







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
华北地台西缘早—中寒武世过渡期核形石:微生物群落对浅海缺氧环境的响应
[张文浩1  , 史晓颖
, 史晓颖1, 2 , 汤冬杰2 , 王新强1, 2 ]
, 史晓颖, 汤冬杰|
|

通讯作者简介 史晓颖,男,1956年生,博士生导师,主要从事地层古生物及沉积学方面的教学与研究。E-mail:shixycugb@cugb.edu.cn。
第一作者简介 张文浩,男,1987年生,博士研究生,主要从事地球生物学与沉积学方面的研究。E-mail:wenhaocugb@aliyun.com。电话:13426430299。
地质记录表明在早—中寒武世过渡期发生了一次重要的生物灭绝事件,但对其成因的认识仍存在分歧。作者试图通过对华北地台该时段广泛发育的核形石微组构和矿化过程研究,揭示微生物群落对灾变事件和海洋环境变化的响应。研究发现核形石内富含细菌化石;包壳的微组构和有机矿化特征表明其形成于细菌硫酸盐还原( BSR)作用活跃的高碱度海水条件,密集的莓状黄铁矿微粒和异养细菌残余指示为缺氧环境;而沉积相分析显示核形石发育于浅海陆棚背景。研究认为,同期的核形石及其他可对比微生物岩不仅在华北广泛分布,在其他大陆和板块上也有良好记录,表明在这个生物灾变期浅海环境有广泛的微生物群爆发,并可能与大火成岩省喷发、全球气温升高以及深部缺氧海水向陆棚侵进引起的浅海广泛缺氧相关。早—中寒武世过渡期广泛发育的微生物岩可能记录了底栖动物大量灭绝后,微生物群在缺氧水体中的快速繁盛与生态扩张过程。
About the corresponding author Shi Xiaoying,born in 1956,is a professor in China University of Geosciences(Beijing). E-mail: shixycugb@cugb.edu.cn.
About the first author Zhang Wenhao,born in 1987,is a Ph.D. candidate of paleontology and stratigraphy. He is mainly engaged in biogeology and sedimentology. E-mail:wenhaocugb@aliyun.com.
Geological records showed that an important biological extinction event happened during the Early-Middle Cambrian transition period, but there are controversial views about its orign. In order to reveal the response of microbial community to the biotic crisis and the marine environment change,microfabrics and organo-minerals in the oncoid cortices formed in that period at the North China Platform were studied. The study revealed that abundant microbial fossils exist in the oncoids. Study of microfabrics and organomineralization patterns in the oncoid cortices suggested that the oncoids were likely formed in seawater with high alkalinity and active bacterial sulfate reduction(BSR). Rich pyrite framboids and purported heterotrophic remains in the cortices represented an anoxic bottom-water condition,and facies and depositional sequence analyses suggested a shallow shelf environment. A wide correlation indicated that the time-equivalent oncoids or related microbialites were widespread in the North China Platform,and also well recorded in some other continents or plates. This may imply an extensive bloom of microbial communities in the aftermath of terminal Early Cambrian biotic crisis,which was likely related to a pervasive anoxia in shallow marine environments caused by LIP(Large igneous province) eruptions,and subsequently global warming and invasion of anoxic deep seawater onto the shelves at that time. The broad distribution of oncoids and other related microbialites at the Early-Middle Cambrian transition may have reflected an ecological response and rapid expansion of microbial community to the high-stressed environments where benthic metazoans had been largely suppressed during the biotic crises.
