
{kind=link}
{kind=link}
{kind=link}
{kind=link}
志留纪—早泥盆世维管植物的系统发育、幕式演化和地理分布:植物大化石证据
[薛进庄 , 郝守刚]
, 郝守刚]
, 郝守刚]
|
|

第一作者简介 薛进庄,男,1981年生,北京大学地球与空间科学学院副教授,主要从事早期维管植物演化的相关研究。E-mail: pkuxue@pku.edu.cn。
尽管近年来对志留纪—早泥盆世陆生维管植物系统发育的认识取得了很大进展,但是目前对多囊蕨类、莱尼蕨类、工蕨类的界定和类群划分尚有分歧,工蕨类和石松类的姐妹群关系仍有疑问,真叶植物基部类群有待深入研究。志留纪至早泥盆世的维管植物演化可划分为 2幕,即前布拉格期序幕阶段和布拉格期爆发阶段。维管植物主要分支在第 1幕均处于萌芽状态,仅出现个别先驱型分子;第 2幕为维管植物分支爆发式演化的主幕。在布拉格期,规则叶序、孢子叶、似叶球状结构等石松类的典型性状最早显现,真叶植物发生明显的分支分化,大型叶及其同源器官(枝叶复合体)多次独立起源,孢子囊和叶性结构的发育相关性在多个支系中趋同演化出现。古植物地理方面,在东北冈瓦纳区的华南地区,工蕨类以孢子囊螺旋排列的类型为主,真叶植物在布拉格期显示出高度的多样性和分异度;在劳俄区,工蕨类中孢子囊成列排列的类型较为丰富,真叶植物的大量出现要晚于华南地区。羊角蕨纲、拟库克逊蕨类、莱尼蕨目的分支演化可能主要发生在南劳俄和西北冈瓦纳等中、高纬度地区。工蕨类、石松类和真叶植物的分支演化和地理辐射可能遵循“走出热带”模式,它们在低纬度地区最早发生分化并快速辐射;维管植物主要分支在布拉格期的爆发式演化,主要是热带植物群革新演化的贡献。
About the first author Xue Jinzhuang,born in 1981,is an associate professor of Peking University. His current research focuses on the evolution of early vascular plants. E-mail: pkuxue@pku.edu.cn.
Much progress has been made in the understanding of the phylogeny of Silurian-Early Devonian vascular land plants during the past years. However,no consensus has been reached considering the delimitation and subdivision of polysporangiophytes,rhyniopsids,and zosterophyllopsids;the sister-group relationship between zosterophyllopsids and lycopsids is questioned;detailed studies of basal euphyllophytes are needed. The evolutionary history of the Silurian-Early Devonian vascular plants can be divided into two phases:The pre-Pragian prelude phase and the Early Devonian Pragian explosion(burst)phase. The first phase is characterized by initial emergences of representatives of vascular plant major clades. The second is the main phase which witnessed explosive radiation of vascular plant clades. During the Pragian,typical features of lycopsids such as regular phyllotaxy,sporophylls,and strobilus-like structures appeared;the diversification of euphyllophyte lineages occurred;megaphylls and their homologous structures(branch-leaf complexes)evolved multiple times independently;and developmental interrelationship between sporangia and leafy structures convergently established in different lineages. In South China of the northeastern Gondwanan phytogeographic unit,zosterophyllopsids were dominated by those with spirally arranged sporangia and euphyllophytes showed high diversity and disparity during the Pragian,while in the Laurussian phytogeographic unit,zosterophyllopsids with rowed sporangia were abundant and the appearance of comparable euphyllophytes occurred later than in South China. The cladogenesis of horneophytopsids,cooksonioids,and rhynialeans might have mainly occurred at middle and high paleolatitudes such as southern Laurussia and northwestern Gondwana. The cladogenesis and biogeographic radiation of zosterophyllopsids,lycopsids,and euphyllophytes may accord with the “out of the tropics”model,that is,these groups diversified and rapidly radiated earlier at low paleolatitudes;the explosion of vascular plant clades in Pragian was mainly contributed by the evolutionary novelty of tropical floras.
