


{kind=link}
{kind=link}
河北省白洋淀地区中全新世丽蚌动物群及其生态环境意义
郭书元, 詹伟, 王强. (2014)河北省白洋淀地区中全新世丽蚌动物群及其生态环境意义[J]. 古地理学报, 16(3): 335-346
Guo Shuyuan, Zhan Wei, Wang Qiang. (2014)Middle Holocene Lamprotula fauna from Baiyangdian area, Hebei Province and its environmental significance [J]. Journal Of Palaeogeography, 16(3): 335-346. DOI:10.7605/gdlxb.2014.03.029
Permissions
Guo Shuyuan, Zhan Wei, Wang Qiang. (2014)Middle Holocene Lamprotula fauna from Baiyangdian area, Hebei Province and its environmental significance [J]. Journal Of Palaeogeography, 16(3): 335-346. DOI:10.7605/gdlxb.2014.03.029
Copyright©2014, 《古地理学报》编辑部
版权所有.《古地理学报》编辑部
河北省白洋淀地区中全新世丽蚌动物群及其生态环境意义
第一作者简介 郭书元,男,1937年生,1960年毕业于北京地质学院地层古生物专业,现为中石化华北分公司教授级高级工程师,主要从事地层古生物、沉积、构造、盆地分析和油气勘探方面的研究。
摘要
丽蚌动物群的分布受气候和地理环境控制,研究地层中的丽蚌动物群可以恢复当时的古环境。在河北省白洋淀地区全新统共发现 13种珠蚌科( Unionidae)的化石,属Lamprotula-Cuneopsis-Unio组合,称为白洋淀丽蚌动物群。该动物群与长江中下游流域太湖、鄱阳湖、洞庭湖及其周围水域的现代丽蚌动物群和山东王因、河南西水坡中全新世丽蚌动物群相比,虽然分异度低,但其组合特征相似,同属东洋界中印亚界动物地理区,生活于亚热带湿热气候下的河湖环境中。推测白洋淀丽蚌动物群的生活时代应为中全新世早期,当时北亚热带和东洋界的北界应在白洋淀以北的京津—河套一带。
关键词:
白洋淀地区; 全新世; 丽蚌动物群; 东洋界; 古环境
中图分类号:Q915.5
文献标志码:A
文章编号:1671-1505(2014)03-0335-12
Middle Holocene Lamprotula fauna from Baiyangdian area, Hebei Province and its environmental significance
About the first author Guo Shuyuan,born in 1937,graduated from Beijing Institute of Geology in 1960. Now he is a professor of North China Petroleum Bureau,SINOPEC,and he is engaged in stratigraphy,palaeontology,sedimentary basin analysis and petroleum exploration.
Abstract
As the distribution of Lamprotula fauna was controlled by climate and environment,the study on Lamprotula fauna might reconstruct the palaeoenvironment at that time. 13 species of the Holocene Unionidae fossils were found in Baiyangdian area of Hebei Province,which belonged to Lamprotula-Cuneopsis-Unio assemblage named as Baiyangdian Lamprotula fauna. Compared that fauna with the modern Lamprotula fauna living in Taihu Lake,Poyang Lake,Dongting Lake and their surrounding water in the Yangtze River downstream watersheds,and the Middle Holocene Lamprotula fauna found in Wangyin of Shandong Province and Xishuipo of Henan Province,they all belonged to the China-India Subrealm of Oriental Realm and in lacustrine environment in subtropical humid climate zone,although Baiyangdian Lamprotula fauna contained certain similar characteristics of the mentioned faunas with low diversity in assemblage during the early Middle Holocene. It was indicated that the north boundary of the North Subtropics and the Oriental Realm was located in Beijing-Tianjin area to the north of Baiyangdian,and extended westwards to Hetao area of Inner Mongolia Autonomous Region.
