



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
晋冀低山丘陵区人工扰动植被花粉组合特征
[李洁1, 2, 3  , 许清海
, 许清海1, 2, 3 , 曹现勇1, 2, 3, 4, 5 , 田芳4, 5 , 丁伟6 , 梁剑2, 3 ]
, 许清海, 曹现勇|
|

通讯作者简介 许清海,男,1950年生,河北师范大学教授,主要从事孢粉学与全球气候变化研究。通讯地址:河北省石家庄市河北师范大学泥河湾考古研究院;邮政编码:050024。E-mail:xuqinghai@mail.hebtu.edu.cn。
第一作者简介 李洁,女,1989年生,河北师范大学硕士研究生,主要从事孢粉学与全球气候变化研究。通讯地址:河北省石家庄市河北师范大学泥河湾考古研究院;邮政编码:050024。E-mail:lijie2011geo@126.com。
54个晋冀低山丘陵区人工和人工扰动植被表土花粉样品分析表明:花粉组合以草本植物花粉( 74.5%)为主,乔木花粉含量低于 20%。人类活动较强的农田以谷物禾本科、蒿属和藜科花粉为主,人类活动较弱的荒地以蒿属、藜科和杂草禾本科花粉为主。人类活动强度不同,指示种不同,孢粉浓度亦不同;农田指示种为谷物禾本科、葡萄科和胡桃科,孢粉浓度约 4380粒 /g;而蒿属、蓼科和蔷薇科则为荒地指示种,孢粉浓度约 10983粒 /g。农作物种植方式不同,谷物禾本科花粉含量不同,单一禾本科作物种植区谷物禾本科花粉含量( 40.2%)高于多种作物混作区( 18.8%)和荒地( 3.8%)的含量。谷物禾本科含量的变化可以为利用化石花粉提取和检测人类活动信息提供依据,也可为利用地层中谷物禾本科花粉判断古代农业活动提供依据。
About the corresponding author Xu Qinghai,born in 1950,is a professor in Hebei Normal University. He is mainly engaged in researches of palynology and global climate change. E-mail:xuqinghai@mail.hebtu.edu.cn.
About the first author Li Jie,born in 1989,is a master candidate of Hebei Normal University. She is mainly engaged in researches of palynology and global climate change. E-mail:lijie2011geo@126.com.
The pollen assemblages of 54 surface samples from human disturbed vegetation in hilly areas of Shanxi and Hebei Provinces show that:Herbaceous pollen types are dominant in pollen assemblages(average 74.5%),arboreal pollen percentages are lower than 20%. Cereals, Artemisia,and Chenopodiaceae are the major herbaceous pollen types in farmlands where human impacts are much stronger. While Artemisia,Chenopodiaceae and Poaceae are the major herbaceous pollen types in wastelands. Different human activity intensities lead to different indicator species and different pollen concentrations. Cereals,Vitaceae and Juglandaceae are the indicator pollen types of farmlands whose pollen concentrations are about 4380 grains/g,while Artemisia,Polygonaceae and Rosaceae are the indicator pollen types of wastelands whose pollen concentrations are about 10983 grains/g. Different agricutural practices produce different pollen types with different pollen content of cereals. The pollen percentages of cereals are higher(40.2%)in single cereal areas than those in mix-planted areas(18.8%)and wastelands(3.8%). The pollen contents of cereals in different crops will give a reference to obtain the messages of human activities in lands as well as to distinguish the possible old cultivation methods.