志留纪— 泥盆纪维管植物在陆地上的起源和演化分异是地球生命史中的重要事件, 对地球环境和陆地生态系统产生了深远的影响。期间, 古老的维管植物类群如莱尼蕨类、工蕨类、三枝蕨类等, 以及现代类群如石松类、楔叶类、真蕨类及种子植物的先驱分子开始出现, 初步形成复杂的、多样化的生态系统; 植物的属种多样性急剧倍增(Knoll et al., 1984; Wang et al., 2010; Xiong et al., 2013), 一些关键革新性状如根、叶、次生木质部、异孢、乔木状习性和种子结构等快速演化出现(Bateman et al., 1998; 郝守刚等, 2002; Gerrienne et al., 2004, 2011; Hao et al., 2010; Hao and Xue, 2013a, 2013b)。因此, 志留纪— 泥盆纪的植物化石对于重建维管植物系统发育树、探讨维管植物重要类群的起源和早期演化、多样性的建立起着至关重要的作用(Kenrick and Crane, 1997; Crane et al., 2004; Hao and Xue, 2013a)。一方面, 化石可以用于标定现代植物类群起源和分化的时间序列, 揭示植物革新性状获得的模式和过程; 另一方面, 一些独特的植物类群仅在化石记录中可见, 如泥盆纪的工蕨类、前裸子植物、种子蕨等, 它们提供了仅通过研究现生植物无法认识到的生物多样性。
自Dawson(1859)、Kidston和Lang(1917)等早期的开创性工作以来, 百余年来对泥盆纪植物化石的研究主要集中在劳俄古陆(西欧和北美地区)。依据这些地区丰富的化石材料, 人们揭示了诸多早期维管植物的宏观演化规律(Knoll et al., 1984; Edwards and Davies, 1990; Kenrick and Crane, 1997)。这些研究无疑对世界其他地区同期植物群的研究提供了可供对比的参考依据; 但仅依据单一古陆的材料所揭示的模式, 对于认识全球植物的演化过程, 显然是不充足的(郝守刚等, 2002)。近几十年来, 对华南、澳大利亚、南美、中国西北准噶尔等地区的志留纪— 泥盆纪维管植物群的研究, 极大地补充和丰富了人们对早期维管植物演化分异的认识。特别是过去几十年的系统研究, 表明华南地区是早期维管植物辐射演化的中心之一(徐仁, 1966; 李星学和蔡重阳, 1977; 李承森, 1982; Li, 1992; 蔡重阳和王怿, 1995; Hao and Gensel, 1998; 郝守刚和王德明, 2003; 杨楠和李承森, 2009; Hao and Xue, 2013a; 王德明和熊聪慧, 2014)。对一系列保存精美、类型丰富的化石植物群如早泥盆世坡松冲植物群、中泥盆世西冲(海口)植物群、晚泥盆世五通植物群等的详细研究, 不仅揭示了华南板块上维管植物及植被的演化历史, 也为重建维管植物的系统发育树、揭示早期植物在不同大陆之间的地理辐射提供了丰富的资料(Hao and Gensel, 2001; 王怿等, 2006)。
以往的研究已对志留纪— 泥盆纪植物群的化石产地、古地理分布、生物地层学等多个方面进行了详细的总结和论述, 因此无需赘述。本文旨在总结近年来对志留纪— 早泥盆世维管植物及其内部类群的系统发育、分支演化、多样性演化与地理辐射方面取得的最新研究进展, 对散见于各类文献的论述做出梳理, 进而总结目前研究中的难点及关键问题之所在, 以期促进相关研究的深入。
本文所涉及到的年代地层名称、绝对年龄值均采用国际年代地层表2013年中文、 英文版(http://www.stratigraphy.org/index.php/ics-chart-timescale)。
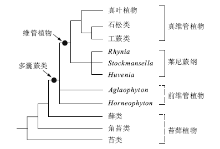
