东海早白垩世晚期大盛群中发现的大量鸟脚类、蜥脚类以及鸟类足迹(Li et al., 2005; Xing et al., 2010; 邢立达等, 2010)。这一发展特征与冀北— 辽西地区相似, 即晚侏罗世— 早白垩世土城子期, 燕山地区以小型兽脚类Grallator占绝对主导(Zhen et al., 1989), 仅在延庆和尚义地区发现少数鸟脚类和蜥脚类足迹(柳永清等, 2012; 张建平等, 2012), 而到早白垩世早期, 滦平地区义县组发现的恐龙足迹中, 鸟脚类比例明显上升(You and Azuma, 1995)。上述足迹分布特点表明, 晚侏罗世— 早白垩世早期, 整个中国东部地区以小型兽脚类占绝对优势, 到早白垩世晚期, 蜥脚类和鸟脚类以及鸟类逐渐繁盛。
中生代晚期华北东北部地区发育2个重要生物群, 即燕辽生物群(165~152 Ma)和热河生物群(140~120 Ma)。燕辽生物群主要分布在冀北、辽西以及内蒙东南部, 而热河生物群早期分布于辽西、冀北、内蒙古东南部、中国东北部地区, 并且随后向蒙古国、西伯利亚南部、朝鲜和日本半岛等东亚地区辐射(Zhou, 2006)。一些学者对这2个生物群少数物种进行了对比研究, 提出燕辽生物群是前热河生物群的设想(张俊峰, 2002; Meng et al., 2006; Zhou et al., 2010), 但由于缺乏过渡时期的骨骼化石证据, 未能给出确切定论。随着近些年大量晚侏罗世— 早白垩世恐龙足迹的发现, 弥补了因骨骼化石缺乏带来的不足, 为研究晚侏罗世— 早白垩世过渡时期生物群演化提供了新素材。
晚侏罗世— 早白垩世, 华北北部地区广泛发育一套土城子组(154~137 Ma)紫红色粗碎屑沉积(许欢等, 2011; Xu et al., 2012), 之前多认为土城子期, 整个华北北部为荒原环境, 而近些年大量恐龙足迹化石的发现正在改变着这一看法。在辽西朝阳、北票, 冀北承德、赤诚、尚义以及北京延庆等地土城子组中均发现大量恐龙足迹(张永忠等, 2004; 舒柯文等, 2009; 张建平等, 2012; Xing et al., 2012b; 柳永清等, 2012; 张建平等, 2012)和部分鸟类足迹(Lockley et al., 2006b), 其中以小型兽脚类Grallator为主。由于这些足迹大部分产自土城子组上部, 推测可能已经进入Berriasian— Valanginian。相比之下, 在山东蒙阴上侏罗统三台组中, 发现了小型鸟脚类足迹, 这是山东目前发现的最早的恐龙足迹化石(李日辉等, 2002)。虽然晚侏罗世— 早白垩世中国东部地区没有发现较多恐龙骨骼化石, 但大量恐龙足迹的发现, 表明这一时期广泛存在着恐龙的生存, 很可能是燕辽生物群的延续。
早白垩世中期热河生物群萌发, 在冀北— 辽西以及内蒙古东南部等地区的义县组、九佛堂组中发现了大量脊椎动物骨骼化石, 其中恐龙类的有效命名至少30属35种(周忠和和王原, 2010), 而足迹化石较少, 这可能与火山活动频繁、沉积水域时常受到改造有关。山东莱阳群(特别是水南组)地质年代略晚于冀北— 辽西地区义县组, 可能对比九佛堂组, 其中发育的叶肢介Yanjiestheria sinensis、鱼类Lycoptera sinensis是热河生物群中的典型分子(陈丕基等, 1980), 其次还产有丰富的腹足、昆虫和植物化石, 而恐龙骨骼化石稀少。诸城黄龙沟杨家庄组及莱阳龙旺庄组(李日辉和张光威, 2000; Xing et al., 2012a, 2012b)发现大量的恐龙足迹化石则说明, 热河生物群晚期恐龙仍然十分繁盛, 不仅以群居性的中— 小型兽脚类为主, 而且早白垩世晚期沂沭断裂带大量恐龙足迹, 包括兽脚类、蜥脚类、鸟脚类和鸟类(Li et al., 2005, 李日辉等, 2005a, 2005b, 2008; 柳永清等, 2011; Xing et al., 2012a)的发现又进一步表明, 中国北方东北部早白垩世晚期恐龙种类的丰度进一步增加, 恐龙动物群极为庞大。
