








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
杭州湾庵东浅滩现代沉积物中的生物遗迹
[宋慧波1, 2  , 于会新
, 于会新1 , 王海邻1 , 范代读3 , 胡斌1, 2 , 王芳1 ]
, 于会新, 王芳|
|

通讯作者简介 胡斌,男,1952年生,湖北武汉人,河南理工大学资源环境学院教授,博士生导师,主要从事古生物学、遗迹学和沉积学教学与研究工作。E-mail: hub@hpu.edu.cn。
第一作者简介 宋慧波,女,1979年生,山西长治人,河南理工大学资源环境学院副教授,博士,主要从事古生物学、遗迹学和沉积学教学与研究工作。E-mail: songhuibo2005@hpu.edu.cn。
杭州湾庵东浅滩沉积物中发现有多种造迹生物,主要有蝼蛄、泥甲虫、宁波泥蟹、珠带拟蟹守螺、泥螺、弹涂鱼、蚂蚁和鸟类等,营造的生物遗迹主要为觅食迹、居住迹和爬行迹(爬迹、拖迹和鸟足迹)等。分析表明,浅滩不同亚环境的造迹生物及其遗迹的主要类型和分布特征具有明显的差异性: ( 1)在潮上带植被茂密的草滩环境中,以层面上的生物觅食迹、居住迹和鸟足迹为主,造迹生物主要有蝼蛄、泥甲虫、蚂蚁和鸟类等,其中以蝼蛄的潜穴最为丰富。( 2)在高潮滩的积水泥沼环境中,有大量近海喜盐植物,生物遗迹主要有宁波泥蟹的“ J”形、“ W”形和“ Y”形潜穴、弹涂鱼的羽状爬迹、珠带拟蟹守螺在层面上营造的弧形、圆圈状的觅食迹和“ C”形的条带状生殖迹。( 3)在中、低潮间带的粉砂质环境中,以层面上珠带拟蟹守螺不规则的觅食迹为主。生物遗迹的分布与浅滩沉积底质沉积物粒度与含水量等特征密切相关。本研究不仅丰富了杭州湾浅滩沉积的新遗迹学研究,而且对古代潮汐沉积及其环境研究有重要的借鉴意义。
About the corresponding author Hu Bin,born in 1952,is a professor and Ph.D. supervisor of Henan Polytechnic University. He is mainly engaged in teaching and researches on paleontology,ichnotology and sedimentology. E-mail: hub@hpu.edu.cn.
About the first author Song Huibo,born in 1979,is an associate professor and Ph.D. of Henan Polytechnic University. She is mainly engaged in teaching and researches on paleontology,ichnology and sedimentology. E-mail: songhuibo2005@hpu.edu.cn.
Various trace-makers have been found in the shoal deposits of Andong area,Hangzhou Bay,including mole crickets( Gryllotalpa unispina),mud beetles(Coleptera),mud crabs( Ilyplax ningboensis), Cerithideopsilla cingalata, mud snails( Bullacta),mudskippers or amphibious fishes( Periophthalmua cantonensis),ants and birds. The most abundant traces can be ascribed to Fodinichnia,Domichnia,Repichnia and Pascichnia. Analysis shows that significant variations in type and diversity of traces and their makers are present over the different shoal zonations. (1)The highly-vegetated supratidal flat is featured by the abundance of bird tracks,and feeding and dwelling traces of mole crickets,mud beetles and ants,in which the feeding burrows of mole cricket are the most common. (2)In the upper intertidal flat where a lot of halophilic plants dominate on the muddy substrates,the biogenic traces include the J-,W-or Y-shaped burrows by mud crabs,the pinniform crawling traces by mudskippers,and the arc-shaped,donut-shaped grazing trails and C-shaped reproduction traces by mud snails ( Cerithideopsilla cingalata).(3) In the silty substrate of the middle-lower intertidal zones,the biogenic traces are characterized by the irregular grazing trails of mud snails ( Cerithideopsilla cingalata) on the bedding plane. It is also found that the distribution of biogenic traces is highly related to the grain sizes and water-saturation of the substrates. This study provides not only a neoichnological approach to tidal-flat sedimentary in the Hangzhou Bay,but also a meaningful modern analogue for sedimentary facies and paleo-environmental interpretations of ancient tidal flats.
