{kind=link}
{kind=link}
{kind=link}
安徽巢湖下三叠统含巢湖龙动物群地层微相特征及古环境
[张钰莹1, 2  , 江大勇
, 江大勇1 , 付宛璐1 , 季承3 , 孙作玉1 ]
, 江大勇, 付宛璐|
|

通讯作者简介 江大勇,男,1969年生,北京大学地球与空间科学学院教授,主要从事古脊椎动物学和综合地层学研究。E-mail: djiang@pku.edu.cn。
第一作者简介 张钰莹,女,1987年生,北京大学地球与空间科学学院硕士,主要从事碳酸盐岩微相研究。E-mail: yycug@163.com。
安徽巢湖马家山剖面下三叠统南陵湖组中段上部—上段下部第 598— 630自然层中产出早三叠世巢湖龙动物群化石。通过对赋存巢湖龙动物群化石的第 598— 630自然层的微相类型和稳定碳氧同位素进行分析,揭示了该时期的沉积环境及其变化。在该段地层中共识别出 4种微相类型,分别是红色—灰色瘤状厚层含骨针—双壳类生屑泥晶灰岩、灰色含生屑泥质泥晶灰岩、青灰色纹层状泥质泥晶灰岩和浅灰色含泥质泥晶灰岩,反映当时沉积环境从早到晚由外缓坡脚—深海盆地边缘渐变为外缓坡环境。稳定碳氧同位素分析表明,该段地层中δ 13C值与 δ 18O 值普遍偏低。综合微相、沉积相及稳定碳氧同位素特征,推断巢湖龙动物群生活在安静的外缓坡环境中,伴生的生物种类及数量相对比较稀少,海水温度总体偏高。
About the corresponding author Jiang Dayong,born in 1969,is a professor at School of Earth and Space Sciences,Peking University. Now he is engaged in vertebrate palaeontology and integrated stratigraphy. E-mail: djiang@pku.edu.cn.
About the first author Zhang Yuying,born in 1987,is a master at School of Earth and Space Sciences,Peking University. Now she is engaged in carbonate microfacies. E-mail: yycug@163.com.
The Early Triassic Chaohusaurus fauna was found at No.598-630 layers at the top of the Middle Member-the bottom of the Upper Member of the Nanlinghu Formation in Majiashan section of Chaohu area,Anhui Province. Based on the microfacies characteristics and the carbon and oxygen isotope analysis,the sedimentary environment of the fossil beds where the Chaohusaurus fauna had occurred was studied. 4 microfacies types were recognized from the studied layers,which contained the red-grey thick nodular limestone with spicules-bivalves,the gray bioclastic-argillaceous micrite limestone,the gray laminated argillaceous micrite limestone,and the light gray micrite limestone,respectively. It was indicated that the sedimentary environment of the fossil beds was from the foot of the outer ramp-abyssal basin to the outer ramp. The values of the δ13C and δ18O are generally low in the whole of fossil beds. Combined with the above characteristics,it might be inferred that the Chaohusaurus fauna was living in the outer ramp zone with low-energy,less numbers and types of the biocoenose,and high seawater temperature.