中国中西部地区, 早白垩世晚期恐龙动物群也极为壮观和繁盛, 如鄂尔多斯盆地东北部的内蒙古等地东胜组和伊金霍洛组中同时保存有恐龙骨骼和足迹化石, 足迹化石以蜥脚类为主, 其次为兽脚类和鸟类(Russell and Dong, 1993; Russell and Zhao, 1996; Lockley et al., 2002; 李建军等, 2006)。甘肃地区发现的早白垩世恐龙包括兽脚类、蜥脚类、鸟脚类、禽龙类和角龙类等(You et al., 2005; 尤海鲁等, 2006, 2008; 尤海鲁和罗哲西, 2008), 尤海鲁和罗哲西(2008)认为公婆泉盆地新民堡群恐龙和辽西热河生物群恐龙关系密切。准噶尔盆地西北缘下白垩统吐谷鲁群中产有虚骨龙类、驰龙类、巨齿龙类、圆顶龙类和剑龙类化石(赵喜进, 1980)。上述各地区恐龙发育和演化表明, 早白垩世开始, 整个中国北方皆发育着大规模的恐龙动物群, 生活的古地理区域也十分广泛。
与此同时, 东亚韩国和日本半岛早白垩世中晚期, 甚至到晚白垩世也同样广泛发育着以恐龙为代表的动物群。韩国恐龙足迹主要分布在南部沿海一带白垩系庆尚超群河流— 湖相沉积中, 其中以津东组(湖相)足迹化石最为发育。最近Jwa等(2009)对庆尚盆地津东组和下伏Haman组之间凝灰岩进行的锆石U— Pb定年, 获得了96~97, Ma的年龄, 表明韩国足迹化石时代主体上为晚白垩世。日本晚侏罗世— 早白垩世弧后盆地河流相沉积中也发育恐龙类、翼龙类和鸟类足迹(Matsukawa et al., 1995, 1997, 2005; Lockley and Matsukawa, 1998; Azuma et al., 2002; Fujita et al., 2003), 其中以Tetori群足迹化石最为发育, 时代为早白垩世中晚期(Kusuhashi et al., 2006), 与山东早白垩世足迹时代相近。日本恐龙足迹中以鸟脚类为主, 其次为小— 中型纤细兽脚类(Matsukawa et al., 2005), 其中晚侏罗世兽脚类足迹Schizograllator和辽西土城子组的Grallator、山东诸城杨家庄组的Paragrallator较为相似。因此, 东亚晚中生代应存在一个大的以恐龙等脊椎动物为主的生物群, 其应涵盖了燕辽生物群与热河生物群及其生物组合。
1)山东诸城黄龙沟恐龙足迹点包含兽脚类、蜥脚类和疑似鸟脚类足迹, 其中以兽脚类为主。兽脚类足迹进一步可分为大型兽脚类足迹、似跷脚龙足迹(Paragrallator)和东方强壮百合龙足迹(Corpulentapus lilasia)。共识别出行迹63条, 对其中50条进行相关数据实测, 并获得了识别3类兽脚类足迹的经验值。
2)足迹赋存于下白垩统下部莱阳群杨家庄组砂岩中, 大比例尺实测剖面沉积相分析显示, 足迹点由浅湖亚相逐渐演变为滨湖亚相, 水体逐渐变浅, 气候干旱。多期波痕波脊走向指示湖岸线近于东西方向, 波痕陡坡面指向呈现出北— 北西的循环式变化, 可能与季节性风向变化有关。泥裂和波痕的分布以及恐龙的分布情况表明, 足迹点以南/西南为盆地中心方向, 以北/东北为湖岸方向。
3)似跷脚龙足迹(Paragrallator)和东方强壮百合龙足迹(Corpulentapus lilasia)特征性的分布说明东方强壮百合龙类恐龙生存区域更靠近湖岸。恐龙足迹中大量的叠盖现象表明黄龙沟足迹点足迹不是同时, 而是多次留下, 小型兽脚类足迹中较多近于平行的行迹体现了小型兽脚类恐龙集群性的生活习性。
4)速度研究显示兽脚类大多处于高速奔跑状态, 而蜥脚类和疑似鸟脚类处于慢速行走状态。兽脚类与蜥脚类、疑似鸟脚类数量差异完全超出了正常捕食与被捕食比例范围, 表明胶莱盆地早白垩世以兽脚类为主, 早白垩世晚期蜥脚类、鸟脚类以及鸟类逐渐繁盛, 这一特征与华北北部地区较为一致。
5)对比了中国北方、韩国、日本地区白垩纪足迹以及骨骼发育情况, 认为晚中生代中国北方以及东亚地区发育一个以恐龙为主的大型陆地动物群。
致谢 中国地质科学院地质研究所姬书安研究员、刘燕学研究员、江小均博士、王旭日博士参加了2010年部分野外工作, 审稿人为本文提出了有益的修改意见, 编辑们在本文的编辑过程中付出了辛勤的劳动, 在此一并表示感谢。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|