沉积物中生物遗迹的记录是生物与环境相互作用的结果, 也是沉积环境及生态环境的良好指示标志。对现代生物遗迹的研究是通过对现代各类生物的造迹活动进行详细的观察和描述, 了解各类造迹生物的活动规律、遗迹特点与分布特征及与沉积底层的关系, 进而分析其受控的环境因素, 从而对古遗迹学的研究有所借鉴和指导(胡斌等, 1997; Genise et al., 2000; Gingras et al., 2004; Wetzel, 2008)。早在20世纪30年代德国科学家Othenio Abel和Rudolf Richter教授等对德国西北部北海(Wadden Sea)和波罗的海潮坪上的现代底栖生物遗迹做了大量观察和研究工作, 并出版了《北海潮坪的实证古生物学》一书(Schä fer, 1962), 为生物遗迹学和海洋沉积学的发展做出了重要贡献, 这也是最早有关现代海岸潮坪生物遗迹群落的研究。到20世纪70— 80年代, 以James Howard和Robert Frey为首的新遗迹学研究者, 对美国北卡罗莱纳(Frey, 1970)和佐治亚海岸等潮坪、河口、盐沼环境下的底栖生物遗迹进行了系统研究(Frey and Basan, 1978; Frey and Howard, 1986), 并对韩国仁川现代潮坪的生物遗迹进行了研究(Frey et al., 1987)。尤其是德国图宾根大学的Seilacher教授, 20世纪50— 60年代就在Wadden海开展了较长时间的现代生物遗迹研究, 详细描述了造迹生物的造迹过程, 这在遗迹学行为分类和遗迹化石生态学分类的发展上具有里程碑的作用, 并提出了建立遗迹相的基本原则(Seilacher, 1954, 1964, 1967; McMenamin, 1994; Goldring et al., 2000)。由此可见, 国外在上世纪已有大量针对河口湾和潮坪沉积物中现代生物遗迹的系统研究。中国较早的新遗迹学研究包括范振刚(1976)对青岛潮间带底内蟹类生态及洞穴进行的观察和分析, 王慧中(1985)研究了江浙一带现代海滩的生物扰动构造并讨论了其指相意义, 王珍如等(1988, 1994)对青岛、北戴河及北部湾等地潮间带现代底内生物的遗迹进行了观察和描述, 王英国(2000)对渤海湾西岸大石河河口湾遗迹的生态学进行了详细研究, 胡斌等(2012)考察和研究了黄河中下游及黄河三角洲现代生物遗迹群落。总体上讲, 中国新遗迹学包括现代海岸潮坪沉积环境中的生物遗迹研究起步较晚, 与西方发达国家相比, 还有较大差距。就杭州湾庵东浅滩现代沉积方面的研究而言, 前人已做过大量沉积学方面的工作(李炎和谢钦春, 1993; 郭艳霞等, 2004; 范代读等, 2005), 但该区域有关现代生物遗迹的研究几乎是空白, 迄今未见正式报道。
庵东浅滩位于杭州湾南岸浅滩的突出段, 是杭州湾内近数百年的冲淤过程中形成的主要淤积区之一。作者以浙江省慈溪市庵东镇(30° 21.759'N, 120° 10.533'E)一带的砂泥质浅滩为研究对象。在综合分析前人沉积学研究资料的基础上, 运用遗迹学的研究方法, 较详细地观察和描述了该区典型沉积单元中潮上带和潮间带的造迹生物及其遗迹, 并分析了生物遗迹的空间分布与底质沉积物成分、结构和含水率的关系。本研究一方面为庵东浅滩生物成因沉积构造研究提供新资料, 另一方面也能为地史时期河口湾沉积环境与古生态的研究提供现代生物学和生态学借鉴。