巢湖龙动物群中含有目前已知的地层层位最低的、时代最老的鱼龙化石之一:龟山巢湖龙(Chaohusaurus geishanensis Young and Dong, 1972)(Young and Dong, 1972; Motani and You, 1998), 并伴生有鱼类、菊石和瓣鳃类等化石(Tong and Zakharov, 2004; Tong et al., 2004, 2006), 其产出于上三叠统奥伦尼克阶斯帕斯亚阶的南陵湖组上段, 岩性主要为灰色薄层灰岩与泥灰岩互层, 属牙形石Neospathodus triangularis带和菊石Subcolombites带(陈烈祖, 1985; Tong and Yin, 2002; Tong and Zakharov, 2004; Tong et al., 2004, 2006; 童金南等, 2005a, 2005b)。
前人对于安徽巢湖下三叠统研究非常系统详细(左景勋等, 1995, 2003, 2006, 2013; 童金南, 1997; 李双应, 2005; 童金南和殷鸿福, 2005a, 2005b, 2009; Li et al., 2007; Jiang et al., 2009; Tong and Zhao, 2011; 浦万财等, 2012; 罗锐和张利伟, 2013), 但含巢湖龙动物群化石层段的沉积环境背景研究则比较少, 尤其是斯帕斯亚期古环境的精细研究不足。因此, 安徽巢湖早三叠世巢湖龙动物群产出地层层段微相特征及古环境背景是文中的研究重点。马家山剖面位于安徽省巢湖市西北郊(图 1), 距市区约2 km。作者在对该剖面系统进行的野外地层观察、描述、测量和采样的基础上, 对磨制的岩石薄片进行了观察、描述、鉴定和微相分析。通过对碳酸盐岩的颗粒类型及特征、微相类型、镜下化石组合及其变化进行分析, 并结合碳氧同位素特征, 初步探讨了巢湖龙动物群化石产出层段地层所反映的沉积环境背景。
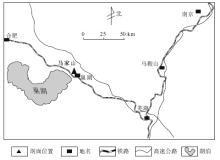
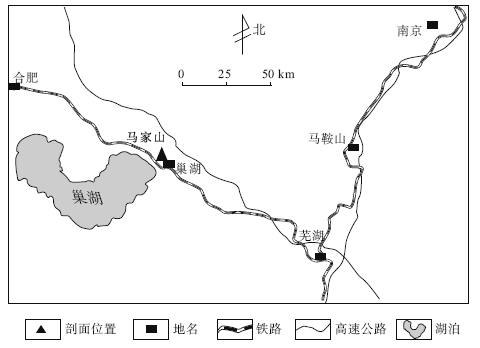 | 图1 安徽巢湖含早三叠世巢湖龙动物群的马家山剖面位置Fig.1 Location of Majiashan section with the Early Triassic Chaohusaurus fauna in Chaohu area, Anhui Province |
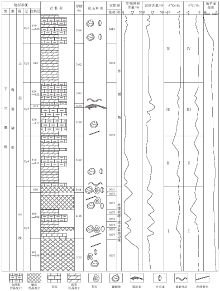
南陵湖组位于下三叠统奥伦尼克阶斯帕斯亚阶(童金南, 1997)。巢湖马家山剖面出露南陵湖组上段及中段上部第598— 1000自然层, 文中主要分析含巢湖龙动物群化石的南陵湖组中段上部第598— 610自然层及南陵湖组上段下部第611— 630自然层(图 2)。研究区南陵湖组中段上部地层主要为瘤状灰岩与泥灰岩互层, 产出假色尔特菊石Pseudoceffifes sp., 克氏蛤Claraia sp., 海浪蛤Posidonia sp., 正海扇Eumorphofis sp.及牙形类化石; 南陵湖组上段底部地层主要为泥质灰岩与泥灰岩互层, 产出亚哥伦布菊石Subcolumbifes sp.、王氏克氏蛤Claraia wangi(Paffe)、海浪蛤、正海扇及牙形类化石(Tong and Yin, 2002; Tong and Zakharov, 2004; Tong et al., 2004, 2006; 童金南等, 2005a, 2005b; Tong and Zhao, 2011), 并报道有出现层位最低的中生代海生脊椎动物化石巢湖龙Chaohusaurus等(Young and Dong, 1972; 陈烈祖, 1985)。马家山剖面第598— 630自然层自上而下的岩性特征描述如下(Cy代表地层分层):