微生物岩作为微生物群落与环境相互作用的产物(Riding, 2000; Shi et al., 2008), 其地质分布和丰度变化不仅反映了微生物群演化及其与后生动物的相互作用(Riding and Liang, 2005), 而且也反映了对环境变化的响应过程(Fischer, 1965; Dupraz et al., 2009; Harwood and Sumner, 2011)。大量的研究表明, 古代微生物岩变化及其微组构特征能够用于古海洋环境(如, 水深、温度、碳酸盐饱和度与缺氧条件)的分析(Riding, 2001; Woods and Baud, 2008; Kershaw et al., 2012; Mata and Bottjer, 2012; Tang et al., 2013)。尤其在显生宙, 微生物岩的广泛发育往往与生物灾变事件和高应力环境相伴生(Whalen et al., 2002; Wang et al., 2005; Kershaw et al., 2007, 2012), 并常被解释为由于底栖动物大量减少、高适应性微生物在高应力环境条件下迅速繁衍和生态扩张的结果(Fischer and Arthur, 1977; Schubert and Bottjer, 1992; Bottjer et al., 1995; Wood, 2000; 王永标等, 2005; Kershaw et al., 2012)。
核形石(oncoids)是指直径大于2 mm、内部具不完整或不规则同心层的碳酸盐球状体, 通常由核心(nucleus)和包壳(cortex)两部分组成。它的形成与微生物活动密切相关(Jones and Renaut, 1997; Hä gele et al., 2006; Shapiro et al., 2009), 并曾被称为“ 球状叠层石” (Logan et al., 1964)或“ 藻球” (Riding and Voronova, 1982)。由核形石堆积形成的核形石灰岩(oncolite)常被视为微生物岩的一种类型(e.g. Peryt, 1983; Tucker and Wright, 1990; Riding, 1991)。核形石在古元古代至新生代地层中均有记录(Li et al., 2000; Schaefer et al., 2001; Shi and Chen, 2006; Reolid and Nieto, 2010); 在现代盐湖(Jones and Renaut, 1994)、热泉(Renaut et al., 1996)、河流(Hä gele et al., 2006)、洞穴(Jones and Renaut, 2010)、深海底(Wang et al., 2012b)等底栖动物贫乏或寡营养环境中也有发现, 并往往具有形态、微组构和矿物组成等特征上的多变性。