庵东镇位于杭州湾南岸浅滩的凸出段, 为强潮环境下凸岸边滩式浅滩堆积体。该浅滩东邻湾口潮汐通道, 西至钱塘江河口, 呈扇形向北突出, 是现代杭州湾浅滩的主体(图 1)。该区地处中纬度亚热带季节性气候区的滨海地带, 气温受冷暖气团交替控制以及杭州湾海水调节, 四季分明, 温暖湿润。年平均气温约为16 ℃; 因受海洋性季风影响, 该地区雨量充沛, 多年平均降水量为1351.1 mm; 年平均日照约2078.3 h, 年平均湿度为18%。该区大部分为围合的浅滩, 拥有潮上带浅滩宽2000~3000 m, 潮间带浅滩宽7000~8000 m。受潮汐作用和波浪共同作用的影响, 庵东浅滩在平面分布上可划分为潮上带、潮间带和潮下带3种地貌单元。浅滩沉积物以粉砂、黏土质粉砂为主, 含盐量较高。
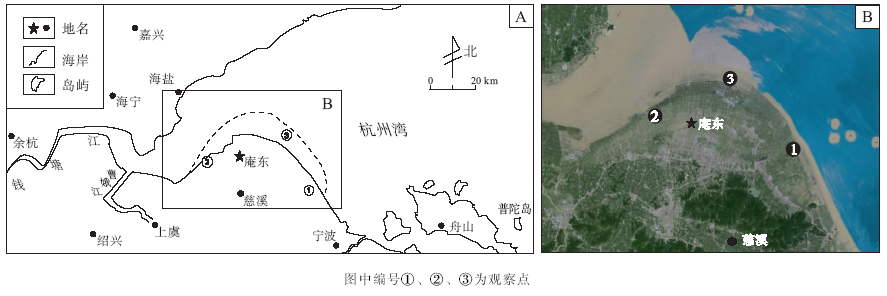 | 图1 杭州湾庵东地区现代潮坪及考察点位置Fig.1 Location of modern tidal flats and fieldwork sites in Andong area of Hangzhou Bay |
杭州湾湾顶地区水浅流急, 落潮时高含沙量水体下泄至庵东浅滩时, 因断面展宽, 水流扩散, 挟沙力降低, 泥沙大量落淤形成宽广的浅滩(刘苍字和董永发, 1990)。庵东浅滩呈扇形向海湾突出, 滩面淤涨受制于涨落潮动力强弱的控制。受长江入海泥沙大量输入的影响, 该区粉砂组分相对集中, 并成为浅滩沉积的主体。
根据庵东浅滩滩面物质及微地貌特征, 可划分为草滩、泥滩和粉砂滩3种微地貌单元, 结合潮位变化可细分为:(1)潮上带盐沼或淡水沼泽(图 1, ①), 在高潮线之上, 因经常遭受人类活动影响, 如围垦后再直接受到潮汐作用的影响, 受雨水冲洗等影响使其表层沉积物含砂量增加。(2)高潮泥沼环境(图 1, ②), 水动力较弱, 低洼处易积水, 生长有近海喜盐植物, 以泥质沉积为主, 且富含有机质。(3)中— 低浅滩(图 1, ③)为粉砂滩, 主要由灰色粉砂构成, 黏土体积分数为3%~12%, 砂体积分数为5%~20%, 见水平纹层。层面上可见少数生物潜穴孔, 自西向东沉积物粒度变细。
受围垦影响, 当前研究区内浅滩大部分为平缓的粉砂滩, 仅在近岸塘脚处有范围较小的草滩和泥滩。在杭州湾跨海大桥以东, 浅滩东侧的地貌组合向粉砂滩过渡。随着浅滩宽度的减小, 平均坡度渐增至0.08%左右, 泥滩与粉砂滩分界线向低潮位方向移动, 位于小潮平均高潮位附近。
庵东浅滩沉积物包括粉砂与黏土质粉砂2大类。黏土质粉砂分布于近岸一侧; 细砂质粉砂分布于滩坡及外侧冲刷槽处, 粉砂质沉积广泛分布于浅滩。沉积物中黏土、粉砂和细砂3种粒级成分中, 粉砂含量为45%~99%, 是优势成分; 黏土含量达1%~55%, 大部分小于40%; 细砂含量为0%~80%, 大部分小于5%。浅滩样品多具有单峰的粒度分布曲线, 中值粒径Md介于5~7.4ϕ , 以5.5ϕ 为多; 分选系数QD为0~1.6, 以0.4居多; 偏态SK为-0.2~+1.0, 以零偏态居多。样品的分选系数和偏态均与中值粒径呈正相关关系。样品的正态概率累积曲线普遍存在着对应于推移、悬移一跃移和絮团状悬移3种运动状态的组分, 其粗细截点多位于4.5ϕ 和5.5ϕ (李炎和谢钦春, 1993)。显然, 在庵东浅滩的2大沉积类型中, 粉砂沉积类型具较粗的中值粒径, 较好的分选、零偏态, 以悬移一跃移运动占优势等特征。黏土质粉砂沉积中值粒径较细, 分选较差, 正偏态, 以悬移一跃移和絮团悬移为主要的搬运形式。