Key words:
Baiyangdian area; Holocene; Lamprotula fauna; Oriental Realm; palaeoenvironment
白洋淀及其周围水域位于华北平原腹地的河北省中部。1957年和1959年, 林振涛和刘月英调查白洋淀地区的现生蚌类时, 在流入白洋淀的唐河古河道砂层及湖底的第四系中采集到蚌类化石, 分别是Unio douglasiae(圆顶珠蚌), Arconaia mutica(反扭蚌), Cuneopsis capitata (巨首楔蚌), Lamprotula zonata(环带丽蚌), L. rochechouarti(猪耳丽蚌), L. spuria(拟丽蚌), L. tortuosa(失衡丽蚌), L. leai(背瘤丽蚌)等8种(林振涛和刘月英, 1963)。2001年, 王强在作华北第四系地质调查时, 在安新县唐河全新统发育平行层理、板状交错层理和槽状交错层理的古河道边滩砂层中(图1), 采集到一批蚌类化石, 经郭书元鉴定有Unio douglasiae, U.tschiliensis(河北珠蚌), Arconaia lanceolata(矛形扭蚌), A.mutica, Cuneopsis heudei(圆顶楔蚌), C. pisciculus(鱼尾楔蚌), Lamprotula (Lamprotula)fibrosa(丝绢丽蚌), L.(L.)zonata, L.(Odhnerella)rochechouarti和 L.(Sinolamprotula)leai 等10种(郭书元, 2012)。因此, 在白洋淀全新统共采集到蚌科化石13种:Unio douglasiae, U.tschiliensis, Arconaia lanceolata, A. mutica, Cuneopsis capitata, C.heudei, C.pisciculus, Lamprotula (Lamprotula) fibrosa, L.(L.)zonata, L.(Odhnerella)rochechouarti, L.(Parunio)spuria, L.(P.)tortuosa, L.(Sinolamprotula)leai等(图 2)。文中主要根据该化石群的组合特征, 恢复了白洋淀地区中全新世的动物群及其生态环境。
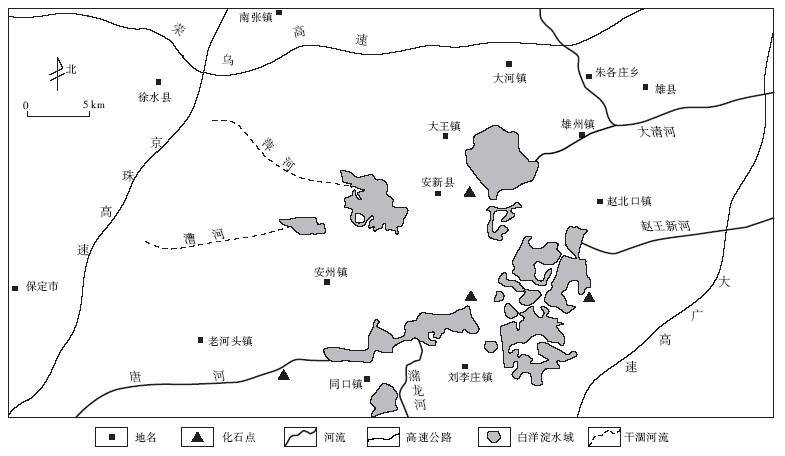 | 图1 河北省白洋淀丽蚌动物群发现位置Fig.1 Discovery location of Baiyangdian Lamprotula fauna in Hebei Province |
 | 图2 河北省白洋淀丽蚌动物群中的典型化石照片Fig.2 Photos of typical fossils from Baiyangdian Lamprotula fauna in Hebei Province |
1 白洋淀丽蚌动物群特征
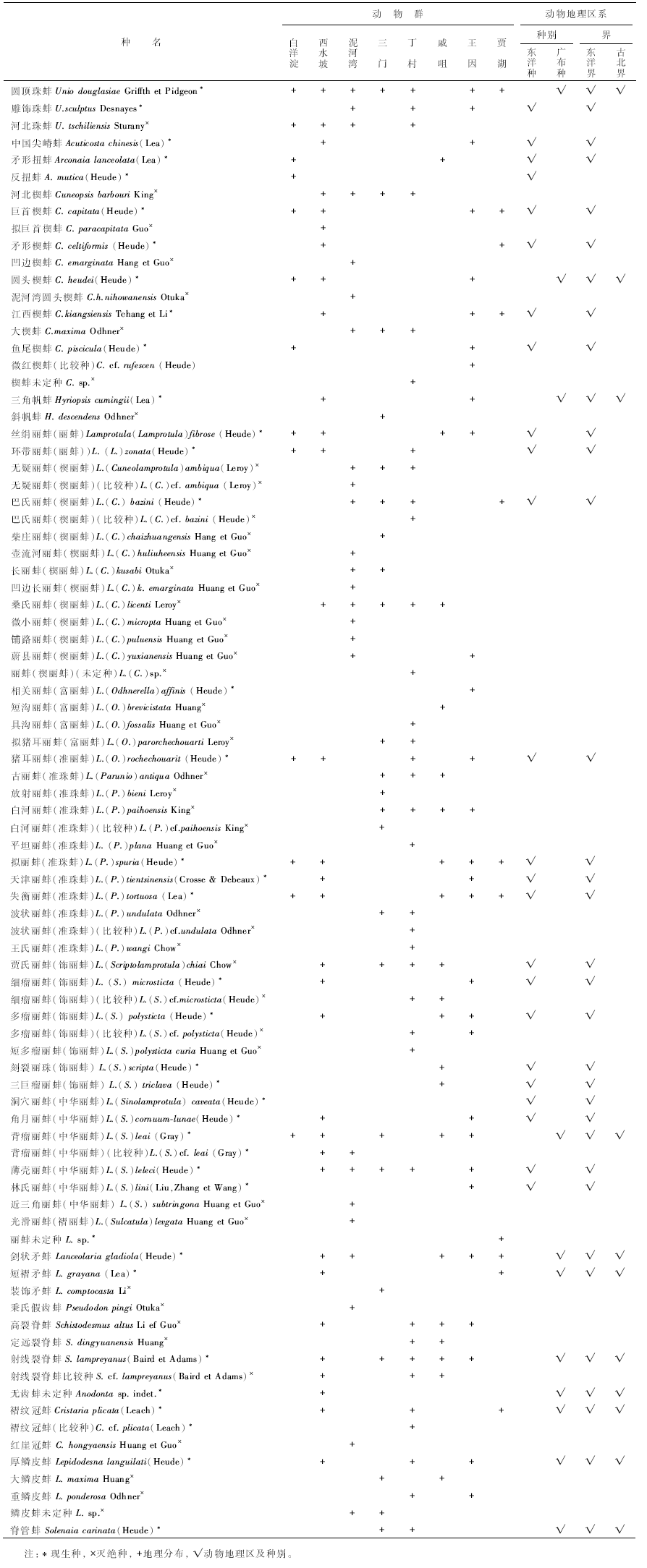
白洋淀地区已发现的全新世13种蚌类化石均属于珠蚌科的珠蚌亚科, 其中:6种属于Lamprotula(丽蚌属)的4个亚属, 占总种数的46.2%; 3种属于Cuneopsis(楔蚌属), 占总种数的15.3%; Unio(珠蚌属)和Arconaia(扭蚌属)各2种(表 1)。因此, 该动物群属于Lamprotula-Cuneopsis-Unio组合, 称为白洋淀丽蚌动物群。该动物群分异度低, 可能与未系统采集化石有关。
| 表1 河北省白洋淀丽蚌动物群地理分布及动物地理区 Table1 Geographical distribution and animal geographical province of Baiyangdian Lamprotula fauna in Hebei Province |