地质记录表明, 在早— 中寒武世之交发生了显生宙第一次重大的生物集群灭绝事件(Zhuravlevand and Wood, 1996; Palmer, 1998; Erwin, 2001; Hallam, 2005), 并伴生有显著的δ 13C(Zhu et al., 2006)、δ 33S同位素(Hough et al., 2006)偏移和海水 87Sr/86Sr 值(Montañ ez et al., 2000)变化。虽然以往对灾变期标志性的三叶虫和古杯类生物群变化有较好的研究, 但对微生物群的变化了解甚少, 且对该生物灭绝原因的认识存在分歧。作者主要报道内蒙古乌海地区毛庄组上部至徐庄组底部大量发育的核形石, 重点讨论它们与生物灭绝及相关事件的关系, 并通过核形石微组构和矿化机制的研究揭示微生物对危机期海洋环境变化的响应, 从而进一步认识微生物与环境的相互作用过程。
研究的核形石主要来自内蒙乌海地区(图 1-A)毛庄组上部至徐庄组下部(图 1-B-①), 少量来自宁夏青龙山地区的毛庄组上部(图 1-B-②)。研究区位于华北地台西缘, 属中朝板块西部大陆边缘的一部分(王鸿祯等, 2000)。与华北地台多数地区一样, 本区的寒武系是在新元古代“ 雪球地球” 结束之后, 随着Rodinia超大陆全面解体和全球海平面上升, 于早寒武世晚期开始接受沉积(史晓颖等, 1997; Meng et al., 1997; Shi et al., 1999; 王鸿祯等, 2000)。在华北地台, 寒武系底部普遍存在重大的区域不整合, 缺失震旦系至下寒武统下部。寒武系总体上以陆棚浅海碳酸盐岩为主, 下部含较多陆源碎屑, 横向岩相分异不太强烈(项礼文等, 1999; 王鸿祯等, 2000); 已建立了良好的生物地层(项礼文等, 1981, 1999; 安太庠, 1987; 彭善池, 2009)和层序地层序列(史晓颖等, 1997; Meng et al., 1997; Shi et al., 1999; 王鸿祯等, 2000)。
 | 图1 华北地台早— 中寒武世过渡期沉积古地理及含核形石地层对比Fig.1 Oncolite distribution and related stratigraphic succession at the Early-Middle Cambrian transition in North China Platform |
在研究的剖面上, 毛庄组与下伏中元古界蓟县系厚层含硅质条带白云岩呈不整合接触。早— 中寒武世以缓坡沉积为主, 从毛庄组到徐庄组陆源碎屑逐渐减少, 而灰岩层明显增多; 结合区域地层发育及沉积相特征分析, 这可能反映了随海侵逐步扩大, 沉积环境由近岸向浅水陆棚的转换。
在乌海剖面上(图 1-B-①), 核形石灰岩极为发育, 呈中— 厚层状产于毛庄组上部和徐庄组底部, 含核形石地层序列厚达12 m; 分别出现在三级层序的高位体系域(HST)上部和海进体系域(TST)下部。据前人的生物地层资料(安太庠, 1987; 内蒙古自治区地质矿产局, 1991)、层序地层对比(史晓颖等, 1997; Meng et al., 1997; Shi et al., 1999)和三叶虫化石带(项礼文等, 1999; 王鸿祯等, 2000; 彭善池, 2009)约束, 核形石的产出层位相当于华北习用的底栖三叶虫Yaojiayuella带上部至Hsuzhauangia带下部(项礼文等, 1999; 王鸿祯等, 2000)。