


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
具彩色斑纹颅形贝在广西桂林下石炭统黄金组中的发现:兼论颅形贝属的地质地理分布*
[陈智敏 , 孙元林]
, 孙元林]
, 孙元林]
|
|

第一作者简介 陈智敏,女,1987年生,北京大学地球与空间科学学院在读硕士研究生。E-mail:echo.chen.0112@gmail.com。
文中描述了产自广西桂林地区下石炭统黄金组下部的颅形贝属一新种,桂林颅形贝(Cranaena guilinensis)。新种以平直的前结合缘和背壳上发育中槽为特征。在4枚标本上发现保存有放射状的彩色条带,表明该种当时生活在温暖海域的浅水环境。对该属70个种的地质地理分布和生物多样性变化的初步分析表明,该属可能起源于早泥盆世欧美大陆西北缘的老世界区,之后的地理分布和生物多样性发展以北美地区为中心,经历了中泥盆世—晚泥盆世早期和早石炭世2次比较明显的辐射演化、迁移扩散高峰和晚泥盆世晚期的1次严重衰退。第1次高峰是中泥盆世—晚泥盆世早期,该属的生物多样性达到巅峰,生物地理分布范围扩大到欧美大陆之外的西伯利亚板块、哈萨克斯坦板块和华南板块等;第2次高峰是早石炭世,该属的生物多样性虽不及前一次,但获得了最广泛的地理分布,不仅在北方大陆有分布,而且已进入到冈瓦纳大陆边缘。晚泥盆世晚期该属的1次严重衰退显然与F/F灭绝事件有关。早石炭世之后,该属进入衰退阶段,最终在二叠纪初灭绝。
About the first author Chen Zhimin,born in 1987,is a graduate student for master degree at Peking University.E-mail:echo.chen.0112@gmail.com.
Cranaena guilinensis is a new species of Cranaena from the lower part of the Huangjin Formation(Visean,Lower Carboniferous)of Guilin,Guangxi.It is characterized by a rectimarginate anterior commisure and a dorsal sulcus.Four specimens of this species show radiate color bands on shell surface,suggesting that it lived in a shallow water of warm marine realm.Preliminary study on the geological and geographic distribution and biodiversity pattern of 70 species of the genus indicates that this genus probably originated in area of the Old World Realm peripheral to the Euramerican Continent of the Early Devonian and experienced two times of radiation and dispersal during the Middle to early Late Devonian and Early Carboniferous,respectively,and a biodiversity crisis in the Famennian of Late Devonian.The genus reached to the zenith of its biodiversity during the Middle to early Late Devonian and its geographic distribution also expanded to plates of Siberia,Kasakstan and South China.Although the biodiversity of the genus in the Early Carboniferous is less than that in the Middle to early Late Devonian,the genus obtained the