
{kind=link}
{kind=link}
{kind=link}
辽西早白垩世离龙类地理地史分布及其演化
[张伟 , 高克勤]
, 高克勤]
, 高克勤]
|
|

通讯作者简介 高克勤,1955年生,北京大学长江学者,特聘教授,主要从事两栖、爬行动物演化研究。电话:010-62754883;E-mail:kqgao@pku.edu.cn。
第一作者简介 张伟,男,1989年生,2009年毕业于中国地质大学(武汉),目前于北京大学地球与空间科学学院攻读理学硕士学位,研究方向为古生物地层专业。电话:15296176469;E-mail:lingyuexiaofeng@126.com。
离龙类是一在形态和生态适应上高度特化的水生—半水生爬行动物双孔亚纲的灭绝支系。离龙目的地理分布局限于北半球,地史分布从侏罗纪一直延续到中新世。辽西是中国最为重要的离龙类化石产地,化石点主要分布于凌源、义县、朝阳和建昌 4个地区,尤其以义县地区化石点的分布最为密集。目前辽西已经发现的离龙类包括 3科 4属 5种,其中潜龙和满洲鳄 2属的化石仅发现于义县组,而伊克昭龙与戏水龙 2属的化石则仅见于九佛堂组。化石证据表明,白垩纪离龙类在辽西范围内的延展分布呈现一个自西向东、由南向北的地理地史演化过程。化石证据也表明辽西的离龙类在较短的时期和较窄的地域范围内经历了一个快速分异演化的过程。长颈潜龙类和短吻满洲鳄类在辽西同时首现于凌源义县组大新房子层,并先后灭绝于义县组的大康堡层和金刚山层。潜龙类最后出现的大康堡层的火山凝灰岩层密集保存了数以千计的潜龙类个体,可能代表该类群于早白垩世发生了与火山喷发活动紧密相关的集群灭绝事件。而后,短吻戏水龙和长吻伊克昭龙同时首现于九佛堂组第 2段,并于该组第 3段沉积时期内灭绝。整个辽西地域内的离龙类的分布从 126~120 Ma,在经历了大约 6 Ma的快速分异演化之后,均于早白垩世阿普第期灭绝。
About the corresponding author Gao Ke-Qin,born in 1955,is a professor of paleontology at the School of Earth and Space Sciences,Peking University. His current research focuses on the evolution of amphibians and reptiles based on fossils from the Mesozoic in China. E-mail:kqgao@pku.edu.cn.
About the first author Zhang Wei,born in 1989,graduated from the China University of Geosciences(Wuhan)in 2009. He is a master's graduated student at the School of Earth and Space Sciences,Peking University. His thesis research is in the field of paleontology. Tel:15296176469;E-mail:lingyuexiaofeng@126.com.
The Choristodera,an extinct and specialized clade of aquatic-semiaquatic diapsid reptiles,had an evolutionary history in close association with the continental ecosystems of Jurassic through Miocene on northern hemisphere. During the Early Cretaceous in western Liaoning Province of China,choristoderes went through a rapid evolution with diverse ecologic adaptations,as evidenced by their extensive fossil record from the Yixian and Jiufotang Formations. Choristoderan fossils have been found from localities within the Lingyuan,Yixian,Chaoyang and Jianchang areas,with a significant concentration of Hyphalosaurus localited at the Yixian area. The found Choristoderan fossies can be divided into five species of four genera in three families. In terms of geological range,fossils of Hyphalosaurus and Monjurosuchus are simply restricted to the Yixian Formation,while those of Ikechosaurus and Philydrosaurus are confined to the Jiufotang Formation. The extended distribution of the Early Cretaceous choristoderes in western Liaoning indicates a dispersal pattern of the group from west to east and from south to north. And last occurrence of Hyphalosaurus choristoderes in western Liaoning is recorded with thousands of fossil specimens preserved in tuffaceous deposits of the Dakangpu beds of the Yixian Formation. Such an unusual concentration of fossils indicates a possible mass extinction event of this clade during the Early Cretaceous Aptian Age. All choristoderes were wiped out from the western Liaoning during the Early Cretaceous Aptian Age.