在该动物群中, Lamprotula是其第一优势属, 是该动物群标志属。该属是东亚、南亚地区特有的种群, 在中国有6个现生亚属, 白洋淀丽蚌动物群出现有4个:L.(Lamprotula)(丽蚌亚属), L. (Odhnerella)(富丽蚌亚属), L.(Parunio)(准珠蚌亚属)和L. (Sinolamprotula)(中华丽蚌亚属), 其中L.(Lamprotula)和L.(Parunio)各2种, 为优势亚属, L.(S.)leai, L.(P.)spuria和L.(P.)tortuosa为优势种。Cuneopsis是该动物群的另一优势属, 有3种, C. heudei为优势种。Unio属中以现生种U. douglasiae个体最多, 居该动物群之首。Arconaia属有2种但个体很少, 为稀有种。该动物群的优势种是Unio douglasiae, Cuneopsis heudei, Lamprotula (Parunio)spuria, L.(P.) tortuosa, L.(Sinolamprotula)leai等。
白洋淀丽蚌动物群的13种蚌类化石中, 除Unio tschiliensis外, 均为现生种, 其中一类是地质历史中分布过的种, 包括Unio douglasiae, Cuneopsis capitata, C. heudei, C. pisciculus, Lamprotula (Lamprotula)fibrosa, L.(L.) zonata, L.(Odhnerella)rochechouarti, L.(Parunio)spuria, L.(P.)tortuosa, L.(Sinolamprotula)leai等10种; 另一类未曾在地质历史时期中分布过, 包括Arconaia lanceolata和A.mutica。根据该动物群12个现生种的地理分布(表 1), 可分为4类:(1)广布型:仅Unio douglasiae, 分布几乎遍及中国全境, 北可至俄罗斯远东地区、朝鲜半岛、日本, 南可至东南亚; 该种适应性强, 既适应潮湿气候, 亦适应西北地区干旱气候。(2)亚广布型:包括Cuneopsis heudei和Lamprotula(Sinolamprotula)leai, 分布于华北、淮河和长江中下游流域, 北可至河北白洋淀地区, 南至浙江、江西和湖南等省份, 适应于四季分明的温带中南部和湿热的亚热带。(3)暖温带— 亚热带型:仅Lamprotula(Lamprotula)fibrosa, 分布于淮河下游和长江中、下游流域, 其适应暖温带和亚热带的湿热气候。(4)亚热带型:有Arconaia lanceolata, A. mutica, Cuneopsis capitata, C. pisciculus, Lamprotula (Lamprotula) zonata, L. (Odhnerella)rochechouarti, L.(Parunio)spuria, L. (P.)tortuosa等8种, 分布于长江中、下游流域的江南地区, 只适应亚热带的湿热气候。另外, 该动物群12个现生种中9个种为东洋种(表 1), 故其地理区系应为东洋界中印亚界(林振涛, 1962; 张璽等, 1965; 刘月英等, 1979)。
2 与中国现代瓣鳃类动物群的比较
2.1 与长江中下游流域现代丽蚌动物群的对比