庵东浅滩沉积层序中以非生物成因沉积构造为主, 其中较为常见的有薄层砂泥交互层理、波痕、泥裂、雨痕和冲刷— 充填构造等沉积单元(图 2)。在高潮滩及潮上带的沉积物中发育的砂泥薄交互层理(图 2-A), 是由涨、落潮急时形成的砂质沉积与憩流时的泥质沉积组合而成的, 见有大量植物根系。浅滩波痕(图 2-B)是波浪、 潮流等作用于非黏质沉积物而形成的一种微地貌形态, 在中、低浅滩发育较普遍, 且范围大。泥裂常见于潮上带和高潮滩(图 2-C), 是泥质沉积物长期暴露、 干涸、 收缩而成; 雨痕(图 2-D)是雨点打落在未固结的泥质、砂质沉积物表面, 所产生圆形或椭圆形的凹穴。
 | 图2 杭州湾庵东地区浅滩上的常见沉积构造Fig.2 Common sedimentary structures of shoal deposits in Andong area of Hangzhou Bay |
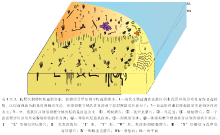
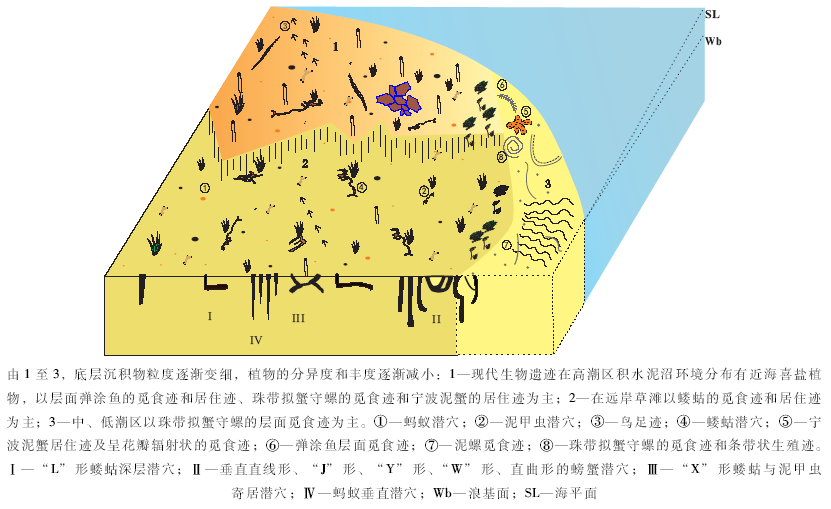
钱塘江及周边支流携带的大量悬移物沉积在湾中庵东浅滩附近, 形成泥砂质底质, 有机质丰富。垂直分布上, 潮区生物量的差异明显:潮上带丰度及分异度极低, 潮间带相对丰富, 造迹生物门类较多且丰度高, 体态偏小, 多以食泥性类为主。这是由于潮汐动力和海浪的水动力作用共同影响, 致使泥砂滩的环境变化明显。落潮后, 由于高、中浅滩的水分流失较快, 条件恶劣, 不利于生物群落的生活, 因此绝大多数生物会将身体潜入松软泥砂中或钻入底质内营穴而居。高潮滩砂质颗粒较粗, 水位低, 营养成分少, 生物多集中在深度大、潮湿的单轴型潜穴中, 摄食底质表面有机物, 并在穴孔附近留下圆形食渣泥球; 觅食迹造迹生物多具带状分布特征(王珍如等, 1994)。造迹生物类型主要包括底表型和底内型2类底栖生物。底表型主要包括腹足类; 底内型以双壳类、节肢类和甲壳类为主。
潮上带远岸草滩造迹生物相对于潮间带造迹生物门类偏少, 主要造迹生物为蚂蚁(膜翅目蚁科)、泥甲虫(鞘翅目长泥甲科)和蝼蛄(直翅目蝼蛄科)以及鸟类(图 3-A), 并发现波痕、雨痕和泥裂等层面构造。其中常见蚂蚁的觅食迹和居住迹、泥甲虫的居住迹、蝼蛄的觅食迹和居住迹等。蚂蚁一般会在地下筑巢, 在出入口处拱起一个由小的圆形土壤颗粒堆积成的小土丘, 像火山堆, 中间有个洞口(图 3-C), 洞口直径为0.5~0.8㎝ , 旁边堆起的土丘直径为5~10 cm, 其洞口也可用于通风; 泥甲虫喜湿, 多钻入土壤浅层来觅食泥土中的腐殖质, 在层面上形成圆柱状隆起、与蝼蛄的潜穴外形上很相似, 有时会相伴而生, 但其潜穴外壁较蝼蛄潜穴外壁光滑, 且潜穴直径仅为蝼蛄潜穴直径的1/2, 宽为0.5~0.8 cm, 深度为0.5 cm, 整体规模约30 cm, 呈直线或分枝状结构(图 3-D), 分枝末端有小孔, 便于通风, 该居住迹同时也是泥甲虫的觅食迹。在潮上带远岸草滩潮湿处还发现有少量蝼蛄的小规模潜穴(图 3-B)。