离龙类(choristoderes)是生活在中生代到新生代的陆生爬行动物的一个灭绝支系, 主要营半水生— 水生生活(高克勤, 2007)。离龙类的化石最早发现于美国蒙大拿州晚白垩世地层中, 以零散的椎骨化石代表一类在体型和生态适应上都类似现生鳄鱼的史前爬行动物。这一史前爬行动物由美国古生物科学家Cope(1876)命名为鳄龙属(Champsosaurus), 并据此建立了离龙目(Choristodera)。目前全球范围已经明确地归入离龙目的化石代表11个属, 并可以粗略地归纳为3个生态— 形态类型, 其中包括有长吻型的鳄龙属(Cope, 1876)、西莫多龙属(SimoedosaurusGervais, 1877)、车尔龙属(TchoiriaEfimov, 1975)和伊克昭龙属(IkechosaurusSigogneau-Russell, 1981); 短吻型的栉颌鳄属(CteniogenysGilmore, 1928; Evans, 1989, 1990, 1991)、满洲鳄属(MonjurosuchusEndo, 1940; Gao et al., 2000)、复活鳄属(LazarussuchusHecht, 1992; Evans and Hecht, 1993)和戏水龙属(PhilydrosaurusGao and Fox, 2005); 以及高度适应水生生活的长颈型的潜龙属(HyphalosaurusGao et al., 1999)、庄川龙属(ShokawaEvans and Manabe, 1999)以及它们的近亲Khurendukhosaurus(Sigogneau-Russell and Efimov, 1984)。
离龙类在地史分布上最早可能的化石记录为英国晚三叠世的厚椎龙Pachystropheus(Storrs and Gower, 1993)。但是由于缺少关键的头部骨骼化石, 目前对于其能否确切归入离龙类还有较大疑问(Gao and Fox, 1998; Matsumoto and Evans, 2010)。目前确切的离龙类的最早化石记录, 为1990年发现于英国中侏罗世地层中的归入栉颌鳄属的化石(Evans, 1990), 而离龙类最晚的化石记录则为产自捷克共和国中新世地层中的归复活鳄属的化石(Evans and Klembara, 2005)。就此而言, 整个离龙类演化支系从侏罗纪到中新世经历了至少140 Ma的演化历史。再者, 如果英国晚三叠世的厚椎龙能够被确认归入离龙类, 那么这一史前爬行动物特殊类群的演化历史将会跨越近190 Ma。
离龙类研究在中国开展得相对较晚, 中国第1个正式在文献发表中归入离龙类的化石类型为孙氏伊克昭龙(Ikechosaurus sunailinaeSigogneau-Russell, 1981), 其化石标本产自内蒙古鄂托克旗的早白垩世地层, 起初曾被杨钟健(1964)鉴定为鳄鱼类始马来鳄属(Eotomistoma)。但是经过法国古生物科学家Sigogneau-Russell(1981)的重新研究, 根据其颌骨及牙齿特征, 将其重新命名为伊克昭龙并明确地将其归入了离龙类。然而, 中国离龙类最主要的化石发现还是在辽西。自上世纪90年代以来, 在辽西的凌源、义县等地相继发现了大量的离龙类化石, 为这一门类的演化研究提供了宝贵的原始材料。
辽西是早白垩世热河群(Jehol Group)的主要分布区域, 而著名的热河生物群(Jehol Biota)是中国乃至世界上化石门类最为齐全、保存状态最为完好的早白垩世生物群(Zhou et al., 2003)。自上世纪90年代以来, 一系列令人震惊的古生物化石发现由此而来, 其中又以带羽毛的恐龙和孔子鸟等最为著名(Xu et al., 2000; Ji et al., 2001; Norell et al., 2002; Chang et al., 2003; 季强等, 2004a), 而这些化石的发现有力地推动了兽脚类恐龙演化及鸟类起源的研究。离龙类是热河生物群的重要组成部分, 而在辽西早白垩世的湖泊生态系统中, 相对于狼鳍鱼等水生类群而言, 离龙类显然是其中的顶级捕食者(周忠和和王原, 2010)。通过离龙类的研究, 能够为恢复热河生物群的生态环境和整体生物群面貌提供良好的佐证。在辽西, 离龙类化石的个体数量和种属数目都比较丰富, 显示其在较短的时间和狭窄的地理范围内经历了快速分异演化的过程。而在北美和欧洲, 早白垩世的离龙类的化石记录则较为稀少。就此而言, 辽西离龙类的研究进展揭示了离龙类演化历史中的一个重要环节。