长江中下游流域, 尤其是江南地区鄱阳湖、洞庭湖和太湖及与其相通的周围水域, 是现代丽蚌动物群的主要分布区, 各个湖区的动物群各具特色, 分别称为鄱阳湖丽蚌动物群、洞庭湖丽蚌动物群和太湖丽蚌动物群(表 2)。其共同特征是:(1)分异度高。鄱阳湖和洞庭湖地区丽蚌动物群分异度尤高, 分别为15属50种和亚种、13属41种和亚种, 均属于珠蚌科的珠蚌亚科和无齿蚌亚科; 太湖地区丽蚌动物群分异度稍低, 有12属25种和亚种, 属于珠蚌科。(2)Lamprotula为第一优势种群。在鄱阳湖、洞庭湖和太湖地区的丽蚌动物群中, Lamprotula分别包括6亚属16种、6亚属14种和2亚属5种, 其优势度依次为32.0%、34.1%和20.0%; 其他优势属为Unio, Cuneopsis, Acuticosta, Lancelaria和Anodonta。(3)东洋种的数量较多。在鄱阳湖、洞庭湖和太湖地区丽蚌动物群中分别为37种、28种和12种, 占比依次为74.0%、70.0%和48.0%, 且均为中国特有种。(4)主要分布于长江中、下游流域的江南亚热带地区。因此, 上述动物地理区系均归于东洋界, 属于中印亚界华中区, 其生存于亚热带潮湿气候下的河湖环境中(林振涛, 1962; 张璽等, 1965; 刘月英等, 1979)。
| 表2 白洋淀丽蚌动物群与长江中下游流域现代丽蚌动物群的对比 Table 2 Comparison of Baiyangdian Lamproyula fauna to modern Lamproyula fauna in middle-lower reaches of Yangtze River |
比较而言, 白洋淀丽蚌动物群虽然分异度远远低于上述3个长江中下游流域现代丽蚌动物群, 但其组合面貌相似, 尤其与洞庭湖和鄱阳湖地区的丽蚌动物群更接近。白洋淀丽蚌动物群的4个属均出现于3个长江中下游流域现代丽蚌动物群中, 第一优势属均为Lamprotula, 而Cuneopsis和Unio亦为共同优势属; 12个现生种全部出现于鄱阳湖丽蚌动物群中, 9个现生种出现于洞庭湖丽蚌动物群中, 4个现生种出现于太湖丽蚌动物群中, 分别占该3个动物群种数的100%﹑75.0%和36.7%; 白洋淀丽蚌动物群的5个优势种全部与鄱阳湖丽蚌动物群共有, 4个优势种与洞庭湖和太湖丽蚌动物群共有; 9个东洋种全部出现于鄱阳湖丽蚌动物群中, 6个东洋种出现于洞庭湖丽蚌动物群中, 2个东洋种出现于太湖丽蚌动物群中(表 2)。因此, 推测白洋淀丽蚌动物群与长江中下游流域现代丽蚌动物群的动物地理区系和生存环境相同。
2.2 与华北地区现代瓣鳃类动物群对比
华北地区现代瓣鳃类动物群以河北白洋淀和山东南四湖的瓣鳃类动物群为代表, 其共同点是:(1)分异度较低。南四湖及与其相相通的水域有16个种和亚种, 包括贻贝科1属1种, 珠蚌科9属11种, 其中珠蚌亚科7属8种(Unio douglasiae, Acuticosta chinensis* , Cuneopsis heudei* , Hyriopsis cumingii* , Lamprotula(Sinolamprotula)leai, Lanceolaria gladiola, L. grayana, Schistodesmus lampreyanus), 无齿蚌亚科2属3种和亚种(Anodonta arcaeformis, A. woodiana, Cristaria plicata)。白洋淀及其周围水域共有14个种和亚种, 均属珠蚌科, 其中珠蚌亚科5属6种(Unio douglasiae, Cuneopsis heudei* , Hyriopsis cumingii* , Lamprotula (Sinolamprotula) leai, Lanceolaria gladiola, L. grayana), 无齿蚌亚科1属4种4亚种(Anodonta arcaeformis, A.a.flavotina, A. euscaphys* , A. fluminea* , A. woodiana woodiana, A. w. elliptica* , A. w. pacifica* , A. w. piscatora等)(带* 者为中国特有种, 其余种皆广布于东亚、东南亚)。