Kenrick和Crane(1991)认为Cooksonia、Nothia和Aglaophyton不具真正的管胞结构(即抗腐蚀的、木质化壁的输水细胞, 具环纹、螺纹、网纹等次生加厚), 所以属“ 前维管植物级别” (protracheophyte grade), 它们与维管植物共同构成一个分支— — 多囊蕨类(Polysporangiophytes), 以分枝的、具多个孢子囊的孢子体为特征。Kenrick和Crane(1997)的分析将Aglaophyton和Horneophytopsida(羊角蕨纲)置于多囊蕨类的基部位置(图 1), 认为它们属于前维管植物, 而不是真正的维管植物。但是, 上述几个属的归属问题、以及究竟哪些植物属于非维管多囊蕨类(non-vascular polysporangiophytes)仍是不确定的。Cooksonia属下的一些种, 已被证明具有管胞, 如:Cooksonia pertoni具有C-型管胞, 与常见的环纹和螺纹管胞类似, 但是其侧壁不穿孔、成分未知(Edwards, 2003); Aglaophyton的输水细胞壁虽然没有次生加厚的直接证据, 但是细胞壁表面呈现出一些网状的、或规则六边形的形式, 表明可能具有次生加厚(Edwards, 1986)。Kerp等(2001)认为Nothia的输水细胞为纺锤形、具有加厚壁和渐尖的顶端, 与管胞似有不同。Kenrick和Crane的(1997)的分支分析根据Nothia具有侧生孢子囊的特点, 将该植物置于工蕨类之中, 否定了他们1991年认为Nothia属于“ 前维管植物” 的观点。Kenrick和Crane(1997)认为羊角蕨纲的代表植物Horneophyton的输水细胞是光滑壁的, 没有次生加厚, 因此不是管胞。但是Edwards(2004)认为, 在Kidston和Lang(1920)的原始描述中, 已显示了Horneophyton的管胞具有螺纹或不规则的加厚, 尽管在腐蚀的样品中, 其管胞壁是光滑的。Aglaophyton、Horneophyton和Nothia这3种植物仅发现于莱尼燧石层中(布拉格期— 最早埃姆斯期), 浸解腐蚀以及硅化等埋藏学过程可能已改变了其输水细胞的细节特征(Edwards, 2004)。另2种被归入羊角蕨纲的植物Caia和Tortilicaulis, 其解剖结构未知。因此, Kenrick和Crane的(1991, 1997)提出的“ 前维管植物” 是否真实存在?它们的输水细胞是否与真正的管胞类似、或在一些微细结构上是否有同源性?这些问题仍有很多争议。另外, Aglaophyton、Horneophyton和Nothia这3种植物的时代为布拉格期— 最早埃姆斯期, 远晚于真维管植物的最早化石记录(志留纪温洛克世— 罗德洛世), 在时序上似乎也于“ 前维管植物” 这一演化级别不符。由于“ 前维管植物” 的不确定性, 多囊蕨类这一分支的概念还有待更深入的研究。
 | 图1 多囊蕨类、前维管植物、维管植物的系统发育关系(综合Kenrick和Crane, 1991, 1997; Kenrick, 2000; 但见文中讨论)Fig.1 Phylogenetic relationships of polysporangiophytes, protracheophytes and tracheophytes(summarized from Kenrick and Crane, 1991, 1997; Kenrick, 2000) |
Banks(1968, 1975)通过比较形态学研究, 将繁盛于泥盆纪早期的、传统意义上的裸蕨类(psilophytes)划分为3个类群:莱尼蕨亚门(Rhyniophytina)、工蕨亚门(Zosterophyllophytina)和三枝蕨亚门(Trimerophytina)。但是随着资料的积累, 莱尼蕨亚门、三枝蕨亚门已被表明不是单系类群, 工蕨亚门的范畴和界定也存有疑问(Kenrick and Crane, 1997; Hao and Xue, 2013a)。传统上, 通常认为莱尼蕨类具二分叉的茎轴、顶生孢子囊、具三缝孢, 包括Rhynia、Aglaophyton、Horneophyton、Cooksonia等植物, 但不同作者的分类系统中, 所包含的植物属种不尽相同(参见Kenrick and Crane, 1997的Table 4.2)。维管组织信息不明、形态上也具二分叉茎轴、顶生孢子囊的植物, 被称为拟莱尼蕨类(rhyniophytoids; Edwards and Edwards, 1986)。