离龙类在世界范围内的宏观地理地史分布已有先前研究者(高克勤, 2007; Matsumoto and Evans, 2010)作了详细研究, 但具体到辽西范围内离龙类的地理地史分布目前还缺乏一个细致的地域性的分析。作者在目前已有化石资料的基础上对辽西离龙类的地理地史分布进行分析研究讨论。与此同时, 根据不同的生态— 形态型的离龙类在地理和地史上的分布形式, 探讨其在辽西地域范围内快速分异演化的过程。
离龙类的研究在中国开展相对较晚。虽然隶属离龙类的楔齿满洲鳄(Monjurosuchus splendensEndo, 1940)的化石在上世纪30年代末就已经在辽西凌源发现, 但原始研究中将其归入与恐龙和鳄鱼类相关的槽齿类(Thecodontia)。另外, 由于正型标本在抗日战争期间遗失, 满洲鳄真正与离龙类的真实隶属关系经历了60年之后, 直到本世纪初随着新标本的发现才得以确认。Gao 等(2000)基于与楔齿满洲鳄正型标本同一层位发现的新标本的研究, 正式将满洲鳄归入了离龙目。到目前为止, 辽西范围内已经发现并有效命名的离龙类包括3科4属5种(表1)。其中凌源潜龙(Hyphalosaurus lingyuanensisGao et al., 1999)是中国发现的第1个产自中生代湖相沉积中的长颈型水生爬行动物, 而朝阳戏水龙(Philydrosaurus proseilusGao and Fox, 2005)则代表一个与楔齿满洲鳄紧密相关的短吻中小型半水生类型。
| 表1 辽西离龙类的系统分类 Table1 Classification of choristoderes from western Liaoning Province |
另外, 高春玲等(2005)命名过一个归入西莫多龙科的新属种朝阳辽西龙(Liaoxisaurus chaoyangensis), 其报道的化石地点和层位为辽宁省朝阳市义县头道湾乡毛家沟村九佛堂组。但是这一产出地点是存在问题的:在义县并不存在头道湾乡和毛家沟村, 义县也并不归朝阳市管辖, 出现这一问题的原因很有可能是原作者的笔误, 而化石标本真实的产地有待进一步考证。而且产自九佛堂组的朝阳辽西龙与先前已经命名发表的皮夹沟伊克昭龙在解剖形态上极为相似, 鉴定特征上难以区分。因此, 朝阳辽西龙作为一个独立支系的属种命名的有效性目前是存在疑问的(Matsumoto and Evans, 2010)。
就离龙类的生态— 形态适应类型而言, 辽西发现的楔齿满洲鳄和朝阳戏水龙为半水生短吻生态— 形态类型, 潜龙类为高度适应水生的长颈型离龙类, 而伊克昭龙则为类似现生长吻鳄的长吻生态— 形态类型。这反映出早期演化阶段中, 离龙类在辽西白垩纪湖沼深水的环境中分化出了不同的食性生态适应, 是离龙类快速分异演化的结果。此外, 近年中关于辽西离龙类的报道还包括游泳脚蹼在内的皮肤特征的保存(Gao et al., 2000), 软壳蛋胚胎(季强等, 2004b; Gao and Ksepka, 2008; Hou et al., 2010), 异常的嗜食同类生态行为(汪筱林等, 2005)以及卵胎生的生殖方式(Ji et al., 2010)等。这些发现为离龙类的深入研究提供了宝贵的材料, 同时揭示出了离龙类在亚洲早期演化的重要信息。
辽西所有的离龙类化石均发现于下白垩统义县组与九佛堂组。然而, 离龙类化石在义县组与九佛堂组的属种分布截然不同, 其中满洲鳄属、潜龙属目前只发现于义县组, 而戏水龙属和伊克昭龙属则仅发现于九佛堂组(图 1)。到目前为止, 辽西离龙类产出的主要化石沉积层有义县组内的罗家沟层、大新房子层、大康堡层、金刚山层和九佛堂组内的皮家沟层、上河首层、喇嘛沟层、原家洼层。
 | 图1 辽西离龙类化石层对比Fig.1 Stratigraphic correlation of choristodere-bearing beds in western Liaoning Province |