(2)白洋淀动物群的代表属Lamprotula仅出现L.(Sinolamprotula)leai 这1个广布种, 是该动物群的次要分子。(3)除Acuticosta chinensis为东洋种外, 其余均为广布种, 其动物地理区属古北界, 其中南四湖现代瓣鳃类动物群属古北界华北区黄淮平原亚区, 白洋淀现代瓣鳃类动物群属华北区海河亚区, 其均生活于四季分明的中温带, 冬季较冷、气候较干燥(林振涛和刘月英, 1963; 刘月英等, 1986)。
将白洋淀丽蚌动物群与上述2个华北地区现代瓣鳃类动物群相比可知, 虽然3者分异度均低, 亦有共同的属种, 但是组合面貌却完全不同, 不仅Lamprotula在华北这2个瓣鳃类动物群中均不是优势属, 仅见1个广布种L.(S.)leai, 而且这2个现代瓣鳃类动物群中仅南四湖动物群出现1个东洋种Acuticosta chinensis。因此, 白洋淀丽蚌动物群和现代华北地区瓣鳃类动物群不属于同一动物地理区, 后者为古北界; 气候环境亦完全不同, 后者属于北温带气候。
3 与中国第四纪丽蚌动物群的对比
中国第四纪丽蚌动物群分布较多。早更新世早期以河北泥河湾丽蚌动物群为代表, 早更新世中期以豫西、晋南、山西襄汾和陕西渭南三门丽蚌动物群为代表; 中更新世以山西襄汾丁村丽蚌动物群为代表; 晚更新世以苏北泗洪和皖北五河、定远一带的戚咀丽蚌动物群为代表; 中全新世已发现3个新石器时代遗址的丽蚌动物群:山东兖州王因丽蚌动物群、河南濮阳西水坡丽蚌动物群和河南舞阳贾湖丽蚌动物群(表 3)。
| 表3 白洋淀丽蚌动物群与中国其他第四纪丽蚌动物群对比 Table 3 Comprison of Baiyangdian Lamprotula fauna to other Quaternary Lamprotula fauna in China |
3.1 与更新世丽蚌动物群的对比
将白洋淀丽蚌动物群与中国更新世早、中期的泥河湾、三门和丁村丽蚌动物群相比发现:其均以Lamprotula为第一优势属和标志属, Unio和Cuneopsis为优势属, 组合特征近似; 但中国更新世丽蚌动物群具古老的组合面貌, 其灭绝率很高, 而白洋淀丽蚌动物群仅有1个灭绝种, 灭绝率0.7%; 同时中国更新世丽蚌动物群的分异度远高于白洋淀丽蚌动物群。
早更新世早期泥河湾丽蚌动物群共发现有24种, 均属珠蚌科, 其中Lamprotula(13种)的优势度高达54.0%, 但其分化程度低, 仅3个亚属且1个已经灭绝, 古老的亚属L. (Cuneolamprotula)有9种, 其中8种已灭绝。该动物群灭绝种20个, 灭绝率高达83.3%, 显示了古老面貌, 与白洋淀丽蚌动物群的面貌极其不同(黄宝玉和郭书元, 1982)。
早更新世中期三门丽蚌动物群共发现有23种, 均属于珠蚌科, 其中珠蚌亚科6属20种, 无齿蚌亚科2属3种, Lamprotula(14种)的优势度高达60.9%, 其分化程度较高, 有5个现生亚属, 其中古老的L.(Cuneolamprotula)有5种, 而且均已灭绝。该动物群包括17个灭绝种, 其灭绝率高达73.9%, 亦显示了古老性质(李云通, 1965; 黄宝玉和郭书元, 1982; 黄宝玉等, 1991)。
中更新世丁村丽蚌动物群共发现有35种, 均属珠蚌科, 以珠蚌亚科为主, Lamprotula(17种)的优势度高达57.1%; 其分化程度高, 与现代丽蚌动物群有6个亚属相同, 但其中以古老的L. (Cuneolamprotula)占优势(5种, 4种已灭绝)。该动物群灭绝种24个, 灭绝率为68.6%, 显示了较古老的组合面貌(周明镇, 1958; 黄宝玉等, 1991)。