Kenrick和Crane(1997)的分支分析认为应该摈弃传统分类学所定义的莱尼蕨亚门; 他们识别出一个由Rhynia、Stockmansella和Huvenia构成的单系类群, 并以之为代表, 定义了一个新的“ 莱尼蕨纲” (Rhyniopsida)(图 1)。该纲的特征包括:Rhynia-型分枝; 孢子囊基部具离层; 孢子囊以囊托着生; 独特的S-型管胞。该“ 莱尼蕨纲” 与真维管植物(Eutracheophytes)互为姐妹群, 共同构成维管植物支系(Tracheophytes, vascular plants)。Gerrienne等(2006)建议将Kenrick和Crane(1997)的“ 莱尼蕨纲” 改名为“ 副维管植物门” (Paratracheophyta), 这有利于避免混乱。“ 副维管植物门” 中, 除了上述3个属之外, 还包括配子体Remyophyton和Sciadophyton, 以及形态属Sennicaulis、Taeniocrada的一些种(Gerrienne et al., 2006)。
Cooksonia曾被作为莱尼蕨类的一个重要分子, 但长期以来, 该属的定义并不严格, 属下种类繁多, 形态变异复杂。Gonez和Gerrienne(2010a)对其属征进行了修订, 主要特征包括顶生孢子囊喇叭形, 孢子囊顶端平圆、由1层等径的细胞构成。模式种为Cooksonia pertoni。其他可归入的种包括C.hemisphaerica、C.cambrensis、C.bohemica、C.paranensis和C. banksii。早泥盆世洛赫考夫期的化石表明C. pertoni具C-型管胞(Edwards et al., 1992; Edwards, 2003), 类似于C.pertoni的孢子囊最早出现于志留纪温洛克世晚期地层中(Homerian期, 约430 Ma; Edwards and Feehan, 1980), 因此Cooksonia被认为是最早的真维管植物(Edwards, 2003; Gonez and Gerrienne, 2010a)。随后, Gonez和Gerrienne(2010b)以Cooksonia的另一种C. caledonica为模式, 建立了新属Aberlemnia和新组合Aberlemnia caledonica。Aberlemnia的孢子囊圆形、卵形或肾形, 孢子囊远端边缘具开裂带。
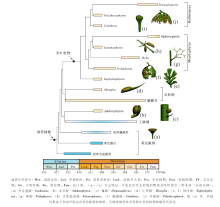
莱尼蕨类(Banks, 1975)形态特征简单, 植物体纤细, 大多保存不好, 因此可对比的性状较少, 使得对它们谱系关系的认识争议很大。Kenrick和Crane(1997)把Aglaophyton置入前维管植物等级, 但它与Rhynia非常相似(Edwards, 2004)。Kenrick和Crane(1997)、Gonez和Gerrienne(2010a)的分析认为Aberlemnia、Cooksonia与工蕨纲+石松纲的关系更近。Hao和Xue(2013a)的分支分析提出了不同的解释:Cooksonia、Aglaophyton、Rhynia、Aberlemnia相继以并系方式位于维管植物的基部位置, Aberlemnia与Renalia、Hsü a关系较近。由于莱尼蕨类谱系关系的复杂性, 目前很难形成一个严格基于谱系关系的分类系统(比如要求分类单元必须是单系的), 现有的分类都有一定的人为性。Hao和Xue(2013a)的分类中, 莱尼蕨类被作为一个纲, 下分莱尼蕨目(Rhyniales)、拟库克逊蕨类(cooksonioids)、拟肾囊蕨类(renalioids)和其他有疑问的植物(图 2)。莱尼蕨目以Rhynia和Aglaophyton为代表; 拟库克逊蕨类以Cooksonia pertoni和C.paranensis为代表; 而拟肾囊蕨类以Aberlemnia、Renalia、Hsü a为代表。未来的研究难点, 仍在于对上述植物性状特征的补充和完善。