根据与华南深水相区浮游三叶虫化石带的对比关系(彭善池, 2009), 本区核形石的主要产出层位大致相当于华南浮游三叶虫Ovatoryctocara granulata带(都匀阶顶部)至Oryctocephalus indicus带(台江阶下部), 属早— 中寒武世过渡期, 并可能主要为第五阶下部, 推测年龄在510~507 Ma之间(图 2)(Haq and Schutter, 2008; Walker and Geissman, 2009; Cohen et al., 2012)。
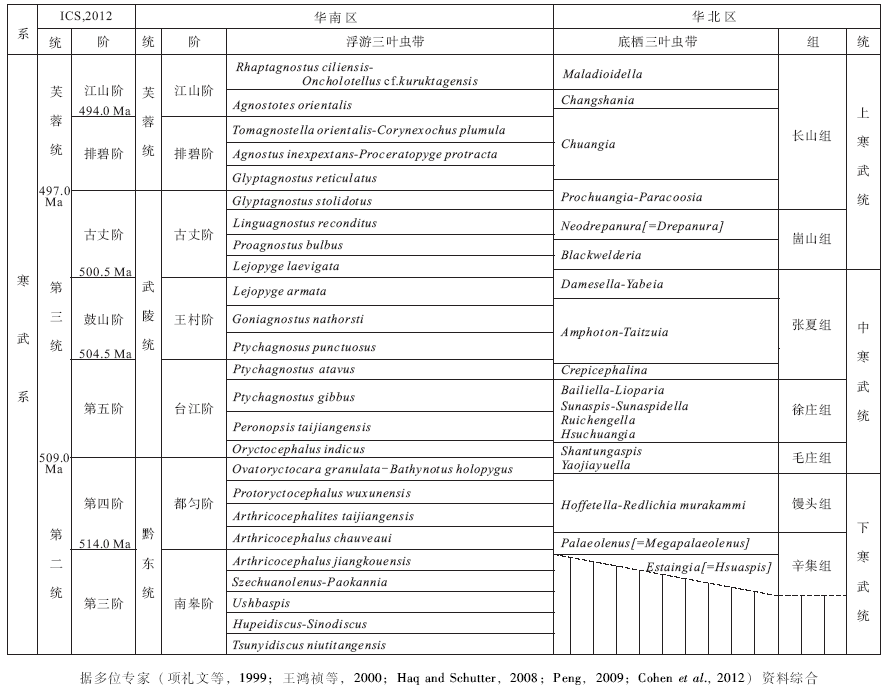 | 图2 华北寒武系地层划分及其国际年代地层对比关系Fig.2 The Cambrian stratigraphic subdivisions of North China and their chronostratigraphic correlation |
内蒙古乌海剖面毛庄组含2个沉积层序(图 1-B-①)。下部层序厚度较小, 底部灰褐色粉砂岩代表海进初期沉积; 中部由灰黄色泥页岩和薄层灰岩交互的米级副层序构成(图 3-A), 构成TST的主体; 灰岩中局部含鲕粒和少量生物碎屑, 表明主要形成于中缓坡环境。HST由中— 厚层细粒含长石石英砂岩组成, 见有小型交错层理, 可能形成于能量较高的潮下带上部或浅缓坡环境。毛庄组上部层序的TST下部以黄绿色页岩居多, 夹薄层泥晶灰岩(图 3-B), 含少量生物碎屑。向上灰岩层明显增多(图 3-C), 并夹有颗粒灰岩和瘤状灰岩层, 主要形成于深— 中缓坡环境; 中部的瘤状灰岩代表最大海泛面(MFS)附近的较深水沉积。该层序的HST以灰岩为主, 下部灰岩层中夹有较多鲕和颗粒灰岩(图 3-D), 而上部则以核形石灰岩为主(图 3-E, 3-F, 3-G), 间夹颗粒灰岩(图 3-F)和薄层微晶灰岩。可能主要形成于浪基面附近的潮下带环境, 空间上与鲕滩环境相邻; 层序顶部为明显的Ⅰ 型层序界面。徐庄组也含有2个三级层序, 其内米级副层序发育良好。总体以薄— 中层状泥晶灰岩为主, 夹多层生物扰动灰岩和竹叶状砾屑灰岩, 并与黄绿色页岩交互构成多个次级旋回。从总体上看, 徐庄组2个三级层序的沉积水深明显大于毛庄组, 主要发育于正常浪基面和风暴浪基面之间的内陆棚环境。该组中核形石主要出现在下部三级层序的近底部(图 1-B-①), 个体略小于毛庄组上部的核形石。
 | 图3 内蒙古乌海地区寒武系毛庄组主要沉积相特征Fig.3 Characteristics of major sedimentary facies of the Cambrian Maozhuang Formation in Wuhai area, Inner Mongolia |
虽然核形石灰岩与鲕灰岩在毛庄组上部地层序列中相邻, 但核形石与鲕粒很少在同层内共生出现。在核形石灰岩中, 基质以微晶— 泥晶胶结物占主导, 而鲕灰岩的基质中亮晶胶结物有较高比例, 这可能表明鲕灰岩形成的水动力条件较强。在核形石的核心(图 4-A)与围岩(图 4-B, 4-C, 4-D)中普遍存在生物碎屑, 但其包壳中完全缺乏碎屑颗粒, 含有大量保存良好的微生物丝状体。这可能表明核形石主要发育于鲕滩附近的低洼地带, 水动力条件要明显弱于鲕灰岩发育的环境。