widest geographic distribution,not only in continents of North Hemisphere,but also in the peripheral areas of the Gondwana Continent.The Famennian crisis is obviously related with the F/F mass extinction event.After the Early Carboniferous the genus began to decline and was finally extincted in the beginning of Permian.
在化石记录中, 具有彩色斑纹的化石比较罕见, 因此, 这类化石的发现和研究, 具有潜在的生态学、分类学和生物地理学方面的意义。保存有彩色斑纹的有铰类腕足动物化石记录最早可追溯到早泥盆世早期(Balinski, 2010), 到目前为止, 已在75种古生代有铰类腕足动物化石中发现保存有彩色斑纹, 包括1种扭月贝类、2种戟贝类、1种正形贝类、1种直房贝类、9种小嘴贝类、8种无窗贝类、6种石燕贝类和47种穿孔贝类(Blodgett et al., 1983, table 1; Sun et al., 1999, table 1; Modzalevskaya, 2007; Balinski, 2010; Baranov and Blodgett, 2011)。在已发现的75种具有彩色斑纹的有铰类腕足动物化石中, 泥盆纪的种类超过57%(43种), 二叠纪的种类占30%(23种), 而石炭纪的种类仅占12%(9种)。但在中国只在晚二叠世的腕足动物化石中有过1例报道(Sun et al., 1999)。文中所记述的是在桂林地区下石炭统韦宪阶黄金组下部地层中发现的保存有放射状彩色条纹的颅形贝, 为中国华南地区石炭纪有铰类腕足动物中的首次发现。该颅形贝本身也是在华南地区韦宪阶中首次发现。
颅形贝属是穿孔贝目隐弧贝超科(Cryptonelloidea)中一个较为原始的代表, 其初现于早泥盆世, 中泥盆世和早石炭世早期曾广泛分布于老世界区和东美区, 最终灭绝于二叠纪初。在该属中, 前人描述和命名了近70种, 文中亦对该属在泥盆纪和石炭纪的地质地理分布及生物多样性变化规律进行了初步的总结。
在桂林东南磨盘山— 华侨农场一带(图1), 下石炭统黄金组广泛出露, 构成了一个北东— 南西向的向斜的核部, 与下伏的下石炭统英塘组白云岩(鸡冠山白云岩)呈假整合接触(两组之间存在古喀斯特面)。据区域地质调查资料, 黄金组在桂林磨盘山地区总厚近700 m, 主要由灰色至深灰色含中厚层生物碎屑灰岩和微晶灰岩构成, 常夹中薄层泥岩及泥质灰岩, 中上部常含燧石结核和条带及白云质条带, 并含有丰富的珊瑚、腕足类等化石。
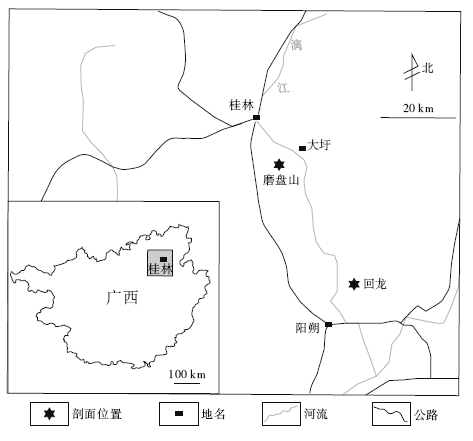 | 图1 广西桂林下石炭统黄金组颅形贝采样点位置Fig.1 Sampling position of Cranaena from the Lower Carboniferous Huangjin Formation in Guilin, Guangxi |
文中所研究的材料来自广西桂林市东南约15 km处磨盘山南侧的华侨农场剖面和桂林市南约50 km的回龙剖面(图 1)下石炭统黄金组中。华侨农场剖面的化石产出在距黄金组底界之上约50 m层段内, 其中, 在黄金组底界之上约35 m处的灰岩层面上共采集到14枚Cranaena的标本, 有4件壳体上保存有彩色斑纹的痕迹。除此之外, 与之共生的尚见有腕足类Martinothyris?sp.和Punctospirifer sp., 以及一些头足类、腹足类、珊瑚和棘皮动物碎片等。回龙剖面的化石材料采自距黄金组底界之上80~100 m的含硅质团块结核的灰岩中, 在实验室中采用酸解的方法获取了15枚硅化的标本。依据这些材料, 命名一新种 Cranaena guilinensis。华侨农场剖面描述如下:
10.中厚层泥粒灰岩夹泥岩。含腕足类Megachonetes sp.等。5 m(未到顶)
9.中薄层泥晶— 泥粒灰岩与泥岩互层, 夹薄层颗粒灰岩。含腕足类Delephinae sp. megachonetes sp.等。5.1 m
8.中厚层泥粒灰岩夹泥岩和泥灰岩。含腕足类Megachonetes sp., Cranaena guilinensis sp.nov. martinothyris?sp.和Punctospirifer sp.等。5.3 m