辽西地区义县组和九佛堂组含化石层位的地史时代曾经颇有争议。自上世纪80年代以来, 前人曾经用Rb-Sr、K-Ar、U-Pb、 40Ar/39Ar 等多种同位素测年方法做出过几十个年龄值, 数据从147 Ma到109 Ma不等(季强等, 2004a; 王五力等, 2004)。近期研究中, Chang 等(2009)对辽西义县组年代做了较为详细的分析, 根据其高精度的 40Ar/39Ar 同位素测年的结果, 义县组从底部到接近顶部的绝对年龄应该在129.7± 0.5~122.1± 0.3 Ma之间, 其地质年代应该为早白垩世巴列姆期(Barremian)到阿普第期(Aptian)早期。九佛堂组的同位素测年数据则相对较少, He 等(2004)根据朝阳上河首岩样得出的 40Ar/39Ar 同位素年龄约为120 Ma, 其地质年代应该为阿普第期早期。从前人的地层对比结论来看, 义县组的大新房子层和罗家沟层可能是辽西所有含离龙类化石层中最下部的层位, 在区域上可以与北票尖山沟层、义县砖城子层相对比(季强等, 2004a; 王五力, 2004)。其中大新房子层最早由陈丕基等(1980)建立于辽宁凌源小城子乡大新房子一带, 其分布向南延伸经过范杖子、大王杖子至帽子山。大新房子层是热河生物群化石产出的重要层位, 绝对年龄值为123~126 Ma(季强等, 2004a; 王五力, 2004; 张宏等, 2006)。凌源地区的楔齿满洲鳄和凌源潜龙化石均产自该层位。而罗家沟层由季强等(2004)建立于建昌县要路沟乡罗家沟村, 主要分布于要路沟、罗家沟及魏家岭一带。除离龙类之外, 罗家沟层还产有鱼类、鸟类、龟类、恐龙、昆虫、介形类、叶肢介、植物等化石(张立君等, 2012)。有关建昌地区的同位素测年数据较少, 只有郑月娟等(2011)对建昌盆地义县组上部英安岩岩样的 40Ar/39Ar 同位素分析, 得出的年龄约为120 Ma, 而建昌地区义县组下部的罗家沟层, 目前还没有同位素测年的数据。
关于义县地区的白台沟潜龙化石的分布层位曾存在2种不同的意见。季强等(2004b)将白台沟潜龙的发现层位记述为下白垩统九佛堂组。同年, 王五力等(2004)通过对辽西义县阶地层的详细研究, 将白台沟潜龙的发现层位归入了义县组, 并且确切将其限定为大康堡层。此后的文献中, 分别引用了上述的2种观点(Gao and Ksepka, 2008; Hou et al., 2010; Ji et al., 2010; Matsumoto and Evans, 2010)。然而, Ji 等(2011)根据白台沟潜龙的化石原产层位的 206Pb/238U同位素测年结果, 破台子一带化石层位的绝对年龄应该在126.47± 0.87 Ma, 从而改变了其产自九佛堂组的观点, 因此白台沟潜龙出自义县组目前是被广为接受的。大康堡层由陈丕基等(1980)建立, 是义县地区大凌河两岸的王家沟— 破台子— 王油匠沟— 白台沟一带的众多离龙类化石点的主要层位, 其层序关系处于义县组砖城子层与朱家沟层之间(王五力等, 2004)。除潜龙类外, 大康堡层还出产有禽龙类恐龙锦州龙(JinzhousaurusWang and Xu, 2001)、翼龙类、龟类、鸟类、叶肢介、昆虫、虾及植物化石等, 而在整个辽西广泛分布的狼鳍鱼化石目前在大康堡层并没有相关的报道。
整个义县组最上部的一个出产离龙类化石的沉积层为金刚山层, 由室井渡(1940)首先建立于辽宁义县金刚山— 枣茨山一带, 也是研究热河生物群的经典层位之一。金刚山层出产的离龙类化石仅有满洲鳄一类, 没有潜龙类或者其他离龙类。除了离龙类之外, 金刚山层还产有蜥蜴类矢部龙(YabeinosaurusEndo and Shikama, 1942)、翼龙类、满洲龟、反鸟类、鱼类、叶肢介、昆虫、木化石等。金刚山层的具体年龄, 前人曾做过不少研究, 包括Smith 等(1995)、季强等(2004a)、王五力等(2004)和张宏等(2005)均对其做过同位素测年。综合其研究结果, 金刚山层的绝对年龄应该在122 Ma左右。
在义县组内广泛分布的满洲鳄和潜龙2属并没有延续到九佛堂组, 而九佛堂组内发现的离龙类化石被分别归入了戏水龙属和伊克昭龙属。从地层对比的结果来看, 朝阳地区九佛堂组第1段的上河首层和喇嘛沟层可能分别是戏水龙和伊克昭龙两属分布的底限, 而朝阳地区九佛堂组第3段的原家洼层和义县地区的皮家沟层则分别为戏水龙和伊克昭龙属地史分布的上界, 同时也是整个辽西地域内离龙类延展分布的上限, 在此之上的阜新组至今一直没有离龙类化石的报道。从更广的区域上来看, 九佛堂组远非离龙类在亚洲分布的上界, 离龙类在亚洲的演化历史可以延伸到古近纪。Averianov(2005)从哈萨克斯坦的古新世— 始新世地层中报道过西莫多龙属未定种(Simoedosaurus sp.)的化石发现。而在中国境内, 内蒙古孙氏伊克昭龙的产出层位罗汉洞组在地质时代上与九佛堂组相近, 不过目前尚缺乏可用的同位素测年数据, 因此中国离龙类的分布上界有待进一步的研究和探讨。