上覆地层:下三叠统奥伦尼克阶斯帕斯亚阶南陵湖组上段中部
整合
下三叠统奥伦尼克阶斯帕斯亚阶南陵湖组上段下部
Cy1:自然层626— 630层, 灰色泥灰岩与中层泥质灰岩互层厚266 cm
630: 灰色泥灰岩, 含极少量双壳碎屑、钙球等 厚27 cm
629: 灰色中层泥质灰岩, 含少量生物碎屑厚34 cm
628: 灰色泥灰岩, 同630层相似 厚12 cm
627: 顶部深灰色泥质灰岩, 向下渐变为深灰色泥灰岩厚42 cm
626: 灰色含泥质灰岩和泥灰岩互层, 共11个旋回 厚151 cm
Cy2: 自然层625层, 土黄色泥灰岩 厚362 cm
625: 土黄色泥灰岩, 几乎不含生物化石 厚362 cm
Cy3: 自然层617— 624层, 灰色厚层— 巨厚层泥质灰岩与灰色泥灰岩互层 厚453 cm
624: 浅灰色巨厚层泥质灰岩, 表面有纹层, 泥质含量较低, 几乎不含生物化石 厚160 cm
623: 泥灰岩, 夹薄层灰岩, 含少量遗迹化石 厚4 cm
622: 上部20.5 cm为浅灰色中层泥质灰岩; 下部含2个旋回, 第1个旋回顶部1 cm泥灰岩, 之下6 cm灰色薄层泥质灰岩, 纹层发育; 第2个旋自上向下依次为1 cm泥灰岩, 8 cm灰色薄层泥质灰岩, 2 cm泥灰岩 厚39 cm
621: 灰色泥灰岩, 主要爬行动物产出层, 单层厚1.5 cm 厚30 cm
620: 灰色厚层泥质灰岩, 不含生物化石 厚93 cm
619: 灰色泥灰岩, 纹层状构造, 不含生物化石 厚12 cm
618: 灰色巨厚层泥质灰岩, 近底部见泥灰岩, 含少量破碎生物化石 厚100 cm
617: 泥灰岩, 含薄壳或海绵骨针碎屑 厚15 cm
Cy4: 自然层611— 616层, 中薄层泥质灰岩与黄— 灰色泥灰岩互层 厚522 cm
616: 灰色含泥质灰岩, 同618层 厚150 cm
615: 顶部灰色泥灰岩, 底部黄色泥灰岩, 中间夹薄层(1~2 cm)灰岩夹层 厚54 cm
614: 灰色中薄层泥质灰岩, 几乎不含生物化石 厚105 cm
613: 灰色泥灰岩, 含较多钙球, 少量瓣鳃碎屑化石 厚132 cm
612: 岩性渐变, 顶部20 cm为深灰色泥灰岩, 中部25 cm为泥灰岩, 底部15 cm为薄层灰岩 厚60 cm
611: 灰色中层含泥质灰岩, 含少量遗迹化石厚22 cm
整合
下三叠统奥伦尼克阶斯帕斯亚阶南陵湖组中段上部
Cy5: 自然层598— 610层, 瘤状灰岩与泥灰岩互层, 夹泥质灰岩与灰岩 厚833 cm
610: 黄色瘤状灰岩, 含少量钙球及双壳碎屑化石 厚54 cm
609: 青灰色泥灰岩, 不含生物化石 厚20 cm
608: 青灰色瘤状灰岩, 风化呈土黄色, 生物碎屑含量较大, 类型丰富, 主要为钙球, 瓣鳃碎屑及海绵骨针 厚132 cm
607: 青灰色泥灰岩, 不含生物化石 厚36 cm
606: 灰色泥质灰岩, 不含生物化石 厚45 cm