7.泥岩与中薄层泥粒— 泥晶灰岩互层。4.9 m
6.中厚层泥晶— 泥粒灰岩夹泥岩, 底部有白云岩化。含腕足类、珊瑚等化石。5.6 m
5.灰色中厚层颗粒灰岩— 泥粒灰岩夹泥岩, 富含腕足类、珊瑚等化石。3.3 m
4.深灰色中厚层泥晶— 泥粒灰岩, 夹颗粒灰岩和泥岩。4.5 m
3.深灰色薄层泥质灰岩和泥晶灰岩, 夹颗粒灰岩和泥岩, 富含腕足类、珊瑚等化石。5.9 m
2.灰色中厚层颗粒灰岩与深灰色泥晶— 泥粒灰岩互层, 夹泥岩。含腕足类Delephinae sp., Cranaena guilinensis sp.nov.和珊瑚等化石。6.29 m
1.深灰色泥粒灰岩与瘤状灰岩互层, 夹生物碎屑灰岩和泥质灰岩, 富含腕足类、珊瑚等化石。5.9 m
— — — — — — 假整合— — — — — —
下伏地层:下石炭统英塘组白云岩及白云质灰岩
桂林地区下石炭统黄金组包含了从韦宪期到谢洛霍夫期的沉积物, 与黔南地区的祥摆组— 上司组相当。文中的研究层段与黔南地区相比, 大体相当于祥摆组, 属于韦宪阶的下部。根据Hance等(2011)对华侨农场剖面的研究, 杜内阶/韦宪阶界限位于英塘组上部的白云岩内, 黄金组下部50余米的层段属于MFZ12有孔虫化石带, 在时代上属于韦宪早期。
穿孔贝目 Terebratulida Waagen, 1883
隐弧贝超科 Cryptonelloidea Thomsom, 1926
颅形贝科 Cranaenidae Cloud, 1942
颅形贝属 Cranaena Hall & Clarke, 1893
模式种 Terebratula romingeri Hall, 1863
属征 壳体小至中等大小, 近圆形至长卵形; 侧视不等双凸, 腹壳凸度强于背壳; 腹中槽和背中隆发育或不发育; 前结合缘平直至微弱单褶型; 茎孔过中窗型; 齿板短; 铰板联合成主板, 空悬, 有穿孔; 三角形(deltiform)腕环。
桂林颅形贝(新种) Cranaena guilinensis sp.nov.(图 2)
词源 桂林, 化石产出的地区
材料 25枚壳体
描述 壳体小, 轮廓呈略纵向延长的圆五边形; 侧视不等双凸, 腹壳强凸, 背壳缓凸; 最大壳宽位于壳长的1/2处。壳体前边缘微微内凹, 前结合缘平直。腹壳喙部弯曲近90° , 肉茎孔形状近卵圆形, 过中窗型; 腹三角孔覆有三角双板; 腹壳壳面前1/3处发育宽缓的中槽。背壳无中隆, 前缘发育微弱的中槽与腹中槽对应。壳体具疹, 疹孔密度大约在235个/mm2。
腹壳内部肉茎领发育, 齿板薄片状, 短, 略向外延伸。背壳内部内铰板略向背凹, 并相向延伸联合形成主板, 空悬无中隔板支持, 后端穿孔。腕环着生在外铰板和内铰板交界的前缘, 其侧带沿侧前方延伸约为背壳长度的1/5~1/4, 前带不清楚。
 | 图2 广西桂林黄金组中的桂林颅形贝(新种)1a~1d— PKUM02-0631的腹视、背视、左侧视和右侧视, 示表面的放射状彩色条带; 2a~2e— PKUM02-0632的腹视、背视、侧视、前视和后视; 3, 4— PKUM02-0633和PKUM02-00634的腹视, 示表面的放射状彩色条带; 5a~5c— PKUM02-0635的腹视、背视和前视, 示壳面的放射状彩色条带; 6— 硅化的壳体后部内视(上为腹壳), 示齿板和主基构造, PKUM02-0636; 7a~7e— PKUM02-0637的腹视、背视、后视、前视和侧视; 8a~8e— 标本PKUM02-0638(正型)的腹视、背视、后视、前视和侧视; 9a~9c— PKUM02-0639的腹视、背视和侧视; 10a~10e— PKUM02-0640的腹视、背视、侧视、前视和后视; 11a~11e— PKUM02-0641的腹视、背视、侧视、前视和后视(幼年个体); 12a~12e— PKUM02-0642的腹视、背视、侧视、前视和后视(幼年个体); 13— PKUM02-0643的背壳内视, 箭头指示腕环; 14— PKUM02-0644腹壳壳面的疹孔Fig.2 Cranaena guilinensis sp.nov.from the Huangjin Formation of Guilin, Guangxi |