 | 图4 内蒙古乌海地区寒武系核形石灰岩中的生物碎屑(薄片显微照片)Fig.4 Photomicrographs showing biodetritus in oncolite of the Cambrian in Wuhai area, Iner Mongolia |
总体上看, 本区核形石主要形成于正常浪基面附近相对低能的浅海环境(张文浩等, 2014)。就其在三级沉积层序中的产出层位分析, 与其他地区报道的不同时代核形石相近(Van Buchem et al., 1996; Whalen et al., 2000a, 2000b; Gradziń ski et al., 2004), 主要出现在沉积层序的HST上部至TST下部, 这也可能表明核形石的发育与海平面变化及水深条件有较密切的相关性(Riding, 2006)。
核形石的微观和超微结构特征作者已有详细描述(张文浩等, 2014), 其显著的特征之一是构成核形石包壳的亮、暗交互纹层在矿物组成与微生物类型上有明显差异。亮纹层以微亮晶质为主, 含大量个体较大的光合自养蓝细菌(如Girvanella和Renalcis), 并具良好的钙化外鞘(图 5-A, 5-B)。而暗纹层则以微晶质为主, 其中含个体微小的异养细菌化石(图 5-C, 5-D), 多保存为残余, 莓状黄铁矿微粒在暗纹层中很丰富(图 5-E, 5-F)。这可能表明, 亮纹层的形成主要由光合自养微生物的钙化作用所导致(Arp et al., 2001), 并与微生物代谢消耗CO2, 增加碱度, 从而使微环境中碳酸盐饱和度升高, 造成微生物的矿化与碳酸盐沉淀密切相关(Arp et al., 2001; Dupraz et al., 2004, 2009)。而暗纹层可能与细菌硫酸盐还原作用(BSR)分解有机质产生HC
 | 图5 内蒙古乌海地区寒武系核形石包壳的微组构特征Fig.5 Microfabric features of oncoid cortex of the Cambrian in Wuhai area, Inner Mongolia |
核形石的地层分布在研究区限于毛庄组顶部至徐庄组底部, 其包壳主要由Girvanella等微生物及其有机矿化产物所形成。这种核形石在华北地台分布广泛, 产出层位一致(图 1-B)。在山东(史晓颖等, 1999)、山西(高建平和朱士兴, 1998)、北京周边(高冉等, 2011)、河北和辽宁(史晓颖等, 1997; 王鸿祯等, 2000)以及陕西陇县、宁夏苏峪口和青龙山地区(本文)均有发现(图 1)。此外, 在北美和澳大利亚(Youngs, 1978; Sundberg, 2005; Powell et al., 2006)、约旦(Elick et al., 2002)、西班牙(Á lvaro et al., 2000; Perejó n et al., 2012)以及阿根廷(Gomez et al., 2007)等地的早— 中寒武世之交也有报道, 且大都发育于浅海陆棚的沉积环境中。