605: 青灰色泥灰岩, 不含生物化石 厚91 cm
604: 灰色瘤状灰岩, 含较多介形虫化石, 少量瓣鳃碎屑 厚32 cm
603: 深灰色泥灰岩, 不含生物化石 厚15 cm
602: 白灰色微薄层灰岩, 含较多丝状体化石, 定向排列 厚45 cm
601: 若干旋回, 每个旋回顶面为青灰色瘤状灰岩, 厚约10 cm, 内部有微薄层灰色泥灰岩与灰岩互层 厚40 cm
600: 顶部33 cm为灰色泥质瘤状灰岩, 风化呈暗肉红色, 底部为灰黑色泥灰岩, 含大量钙球化石, 少量介形虫及瓣鳃碎屑化石 厚52 cm
599: 灰色泥质瘤状灰岩, 含大量钙球化石, 少量介形虫及瓣鳃碎屑化石 厚61 cm
598: 红色厚层瘤状灰岩, 含大量瓣鳃碎屑、介形虫、钙球化石 厚200 cm
整合
下伏地层:下三叠统奥伦尼克阶斯帕斯亚阶南陵湖组中段中部
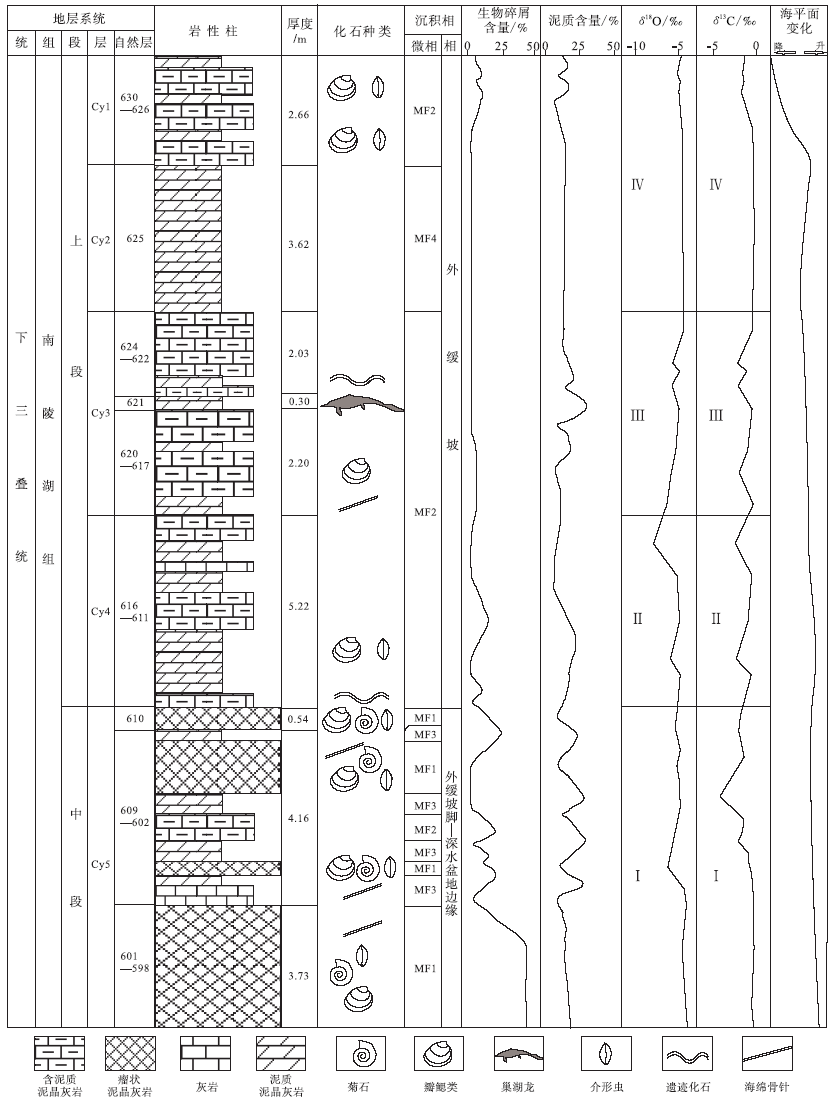 | 图2 安徽巢湖马家山剖面南陵湖组第598— 630自然层地层综合柱状图Fig.2 Intergrated stratigraphic column of No.598-630 layers of the Nanlinghu Formation in Majiashan section of Chaohu area, Anhui Province |
文中在对碳酸盐岩进行分类命名时采用了Dunham(1962)的分类方案。通过对岩石薄片中基质类型、颗粒类型及特征、化石特征等的观察和描述, 对安徽巢湖马家山剖面南陵湖组中段上部— 上段下部地层的微相类型(MF)进行了区分。马家山剖面南陵湖组中上段碳酸盐岩基质以泥晶为主, 生物碎屑颗粒含量较少, 与Flü gel(2004)总结的古生代/中生代碳酸盐岩缓坡的常见微相类型能够较好地对比, 从而确定马家山南陵湖组碳酸盐岩形成于缓坡相(Burchette and wright, 1992)。南陵湖组中段顶部— 上段底部第598— 630自然层的碳酸盐岩可划分为4种主要的微相类型。
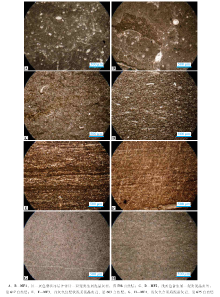
含多种生物化石碎屑, 保存较破碎, 含量15%~40%, 瘤状结构(图 3-A, 3-B)。生物碎屑类型多样:双壳类壳体极薄, 破碎程度高, 含量5%~15%; 海绵骨针长0.2~0.5 mm, 非定向排列; 钙球呈圆球状, 内部亮晶胶结物呈波状消光, 直径0.1~0.3 mm, 无法判断内部结构, 含量10%~25%; 介形虫呈椭圆至圆球状, 直径0.2~0.3 mm。此微相类型与标准微相RMF3(有多种生物的生屑颗粒质灰泥石灰岩)相似(Flü gel, 2004), 代表外缓坡底部— 盆地边缘的沉积环境(郭福生, 1989; 朱洪发和王恕一, 1992; 董兆雄和朱晓惠, 2002)(其地层分布见图2)。
 | 图3 安徽巢湖马家山剖面南陵湖组第598— 630自然层中碳酸盐岩微相类型Fig.3 Microfacies types of No.598-630 layers of the Nanlinghu Formation in Majiashan section of Chaohu area, Anhui Province |