孢粉是研究全球变化的重要代用指标之一, 主要用于古气候重建及古植被模拟(许清海等, 2004a, 2004b; 孙雄伟和夏正楷, 2005)。建立表土花粉与气候的关系, 是依据孢粉来定量恢复古植被、古气候的基础(张玉兰, 2006; 王学丽等, 2008)。以往表土孢粉研究主要选择人类活动干扰较少、自然植被保存较好的地区探讨孢粉— 植被— 气候的关系(秦大河, 2005; 朱诚等, 2006), 但工业革命以来, 人类活动对气候和植被的影响越来越显著, 预测未来气候和植被变化, 除要考虑自然变化的规律和特征外, 还需考虑人类活动对气候和植被的影响。因此越来越多的孢粉工作者开始关注人类活动影响下的表土孢粉研究(Hjelle, 1997; 邓韫等, 2002; 李月丛等, 2003, 2005; 许清海等, 2004b, 2006; Buttler and de Beaulieu, 2005; Court-Picon et al., 2005; Mazier et al., 2006), 以检测人类活动对气候和植被的影响。
华北地区人类活动历史悠久, 人口稠密, 自古以来就是中华民族先人活动的重要地区(史金波, 2011), 是受人类活动影响强烈的地区之一。区域内地带性植被经过长期大规模人类活动的影响, 发生了巨大变化, 大部分丘陵地区的原有植被已被人工或人工扰动后的次生植被替代, 平原区则完全被人工植被替代(上官铁梁, 1985; 《山西森林》编辑委员会, 1992; 《河北植被》编辑委员会和河北省农业区划委员会办公室, 1996)。丘陵区的孢粉组合既受农作物影响, 又受人工扰动次生植被影响, 其孢粉组合可能与平原区人类活动破坏植被的初始阶段相近(丁伟等, 2011), 因而研究低山丘陵区人工与人工扰动次生植被孢粉组合特征对检测早期人类活动具有重要意义, 并且取得了一定的成果(刘鸿雁, 2002; 庞瑞洺等, 2010; 杨士雄等, 2010; 丁伟等, 2011; 黄小忠等, 2011; 李曼玥等, 2012), 但前人研究未能明确指出能够指示人类不同活动强度下的花粉类型。因此, 本研究以河北省西部、山西省北部及西部低山丘陵区表土样品为研究对象, 结合多响应置换分析和指示种分析方法, 探讨人工和人工扰动次生植被孢粉组合特征, 揭示对人类活动影响强度有指示意义的花粉类型和花粉组合特征, 为利用化石孢粉资料探讨人类活动对区域古植被和古气候的影响、提取和检测人类活动信息提供依据。
晋冀低山丘陵区位于中国北部(36° 20'~40° 8N, 111° 14'~114° 48'E, 海拔87~1707 m), 行政区主要包括河北省西部、山西省北部及西部, 地貌区划主要有冀西北山间盆地、大同盆地、吕梁山地等(张新时, 2007)。气候为温带大陆性季风气候, 年均气温5~13 ℃, 年均降水量400~600 mm, 降水主要集中在7~8月份(焦北辰, 1984)。研究区自然植被主要为暖温带落叶阔叶林, 以油松(Pinus tabuliformis)、栓皮栎(Quercus variabilis)、辽东栎(Quercus liaotungensis)、山杨(Populus davidiana)等为主。由于人类活动的影响, 林地主要分布在海拔较高的山地, 海拔较低的丘陵区主要为灌草丛, 种类较贫乏, 主要为荆条(Vitex negundo)、酸枣(Ziziphus jujuba)、白羊草(Bothriochloa ischaemum)、黄背草(Themeda triandra)等。在村庄、路旁散生有杨(Populus)、榆(Ulmus)、柳(Salix)、槐(Sophora japonica)、泡桐(Paulownia fortunei)等乔木。