 | 图3 杭州湾庵东地区潮上带远岸草滩上常见的现代生物遗迹Fig.3 Common modern biogenic traces in grass pool in supratidal zone far away from shoreline in Andong area of Hangzhou Bay |
潮上带的近岸边滩的生物丰度相对远岸草滩要大, 造迹形态也多样。当大的潮水在这里退去, 其所携带的有机质大量沉积, 底质较松散, 便于生物在此造穴营生。造迹生物主要为蝼蛄Gryllotalpa unispina(直翅目), 其次, 由于大潮的冲刷, 在潮上带的近岸草滩还发现有少量的宁波泥蟹Ilyplax ningpoensis(十足目)、弹涂鱼Periophthalmua cantonensis(鲈形目)和泥甲虫(鞘翅目)等。生物遗迹中蝼蛄的觅食迹和居住迹丰度极高, 并伴有泥甲虫的觅食迹和居住迹, 少量的宁波泥蟹居住迹、弹涂鱼的爬迹和鸟足迹等。下面重点介绍蝼蛄的居住迹。
蝼蛄的活动易受土壤湿度及食物的影响。它在近岸边滩表层比较松软的土壤中以咬食植物幼根和嫩茎为食, 所以其潜穴及穴道分枝点多分布在植物根旁。在底层层面上形成“ F” 形(图 4-A)、“ M” 形(图 4-B)、“ X” 状(图 4-C)、弯曲线形(图 4-D)或分枝网络状(图 4-E)等5种形态的潜穴, 潜穴直径2~6 cm, 潜穴规模0.3~3 m。潜穴内壁具平行锯齿状纹饰, 并且大小个体不同的蝼蛄潜穴交叉存在。土壤类型又极大地影响着蝼蛄的分布和密度。成年蝼蛄喜欢在轻度盐咸化及沿河两岸、灌渠两旁, 这些地方土壤土质松软, 保温保湿性能良好, 昼夜温度变化小, 环境比较稳定, 适合蝼蛄生活(赵月亮, 1974)。此外, 底层含水率的不同也会对蝼蛄潜穴的形态产生影响, 潮上带近岸边滩的蝼蛄潜穴, 顶层土壤会成胶结的瘤状结构(图 5-B); 而潮上带远岸草滩发现的潜穴顶层则孔隙较大, 多呈松散分离的块状(图 5-A); 且在远岸草滩的蝼蛄潜穴结构较简单, 而在近岸边滩发现的潜穴结构复杂、形态多样。在潮上带的近岸边滩的暂时性水渠的底层面上有弹涂鱼的爬迹, 该遗迹中间是单轴弯曲线型沟痕, 两侧为对称的呈羽状排列的小叶形沟痕(图 5-C)。
 | 图4 杭州湾庵东地区潮上带近岸边滩上蝼蛄造的不同形态遗迹Fig.4 Diverse burrows excavated by mole cricket in shoal located in supratidal zone near shoreline in Andong area of Hangzhou Bay |
 | 图5 杭州湾庵东地区潮上带不同含水量底层中的蝼蛄潜穴及暂时性水渠内弹涂鱼造的爬行迹Fig.5 Burrows of mole cricket on substrates with different water content and crawling traces of mudskipper generated in temporary channels located in supratidal zone near shoreline in Andong area of Hangzhou Bay |