除了离龙类之外, 朝阳附近九佛堂组上河首层出产有大量的鱼类、鸟类、兽脚类恐龙、翼龙类等脊椎动物化石, 喇嘛沟层出产有鱼类、鸟类、翼龙类和龟类化石, 原家洼层则出产鱼类、恐龙类、鸟类和翼龙类化石等。义县附近九佛堂组皮家沟层产有鱼类化石。
综上所述, 离龙类化石在辽西的地层分布仅限于下白垩统义县组和九佛堂组。从126~120 Ma的地质时限中, 离龙类在辽西至少经历了约6 Ma的演化历史, 并且快速地分异演化出3个不同的生态— 形态类型(见下文)。具体而言, 满洲鳄和潜龙类的化石几乎同时出现于义县组下部的大新房子层。从这2个属的离龙类在义县组内的延续时间来看, 满洲鳄要稍长于潜龙类(图 1)。潜龙类在义县组内最后的化石记录出现于义县地区的大康堡层, 极为繁盛, 出产有数以千计的标本。在此之上的主要化石沉积层金刚山层则仅出产满洲鳄, 没有潜龙类化石的发现。而且不仅义县地区, 整个辽西地区之后就再也没有潜龙类的记录。由上可知, 辽西的满洲鳄和潜龙类均在义县组的沉积时限内灭绝, 具体时代应该在早白垩世阿普第期早期, 而潜龙类先于满洲鳄灭绝。与此不同, 伊克昭龙和戏水龙的化石则几乎同时出现于九佛堂组第2段, 并且共同于九佛堂组第3段的沉积时限内灭绝, 延续时间近乎相等。整个辽西的离龙类类群均在早白垩世阿普第期灭绝。
辽西早白垩世离龙类化石的地理分布具有一定的规律, 在相当程度上可能反映出离龙类在早白垩世辽西地区的生存和分布状况。目前辽西范围内发现的离龙类化石点分布在凌源、建昌、朝阳和义县4个地区(图 2, 图3)。其中明显地又以义县地区的化石点分布最为密集, 而凌源、朝阳、建昌等地的离龙类化石点则相对较为稀少, 同时各化石点的相对位置分布也较为离散。如前所述, 辽西热河生物群的发现和研究始于20世纪40年代。经过了70多年的研究, 尤其是近20年来辽西化石的大量发现使之有了很高的研究程度。就此而言, 目前已知的辽西离龙类的地理分布不应存在重要的化石点遗漏的情况。换言之, 凌源、建昌、朝阳和义县地区的化石发现, 应该可以反映出早白垩世离龙类在辽西的基本分布格局。同时, 义县组和九佛堂组离龙类在化石点分布上的差异可以从一定程度上反映出早白垩世辽西离龙类在演化过程中的地理分布上的变化。
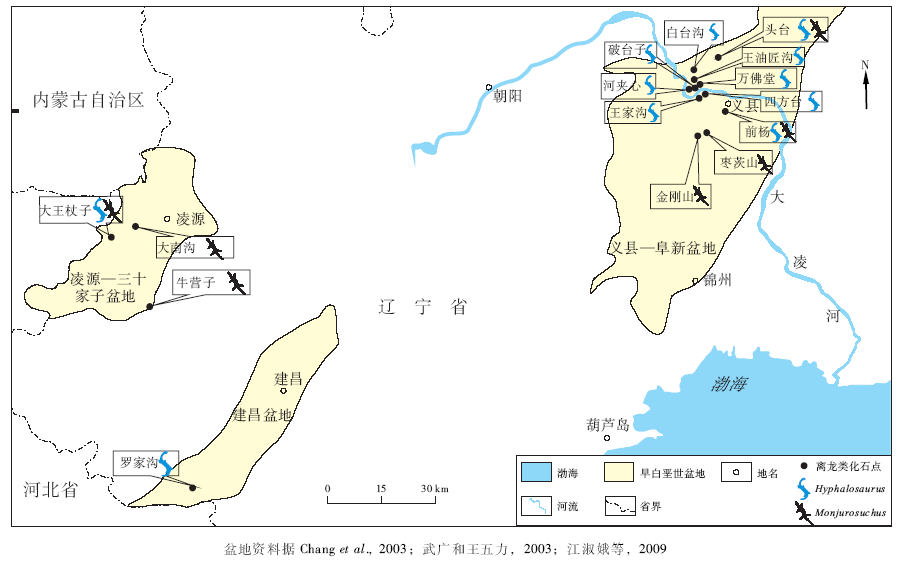 | 图2 辽西义县组离龙类化石地理分布Fig.2 Geographic distribution of choristoderan fossil sites of Yixian Formation in Cretaceous basins of western Liaoning Province |
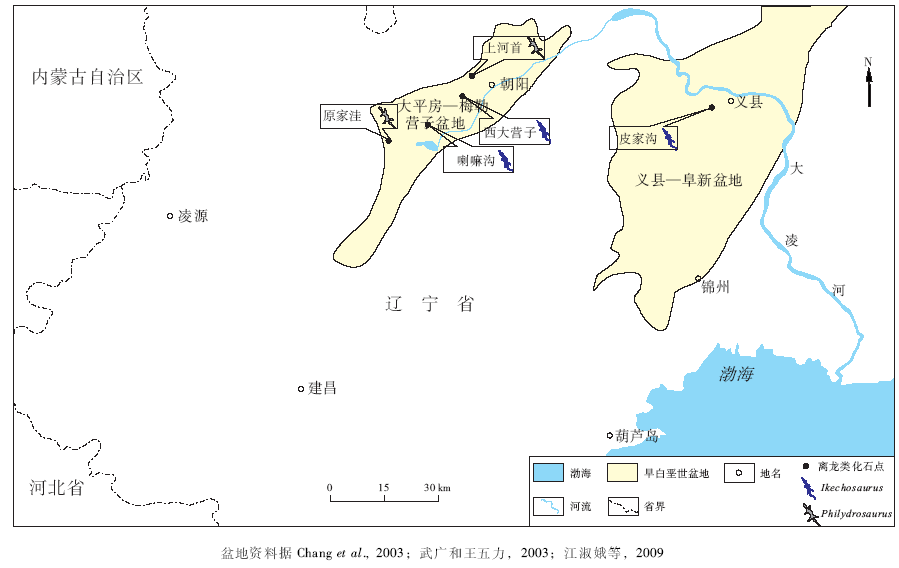 | 图3 辽西九佛堂组离龙类化石地理分布Fig.3 Geographic distribution of choristoderan fossil sites of Jiufotang Formation in Cretaceous basins of western Liaoning Province |
辽西离龙类的化石层多分布于早白垩世构造盆地之中, 分别为凌源— 三十家子盆地、建昌盆地、义县— 阜新盆地和大平房— 梅勒营子盆地。
具体而言, 地质时代较早的义县组离龙类的化石点数量明显较九佛堂组为多, 且分布更为广泛, 主要分布在凌源、建昌和义县3个地区, 分别位于凌源— 三十家子盆地、建昌盆地和义县— 阜新盆地3个构造盆地之中。
早期义县组化石点主要分布于辽西西部的凌源和建昌。其中凌源— 三十家子盆地是热河生物群化石发现和研究的经典地域。楔齿满洲鳄的化石最初就于20世纪30年代发现于凌源西南10.5 km处的大南沟(Endo, 1940)。此外, 已经发现离龙类化石点还有凌源大王杖子(凌源西南约20 km, Gao et al., 1999; 汪筱林等, 2000)和牛营子(凌源南约27 km, Gao et al., 2000)。凌源— 三十家子盆地中产出离龙类化石的沉积层为义县组大新房子层, 为一套火山碎屑— 湖相化石沉积层, 岩性主要为灰色、灰黑色页岩夹黄褐色、黄绿色凝灰岩(汪筱林等, 2000)。大王杖子化石点产出有凌源潜龙和满洲鳄, 牛营子化石点则只出产有满洲鳄化石。除离龙类外, 在凌源大王杖子化石点的重要发现还有最早的有胎盘哺乳类攀援始祖兽(Eomaia scansoriaJi et al., 2002), 带羽毛的恐龙千禧中国鸟龙(Sinornithosaurus milleniiJi et al., 2001), 最早的被子植物之一的中华古果(Archaefructus sinensisSun et al., 2002)等。目前凌源— 三十家子盆地中大王杖子等地发现的主要脊椎动物化石门类有鱼类、离龙类、有鳞类、翼龙类、奔龙类、鸟类、哺乳类(汪筱林等, 2000; Chang et al., 2003; 季强等, 2004a; 王五力等, 2004)(表 2), 其中只有鱼类和离龙类属于水生生活的化石代表。