农作物类型主要有玉米(Zea mays)、谷子(Setaria italica)、高粱(Sorghum vulgare)、小麦(Triticum aestivum)、荞麦(Fagopyrum esculentum)、莜麦(Avena nuda)、大豆(Glycine max)、马铃薯(Solanum tuberosum)、亚麻(Linum usitatissimum)、向日葵(Helianthus annuus)、油菜(Brassica campestris)、白菜(Brassica pekinensis)等。人工种植的经济林主要有苹果(Malus domestica)、桃(Amygdalus persica)、梨(Pyrus)、板栗(Castanea mollissima)、核桃(Juglans regia)、葡萄(Vitis vinifera)等。
2008年7月中旬, 在研究区内共采集表土样品54个, 其中农田样品32个(编号为1~32), 荒地样品22个(农田样品采自农田内, 可定义为人类活动影响强度较大的样品; 荒地样品多采自田间荒地或荒坡上, 可定义为人类活动影响强度相对较弱或人工扰动植被样品)。样点位置见图1(33~54号荒地样品由于取自农田样品附近, 地图上两类样品位置基本重合, 因此图1中显示的是全部农田样点位置), 样点周围植被组成见表1。样品取自0~1 cm的表层土壤中, 并记录每个样点的经纬度、海拔高度, 同时调查样点周围的植被状况。荒地植被年际间变化相对较小, 但农田作物种植类型可能经常发生变化, 因此, 表土花粉组合只能是多年植物类型花粉的混合。采样点的气象数据(年均降水量、年均温)依据国家基准站气象资料插值获得, 并对温度变量进行海拔校正。
 | 图1 晋冀低山丘陵区表土孢粉采样点位置Fig.1 Distribution of sample sites of surface pollen in hilly areas of Shanxi and Hebei Provinces |
| 表1 晋冀低山丘陵区表土孢粉采样点位置及植被状况 Table 1 Location and vegetation conditions of sample sites of surface pollen in hilly areas of Shanxi and Hebei Provinces |
孢粉提取采用常规的酸碱处理和重液浮选方法(Faegri and Iversen, 1989)。每个样品取干重20 g, 加入1粒石松孢子片(27 637± 563粒/片)作为指示剂, 以计算孢粉浓度。花粉鉴定统计在400倍日本Olympus BX51光学显微镜下进行, 每个样品观察统计至少3张薄片, 鉴定统计花粉400粒以上。研究区农作物以玉米、谷子、高粱、小麦、莜麦等禾本科谷物为主。但禾本科花粉形态非常相近, 如何区分谷物禾本科与杂草禾本科花粉始终是孢粉形态学研究的难题。依据作者制作的部分禾本科花粉(小麦、玉米、莜麦、高粱、谷子、白羊草、黄背草、狗尾巴草等)标准片, 参照前人研究成果(Andersen, 1978; 蓝盛银和徐珍秀, 1996; 王学丽等, 2010), 将本研究中的禾本科花粉分为2类:个体直径大于35× 35 μ m、孔径较大、颗粒纹饰较清晰的归为谷物禾本科花粉, 如小麦直径平均63.3× 56.6 μ m、玉米89.9× 80.8 μ m、莜麦41.5× 38.5, μ m、高粱49.0× 44.3 μ m, 谷子花粉个体直径虽小于35× 35 μ m, 但孔径较大, 平均6.2 μ m; 个体直径小于35× 35 μ m、孔径多在4.5 μ m以下、雕纹较模糊的(较谷物禾本科浅)归为杂草禾本科花粉, 如白羊草个体直径29× 26.5 μ m、孔径3 μ m, 黄背草个体直径26.5× 23.5 μ m、孔径3, μ m。