潮间带的生物遗迹分异度及丰度较潮上带高, 尤其是退潮后, 形成地势平坦的小浅滩的生活环境, 生物门类多且个体分布密度大。除了与潮上带相似的生物遗迹外, 在高潮滩还发现了许多不同的造迹生物, 因这里的泥沼环境便于生物造穴, 所以造迹生物多在底层沉积物的浅层营穴居住。在高潮滩的积水泥沼环境中, 地表覆盖有大量喜盐性植物, 造迹生物在这里生活可以敏捷、快速地活动及进行底表觅食, 当遇到外界刺激时也可以迅速地躲藏。主要造迹生物包括珠带拟蟹守螺Cerithidea cingulata(Gmelin)(汇螺科)、泥螺Bullacta exarata(Philippi)(头楯目阿地螺科)、弹涂鱼Periophthalmua cantonensis(Osbeck)(弹涂鱼科鲈形目)、宁波泥蟹和少量的蝼蛄等。潮间带造迹生物遗迹多以运动迹为主, 运动迹的造迹者以珠带拟蟹守螺、泥螺、弹涂鱼为主, 尤其以弹涂鱼的丰度最高, 造迹形态上相差明显, 以拖迹数量较多; 此外还发现有觅食迹、停息迹、逃逸迹和生殖迹等, 该处因受杭州湾半日潮影响较强, 表层生物遗迹易发生变形或被冲刷, 不易保存。
4.2.1 高潮滩的积水泥沼环境中的生物遗迹
1)弹涂鱼的爬迹。弹涂鱼栖息于近岸浅滩或底质烂泥的高潮区, 对恶劣环境的水质耐受力强, 是暖水性的广温、广盐性鱼类, 喜穴居。依靠胸鳍和尾柄在水面、泥滩、岩石上爬行或跳跃; 匍匐于泥涂上觅食底栖硅藻、蓝绿藻, 也食少量桡足类及有机质。退潮后, 弹涂鱼在泥滩上不停爬行、跳跃以进行觅食, 遇到外界刺激后, 会快速钻入底内穴道中, 在穴口附近常伸头观察周围情况(图 6-A)。其爬行的拖迹因底质的不同也会呈现不同的形态(图 6-B— 6-E), 如在较干的软底底质的环境下, 其胸鳍在底质上呈现两边对称的蹼状(图 6-E), 两蹼中间尾柄留下较浅的拖迹; 而在较湿的软底底质下两蹼中间则出现尾柄留下的犁沟状爬迹(图 6-C, 6-D)。居住迹为“ Y” 型, 由上部的“ U” 形穴道和下部的直形穴道组成; 两穴口往往稍高出底表面, 呈锥形丘, 是动物挖穴道时抛出来的沉积物堆积而形成的, 穴道直径不等, 穴口径较小, “ U” 形底横穴道及单行垂直穴道径粗, 穴壁光滑, 具氧化层(张守忠, 2003)。洞穴孔道的深浅和长度依底质的性质而异, 软泥质较深长, 可达50~70 cm, 孔穴一般是独占性的。这些遗迹多出现于高、中浅滩的砂泥滩或泥滩氧化层。此外, 当弹涂鱼遇到外界刺激需要躲避时, 通常也会寄居在螃蟹的潜穴中。
 | 图6 杭州湾庵东地区高潮滩积水泥沼环境中弹涂鱼造的不同形态遗迹Fig.6 Different morphologic traces crawled by mudskipper in a waterlogging quagmire environment of the upper intertidal zone in Andong area of Hangzhou Bay |