讨论 弯曲的腹喙、具齿板、主板空悬穿孔和短小的腕环等特征明确指示当前的新种属于颅形贝属。成年体背壳具有微弱的中槽是新种不同于该属中多数其他种类的主要特征。在颅形贝属中, 只有产自美国德州下石炭统的C.?dorsisulcata具有背中槽(Carter, 1967), 同时在壳体大小和轮廓上与新种十分相似。但C.?dorsisulcata的最大壳宽位于壳体中线之前, 前结合缘呈单褶型, 不同于新种。
产地和层位 桂林磨盘山、回龙一带下石炭统黄金组下部。
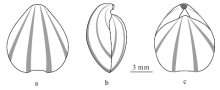
在4枚保存有彩色斑纹的标本中, 腹壳上的彩色斑纹保存较为完整清晰。标本PKUM02-0631的腹壳在正视的情况下可清楚地看到在淡黄白色壳面背景色下的2对始于喙部、向前逐渐变宽的青灰色放射状条带(图 2-1a)。在侧视的情况下, 在腹壳肩部侧缘还可以观察到1条(图 2-1c, 2-1d), 表明该种的腹壳上共覆有3对左右对称、等间距均匀分布的放射状彩色条带(图 3-a, 3-b)。其余3枚标本上, 由于标本的保存关系, 只能见到靠近中央的2对条纹(图 2-3, 2-4, 2-5)。背壳上的彩色斑纹, 只在2枚标本上有残留。在标本PKUM02-0631残损背壳的喙部可观察到2对始自壳顶的放射状条纹的痕迹(图 2-1b); 在标本PKUM02-0635残损背壳的壳面可见到2对左右对称分布的放射状条纹痕迹(图 2-5a), 表明该种的背壳具有至少4条彩色条纹(图 3-c)。可观察到的2对条纹分布型式与腹壳靠近中央的2对条纹相似。在已知保存有放射状彩色条纹的颅形贝属(Cranaena)其他种中, 绝大多数种类背壳和腹壳上的条纹数目是对等的, 因此, 有理由相信Cranaena guilinensis sp.nov.背、腹壳上彩色条纹的数目差异有可能是保存的缘故, 其背壳上也应该是6条, 而不是4条。
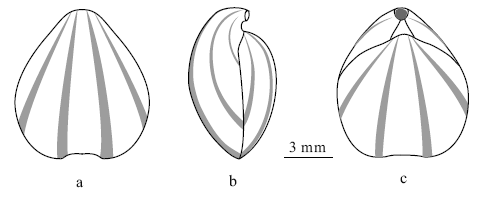 | 图3 桂林颅形贝(Cranaena guilinensis sp.nov.)壳体表面彩色纹饰复原(主要依据标本PKUM02-0631和PKUM02-0635)a— 腹视; b— 侧视; c— 背视Fig.3 Restored color pattern of Cranaena guilinensis sp.nov.(mainly based on samples PKUM02-0631 and PKUM02-0635) |
相对于其他种类的腕足动物而言, 彩色斑纹的保存在颅形贝属(Cranaena)中并不罕见。在该属正式命名的60多个种中, 前人已在15个种中发现保存有彩色斑纹, 其中有2个种为同心环带、11个种为放射状条纹和2个种为放射状斑点(Cooper, 1938; Stainbrook, 1941; Cloud, 1941, 1942; Griesemer, 1965; Tuftin, 1984; Blodgett et al., 1983, table 1; Modzalevskaya, 2007)。但这些记录均只限于泥盆纪的种类。文中所描述的桂林颅形贝(新种)则是目前所知的石炭纪唯一保存有彩色斑纹的颅形贝属化石记录。
彩色斑纹在无脊椎动物化石上的保存具有潜在的分类学、生态学和生物地理意义(Blodgett et al., 1988; Baliń ski, 2010)。
在分类学上, 一些特定的彩色斑纹图案可以作为一些低级分类单元的重要鉴定特征补充, 如Blodgett等(1983, 1988)至少部分地将腕足动物Beachia和?Prionothyri背壳上稳定出现的1对放射状彩色条纹视为Eurythyridinae亚科的重要辅助鉴定特征。对于颅形贝属而言, 前人已在泥盆纪的11个种中发现保存有放射状的彩色条纹, 其中有5个种(Cranaena radiata、C.cooperi、C.thomasi、C.inflata、C.elia)与文中描述的C.guilinensis sp.nov.一样, 具有比较恒定的6条放射状条纹, 但只有C.radiata和C.cooperi的彩色放射状条纹在粗细和分布间距上与C.guilinensis sp.nov.相近。其他几个种上的放射状条纹数目也在4~8条之间变化。从已知的情况看, 尚难以确定放射状条纹数目和粗细, 以及分布间距的变化在这些种类中的分类学价值, 但放射状彩色条纹在所有这些种的单个标本上, 总是在背壳和腹壳上对等分布, 可以说是这些具有放射状彩色条纹颅形贝种类的共同特征。这样分布的放射状彩色条纹或条带一般与壳体内表面的膜脉管主支印痕相对应, 其成因可能与动物的生理反应有关(Cloud, 1942)。