样点位置图用ArcGIS软件绘制, 孢粉图谱由Tilia软件完成。
主成分分析(Principal component analysis, PCA)作为一种多元分析方法, 在表土孢粉研究中可以突出影响孢粉组合的主要孢粉类型, 使研究结果更直观(王琫瑜等, 1996; 李月丛等, 2008)。本研究用PCA方法结合孢粉组合来划分农田和荒地。在PCA分析的基础上, 对环境因子进行冗余分析(Redundancy analysis, RDA), 探讨孢粉组合与气候的关系。PCA和RDA分析采用Canoco4.5软件(王琫瑜等, 1996)完成。
多响应置换过程分析(Multi-response permutation procedures, MRPP)用来检验预先设定的不同组间的差异性(Zimmerman et al., 1985; Stallins, 2002)。本研究采用MRPP方法分析不同强度人类活动影响下(农田、荒地)的孢粉组合是否存在显著差异, 若差异性显著(差异值P< 0.05, P越接近于零, 差异性越显著), 再进行指示种分析(Dufrê ne and Legendre, 1997; 吕浩荣等, 2009)。指示种分析(Indicator species analysis, ISA)用来探究对人类活动干扰强度具明显指示意义的孢粉类型, 并用蒙特卡罗检验(Monte Carlo permutation test)进行显著性检验(Dufrê ne and Legendre, 1997; 吕浩荣等, 2009)。MRPP分析和ISA分析在PC-ORD4.0软件(McCune and Mefford, 1999)下完成。
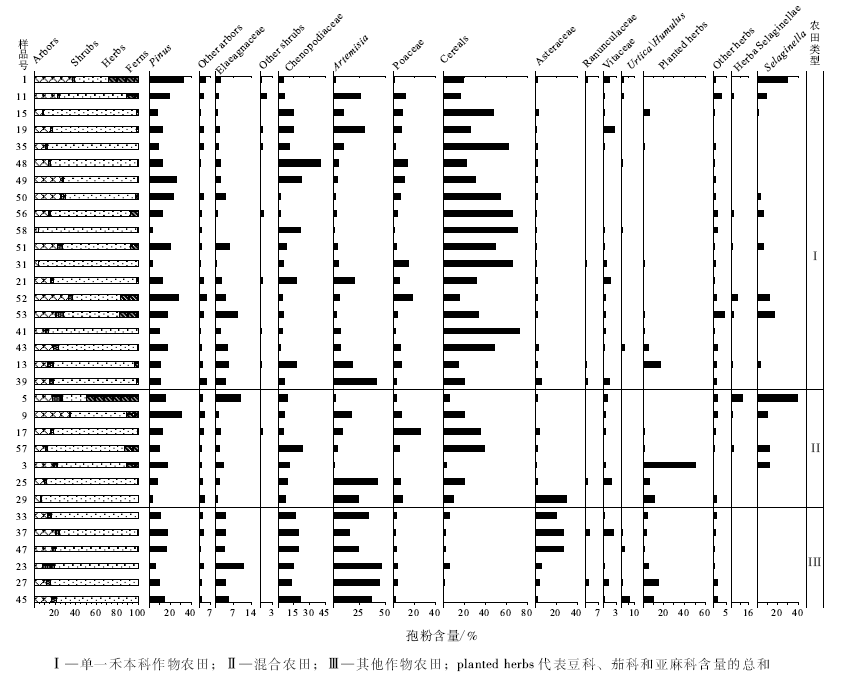
54个样品共鉴定出100个科属植物的孢粉类型。其中乔木植物花粉类型23个(平均17.1%, 文中百分含量均为平均值), 普遍出现且含量较高的类型有松属(Pinus)和栎属(Quercus), 含量较低的是桦属(Betula)、胡桃科(Juglandaceae)、臭椿属(Ailanthus)、杨属(Populus)和柳属(Salix)等; 灌木植物花粉类型15个(3.4%), 主要有胡颓子科(Elaeagnaceae)、鼠李科(Rhamnaceae)、虎榛子属(Ostryopsis)、白刺属(Nitraria)、麻黄属(Ephedra)、牡荆属(Vitex)和木犀科(Oleaceae)等; 草本植物花粉类型57个(74.5%), 常见的有谷物禾本科(Cereals)、杂草禾本科(Poaceae)、蒿属(Artemisia)、藜科(Chenopodiaceae)、蓼科(Polygonaceae)、菊科(Asteraceae)、十字花科(Brassicaceae)、葎草属/荨麻属(Humulus/Urtica)、毛茛科(Ranunculaceae)、唇形科(Lamiaceae)、唐松草属(Thalictrum)、茄科(Solanaceae)、旋花科(Convolvulaceae)和蔷薇科(Rosaceae)等; 蕨类孢子(5.0%)主要为中华卷柏孢子(Selaginella sinensis)和三缝孢(Trilete spore)等。
虽然采集了54个地点的样品, 但有的地点只采集了农田样品, 没有采集荒地样品。为了分析农田样品(指示人类活动强度较强)与荒地样品(指示人类活动强度相对较弱)花粉组合的差异, 选取了研究区22组(1~22号和33~54号)既采集了农田样品、又采集了荒地样品样点的花粉组合进行对比分析, 以探究在不同强度人类活动影响下的花粉组合的差异。