晚更新世戚咀丽蚌动物群共有20种, 均属于珠蚌科, 其中珠蚌亚科4属19种, 无齿蚌亚科1属1种。Lamprotula有13种, 为第一优势属, 优势度为65.0%, 其他属各1种。该动物群中Lamprotula分化程度高, 与现代丽蚌动物群的6个亚属完全相同。其现生种中有10个东洋种, 占总种数的58.8%。白洋淀丽蚌动物群与其相比, 均以Lamprotula为第一优势属、含有高比例的东洋种为共同特征, 差异是后者的分异度较高, 尤其是有7个灭绝种, 灭绝率为35.0%, 亦显示了较古老的面貌(周明镇, 1955; 王淑梅, 1961; 黄宝玉, 1983)。
3.2 与中全新世早期丽蚌动物群的对比
已发现的中国中全新世早期丽蚌动物群有河南贾湖、河南西水坡和山东王因3个, 虽然均为新石器时代遗址出土的动物群, 但是其与长江中下游地区3个现代丽蚌动物群组合面貌极其相似。
将白洋淀丽蚌动物群与中全新世早期山东王因丽蚌动物群和河南西水坡丽蚌动物群相比发现:均以Lamprotuila为第一优势属, Unio, Cuneopsis均为优势属; 白洋淀丽蚌动物群中除Arconaia的2个种外, 均出现于中全新世的山东王因丽蚌动物群和河南西水坡丽蚌动物群中, 而且各动物群中均以东洋种为主体。其差异是后两者的分异度远高于白洋淀丽蚌动物群, 而且含有较多的灭绝种, 而白洋淀动物群仅有1个灭绝种。山东王因丽蚌动物群共有9属30种, 均属于珠蚌科, 其中珠蚌亚科8属29种, 无齿蚌亚科1属1种(表 3), Lampritula为优势属, 多达16种, 优势度为53.3%, Unio和Cuneopsis亦为优势属; 代表属Lamprotula分化程度高, 有6个现生亚属(郭书元等, 2000)。该动物群灭绝种6个, 灭绝率20.0%。河南西水坡丽蚌动物群有32种, 均属珠蚌科, 其中珠蚌亚科7属30种, 无齿蚌亚科3属3种以上(表 3), 代表属Lamprotula有14种之多, 优势度为43.7%, 其他优势属主要有Unio, Cuneopsis, Lanceolaria等; Lamprotula分化程度高, 具有全部的6个现生亚属, 以L. (Parunio), L.(Sinolamprotula)为优势亚属(郭书元和孙长虹, 2003; 郭书元, 2012)。该动物群灭绝种8个, 灭绝率25.0%。
贾湖丽蚌动物群分布于河南舞阳县贾湖新石器时代遗址。该动物群分异度和丰度均低, 可鉴定到种的标本仅11种, 均属珠蚌科, 其中珠蚌亚科4属10种, 无齿蚌亚科仅1属1种(表 3)。其优势属为Lamprotula 有4种, 优势度为36.3%。灭绝种1个, 灭绝率9.1%。Lanceolaria 共2种, 为现生广布种, Unio仅1个现生广布种, 属于Cristaria的标本可能属于现生广布种C.plicata。其中Lamprotula的4个种中有3个为现生种, 均为东洋种, L.sp.可能为灭绝种。Cuneopsis的3个种均为现生种, 其中C. heudei为广布种, 其他2种为东洋种(河南省考古研究所, 2000)。与白洋淀丽蚌动物群相比, 其分异度均低, 组合特征相同, 东洋种均占主要地位; 其差别在于白洋淀丽蚌动物群未发现Lanceolaria的分子, 贾湖丽蚌动物群不含Arconaia的分子。
因此, 白洋淀丽蚌动物群与中全新世早期的3个丽蚌动物群应同属东洋界动物地理区, 生活环境相似, 生存时代亦应相同。
4 古环境和动物地理区系
4.1 古生态
据研究, 早更新世早期泥河湾丽蚌动物群、早更新世中期三门丽蚌动物群、中更新世丁村丽蚌动物群和晚更新世戚咀丽蚌动物群的生活环境为湿热气候条件下的河湖环境, 其主要生活于三角洲前缘分流河道砂质、砂砾质、含砾砂泥质底床和浅湖— 半深湖灰绿色、灰色含砂泥质、深灰色泥质底床环境中(周明镇, 1955, 1958; 李云通, 1965; 黄宝玉和郭书元, 1981, 1982; 黄宝玉, 1983)。山东王因和河南西水坡中全新世丽蚌动物群生存环境与现代长江中下游江南地区相似, 均为亚热带湿热气候条件下的河湖环境中(郭书元等, 2000; 郭书元, 2012)。