对于现生的海生无脊椎动物而言, 鲜艳的色彩和斑纹是生活在温暖水域浅水环境中的一些种类所具有的特征。具有鲜艳色彩和斑纹的现生穿孔贝类通常生活在水深小于50 m的浅水环境中, 其色素源于腕足动物所摄取的藻类(Cusack et al., 1992); 深水型种类的壳体则通常呈白色、奶油色或青灰色(Lee et al., 2006)。古地磁资料显示, 华南板块在石炭纪地处低纬度的赤道附近(王俊达和李华梅, 1998), 因此, 具有彩色斑纹的Cranaena guilinensis sp.nov.的发现, 说明其当时生活在温暖透光的浅水环境, 黄金组的沉积也是在比较温暖的浅海环境中形成的。彩色斑纹同时也可以起到伪装的作用。
在生物地理上, 如果获取的化石记录信息足够完整, 那么各个地质时代具有彩色斑纹的海生无脊椎动物化石的地理分布就可以用于区分当时寒冷的极区水域和温暖的温带— 热带水域。在极区水域的贝类动物壳体通常缺乏彩色斑纹(Nicol, 1967), 而在温带水域贝类动物壳体上彩色斑纹已比较普遍, 在热带水域最为丰富、并且具有最为复杂的图案。这样的地理分布已经被泥盆纪的海生无脊椎动物化石所证实(Blodgett et al., 1988):所有已知保存有彩色斑纹的化石均出现于老世界区(Old World Realm)或东美区(Eastern Americas Realm), 而在被认为是当时处于南半球高纬度地区的马尔维诺卡夫区(Malvinokaffric Realm)则无。虽然, 石炭纪具彩色斑纹的有铰类腕足动物化石纪录比较少, 只在10个种中发现, 包括在美国发现的Mesolobus mesolobus、Chonetinella jeffordsi、Hustedia mormoni和Orthotetes kaskaskiensis(Morningstar, 1922; Stevens, 1965; Nitecki and Sadlick, 1968; Hoare and Sturgeon, 1979), 在英国和俄罗斯发现的Dielasma hastata或 D.cf.D.hastata(Davidson, 1857; Baily, 1875; Kalashinikov, 1968); Martinia glaber, Schizophoria resupinata, Dielasma elongatum, 以及文中描述的Cranaena guilinensis sp.nov.。这些记录都只限于当时的欧美板块和华南板块的范围之内, 而从石炭纪开始, 这2个板块的主体均毫无例外地处于赤道附近(图 4)。在石炭纪的其他无脊椎动物化石中已知有66个属中发现保存有彩色斑纹, 包括3个三叶虫属、8个头足类属、43个腹足类属和12个双壳类属(见Kobluk and Mapes, 1989, tables 1-3, 5)。这些具有彩色斑纹的化石记录主要限于美国中大陆地区和欧洲英伦地区, 少量发现见于比利时和日本。这些地区在石炭纪也都处于赤道附近的低纬度地区。
颅形贝属是穿孔贝目隐弧贝超科(Cryptonelloidea)中一个较为原始的代表, 其以具有空悬穿孔的主板和短小的三角型(deltiform)腕环为特征。该属初现于早泥盆世, 灭绝于二叠纪初。本节所涉及的该属资料包括了经Cloud(1942)系统厘定确认的39种和部分有疑问的种, 以及后人描述命名的20余种, 共计有70种(表 1)。此外, 前人研究中所记录的一些未定种或有疑问的未定种也收集在内, 用于该属的地质地理分布讨论。该属的地质地理分布资料也包括了少量来自因特网上古生物数据库(Paleobiology Database, 缩写PDB)(http://www.paleodb.org)的资料。
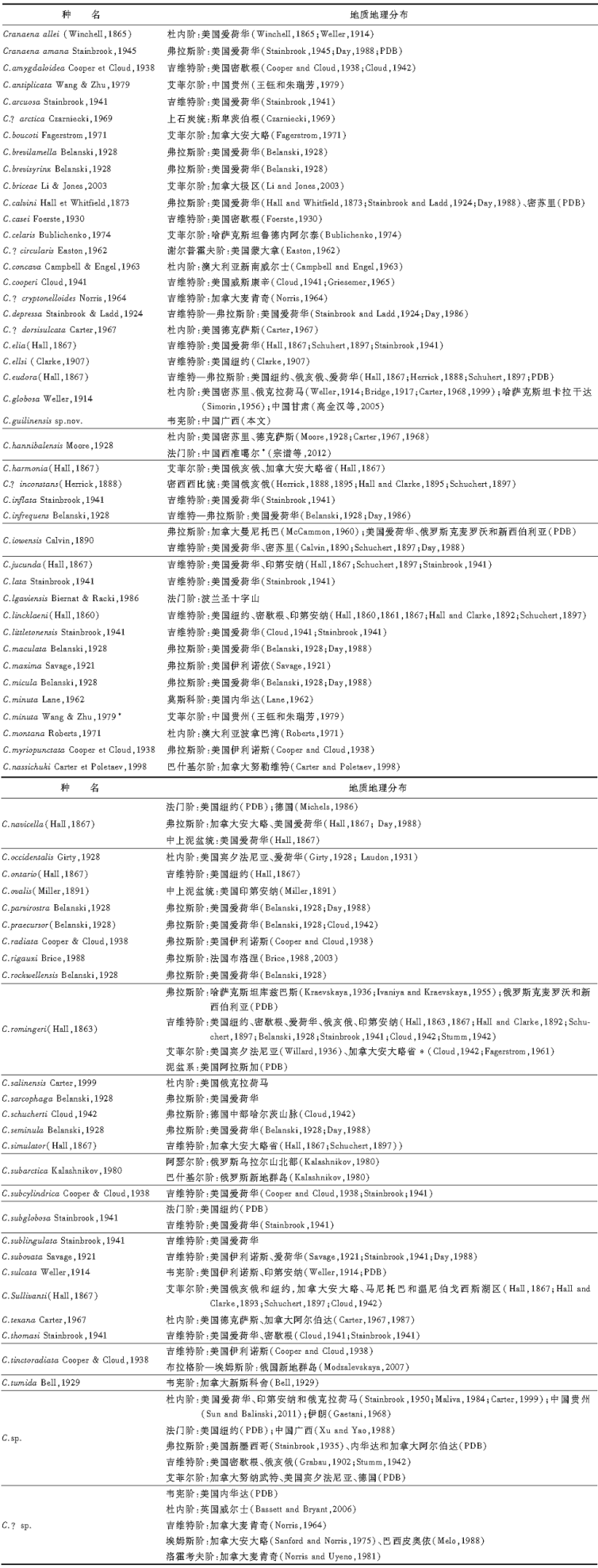
| 表 1 颅形贝属的地质地理分布统计 Table 1 Statistics table of geological and geographic distribution of Cranaena |