 | 图2 志留纪— 早泥盆世维管植物代表类型及相关类群的系统发育关系(据Hao and Xue, 2013a, 有修改)Fig.2 Phylogenetic relationships among representative plants of the Silurian-Early Devonian and related groups(modified from Hao and Xue, 2013a) |
通常以侧生孢子囊等综合特征来界定工蕨类(工蕨亚门或纲)(Banks, 1975), 这一类植物分布广泛, 化石资料丰富, 目前至少包括20余属(Wang and Hao, 2002; Wang, 2007; Taylor et al., 2009; Hao and Xue, 2013a; Cascales-Miñ ana and Meyer-Berthaud, 2013)。工蕨类的化石记录漫长, 最早出现于加拿大极区Bathurst岛的志留纪罗德洛世(Ludlow)(Kotyk et al., 2002), 至早泥盆世布拉格期, 属种多样性达到高峰, 埃姆斯期之后明显衰减, 最晚记录可延续至晚泥盆世(Hueber and Banks, 1979; Cascales-Miñ ana and Meyer-Berthaud, 2013)。
尽管研究历史漫长, 资料积累较多, 但目前对工蕨类的界定、工蕨类内部类群的划分及系统关系仍有很大分歧(Gensel, 1992; Kenrick and Crane, 1997; Hao and Xue, 2013a)。Kenrick 和Crane(1997)的分析认为“ 工蕨纲” 的近裔共性为顶端拳卷、孢子囊两侧排列、横切面椭圆形的木质部束, “ 工蕨纲” 下分1个目、4个科, 其中包括了Hsü a、Barinophyton等分类位置有争议的植物。但是, Kenrick 和Crane(1997)将传统上认为属于工蕨纲的具螺旋排列孢子囊的一些植物— — 包括工蕨属的模式种米尔顿工蕨(Zosterophyllum myretonianum)、古木蕨(Gumuia)和盘囊蕨(Discalis)等— — 排除在他们的“ 工蕨纲” 之外; 他们所列举的“ 工蕨纲” 的特点与传统的沙顿蕨目(Sawdoniales)一致。
Hao和Xue(2013a)最新的研究将米尔顿工蕨、古木蕨和盘囊蕨等这些植物, 都归入工蕨纲, 而将Hsü a、Barinophyton从工蕨纲中排除了出去。工蕨纲的共性包括扁圆形孢子囊侧生、远端开裂, 具椭圆形的、或圆形的外始式初生木质部束, 管胞螺纹加厚、具小穿孔等(Hao and Xue, 2013a)。除这些特征之外, Hao和Xue(2011)通过详细比较目前已知的工蕨类孢子囊结构, 提出工蕨类的另一个(可能的)近裔共性:孢子囊由两层壁构成, 外层为垂周分裂的细胞层(外垂周层), 而内层为平周分裂的细胞层(内平周层)。Niklas和Banks(1990)将工蕨类划分为“ 顶生类群” 和“ 非顶生类群” , 两者的区别在于生殖枝(孢子囊穗或疏松排列的孢子囊区域)是否具有一个顶生孢子囊, 这一特征决定了植物体的发育模式, 即具有顶生孢子囊代表了有限生长模式, 而顶生孢子囊的缺失代表了无限生长。Hao和Xue(2013a)将“ 顶生类群” 和“ 非顶生类群” 分别归为工蕨目(Zosterophyllales)和戈斯林蕨目(Gosslingiales), 各自下分若干个科。