据华北地台寒武系层序地层序列与三叶虫带的产出层位(史晓颖等, 1997; 王鸿祯等, 2000)以及与华南深水相区浮游三叶虫带的对比关系(彭善池, 2009), 可以看出华北地台核形石广泛发育的层位, 与已知的早— 中寒武世之交生物灭绝事件(Zhuravlev and Wood, 1996; Palmer, 1998; Erwin, 2001; Hallam, 2005), 在时间上有很好的一致性(图 2)(Zhuravlev and Wood, 1996; Glass and Phillips, 2006; Mata and Bottjer, 2012)。
在宁夏青龙山毛庄组上部(图 1-B-②), 除发现有大量核形石外(图 6-A), 在共生的暗色页岩夹层中发育保存良好的皱饰构造(属微生物成因构造MISS的一种)(图 6-B)和丰富的莓状黄铁矿微粒。这种MISS构造主要见于前寒武纪(Noffke et al., 2008; Shi et al., 2008), 其发育与保存要求缺氧环境条件(Zhuravlev and Wood, 1996; Mata and Bottjer, 2012)。而莓球状黄铁矿普遍被认为标志着缺氧条件(Soliman and Goresy, 2012; Wang et al., 2012a)。虽然目前发现的这些证据仍然有限, 但似乎表明在毛庄组上部核形石发育期, 浅海环境的确具有缺氧— 贫氧的海底条件。
 | 图6 寒武系毛庄组上部的核形石与共生的微生物席皱饰构造Fig.6 Oncoids and microbial wrinkles in upper part of the Cambrian Maozhuang Formation |
从整体上看, 微生物岩在前寒武系中相当普遍(Riding and Liang, 2005; Tang et al., 2013), 而在显生宙丰度明显下降(图 7)。它们在显生宙的广泛发育往往与缺氧环境密切相关(Riding, 2000; Flü gel and Kiessling, 2002; Woods and Baud, 2008; Mata and Bottjer, 2012)。特别是在灾变期底栖动物大量衰减或灭绝后, 由于缺乏后生动物的觅食消耗, 具有较强适应性的微生物群可在高应力环境中存活并在多种生态域扩展(Kershaw et al., 2012), 从而有更多机会堆积并保存成为微生物岩(图 7)。因而有研究认为, 微生物岩在显生宙一些时段的广泛发育可作为生物灾变事件的标志(Schubert and Bottjer, 1992; Whalen et al., 2002)。如在晚泥盆世F/F以及二叠纪末生物大灭绝后, 微生物岩不仅广泛发育, 其丰度也明显增加(图 7)(Whalen et al., 2002; Pruss et al., 2006; Woods and Baud, 2008; Xie et al., 2010; Ezaki et al., 2012; Kershaw et al., 2012)。