早泥盆世— 中泥盆世艾菲尔期 Modzalevskaya(2007)记述的产自俄罗斯新地群岛(Novaya Zemlya)南岛下泥盆统布拉格阶— 埃姆斯阶底部的Cranaena tinctoradiata, 及Levy 和 Nullo(1970)描述命名的产自南美阿根廷圣胡安省(San Juan)下泥盆统的C.argentina, 是目前已知有正式描述的早泥盆世颅形贝化石记录。需要说明的是, 俄罗斯新地群岛的标本可能是与C.tinctoradiata不同的种。Modzalevskaya(2007)将其归入C.tinctoradiata的理由是其具有与该种模式标本(Cooper et Cloud, 1938, p.449, pl.54, Fig.27-28)相似的褐红色放射状彩色条带和微凹的壳体前缘。但是新地群岛的标本壳体更加细长, 其壳面的放射状彩色条带为6~8条, 而不是4条。
此外, 早泥盆世的颅形贝化石记录还包括产自加拿大大布列颠哥伦比亚省麦肯齐地区洛霍考夫阶“ Delorme” 组(Norris and Uyeno, 1981)、加拿大安大略省哈德逊湾埃姆斯— 艾菲尔阶Kwatoboahegam组(Sanford and Norris, 1975)和巴西皮奥伊省(Piaui)埃姆斯阶Pimenteira组的3个可疑记录。所有这些化石记录, 以及上述的俄罗斯新地群岛和阿根廷圣胡安省的化石记录, 均限于加里东运动之后形成的欧美大陆内以及冈瓦纳大陆的西缘(图 4)。
 | 图4 颅形贝属的地质地理分布1~2— 广西和贵州(Guangxi and Guizhou, China); 3~4— 甘肃和西准噶尔(Gansu and West Jungar, China); 5~7— 卡拉干达、库兹巴斯和鲁德内阿尔泰(Karagandinsk, Kuzbass and Rudny Altai, Kazakhstan); 8~9— 新西伯利亚和克麦罗沃(Novosibirsk and Kemerovsk, Russia); 10~11— 北乌拉尔和新地群岛(North Ural and Novaya Zemlya, Russia); 12— 斯卑茨伯根(Spitsbergen); 13— 加拿大极区(Arctic Canada); 14— 阿拉斯加(Alaska); 15~19— 麦肯齐、努勒维特、阿尔伯达、安大略和新斯科舍(Mackenzie, Nunavut, Alberta, Ontario and Nova Scotia, Canada); 20~26— 内华达、蒙大拿、新墨西哥、德克萨斯、俄克拉荷马、威斯康星和密歇根(Nevada montana, New Mexico, Texas, Oklahoma, Wisconsin and Michigan, USA); 27~33— 密苏里、爱荷华、伊利诺斯、印第安纳、俄亥俄、宾夕法尼亚和纽约(Missouri, Iowa, Illinois, Indiana, Ohio, Pennsylvania and New York, USA); 34— 威尔士(Wales); 35— 法国(France); 36— 德国(Germany); 37— 波兰(Poland); 38— 伊朗(Iran); 39~40— 波拿巴湾盆地和新南威尔士(Bonaparte Gulf Basin and New South Wales, Australia); 41— 西皮奥伊 (Piaui, Brazil); 42— 圣胡安(San Juan, Argentina)Fig.4 Geological and geographic distribution of Cranaena |
在早泥盆世, 加拿大大布列颠哥伦比亚省和俄罗斯新地群岛在腕足动物生物地理区系属性上同属于Boucot等(1969)所划分的老世界区(Old World Realm)中的科迪勒拉— 乌拉尔省; 加拿大安大略省哈德逊湾地区和巴西皮奥伊省则属于东美区(Appalachian Realm); 而阿根廷圣胡安省则介于东美区和马尔维诺卡夫区(Malvinokaffric Realm)的过渡带上, 早泥盆世早期属于马尔维诺卡夫区(Malvinokaffric Realm), 早泥盆世晚期属于东美区(Boucot et al., 1969)。因此, 如果这些化石记录可靠的话, 那么颅形贝属很可能起源于欧美大陆西部的老世界区, 并逐渐沿着当时欧美大陆的边缘迁移扩散, 向北至俄罗斯新地群岛, 向南或东南进入东美区。