工蕨类可能起源于某些拟莱尼蕨类(Gensel, 1992; Hueber, 1992; Gensel and Berry, 2001)。常被作为工蕨类祖先类型的是拟肾囊蕨类如Aberlemnia、Renalia和Hsü a(据Hao and Xue, 2013a的定义), 但这些植物的演化关系和系统位置仍存在不确定性(Kenrick and Crane, 1997; Hao and Xue, 2013a), 从拟肾囊蕨类到工蕨类的演化过程仍是一个谜题。一些工蕨类植物的性状特征不齐全(比如华南的工蕨类植物大多缺乏解剖结构信息), 这也使得目前对它们系统位置的认识有很大局限性。工蕨类在全球不同板块的时空分布模式、在不同陆块上的组成分子和性状演化上的区别和联系, 仍是后续研究的重点。
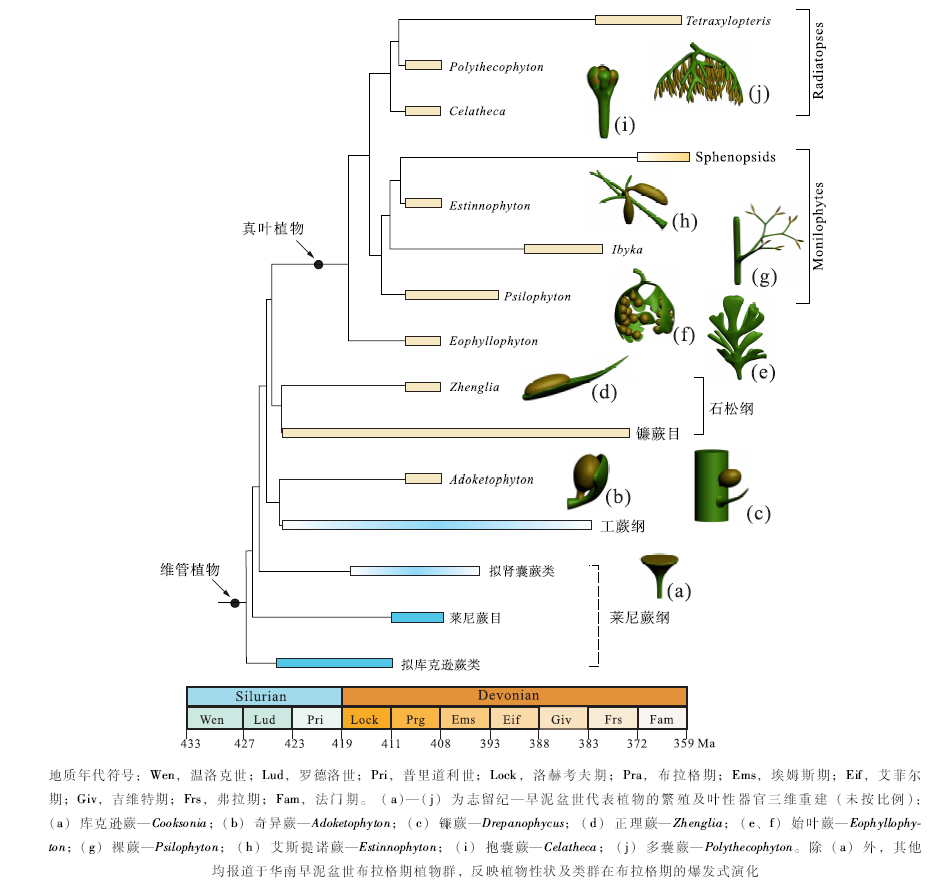
志留纪至早泥盆世的石松类植物较少, 包括Baragwanathia、Drepanophycus、Halleophyton、Hueberia、Asteroxylon、Zhenglia、Leclercqia以及其他若干有疑问的植物, 前4属被归入镰蕨目(Drepanophycales), 而后2属被归入原始鳞木目(Protolepidodendrales)。最早的石松类化石为发现于澳大利亚罗德洛世(Ludlow)地层中的Baragwanathia, 也见于华南布拉格期坡松冲组、加拿大埃姆斯期Sextant组(Lang and Cookson, 1935; Hueber, 1983; Hao and Gensel, 1998)。Drepanophycus常见于世界范围内的早泥盆世地层中(Li and Edwards, 1995), 可延续至中、晚泥盆世(Xu et al., 2013)。Asteroxylon发现于莱尼燧石层植物群中(Kidston and Lang, 1917; Edwards, 2004)。Halleophyton、Hueberia和Zhenglia这3属仅见于云南文山的坡松冲组(Li and Edwards, 1997; Hao et al., 2006; Yang et al., 2009; Xue, 2013)。其中Zhenglia具孢子叶(石松类孢子叶性状的最早记录), 其孢子叶密集排列形成最早的似叶球状结构, 叶基菱形, 规则排列成鳞木型叶序, 这一植物可能和中— 晚泥盆世的乔木状石松有一定演化联系(Hao et al., 2006)。Leclercqia最早出现于加拿大埃姆斯期地层中(Gensel and Kasper, 2005), 至中泥盆世, 其分布非常广泛(Xu et al., 2011), 但在华南尚无报道。
目前, 对石松类与一些可能的祖先类群之间的系统关系仍有争议。Kenrick和Crane(1997)认为石松类和工蕨类是姐妹群关系(图 1), 在分支图上, 一些工蕨植物介于他们的“ 工蕨纲” 和石松纲之间。基于这一框架, Kenrick和Crane(1997)提出“ 孢子囊不育化假说” , 认为在工蕨类的囊穗中, 紧密排列的两个孢子囊中下部的一个发生不育, 转变成石松类的孢子叶。但是, 这一假说只能解释孢子叶的来源, 而无法解释营养叶的起源, 并且孢子囊的不育化缺乏发育生物学的支持。另外, 石松类和工蕨类在志留纪罗德洛世差不多同时期最早出现, 时序上并没有先后之分, 似乎其祖裔相继的演化关系并不成立。Hao和Xue(2013a)的系统学分析中, 工蕨类和石松类的姐妹群关系未得到支持, 而石松类与真叶植物构成姐妹群(图 2)。工蕨类的显著特点是孢子囊扁平(flattened)、远端开裂, 孢子囊壁结构已知的类型都具有两层壁结构(Hao and Xue, 2011)。在早期石松类中, Asteroxylon的孢子囊与工蕨类的孢子囊最为相似(Lyon, 1964), 而其他类型如Drepanophycus、Halleophyton和Zhenglia的孢子囊大多以压型形式保存, 是否扁平难以确定, 但稍晚期的石松类如Leclercqia、Yuguangia等, 孢子囊并不扁平(Bonamo et al., 1988; Hao et al., 2007); Yuguangia仅具单层栅栏状细胞组成的孢子囊壁(Hao et al., 2007), 与工蕨类孢子囊差别很大。拳卷这一特征在工蕨类戈斯林蕨目中常见, 但在工蕨目和早期石松类中很罕见。