表明在灾后浅海生态系统中微生物群落居主导地位(Yang et al., 2011; Mata and Bottjer, 2012)。类似的情况也出现在奥陶纪末生物大灭绝(Sheehan and Harris, 2004)以及晚志留世生物灭绝事件(Calner, 2005)之后。但在三叠纪末和白垩纪末2次大灭绝事件中至今尚未发现显著的微生物岩(图 7), 可能表明在这些时段限制后生动物存活的机制, 也同样制约了某些微生物的发展(Kiessling and Simpson, 2011)。
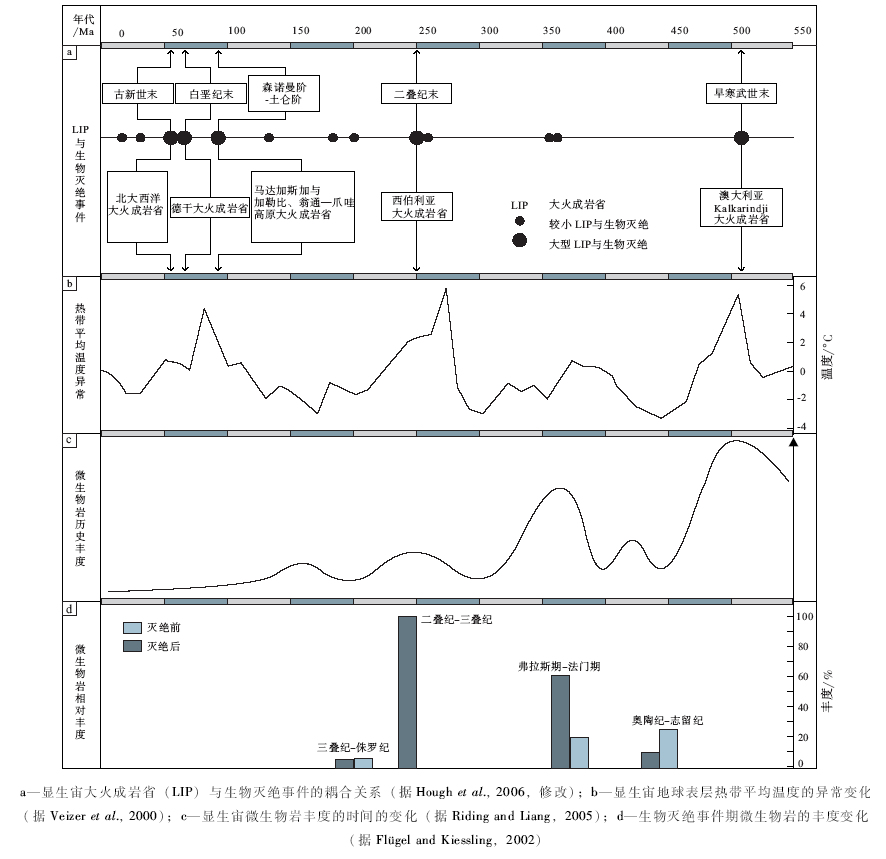 | 图7 显生宙历史中大火成岩省、生物灭绝、温度升高与微生物岩发育的时间对应联系Fig.7 Correlation of LIP, mass extinction, temperature in crisement, and microbialite development in the Phanerozoic |
近年的研究证明, 在澳大利亚发现的面积达2.1× 106km2的大火成岩省(即, Kalkarindji LIP)集中喷发于早— 中寒武世之交的512~507 Ma(Glass and Phillips, 2006; Evins et al., 2009)(图 7-a, 图8)。其规模仅次于著名的西伯利亚(P/T之交)、德干(K/T之交)以及中大西洋(T/J之交)大火成岩省(Wignall, 2001)。同时, 在早— 中寒武世之交约510 Ma附近识别了显生宙最大的硫同位素正向漂移(图 8), δ 34S值高达+50‰ (Hough et al., 2006), 并被解释为由于海洋极低的硫酸盐浓度与广泛缺氧条件相耦合的结果(Hough et al., , 2006)。与此相对应, 在北美、西伯利亚和澳大利亚都记录了明显的海洋碳同位素波动(Brasier et al., 1994; Dilliard et al., 2007; Howley and Jiang, 2010), 并被称为ROECE事件(Zhu et al., 2004)。这个事件在华南(Zhu et al., 2004)、塔里木盆地(Wang et al., 2011)以及华北地台南部(Zhu et al., 2004)也有良好的记录(图 8)。虽然造成地质历史上海洋广泛缺氧条件的原因有很多, 但高温气候条件导致海水溶氧量下降与快速海侵常被视为地质记录中最主要的因素(Erwin, 2001; Wignall, 2001; Hallam, 2005; Meyer and Kump, 2008)。由于Kalkarindji LIP的快速喷发, 大量CO2及其他挥发分被排放到大气圈, 导致气温迅速升高(Evins et al., 2009), 从而降低海水溶氧量。而随之发生的快速海进(Hough et al., 2006; Haq and Schutter, 2008)将深部缺氧海水带入浅水陆棚区, 造成浅海区底层水域广泛缺氧(Hough et al., 2006; Mata and Bottjer, 2012), 从而导致了浅海区底栖动物的灭绝(Zhuravlev and Wood, 1996; Wignall, 2001; Hallam, 2005)。