中国现代丽蚌动物群主要分布于长江中下游流域江南地区大、中型湖泊及与其相通的河流中, 如洞庭湖和与其相通的湘、资、沅、澧4大水系, 鄱阳湖及与其相通的赣江、信江、修水和抚河, 太湖及与其相通的大运河和诒溪等; 另外, 武汉附近的梁子湖、南京附近的水域亦有分布。动物群中珠蚌亚科的多数种类喜流水, 甚至激流, 如Arconaia mutica, Cuneopsis capitata, C. heudei, Lamprotula (Lamprotula)fibrosa, L.(L.)zonata, L.(Odhnerella)rochechouarti, L.(Parunio)spuria, L.(P.)tortuosa等, L.(Sinolamprotula)leai既适应急流亦适应缓流, 而Unio douglasiae不仅喜急流和缓流, 而且亦适应静水湖泊; Lamprotula的一些种类则生活于湖泊三角洲前缘水下分流河道中, 有的则习惯生活于长江与大湖的通道中。就水深而言, 它们均生活于浅水区域(按小于10 m为浅湖), 但水深的要求不同, 有的种适应较浅水域, 如Cuneopsis heudei生活于1~2 m深的水域, Unio douglasiae要求1~3 m水深, Lamprotula的种适应较深水域, 甚至超过3 m。对于水质均要求具有一定矿化度的中性水, 或偏碱性水, 多喜无水草孳生的较清的水。要求水体的底质多为稍硬的泥沙底、沙泥底、沙底, 有的要求硬质砾质底, 无齿蚌多喜软泥底。总之, 长江中、下游现代丽蚌动物群的生存环境属于三角洲分流河道— 湖泊环境(林振涛, 1962; 刘月英等, 1965, 1979, 1980; 张璽和李世成, 1965; 张璽等, 1965)。
白洋淀丽蚌动物群古生态是根据它们产出地层的岩性和现生种的生态资料确定的, 其古生态应与上述动物群相似, 其整体生存环境亦为三角洲前缘分流河道— 湖泊环境。
4.2 古气候
由于白洋淀丽蚌动物群与现代长江中下游流域的丽蚌动物群和中全新世早期山东王因丽蚌动物群、河南西水坡丽蚌动物群组合特征相似, 同为东洋界动物地理区, 故其生存的气候环境亦应同为湿热的亚热带。另外, 在2个新石器时代遗址出土的兽骨和地层中发现的孢粉化石亦说明中全新世早期的气候为湿热的亚热带。中全新世早期山东王因丽蚌动物群共同出土的兽骨有Bubalus bubalus(水牛), Cervus unicolor(水鹿), Elaphurus dacidianus(四不象), Alligotor sinensis(扬子鳄)等热带、亚热带常见分子, 其中Alligotor sinensis现代仅生存于长江中下游流域(周本雄, 2000a, 2000b); 在王因遗址77SYWT4016探方的孢粉样品中分析出Quercus(栎), 其为落叶阔叶乔木, 亦发现生长于热带和亚热带的草本蕨类Pteris(风尾蕨)和Lygodium(海金沙), 还有可能属于Oryza sativa(稻)的禾本科花粉, 其中Pteris、Lygodium在形态上可与现代江苏和安徽等省的Pteris vittata(蜈蚣草)、Lygodium japonicum(海金沙)的孢子比较(孔昭辰和杜乃秋, 2000)。上述动植物的生存环境应是亚热带地区。中全新世早期河南西水坡丽蚌动物群出土的兽骨有Bubalus bubalus, Alligotor sinensis和Cervus unicolor等, 亦反映了亚热带湿热气候环境(郭书元等, 2003; 郭书元, 2012; 吕鹏等, 2012)。对内蒙古伊克昭盟第四系钻井岩心研究认为, 中全新世7.5~6.3 ka BP为大暖期, 6.3~4.2 ka BP为气候波动期, 4.2~2.3 ka BP为暖期(魏东岩, 1997)。因此, 白洋淀丽蚌动物群生存的气候环境应为亚热带湿热气候, 同时说明中国华北北纬39° 一带, 中全新世早期远比现代湿热, 属亚热带气候(赵锡文, 1982; 郭书元等, 2000, 2003; 郭书元, 2012)。