Hueber(1992)倾向于认为石松类和工蕨类各自起源于传统意义上的“ 拟库克逊蕨类” (cooksonioids); 传统意义上的“ 拟库克逊蕨类” 较为混杂, 包含了Hao和Xue(2013a)所定义的拟库克逊蕨类、拟肾囊蕨类以及其他类群。最有可能的模式是, 石松类和工蕨类各自独立从拟肾囊蕨类中演化出现。但是, 不管在哪种模式下, 目前都不能很好地解释石松类小型叶、具单一孢子囊的孢子叶这两个性状的起源问题。这说明, 目前对石松类、工蕨类的一些性状特征及其同源性的认识还存在很大不足, 这也是后续研究的主要任务。
现生的非石松类维管植物被归入真叶植物支系(Pryer et al., 2002)。早泥盆世是真叶植物基部类群初始分化的时代。传统上, 通常认为早泥盆世的三枝蕨类(如著名的裸蕨Psilophyton)是真叶植物的祖先类群, 逐渐演化出真蕨类、楔叶类和种子植物等现代类群(Andrews et al., 1975), 但分支系统学的分析表明, 三枝蕨类是并系的或者多系的(Kenrick and Crane, 1997; Hao and Xue, 2013a), 应该摒弃。Hao和Xue(2013a)的分支分析中, 真叶植物的近裔共性包括:大型叶及其同源器官(多次分叉的小羽片状附枝)、不规则的叶序、孢子囊辐射对称、孢子囊与特化的生殖结构或大型叶相联系。早泥盆世布拉格期的始叶蕨(Eophyllophyton)位于真叶植物的最基部位置。
Beck和Stein(1993)通过广泛研究认为, 在泥盆纪时期, 种子植物和真蕨类、楔叶类已经在解剖结构方面发生了分化, 种子植物来源于“ 辐射原生木质部类群” (radiate protoxylem group), 而后两者来源于“ 持久原生木质部类群” (permanent protoxylem group)。Kenrick和Crane(1997)将这两个类群分别命名为Radiatopses和Moniliformopses(后被简化为Monilophytes, 见薛进庄等, 2014; 下文统一用Monilophytes), 它们互为姐妹群, 共同构成真叶植物。从字面上理解, Radiatopses源自拉丁语“ radiatus” (放射状), 意指原生木质部位于中柱臂的中间面上, 自中柱中央至中柱边缘都有分布, 呈放射状; 而Moniliformopses源自拉丁语“ moniliformis” (串珠状), 意指原生木质部位于中柱臂的边缘部位, 呈串珠状(薛进庄等, 2014)。这两个类群解剖结构特征的分化, 至少可追溯到中泥盆世, 如中泥盆世伊瑞蕨目的Ibyka具“ 持久原生木质部” , 而前裸子植物Tetraxylopteris具“ 辐射原生木质部” 。当然, 也有学者认为Radiatopses和Monilophytes的区别并非如此截然, 有一些中泥盆世植物如Langoxylon兼具了两个类群的一些解剖学特征(Scheckler et al., 2006)。
尽管Radiatopses和Monilophytes是依据解剖学特征建立的, 但是这两个类群的形态学特征特别是生殖结构方面的分化也比较明显, 时间上要更早, 可追溯到早泥盆世(图 2)。华南布拉格期坡松冲植物群中的Estinnophyton、Psilophyton和Pauthecophyton, 其生殖结构可与中泥盆世的伊瑞蕨目、瘤指蕨目植物相比较, 后两个目被归入枝蕨纲, 通常被视为Monilophytes的基部类群(Berry and Stein, 2000; Wang, 2008; 薛进庄和郝守刚, 2008; Xue et al., 2010)。同一植物群中的Polythecophyton和Hedeia, 其生殖结构中孢子囊侧生、羽状排列, 与Radiatopses的基部类群前裸子植物(如无脉树目)的生殖结构相似。坡松冲植物群中的另一真叶植物Celatheca是一个独特的类群, 其生殖结构位于枝顶, 由4个孢子囊组成, 每个孢子囊被叶性结构所包被, 这样一种结构与晚泥盆世的前胚珠或聚合囊相似。上述主要基于生殖结构性状对谱系关系的推断, 尚需从其他方面进行检验。Psilophyton的中柱为实心的圆柱, 初生木质部发生顺序为心始式(Hao and Xue, 2013a), 而其他几种植物的解剖结构了解甚少。Pauthecophyton、Polythecophyton和Hedeia的茎轴系统、叶性器官、原位孢子等有待进一步研究。对这些植物的详细研究对于探讨Radiatopses和Monilophytes的起源、 分化以及一些重要形态学性状的发生等问题至关重要(见2.3节)。