含有少量遗迹化石, 生物化石数量较少(图 3-C, 3-D), 种类与MF1类似, 主要为瓣鳃碎屑、海绵骨针、介形虫等, 但含量相对上一种微相类型比较小, 含量为5%~15%。瓣鳃碎屑保存较差, 多为薄质双壳类, 长0.2~0.5 mm, 含量5%~10%; 海绵骨针保存极差, 弱定向排列, 含量约为5%; 介形虫呈圆球状, 亮晶胶结, 直径0.1~0.2 mm, 含量约为5%。此微相岩石中泥质含量为10%~20%, 分布不均, 曾受到生物扰动。此微相类型与标准微相RMF2(泥质经过掘穴生屑颗粒质泥灰石灰岩)相似(Flü gel, 2004), 在外缓坡常见, 代表较深水的盆地和开阔海陆棚以及深水陆棚的坡脚前端位置(方邺森等, 1987)(其地层分布见图2)。
几乎不含生物碎屑, 由明暗相间的微米级细纹层形成水平纹层, 泥质含量极高(图 3-E, 3-F)。此微相类型与标准微相RMF5(远洋灰泥石灰岩)(Flü gel, 2004)相似, 代表外缓坡底部— 盆地边缘、深水陆棚相沉积(方邺森等, 1987)(其地层分布见图2)。
与MF2相比生物碎屑含量极低, 与MF3相比泥质含量更低(图 3-G, 3-H)。此微相与RMF4(似球粒颗粒质灰泥石灰岩)(Flü gel, 2004)相似, 在外缓坡斜坡脚常见, 代表较深水的盆地和开阔海陆棚以及深水陆棚的坡脚前端位置(其地层分布见图2)。
前人研究(Tong and Yin, 2002; 童金南和殷鸿福, 2005, 2009; Li et al., 2007)认为, 早三叠世下扬子区是一个向北倾斜的碳酸盐岩缓坡台地, 巢湖位于该台地北侧较深水区域, 南陵湖组中上部碳酸盐岩组分以泥晶为主, 含较深水相的菊石、双壳类化石, 沉积环境为安静的盆地边缘— 潮下带斜坡环境。
根据前述微相特征、泥质含量变化情况和野外观察的宏观特征, 结合前人研究成果, 对照Flü gel(2004)的碳酸盐岩缓坡相带模式, 将安徽巢湖马家山剖面南陵湖组第598— 630自然层分为2种沉积相类型。
该沉积相包括MF1、MF2和MF3, 主要是含生物碎屑颗粒瘤状灰岩与纹层状泥灰岩互层, 泥质含量较多, 颗粒主要为生物碎屑, 含丰富的薄壳远洋双壳类、介形虫、钙球、丝状体、海绵骨针等, 非定向排列。为典型的深海盆地边缘沉积环境。
该沉积相包括MF2和MF4, 下部为中— 薄层泥质灰岩与泥灰岩互层; 中部变为厚— 巨厚层泥质灰岩与泥灰岩互层; 上部为中层泥质灰岩与泥灰岩互层。此段地层生物碎屑含量较低, 主要为破碎的双壳类、介形虫等, 部分弱定向排列, 泥质含量总体向上变小(泥质含量变化见图2)。为典型的外缓坡沉积环境, 海水深度向上变浅。
综上, 巢湖马家山剖面南陵湖组中段— 上段第598— 630自然层碳酸盐岩总体沉积特征是生物碎屑种类较少, 以泥晶支撑为主, 代表相对低能的深海外缓坡沉积环境。所识别的2种碳酸盐岩沉积相类型在纵向上形成了向上变浅相序, 即由盆地边缘沉积相向上渐变为外缓坡沉积相。
作者系统采集了研究层段每一自然层的测试样品, 共33块, 室内挑选无后期蚀变、无方解石脉充填、无破碎现象的新鲜岩石样品, 由中国科学院南京地质古生物研究所国家重点实验室同位素质谱仪完成测试, 参比标准:GBW-04405, 分析精度:δ 13C(PDB)和δ 18O(PDB)测定值标准偏差小于0.030及0.080。