2)宁波泥蟹的觅食迹及居住迹。宁波泥蟹体态近长椭圆形, 在潮间带泥滩上分布较多, 其居住迹的穴道多为圆柱状, 伸入底层呈“ J” 形(图 7-C), 穴径的上端较细, 直径为1.5 cm, 中部以下稍粗, 直径为2.5~3 cm, 主穴道的穴口高出底面(图 7-B), 位于圆筒状泥墙顶部, 并在泥墙脚周围筑墙时留下爪痕(图 7-A); 另一类在近底表段分枝, 分枝潜穴具有3个穴口, 形成“ Y” 形(图 7-D)和“ W” 形潜穴; 第3类为直— 曲形, 上端呈半“ U” 形, 下端呈“ J” 形(图 7-E), 总深度为32 cm, 上口径较细, 直径为1 cm, 向下渐宽, 在拐弯处垂直向下呈“ J” 形向下延伸, 半U型拐弯处直径为3 cm, 直至底部较粗。主穴道穴口一般呈圆形, 与底表平齐。在退潮后, 宁波泥蟹在底表从泥砂中摄取有机质, 具群聚性, 在低洼水边处, 边运动边觅食, 遇到外界刺激后会迅速潜入潜穴。泥蟹在退潮后的较长时间里会藏在潜穴内, 以躲避干旱环境。穴口周围有近圆形的食渣泥球及爪痕, 分布分散, 且发现有洞口呈花瓣状放射状的爬迹(图 7-F)。由于中潮区底质多样, 较平坦、含水量大、有机质丰富, 且环境稳定, 适宜于蟹类生长, 其多分布在潮间带中潮坪的砂质泥滩层中。
 | 图7 杭州湾庵东地区高潮滩积水泥沼环境下螃蟹造的不同形态遗迹Fig.7 Various traces generated by crabs in a waterlogging quagmire environment of the upper intertidal zone in Andong area of Hangzhou Bay |
3)泥螺的拖迹。泥螺是典型的潮间带底栖匍匐动物, 多栖息在潮间带的中低浅滩, 泥砂或砂泥的浅滩上, 以中潮带下缘至低潮带上缘最多, 退潮后在浅滩表面爬行, 在阴雨或天气较冷时, 潜于泥砂表层1~3 cm处, 不易被人发现, 以底栖藻类、有机碎屑、无脊椎动物的卵、幼体和小型甲壳类等为食。泥螺行动缓慢, 它用头盘掘起泥砂与身体分泌的黏液混合, 包被在身体表面, 像是一堆凸起的泥沙, 起着拟态保护作用。其运动时会在壳体两侧形成2条平行的脊状凸起, 其拖迹会因底质含水量的不同呈现出不同的形态, 如在富水底层中, 螺体常形成平缓不分枝的带状拖迹(图 8-A), 拖迹的宽度约为0.5 cm。而在弱固结的底质中, 其拖迹呈3条平行的条带状(图 8-B), 中间条带宽约0.2 cm, 两侧条带宽约0.4 cm。类似于珠带拟蟹守螺的犁沟状拖迹。
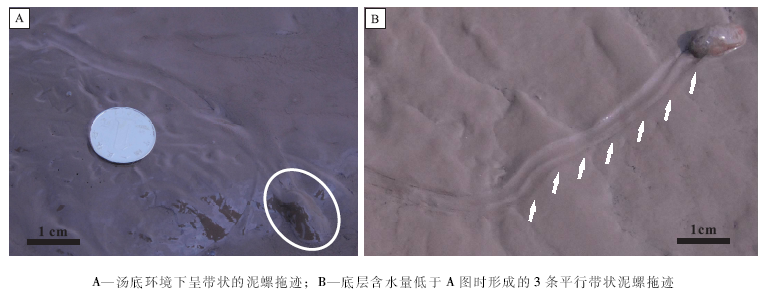 | 图8 杭州湾庵东地区潮间带高潮区不同底质上的泥螺拖迹Fig.8 Different trails produced by mud snails in different substrates of the upper intertidal zone in Andong area of Hangzhou Bay |
4.2.2 中、低浅滩粉砂质沉积环境中的生物遗迹
中、低浅滩的粉砂质沉积环境中以珠带拟蟹守螺的丰度最高, 在滩面上形成大量的运动拖迹、停息迹和生殖迹等。珠带拟蟹守螺在底表沉积物中缓慢匍匐爬行来进行觅食活动。退潮后, 它们多聚集在藻类堆积或低洼积水处, 呈群聚状态。滩面数量分布较多, 主要分布在潮间带中的泥砂或砂泥滩中。发现有运动迹、停息迹和生殖迹等3种较明显的生物遗迹。运动迹呈弧形(图 9-A)或弯曲形(图9-B), 宽度为0.4~0.8 cm, 是该生物在滩面上匍匐运动时拖拉着贝壳在底表沉积物上擦划而形成的漫移拖迹。拖迹的形态呈弯、直、交叉穿切和环形圈状(图 9-C)等, 随壳体运动方向变化而变化, 运动时两侧沙脊微隆起, 中部微凹、属表迹沟痕遗迹。壳尾在两侧沙脊中间, 形成犁沟状。