3.2.1 农田、荒地孢粉含量的差异
农田与荒地样品均以草本植物花粉为主, 但农田中以谷物禾本科花粉占优势, 而蒿属花粉则是荒地的主要成分。农田样品中乔木植物花粉占15.6%, 主要包括松属(13.3%)、桦属和栎属等; 灌木植物花粉含量平均为3.4%, 以胡颓子科(3.2%)为主; 草本植物花粉含量平均为75.5%, 以谷物禾本科(24.1%)、藜科(10.5%)、蒿属(20.2%)、杂草禾本科(5.9%)和菊科(5.6%)为主, 毛茛科、荨麻属/葎草属和十字花科等为常见草本植物花粉类型; 蕨类孢子主要为中华卷柏(5.5%)。荒地样品藜科(17.6%)、蒿属(36.2%)和荨麻属/葎草属(2.2%)等草本植物花粉含量高于农田, 谷物禾本科含量(3.8%)低于农田, 而松属(15.1%)、胡颓子科(3.1%)、杂草禾本科(6.0%)和中华卷柏孢子(3.7%)含量与农田相近。此外, 一些农作物花粉只出现于对应的农田及其周围样品中, 如亚麻科(Linaceae)和向日葵属(Helianthus), 个别花粉类型(如松属)异常值系样点周围有母本植物生长所致(图 2)。
 | 图2 晋冀低山丘陵区农田与荒地表土样品主要花粉类型含量Fig.2 Content of major pollen types in surface pollen samples of farmlands and wastelands in hilly areas of Shanxi and Hebei Provinces |
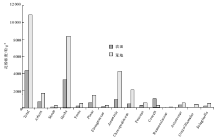
3.2.2 农田、荒地孢粉浓度的差异
不同地点土壤、水分、植物类型、地貌部位等条件的差异, 导致其表土样品的孢粉浓度可能没有可比性, 但众多农田与荒地样品的差异可能反映其孢粉浓度的特征。晋冀低山丘陵区农田孢粉浓度平均约4380 粒/g, 荒地孢粉浓度平均约10983粒/g。农田中除谷物禾本科浓度明显高于荒地外, 其他类型的花粉浓度明显低于荒地, 特别是松属等乔木植物花粉以及蒿属、藜科等草本植物花粉, 但蕨类孢子浓度两者相近(图 3)。
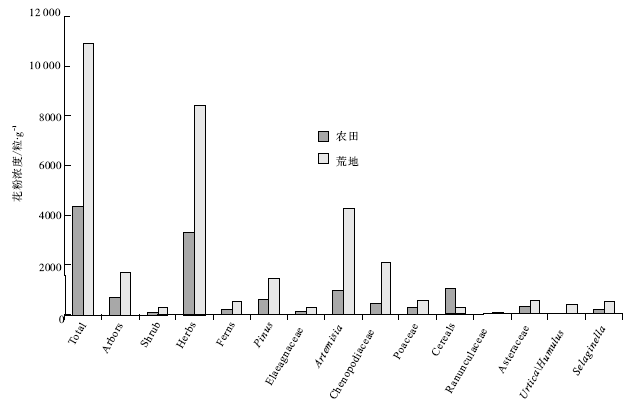 | 图3 晋冀低山丘陵区农田与荒地表土样品主要花粉类型浓度对比Fig.3 Concentration of major pollen types in surface pollen samples of farmlands and wastelands in hilly areas of Shanxi and Hebei Provinces |
农田与荒地孢粉组合特征表明, 农田与荒地样品中占优势的孢粉类型不同, 故是否可以依据某些孢粉类型(花粉类型至少出现在3个样点以上, 且在样点中的平均含量大于1%)来划分农田与荒地是一个值得探讨的问题。因此笔者对54个样品进行了主成分分析。结果显示:第1、第2主轴的特征值分别为0.50和0.20, 第3、第4主轴的特征值为0.14和0.05, 说明孢粉组合主要受第1、第2主轴控制。主要孢粉类型的排序结果(图 4-a)显示, 谷物禾本科、松属、蒿属等在排序空间内的变化比值相对较大, 第1轴上谷物禾本科的得分值最高, 蒿属最低; 第2轴上松属花粉的得分值最高, 谷物禾本科花粉最低。由于谷物禾本科花粉与人类活动关系密切, 因此认为, 第1轴正方向和第2轴负方向可能指示人类活动较强的方向, 而第1轴负方向和第2轴正方向可能指示人类活动较弱的方向。