研究区下三叠统碳酸盐δ 13C值变化于-4.296‰ ~-0.342‰ 之间, 平均值为-1.338‰ , 变化幅度为3.954‰ 。碳同位素变化大致分为4个阶段(图 2), 与4个沉积层段相对应:第Ⅰ 阶段对应南陵湖组中段第598— 610自然层, 碳同位素总体向负漂移, 在第607层达到最低值, 然后向正漂移; 第Ⅱ 阶段对应南陵湖组上段第611— 616自然层, δ 13C值有所波动, 总体呈负漂趋势; 第Ⅲ 阶段对应南陵湖组上段第617— 624自然层, δ 13C值呈正漂趋势; 第Ⅳ 阶段对应第625— 630自然层, δ 13C值变化较小, 总体平稳, 略有负漂趋势。碳酸盐岩中无机碳同位素受多种因素影响, 众多研究结果(Kump, 1991; 王伟等, 1997; 曹长群等, 2002; 左景勋等, 2003, 2006, 2013; Payne et al., 2004)表明, 碳酸盐δ 13C值反映地史时期的生物初级生产力水平, 与生物总量成正比。马家山剖面碳同位素均为负值, 有小幅度波动, 反映了该段在P/T生物大灭绝之后生态系统刚刚重建, 生物稀少。
研究区 δ 18O 值变化范围为-8.884‰ ~-5.023‰ , 平均值为-6.142‰ , 变化幅度为3.861‰ 。δ 18O值变化趋势与δ 13C值变化类似, 也分为4个阶段(图 2), 与4个沉积层段相对应:第Ⅰ 阶段(第598— 610自然层)氧同位素总体向负漂移, 在第604自然层达到最低值, 然后向正漂移; 第Ⅱ 阶段(第611— 616自然层)δ 18O值有所波动, 总体呈负漂趋势; 第Ⅲ 阶段(第617— 624自然层)δ 18O值呈正漂趋势; 第Ⅳ 阶段(第625— 630自然层)δ 18O值变化较小, 总体平稳, 略有负漂趋势。当碳酸盐与介质处于平衡状态时, δ 18O值随温度的升高而下降(张秀莲, 1985; 邵龙义等, 1996)。但是由于 δ 18O 值测定古海水温度的方法受到很多因素影响, 因此文中不利用氧同位素数据定量计算古海水温度, 只分析海水温度变化趋势及大致温度范围。Sun 等(2012)利用牙形石的稳定氧同位素数据恢复出早三叠世华南地区古海水温度, 指出在斯密斯亚期末期海水温度高达40 ℃, 导致了海洋生物的灭绝并阻碍了海洋生物圈的复苏。研究区的 δ 18O 值普遍偏低, 说明斯帕斯亚期延续了斯密斯亚期末期海水的高温度, 在斯帕斯亚期早期海水温度缓慢上升, 而在南陵湖组上段底部对应的斯帕斯亚期中晚期之交可能达到最高温度后缓慢下降, 由此可以判断出巢湖龙动物群可能生活在海水温度偏高的环境中。
1)在安徽巢湖马家山剖面南陵湖组含早三叠世巢湖龙动物群化石的第598— 630自然层中共识别出4种微相类型:红色— 灰色瘤状厚层含骨针— 双壳类生屑泥晶灰岩、灰色含生屑泥质泥晶灰岩、青灰色纹层状泥质泥晶灰岩、浅灰色含泥质泥晶灰岩。该段地层可分为2种沉积相类型, 自下而上为外缓坡脚— 深海盆地边缘和外缓坡。
2)安徽巢湖马家山剖面整段化石层段的δ 13C值均为负值, 说明早三叠世海洋生物稀少。同时, δ 18O值普遍偏低, 很可能说明斯帕斯亚期延续了斯密斯亚期末期海水的高温度, 其中, 斯帕斯亚期早期温度呈现缓慢上升, 而在南陵湖组上段底部对应的斯帕斯亚期中晚期之交达到最高温度后, 海水温度缓慢下降。
3)综合巢湖地区马家山剖面南陵湖组第598— 630自然层沉积微相、沉积相及稳定碳氧同位素曲线特征, 认为巢湖龙动物群生活在安静的外缓坡、且海水温度总体偏高的环境中。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|