| 表2 辽西离龙类主要化石点化石类群对比 Table 2 Comparison of taxonomic components among choristoderan fossil sites in western Liaoning Province |
建昌盆地的离龙类化石报道时间相对较晚。Duan和Zhang(2009)首先报道了从建昌县的魏家岭— 要路沟一带的义县组罗家沟层发现的潜龙类化石。之后张立君等(2012)又对要路沟一带做了详细的地层描述, 在其对上白庙— 罗家沟北山剖面的地层描述中报道有潜龙属未定种(Hyphalosaurus sp.)的化石发现。化石层岩性为湖相深灰色薄层泥质粉砂岩、粉砂质泥岩夹黄灰色凝灰质页岩, 具水平层理。除了离龙类之外, 建昌罗家沟还发现有恐龙、鸟类等多门类的化石(张立君等, 2012)。不过, 该地区的离龙类研究目前还处于半空白的阶段, 已经发现的化石还有待进一步的研究和考证。
晚期义县组的离龙类化石点主要分布于辽西东部义县地区的早白垩世义县— 阜新盆地之中。其中大凌河两岸一系列的化石产地包括白台沟(义县西北约13 km)、四方台(义县西北约7 km)、破台子(义县西北约10 km)、王油匠沟(义县西北约11 km)、王家沟(义县西北约8.5 km)、河夹心(义县西北约11 km)、万佛堂(义县西北约10 km)等化石点。这些位于盆地西北部的化石点仅出产有潜龙类, 而无其他离龙类化石(Chang et al., 2003; 季强等, 2004a; 王五力等, 2004)。化石沉积层为义县组大康堡层, 岩性为湖相的浅灰白色钙泥质粉砂岩、泥质页岩与灰白色含钙粉砂岩互层(季强等, 2004a; 王五力等, 2004)。与此不同的是, 地理位置位于义县— 阜新盆地中部的枣茨山、金刚山(义县西南约22 km)只出产有楔齿满洲鳄化石(Endo and Shikama, 1942; 王五力等, 2004; Gao and Fox, 2005; 汪筱林等, 2005)。化石沉积层为金刚山层, 主要岩性为湖相的灰、灰绿色粉砂质泥页岩与灰白色泥晶灰岩互层(王五力等, 2004)。而义县附近的前杨(义县南2 km, Ji et al., 2011)和头台(义县北约18 km, 季强等, 2004a; Gao and Fox, 2005)则同时出产有满洲鳄和潜龙化石。除了离龙类之外, 义县地区还发现有鱼类、反鸟类、禽龙类、翼龙类、满洲龟等化石(Chang et al., 2003; 季强等, 2004a; 王五力等, 2004), 其中包括有金刚山产出的翼龙蛋及胚胎(Wang and Zhou, 2004)和鸟类胚胎化石(Zhou and Zhang, 2004), 河夹心化石点产出的无尾类纤细宜州蟾(Yizhoubatrachus macilentusGao and Chen, 2004)。
相对义县组而言, 地质时代相对较晚的九佛堂组产出离龙类的化石点较少, 主要分布于义县和朝阳附近, 分别位于义县— 阜新盆地和大平房— 梅勒营子盆地之中。在大平房— 梅勒营子盆地中发现有3个化石点, 其中的朝阳市龙城区上河首化石点(Gao and Fox, 2005)属于上河首层, 而东大道喇嘛沟(段冶等, 2006)和西大营子西北沟(杨雅军等, 2006)的化石沉积层则属于喇嘛沟层。上河首层岩性主要为湖相的灰绿色泥岩与页岩互层, 夹有火山灰(Zhang et al., 2007), 产有离龙类戏水龙属的化石。而西大营子西北沟和东大道喇嘛沟的岩性为湖相的灰色、灰白色粉砂质泥岩、粉砂岩夹泥灰岩、沉凝灰岩、页岩和薄层砂岩。这一层位中出产有离龙类伊克昭龙化石(段冶等, 2006)。