图4-b是所有样品的排序结果, 根据样品的分布位置, 将样品划分为2组:第1组共18个样品, 均为农田样品, 主要分布于第1轴正方向。第2组共30个样品, 除9个农田样品外, 其余为荒地样品, 且主要分布于第1轴负方向和第2轴正方向。第2组中含9个农田样品, 这些农田主要种植非禾本科作物(葡萄、亚麻、豆、向日葵等), 谷物禾本科含量较低(平均6.5%), 花粉组合以蒿属、藜科和松属等为主, 更接近荒地花粉组合特征。PCA分析表明, 禾本科作物农田与荒地样品可以根据主要花粉类型进行划分, 但主要种植非禾本科作物的农田与荒地则较难区分。
 | 图4 晋冀低山丘陵区表土样品主要孢粉类型(a)及采样点(b)孢粉组合PCA排序示意图Fig.4 Ordination results of principal component analysis(PCA)for major pollen types(a) and sample sites(b) of surface pollen samples in hilly areas of Shanxi and Hebei Provinces |
为了解晋冀低山丘陵区人工扰动植被孢粉组合能否反应区域气候特征, 文中在PCA分析的基础上将经度、纬度、海拔、年均降水量、年均温作为气候参数(5个气候因子之间方差膨胀因子值小于20)(Ter Braak and Prentice, 1988; Lu et al., 2006), 与样品孢粉组合进行RDA分析。结果显示, 孢粉组合受多种气候因素的共同影响, 且第1、第2主轴贡献率较高, 两轴与5个环境因子的相关性分别为0.73和0.56, 且第1轴与年均温(0.63)、年均降水量(0.55)呈正相关, 与海拔(-0.62)呈负相关(海拔变化本质反映的是温度的变化, 海拔升高温度降低, 反之海拔降低温度升高); 第2轴与经度(-0.18)呈负相关。RDA分析表明, 样品与气候因子的相关性较好, 且研究区内孢粉组合特征较好地反映了区域温度和降水量的变化。当花粉组合中谷物禾本科、松属和中华卷柏占优势时, 反映区域的气温、降水条件较好(或者海拔高度较低), 当花粉组合中以蒿属和菊科等占优势时, 反映区域气温、降水条件较差(或者海拔较高)。
多响应置换过程分析显示, 农田与荒地样品的差异值P为0.000 003 02, 小于0.05, 表明农田与荒地两者之间存在显著差异。指示种分析显示, 7种花粉类型通过蒙特卡罗检验(P< 0.05)(表 2), 其中谷物禾本科为农田常见花粉类型, 含量明显高于荒地, 为农田指示种, 葡萄科(Vitaceae)、胡桃科在农田中的含量虽较低(介于0.5%~1%之间), 但仍高于荒地中的含量, 也可以作为农田孢粉指示种; 蒿属在荒地中的含量明显高于农田(2倍以上), 为荒地指示种, 蓼科、蔷薇科(Salicaceae)在农田和荒地中含量均较低, 但在荒地中的含量明显高于农田中, 为荒地指示种(表 2)。
| 表2 晋冀低山丘陵地区人工扰动植被主要花粉指示种 Table 2 Pollen indictor species of human disturbed vegetation in hilly areas of Shanxi and Hebei Provinces |
依据农田中作物类型, 将农田分为单一禾本科作物农田(19个, 只种植玉米、莜麦、谷子)、混合农田(7个, 多种农作物混合种植, 主要有玉米、高粱、谷子、黍子、大豆、土豆、亚麻等)和其他作物农田(6个, 主要为向日葵、豆科、胡麻科、茄科等)。结果显示, 不同作物类型农田中谷物禾本科花粉含量差异明显, 其他作物农田谷物禾本科含量(2.4%)明显低于混合农田(18.8%)和单一禾本科作物农田(40.2%)(图 5)。但其他作物农田种植的农作物花粉含量较高, 如种植向日葵的农田中菊科花粉含量平均为24.7%(非向日葵农田中菊科含量仅3.2%), 种植胡麻的农田中胡麻科花粉含量平均为6.3%(非胡麻农田中基本不含胡麻科花粉)。