与早泥盆世相比, 中泥盆世早期(艾菲尔期)颅形贝化石记录的地理分布已有明显的扩大(图 4)。在欧美大陆上, 北美中东部地区的阿帕拉契亚盆地和密歇根盆地分布有4种(C.boucoti、C.harmonia、C.romingeri和C.sullivanti)(Hall, 1867; Hall and Clarke, 1893; Schuchert, 1897; Willard, 1936; Cloud, 1942; Fagerstrom, 1961, 1971), 在加拿大北部的努纳武特(Nunavt)(C.sp., PDB)和北极区(C.briceae), 及德国的莱茵地区(C.sp., PDB)也有分布。同时, 该属也分布到了西伯利亚板块边缘(哈萨克斯坦的鲁德内阿尔泰(Rudny Altai)地区)(C.celaris)(Bublichenko, 1974)和中国的华南地区(C.antiplicata和C.minuta)(王钰和朱瑞芳, 1979)。需要说明的是王钰和朱瑞芳(1979)所描述命名的C.minuta是C.minuta Lane, 1962的后出重名, 需要另行命名替代。在种类上, 颅形贝属也从早泥盆世的2个种增加到了8个种(表 1, 图5), 反映了该属在中泥盆世早期开始的快速辐射演化和扩散(图 4)。在北美的阿帕拉契亚盆地和密歇根盆地, 该属开始成为腕足动物群中的重要组成分子, 并在中泥盆世晚期和晚泥盆世早期持续繁荣。在加拿大北极地区还演化出现了壳面前部具放射褶的褶颅形贝属Costacranaena, 成为该地区艾菲尔期的一个典型土著型分子(Johnson and Perry, 1976; Li and Jones, 2007)。
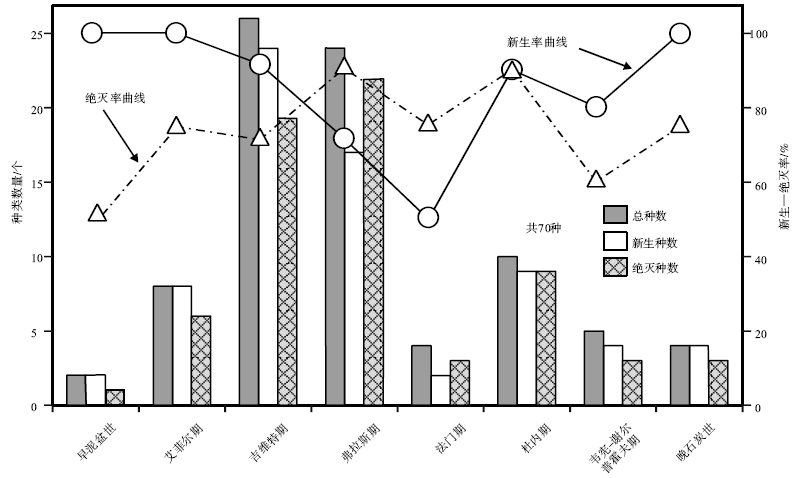 | 图5 泥盆纪和石炭纪颅形贝属的生物多样性及新生— 灭绝率曲线Fig.5 Biodiversity pattern and origination-extinction curves of Cranaena during the Devonian and Carboniferous |
中泥盆世吉维特期— 晚泥盆世弗拉斯期 颅形贝属在中泥盆世吉维特期至晚泥盆世弗拉斯期迎来了其生物多样发展的第1次顶峰。已有的化石记录显示, 先后有43种颅形贝生存在这个时期, 占该属已命名种类的60%以上(表 1, 图5)。其中, 在吉维特期有26种, 其中24种为新生分子; 弗拉斯期有26种, 其中17种为新生分子(表 1, 图4)。
目前所知的吉维特期颅形贝化石记录的分布全部限于北美地区, 有25种来自北美中东部的阿帕拉契亚盆地和密歇根盆地周围地区(Savage, 1921; Belanski, 1928; Cooper and Cloud, 1938; Cloud, 1942; Stainbrook, 1941), 1种来自加拿大西部的麦肯齐地区(Norris, 1964)。在世界其他地方缺失化石记录, 可能与研究程度有关。
弗拉斯期, 颅形贝属的分布仍主要集中在北美中东部的阿帕拉契亚盆地和密歇根盆地周围地区, 当时生活的26种中有24种分布在这个区域, 另外2种在欧美大陆东部的欧洲中部地区(Cloud, 1942; Brice, 1988, 2003)。同时, 该属也扩散到了属于西伯利亚板块范畴的俄罗斯克麦罗沃和新西伯利亚地区(PDB), 以及哈萨克斯坦板块的库兹巴斯盆地(Kraevskaya, 1936; Ivaniya and Kraevskaya, 1955), 显示了比较广泛的地理分布。
晚泥盆世法门期 法门期正式描述报道的颅形贝化石记录有3条, 包括波兰圣十字山地区发现的C.lgaviensis(Biernat and Racki, 1986; Halamski and Baliń ski, 2009)、中国西准噶尔地区发现的C.cf.hannibalensis(宗谱等, 2012)和广西桂林地区的C.sp.(Xu and Yao, 1988)。这些记录均限于法门期的最晚期, 代表了晚泥盆世F/F灭绝事件之后的生物复苏阶段的新生类型。目前尚没有发现法门期早期正式描述报道的化石记录, 仅在古生物数据库(PDB)中记录了在美国纽约有C.navicella、C.subglobosa和C.sp., 在德国北威州有C.navicella和C.sp.。如果这些记录属实的话, 那么在美国和德国分布的这些分子则代表了F/F灭绝事件之后的残存分子。法门期稀少的化石记录也间接地说明了在晚泥盆世的F/F灭绝事件中颅形贝属曾遭受了重创。