大平房镇原家洼(朝阳市西南约30 km, Gao et al., 2007)和义县皮家沟化石点(义县西约5.7 km, Liu, 2004)的化石沉积层的地质时代则相对较晚, 沉积层分别为原家洼层和皮家沟层。其中大平房镇原家洼化石点出产的离龙类为朝阳戏水龙(Gao et al., 2007), 沉积层岩性为湖相灰色、深灰色粉砂质泥岩、页岩、粉砂岩夹泥灰岩和沉凝灰岩, 具水平层理(段冶等, 2006)。义县皮家沟化石点则发现有皮家沟伊克昭龙, 产自湖相的灰色泥岩、页岩、粉砂岩的皮家沟层(王五力等, 2004)。
另外除了离龙类之外, 在朝阳地区的九佛堂组地层中还发现有大量的鸟类、鱼类、兽脚类恐龙、翼龙类化石(Chang et al., 2003; 季强等, 2004a; 王五力等, 2004), 其中尤其以原始热河鸟(Jeholornis prima; Zhou and Zhang, 2002), 赵氏小盗龙(Microraptor zhaoianus; Xu et al., 2000)最为著名。义县皮家沟化石点还产出有鱼类化石。
由上可知, 凌源、义县、朝阳以及建昌这4个地区的主要化石点所出产的离龙类化石多有差异。从属种的分布上来看, 目前已发现有楔齿满洲鳄的化石点有凌源地区的大王杖子和牛营子, 义县地区的头台、前杨、金刚山和枣茨山。凌源潜龙和白台沟潜龙分别发现于凌源地区的大王杖子和义县地区头台乡白台沟村, 除了原始的发现地之外, 潜龙类广泛分布于义县附近的大凌河两岸的许多地点, 具体包括前杨、四方台、破台子、王油匠沟、王家沟、河夹心、万佛堂等地。另外, 在建昌的罗家沟亦曾经有过未曾描述的潜龙类化石的发现。皮家沟伊克昭龙目前唯一的化石发现于义县地区的皮家沟。此外, 段冶等(2006)、杨雅军等(2006)还报道过在朝阳县喇嘛沟和西大营子西北沟的九佛堂组中发现伊克昭龙未定种(Ikechosaurus sp.)化石。朝阳戏水龙目前仅发现于朝阳地区, 其化石点分别在朝阳市龙城区的上河首和大平房镇原家洼。
从不同生态— 形态类型的离龙类的地理分布上可明显看出, 长颈型潜龙类的分布最为广泛, 从辽西离龙类化石点地理纬度上最北的头台(41° 38'42″N, 121° 12'18″E), 到最南端的罗家沟(40° 34'58″N, 119° 29'42″E), 从最东端的前杨(41° 30'54″N, 121° 13'48″E)到最西部的大王杖子(41° 11'24″N, 119° 13'48″E), 都有长颈离龙类的化石的发现。而短吻型离龙类次之, 以长吻型离龙类的分布范围最为狭窄, 目前确切的化石点只有义县皮家沟和朝阳喇嘛沟。而从化石标本的个体数量上来看, 长颈离龙类的化石也是最多的, 由此可知, 长颈潜龙类很有可能是辽西早白垩世3个生态— 形态类型的离龙类中最为繁盛的一个类群。
在辽西发现的3个生态— 形态类型的离龙类之中, 地理分布跨度最大的是长颈潜龙类, 但是从地史延续分布上来看, 长颈离龙类的延续时间要短于中小型短吻离龙类, 后者是辽西唯一一个在义县组和九佛堂组均有化石产出的生态— 形态类型。辽西长吻生态— 形态类型的离龙类(伊克昭龙属)在地史上的出现时间最晚, 同时其地理地史分布也仅限于朝阳和义县地区的九佛堂组。但是在辽西地域之外, 长吻类型在亚洲的演化历史一直延续到哈萨克斯坦的古新世— 始新世(Averianov, 2005), 而在欧美长吻型离龙类的辐射演化始于晚白垩世, 其时代分布的上限止于古新世(Gao and Fox, 1998; Matsumoto and Evans, 2010)。
在辽西地域范围内, 无论是长颈潜龙类抑或是短吻生态— 形态类型的离龙类, 其地史时代相对较老、层位相对较低的化石点多位于辽西西部和南部的凌源与建昌地区, 而地史时代相对较新、层位相对较高的化石点则多位于辽西东部和北部的义县地区和朝阳地区。在辽西地史分布出现最晚的长吻离龙类则仅见于义县和朝阳地区。另外如图2和图3所示, 辽西离龙类从义县组到九佛堂组沉积时限的演化过程中, 其地理分布有一个较为明显的收缩, 到最后仅分布于辽西东部和北部的义县与朝阳的稀少化石点。这说明早白垩世离龙类在辽西范围内总体上呈现一个自西向东、自南向北的延展分布过程。换言之, 适合于离龙类生存的生态环境的发育可能是一个自西向东、自南向北的发展过程。而这一现象的成因机制, 需要有构造地质学、沉积学以及多门类古生物学的综合研究, 不是单一离龙类的地史地理分布可以解释的, 有待后人进一步的分析和研究。