 | 图5 晋冀低山丘陵区不同农田类型表土样品花粉组合特征Fig.5 Assemblages characteristics of major pollen types in surface pollen samples of different farmlands in hilly areas of Shanxi and Hebei Provinces |
孢粉研究人员一直在努力寻找反映人类活动强度的孢粉指示种(Court-Picon et al., 2005; Lamentowicz et al., 2008; 李宜垠等, 2008), 并将生态学的多响应置换分析和指示种分析方法应用在不同群落类型孢粉指示种分析研究上(Herzschuh and Birks, 2010)。多响应置换分析表明, 晋冀低山丘陵区不同强度人类活动下的农田和荒地植物群落孢粉组合存在显著差异, 表明孢粉可以反映不同的人类活动强度和土地利用特征。指示种分析结果显示, 人类活动影响强度不同, 指示种不同。谷物禾本科花粉指示值最高(85.5, 指示值越高, 说明指示性越好, 最高为100), 与研究区主要种植玉米、小麦、谷子、莜麦、高粱等禾本科作物密切相关, 因此, 谷物禾本科对农田指示性最好, 与前人研究结果(Court-Picon et al., 2005; Cao et al., 2010)一致。葡萄是研究区农田中常见的经济作物, 花粉含量虽较低(马艳玲等, 2009; 庞瑞洺等, 2010; 丁伟等, 2011), 却对农田有较好的指示意义(指示值为66.5); 胡桃科中的核桃是研究区村落或附近农田、路旁人工培育栽种的经济林树种, 有的地方农田中栽种的就是核桃树, 其花粉含量虽较低, 但指示值为55.8, 对农田仍有较好的指示意义。荒地由于水分条件差、土壤较贫瘠等原因, 蒿属(指示值为69.9)、蓼科(指示值为36.2)、蔷薇科(指示值为35.0)植物生长较好, 是研究区荒地较好的指示种; 杨柳科为研究区常见的林道树或沿河树种, 广泛分布在公路旁或河边, 虽然林道树为人工种植, 但多远离农田, 常与荒地相伴, 因此对荒地有一定指示意义, 不过其指示值较低(27.2), 且花粉含量(0.05%和0.31%)也较低, 故其对研究区荒地的指示意义有待更多数据证实。
农田中不同作物类型谷物禾本科花粉含量差异明显, 混合农田谷物禾本科花粉含量明显低于单一禾本科作物农田, 应与农作物类型有关, 混合农田为谷物禾本科和其他农作物类型混合种植区, 非谷物禾本科农作物花粉含量的增加, 必然导致谷物禾本科花粉含量的下降(马艳玲等, 2009; 黄小忠等, 2011)。不同地区禾本科作物农田的谷物禾本科花粉含量也存在差异, 可能与区域湿润指数、植被类型、种植方式等不同有关, 如东北和华南地区(杨士雄等, 2010; 李曼玥等, 2012)谷物禾本科花粉含量均高于40%, 可能与东北和华南地区气候相对湿润、地表植被发育、花粉落到地表后不易被氧化有关; 西北(12.2%~17.3%; 马艳玲等, 2009)和华北地区(19.4%~30.6%; 庞瑞洺等, 2010和本文)相对较低, 可能与西北和华北地区气候相对干燥、地表植被欠发育、花粉落到地表后容易被氧化有关。但不论是哪个地区, 离开农田后谷物禾本科花粉含量均迅速下降(大部分低于6.0%), 因此可以根据谷物禾本科花粉含量直接划分农田和荒地, PCA分析中, 谷物禾本科的农田样点较易与荒地分开也证明了这一点。但若农田中主要种植非禾本科作物, 则较难与周边荒地分开, 这是由于该农田中禾本科花粉含量较低, 与荒地接近; 若农田中主要种植十字花科、豆科、茄科等作物, 因十字花科常在花期前收割(邓韫等, 2002; 羊向东等, 2005), 豆科、茄科作物的花粉产量较低(王琫瑜等, 1996; 张丽艳等, 2007; 丁伟等, 2011), 没有出现明显区别于荒地的特征, 因此在PCA分析中较难与荒地分开。