而这种情形却为不强烈依赖氧浓度的微生物群的蓬勃发展创造了条件。在早寒武世末生物灭绝事件期间及其后, 在世界各地广泛出现了贫氧生物群(Zhuravlev and Wood, 1996)和富黄铁矿微纹层黑色页岩(Matta and Bottjer, 2012), 可能反映了这个时期浅海陆棚区广泛缺氧和富营养化的环境条件(Hough et al., 2006; Mata and Bottjer, 2012)。
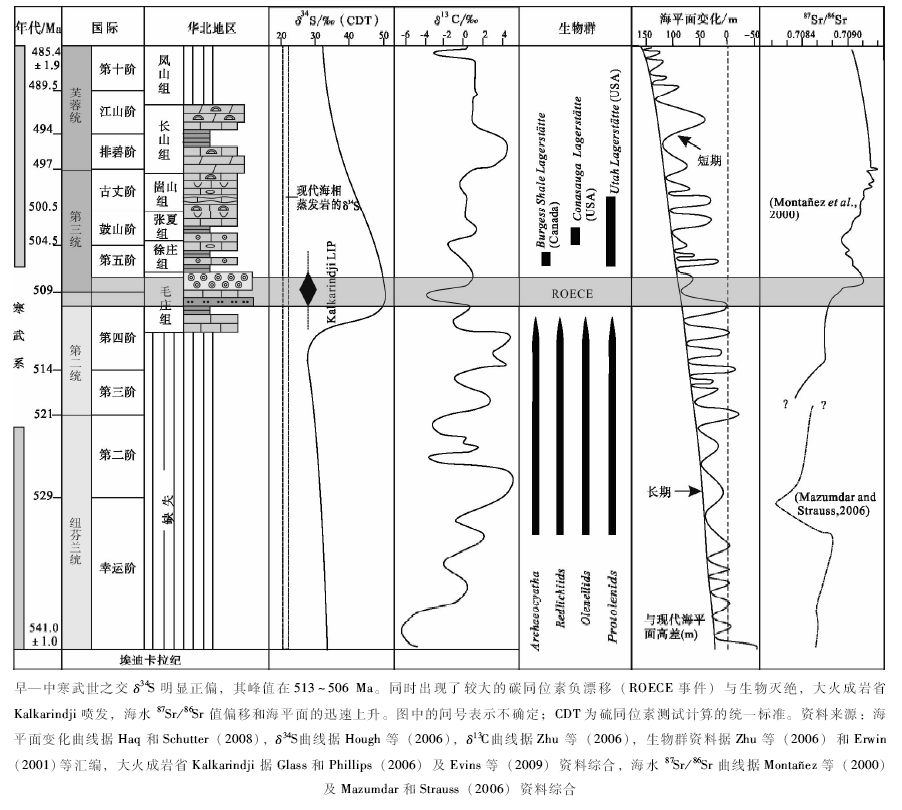 | 图8 早— 中寒武世之交核形石发育与其他地质事件的时间对比Fig.8 Temporal correlation of mass-oncoid occurrence with various events at the Early-Middle Cambrian transition |
如前所述, 早— 中寒武世之交广泛的核形石及其可对比的微生物岩的形成, 首先要求有大量的微生物群发育; 核形石包壳的微组构和快速有机矿化过程要求有活跃的BSR作用; 而包壳纹层中丰富的莓状黄铁矿和异养微生物残余的存在表明为一种缺氧条件。这些证据显示的环境条件与地球化学指标反映的条件一致。海水的低硫酸盐浓度, 最有可能与活跃的BSR作用过量消耗水体中的硫酸盐相关(Visscher and Stolz, 2005; Dupraz et al., 2009)。因此, 在早— 中寒武世之交核形石的广泛发育, 可能反映了微生物群落对该生物集群灭绝事件以及浅海环境普遍缺氧条件的生态响应, 并有可能用于标定该地质事件的层位, 从而进一步提高不同沉积环境背景下的地层对比精度。
早— 中寒武世之交大火成岩省喷发、全球温度升高、海洋缺氧条件扩展、生物灭绝以及微生物岩广泛发育这些事件在时间上的密切相关性(图 8), 以及在显生宙地史中类似情况的多次出现(图 7-a, 7-b, 7-c, 7-d), 可能表明它们之间的确存在着因果关系; 并也有可能说明微生物碳酸盐岩的发育与环境变化有一定的相关性。
华北地台西缘早— 中寒武世之交发育大量的核形石, 主要出现在毛庄组上部和徐庄组底部。核形石包壳内存在丰富的异养微生物残余和莓球状黄铁矿, 表明其可能形成于缺氧的浅海环境。在这个时段, 核形石以及可对比的微生物岩不仅在华北地台广泛发育, 在其他大陆上也有良好记录。它们在时间上与近年发现的大火成岩省喷发、碳同位素负漂移、硫同位素正异常、海水锶同位素偏移和低硫酸盐浓度以及显生宙第1次重要的生物灭绝事件有良好的一致性。可能反映了微生物群落对生物灾变期浅海广泛缺氧环境的生态响应。但这些事件的精确时间标定与确切的相互作用机制仍有待进一步的深入研究。
致谢 中国地质大学(北京)刘典波、李斌、吴金键参加了部分野外工作; 宁夏地矿局郑昭昌、中石油研究院郭彦如高级工程师在野外和室内工作中提供了帮助, 在此致以诚挚的谢意!
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|