从标本的数量上来看, 高度适应水生的长颈型离龙类的化石已有数千件标本发现, 无疑是所有离龙类中标本数目最多的。体型较大的湖沼长吻型的标本记录则显然是辽西3个类型中最少的, 而半水栖的短吻型的标本数量则处于一个中等的水平。这在一定程度上应该能够反映出当时3个生态型的离龙类在个体数量上的差异。另外, 依照现有的热河生物群的化石发现来看, 整个辽西湖泊生态系统中的顶层捕食者应该是离龙类, 其中又以体型较为庞大的伊克昭龙为最(周忠和和王原, 2010)。辽西并没有发现鳄鱼类的化石, 在辽西的湖泊生态系统中显然并没有能够与离龙类进行生态位竞争的顶层捕食者, 再加上温暖适宜的气候和食源较为丰富的水体环境, 这些可能构成了离龙类在早白垩世辽西地域范围内快速分异演化的基本生态条件, 并在较短的地史时限(6 Ma)和较小的地理范围内分化出了3个不同的生态— 形态类型。
虽然长颈类型的潜龙类就个体数目而言可能是辽西乃至世界范围上最为繁盛的离龙类群, 但就离龙类的全球分布而言, 高度适应水生生活的长颈类型无疑是3个生态— 形态类型中地理分布范围最为狭窄的, 也是亚洲特有的一个本地支系(endemic clade)。已经有效命名的长颈类离龙类仅包括辽西的潜龙和日本的庄川龙2个属, 另外与之相关的还可能有发现于蒙古和西伯利亚的Khurendukhosaurus(Sigogneau-Russell and Efimov, 1984; Skutschas, 2008)。这表明长颈型离龙类的地理分布目前仅局限于亚洲东部。与此不同, 无论是长吻离龙类还是中小型短吻离龙类, 其地理分布均跨越欧亚和北美, 广泛分布于劳亚大陆(Matsumoto and Evans, 2010)。在地史分布方面, 长吻型离龙类的演化历史从早白垩世延伸到古新世, 跨越了超过60 Ma的地史时间。短吻中小型离龙类的地史分布最长, 从中侏罗世到中新世跨越了超过130 Ma的地史时间。相比之下, 长颈型离龙类的地史分布局限于早白垩世巴列姆期— 阿普第期早期, 显然是3个生态— 形态类型中地史分布最短的。在辽西, 潜龙类大量幼年个体化石在义县地区大凌河两岸的众多化石点的密集发现, 可能暗示该地区在当时的湖泊中是潜龙类的一个繁殖水域。与此同时, 数量众多的潜龙类化石高度密集地埋藏于辽西大康堡层火山凝灰岩层之中, 也可能意味着长颈离龙类在当时当地发生了与火山喷发相关的集群灭绝事件。而辽西九佛堂组之上的地层中再也没有长颈潜龙类化石的发现。
从Cope(1876)的最早发现开始, 离龙类的化石发现和研究已经有近140年的历史。长期的化石发现和研究表明, 离龙类在中生代乃至新近纪的陆生生态系统中是一个不可忽视的水生— 半水生生态类群。随着20世纪末以来一系列的化石发现与研究, 对于离龙类各支系的解剖特征和演化关系的认识都有了相当程度的进展, 但是包括满洲鳄科(Monjurosuchidae)和潜龙科(Hyphalosauridae)在内的中小型离龙类的演化关系却依旧存在较大的争议。包括Cteniogenys和Khurendukhosaurus在内的一些分类单元所依据的多为保存不佳的零散化石, 依此所做的复原多存在不小的疑问, 因此在较为完整的化石发现之前, 想要彻底解决离龙类内部各演化支系间亲缘关系问题, 显然是比较困难的。目前能够确认的是由包括有鳄龙科和西莫多龙科在内的大型长吻离龙类及其内各属的演化关系, 而其余离龙类各科、属之间的演化关系, 还有待于进一步的分析和研究。
1)辽西的离龙类化石点主要分布于凌源、建昌、朝阳和义县4个地区, 分别位于早白垩世的凌源— 三十家子盆地、建昌盆地、大平房— 梅勒营子盆地和义县— 阜新盆地之中。
2)辽西离龙类化石仅发现于义县组与九佛堂组, 其中满洲鳄和潜龙类仅发现于义县组, 而戏水龙和伊克昭龙仅见于九佛堂组。地史分布表明离龙类在辽西至少经历了约6 Ma的演化历史, 其延续时代为早白垩世巴列姆期至阿普第期早期。
3)在辽西发现的3个生态— 形态类型的离龙类中, 以长颈型离龙类在辽西的地理分布最为广泛, 不过其在义县组内的延续时限要稍短于短吻型的满洲鳄。
4)早白垩世离龙类在辽西范围内总体呈现一个自西向东、自南向北的延展分布过程, 而在义县组到九佛堂组的沉积时限的演化过程中, 离龙类在辽西的分布范围明显收缩。
5)长颈型离龙类在早白垩世的阿普第期早期, 在辽西可能发生了与火山活动紧密相关的集群灭绝事件, 而这一事件最终导致了长颈离龙类在辽西化石记录中的绝迹。
致谢 作者感谢沈阳师范大学张立君教授提供地层剖面及化石分布方面的资料, 感谢周长付教授在离龙类及其化石分布方面提供的参考和帮助。北京大学地球与空间科学学院刘建波教授曾对本文提出过颇具建设性的修改建议, 在此表示衷心感谢。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|