 | 图9 杭州湾庵东地区潮间带中— 低潮区的珠带拟蟹守螺造的不同生物遗迹Fig.9 Different trails and resting traces produced by Cerithideopsilla cingulate of the middle-lower intertidal zone in Andong area of Hangzhou Bay |
退潮后, 因生物暴露于外部时间长或周边环境发生急剧变化, 螺体的前部分或整体会暂时潜入底质中 (图 9-D), 而形成囊状凹陷的表生半浮式停息迹, 或与运动迹相伴随, 从轨迹形态上似串珠状 (图 9-E)。生殖迹为近似“ C” 形或圆形的泥质圆条状卵群 (图 9-F), 粗细均匀, 宽度约为0.1 cm, 长度为0.3~0.5 cm, 属表生遗迹(表迹)。在繁殖季节, 通常以壳顶为中心, 按逆时针方向边运动边产卵, 卵与分泌的黏液全部从体前端右侧排出, 并与底表沉积物粘结在一起, 形成泥条状的卵群。
生态环境影响着造迹生物行为习性及潜穴内部微观结构形态。潮间带退潮后, 地势平坦, 生存环境较稳定, 造迹生物丰富、体态偏小, 多以食泥性类为主。潮汐的规律运动塑造了潮间带环境的3个分区及各种微环境, 影响着造迹生物的分布, 形成以底质表面觅食迹、底内居住迹为主体的遗迹群落(图 10)。底质类型则决定造迹者的群落结构特征, 也就是说, 同一潮区环境中, 底质性质不同会导致造迹生物群落的成分及造迹形态出现明显差异。如蝼蛄在含水量低的底质上丰度低, 且潜穴外壁为松散的块状结构; 而在含水量高的底质环境下分布较集中, 其潜穴外壁则呈胶结的瘤状构造。此外, 水动力和底质粒度也共同影响着生物遗迹的清晰度、改造程度和遗迹的形态变化等。
本次工作对浙江省庵东镇以北浅滩上现代生物遗迹的组成与分布特征进行了初步研究, 主要取得了以下几方面的新认识:
1)研究区浅滩环境中的造迹生物主要为蝼蛄、 泥甲虫、 宁波泥蟹、 珠带拟蟹守螺、泥螺、弹涂鱼、 蚂蚁和鸟类等, 营造的生物遗迹主要为觅食迹、 居住迹、 爬行迹(包括爬迹、拖迹和鸟足迹)等。
2)潮上带以节肢动物蝼蛄造的“ F” 形、“ M” 形、“ X” 形、弯曲线形和分枝网格状的觅食迹和居住迹最丰富, 常伴生有泥甲虫造的树枝分叉状居住迹、蚂蚁潜穴和鸟类足迹, 以及弹涂鱼造的羽状觅食迹和少量宁波泥蟹造的居住迹。
3)在潮间带高潮坪的积水泥沼环境中主要发育弹涂鱼造的多种形态的爬行迹、宁波泥蟹造的觅食迹及居住迹(“ J” 形、“ W” 形和“ Y” 形潜穴)和泥螺造的带状拖迹; 而中— 低潮区则以珠带拟蟹守螺造的弧形、弯曲形、圆圈状的运动迹、停息迹和呈“ C” 形条带状的季节性生殖迹为主。在潮间带高潮区和中— 低潮区, 生物遗迹的丰度和分异度比潮上带高。
4)造迹生物的带状分布具明显的环境指示意义。可根据特有的生物沉积构造来指示沉积速率; 不同的亚环境或微环境具有不同的生物遗迹群落, 从而建立了庵东浅滩现代沉积中生物遗迹分布模式。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|