4.3 古地理
白洋淀流域处于华北平原冀中坳陷的沉降中心, 是华北的汇水区域, 20世纪期间的明水面积达333 km2以上(林振涛和刘月英, 1963), 由西北、西、西南呈放射状流入白洋淀水域的季节性和常年河流曾经多达十余条。但是, 目前仅存8条河流注入白羊淀内, 且入淀径流量很小, 基本都已断流, 2007年湿地和明水面积仅180 km2, 曾几度趋向干淀(庄长伟等, 2011)。这主要是受气候变化控制, 另外人类的围田造地、过度开发地下水使地下水位严重降低, 亦加快了环境的恶化。白洋淀丽蚌动物群生存时期的古地理环境与现代相比, 地貌景观变化不大, 均地处华北平原的腹地, 西面是太行山, 东距渤海比现代要近, 是华北盆地冀中坳陷沉降中心。当时地处亚热带, 气候湿热、雨量充沛, 与其相通的河流比近代多, 且均为常年性河流, 水量亦大得多, 地下水位亦高。充沛的水源、很浅的潜水面使白洋淀的水域比现代宽广很多。适宜的气候、草木繁茂的平原和广阔的河湖水域, 为喜湿热的丽蚌动物群提供了优越的生存环境, 它们可栖息于河道、河口区、口岸内和湖区。
4.4 动物地理区系
由上述可知, 白洋淀丽蚌动物群的动物地理区系应与现代长江中下游流域丽蚌动物群和中全新世山东王因丽蚌动物群、河南西水坡丽蚌动物群相同, 同属东洋界中印亚界华中区。
5 白洋淀丽蚌动物群的时代
白洋淀丽蚌动物群的产出地层表明其生存时代早于现代, 然而由于其组合特征接近于现代长江中下游流域的丽蚌动物群, 其时代亦不会太早。与中国其他地区更新世丽蚌动物群相比, 白洋淀丽蚌动物群组合面貌相差较大, 缺少古老属种, 且后者灭绝率高、组合面貌古老。所以, 白洋淀丽蚌动物群的生存时代晚于更新世, 为全新世。由于该动物群动物地理区系和生态环境与山东王因丽蚌动物群、河南濮阳仰韶文化早期西水坡丽蚌动物群相同, 故应为同时代动物群。后两者的时代为中全新世早期, 14C测年分别为5950± 125 a BP和6400± 135 a BP(郭书元等, 2000; 郭书元和孙长虹, 2003; 郭书元, 2012)。河南贾湖丽蚌动物群产于裴里岗文化期, 时代为中全新世初期(7000 a BP左右)。因此, 白洋淀丽蚌动物群的时代应为中全新世早期。由此可以推断, 中全新世早期北亚热带的北界应在北纬39° 线以北, 可能在京津至河套一线。该时期的动物地理区划亦不同于现代, 东洋界的北界亦可能在天津— 北京, 向西可延至河套地区(郭书元等, 2000; 郭书元, 2012)。
6 结论
1)由于没有系统采集, 导致白洋淀丽蚌动物群分异度底。但是, 其组合面貌与现代长江中下游地区太湖、鄱阳湖和洞庭湖流域的3个丽蚌动物群的组合面貌相似, 应同属东洋界中印亚界华中区, 当时的北界应在北纬39° 一带。
2)白洋淀丽蚌动物群反映的生态环境为湿热气候条件下的三角洲前缘水下分流河道— 湖泊环境。
3)根据白洋淀丽蚌动物群反映的气候、产出层位及其完全可与河南贾湖、河南西水坡和山东王因新石器时代遗址出土的丽蚌动物群对比, 确定其时代为中全新世早期。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
参考文献
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|