农田与荒地样品孢粉浓度对比显示:农田样品孢粉浓度除谷物禾本科花粉明显高于荒地外, 其他植物类型均低于荒地。农田中谷物禾本科花粉浓度较高, 与农作物种植面积较大和种植强度高有关。其他植物类型的孢粉浓度低于荒地, 与孢粉保存环境有关, 因为为了提高农作物产量, 农田中常施用大量化肥、灌溉、翻耕、除草、混种、轮作等手段促进农作物增产, 这都可以改变土壤的酸碱度, 使土壤孔隙增大、氧化作用增强。已有研究表明, 土壤pH值变化、孔隙增大、氧化作用增强都会使花粉保存能力下降(许清海等, 2005a, 2005b), 进而降低农田中表土孢粉的浓度。此外, 花粉来源和产量差异也会引起浓度的差异, 农田花粉来源主要为农田农作物, 荒地花粉来源主要为荒地中的杂草, 有研究认为, 谷物禾本科花粉产量相对低于荒地中的蒿属、藜科植物的花粉产量(Moore, 1978; Birks and Birks, 1980; Bunting et al., 2005; 李月丛等, 2005a; Soepboer et al., 2007), 进而导致农田孢粉浓度较低。
为了更好地探讨农田和荒地孢粉浓度的差异, 将本研究结果与课题组对东北地区、华北南部及华中北部、西北和河北中南部地区的研究结果进行对比(表3)。对比结果(表3)表明, 尽管不同地区人类活动影响强度、作物类型、气候条件不同, 但农田孢粉浓度均低于荒地。另外, 不同地区孢粉浓度也存在一定差异, 农田样品孢粉浓度从高到低依次为:西北地区、华北南部及华中北部、晋冀低山丘陵区、河北中部、东北地区。荒地样品孢粉浓度也是西北地区、华北南部及华中北部高, 不同的是东北地区荒地孢粉浓度高于河北西部及山西北部地区。但总体均表现出丘陵区孢粉浓度大于平原区孢粉浓度的特征。孢粉浓度的差异似乎与气候无关, 而与地貌类型、海拔高度和人类活动有关。相对平原区而言, 低山丘陵区海拔较高、地形复杂、人类活动较弱, 农田多依地形分布、面积相对较小、距荒山杂草植被较近, 接受了较多荒山杂草的孢粉, 因而低山丘陵区孢粉浓度高于平原区。东北荒地孢粉浓度高于河北西部及山西北部地区是否与东北地区气候较湿润有利于花粉保存有关, 有待进一步研究。
| 表3 中国不同区域农田和荒地孢粉浓度(粒/g) Table 3 Pollen concentration(grains/g)in farmland and wasteland form different area in China |
1)晋冀低山丘陵区农田孢粉浓度平均约4380粒/g, 荒地孢粉浓度平均约10983粒/g。农田中除谷物禾本科花粉浓度高于荒地外, 其他植物类型孢粉浓度均低于荒地。
2)多响应置换分析表明, 晋冀低山丘陵区不同强度人类活动影响下的花粉组合存在明显差异, 孢粉组合可以反映人类活动影响强度和土地利用状况。指示种分析表明, 晋冀低山丘陵区谷物禾本科、葡萄科、胡桃科指示人类活动较强的农田, 蒿属、蓼科、蔷薇科指示人类活动较弱的荒地。
3)PCA分析结果显示, 可以依据花粉组合特征区分禾本科作物农田与荒地, 而主要种植十字花科、豆科、茄科等作物的农田则较难与荒地区分。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|