早石炭世 早石炭世杜内期的颅形贝属虽然在生物多样性的发展上不及中泥盆世吉维特期或晚泥盆世弗拉斯期的一半, 只有10种(表 1, 图5), 但在地理分布上却相当广泛, 不但在北美广泛分布(Winchell, 1865; Hall and Clarke, 1895; Herrick, 1888; Schuchert, 1897; Weller, 1914; Bridge, 1917; Girty, 1928; Moore, 1928; Laudon, 1931; Stainbrook, 1950; Carter, 1967, 1968, 1987, 1999; Maliva, 1984), 在英国的威尔士(Bassett and Bryant, 2006)、哈萨克斯坦中部的卡拉干达盆地(Simorin, 1956)、中国祁连地区和华南地区(高金汉等, 2005; Sun and Balinski, 2011)等也有分布, 甚至也分布到了当时古特提斯洋南岸的伊朗(Gaetani, 1968), 澳洲的波拿巴湾盆地和新南威尔士等地(Campbell and Engel, 1963; Roberts, 1971)。反映了颅形贝属在经历了F/F灭绝事件和法门期的缓慢复苏发展阶段后, 在早石炭世早期开始了新的辐射演化和扩散。
早石炭世晚期颅形贝属的化石记录主要限于北美地区(Weller, 1914; Bell, 1929; Easton, 1962), 以及中国华南地区(本文), 共计有4个种。无论在生物多样发展上还是在地理分布范围上, 都较早石炭世早期有了明显的萎缩, 显示出该属的发展开始逐渐进入了衰退的阶段。
晚石炭世 早石炭世之后的颅形贝属化石记录分布十分零星(表 1), 包括见于俄罗斯新地群岛地区巴什基尔阶的C.subarctica(Kalashnikov, 1980)、加拿大努勒维特巴什基尔阶的C.nassichuki(Carter and Poletaev, 1998)、美国内华达地区莫斯科阶的C.minuta(Lane, 1962)和斯卑茨伯根地区上石炭统中的C.?arctica(Czarniecki, 1969)。这些化石记录都只限于欧美大陆及其边缘(图 4)。该属的最后出现见于俄罗斯乌拉尔山北部地区二叠系底部的阿瑟尔阶中(Kalashnikov, 1980)。
从上述颅形贝属的地质地理分布情况不难发现, 该属在早泥盆世出现以后, 在当时欧美大陆的北美东部地区一直比较繁荣, 并有比较连续的化石记录, 在世界其他地方只有零星的发现。受晚泥盆世F/F灭绝事件的影响, 其地理分布和生物多样性的发展经历了两次辐射演化和迁移扩散的高峰。第1次是在中泥盆世— 晚泥盆世早期, 颅形贝属的生物多样性达到了顶峰, 生物地理分布范围也扩大到了欧美大陆之外的西伯利亚板块、哈萨克斯坦板块和华南板块等。第2次是在早石炭世, 颅形贝属的生物多样性虽不及前一次高峰, 但其生物地理分布范围却是该属发展历史上最大的。早石炭世之后, 该属进入衰退阶段, 最终在二叠纪初灭绝。
1)在桂林地区下石炭统韦宪阶黄金组下部地层中发现的桂林颅形贝(Cranaena guilinensis sp.nov.)是华南地区韦宪阶中首次发现的颅形贝属分子, 也是目前已知的华南地区石炭纪有铰类腕足动物化石记录中唯一保存有放射状彩色条纹的化石记录; 该新种以平直的前结合缘和背壳上发育中槽为特征。保存有放射状彩色条带的桂林颅形贝的发现, 指示了黄金组下部的灰岩主要形成于温暖的浅海环境中。
2)对颅形贝属已知的70个种的地质地理分布和生物多样性变化的初步分析表明, 该属可能起源于早泥盆世欧美大陆西北缘的老世界区, 并以北美地区为中心, 经历了中泥盆世— 晚泥盆世早期和早石炭世2次比较明显的快速辐射演化和迁移扩散, 以及晚泥盆世晚期的1次严重衰退。在中泥盆世— 晚泥盆世早期的快速辐射演化中, 该属的生物多样性达到了巅峰, 生物地理分布范围也扩大到了欧美大陆之外的西伯利亚板块、哈萨克斯坦板块和华南板块等地; 在早石炭世的快速辐射演化中, 该属的生物多样性虽不及中泥盆世— 晚泥盆世早期, 但获得了最广泛的地理分布, 不仅分布在北方大陆, 而且进入到冈瓦纳大陆的边缘。晚泥盆世晚期的1次严重衰退显然与F/F灭绝事件有关。早石炭世之后, 该属进入衰退阶段, 最终在二叠纪初灭绝。
致谢 感谢美国Blodgett R.B.博士、Boucot A. J.教授和Isaacson P.教授及波兰Baliń ski A.教授提供相关的参考文献资料。广西区调院殷保安教授在野外工作中给予了指导, 笔者表示衷心的感谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|