地质历史时期的生物多样性一般用已知时间间隔内的分类单元(如科、属、种等)数量、以及新生率、灭绝率等定量指标来衡量。与海相化石相比, 植物化石记录较为零散, 但是通过构建大型的化石数据库以及稀疏标准化等数据处理方法(如Niklas et al., 1983; Hao and Xue, 2013a; Xiong et al., 2013), 可以整合这些零散的化石记录, 降低由于研究程度、取样、埋藏等因素带来的偏差, 为揭示植物多样性动态提供较为可靠的依据。前人在这方面已开展了大量的工作, 但不同的作者所研究样本的时间跨度、所依据的数据来源(如不同板块、不同地区)、所分析的多样性指标不尽相同。
Niklas等(1983)对古生代以来的陆生维管植物多样性进行了系统研究, 认为志留纪至中泥盆世是维管植物的第1个演化阶段, 这一时段的维管植物被称为早期维管植物(early vascular plants), 以形态上简单的原始类型为主。Knoll等(1984)研究了欧美地区志留纪— 泥盆纪的植物大化石、微体化石属级多样性变化, 并分析了灭绝率、新生率等定量指标, 提出志留纪— 泥盆纪的维管植物属级多样性与古生代海洋无脊椎动物多样性变化具有可比性, 都在快速的辐射之后达到动态的平衡。Edwards和Davies(1990)基于欧美地区志留纪— 早泥盆世的属、种多样性, 认为Knoll等(1984)的推论过于大胆, 因为与海洋动物的化石记录相比, 植物化石记录的质量和数量都有很大缺陷。Edwards和Davies(1990)认为, 传统意义上(如Banks, 1975)的拟莱尼蕨类、工蕨类、三枝蕨类的多样性演化速率不同, 它们依次在志留纪— 泥盆纪最早期、晚洛赫考夫期— 早布拉格期、晚布拉格期之后达到多样性峰值。
王怿等(2006)统计了华南地区早期维管植物的多样性, 并与欧美地区的植物多样性曲线进行了对比, 指出两地区若干不同的特点, 特别是在华南地区, 早泥盆世布拉格期已达到多样性的高峰值, 而在欧美地区, 第1次高峰值出现于稍晚的埃姆斯期。Xiong等(2013)对华南植物大化石、微体化石的产地、剖面、属种多样性进行了更为精细的分析, 提出华南维管植物多样性模式与劳俄古陆大体相似, 但在华南地区, 植物的新生率和灭绝率在洛赫考夫期— 布拉格期最高, 之后逐渐递减, 植物多样性的演变与陆地生境的变化(可能与海平面变化有关)、广布属的比例有关。
在整合上述研究的基础上, Hao和Xue(2013a)建立了全球志留纪至早泥盆世的陆生植物(主要是维管植物)大化石和中化石(mesofossil)属种数据库。该数据库包含了劳俄、东北冈瓦纳(华南、澳大利亚)、西北冈瓦纳(巴西、阿根廷)、西伯利亚、哈萨克斯坦(如新疆准噶尔西部)等多个植物古地理区(板块或板块群)的资料, 时间跨度为志留纪兰多维列世(Llandovery)至早泥盆世。结果表明:植物的属种总数、每百万年属的数目、标准化后属的数目反映出同样的趋势, 从志留纪至洛赫考夫期缓慢增加, 至布拉格期急剧倍增。在演化速率方面, 首现属(新生属)、末现属(灭绝属)、每百万年首现属、每百万年末现属在布拉格期都达到高峰。在高级分类单元方面, 布拉格期的纲一级单元可达到11个, 远大于洛赫考夫期纲的数目(6个)。依据定量的多样性动态统计, 以及植物形态学、解剖学的比较研究, 志留纪至早泥盆世的植物演化可划分为两幕:(1)前布拉格期(pre-Pragian)演化阶段; (2)布拉格期的爆发阶段。
前布拉格期的维管植物以莱尼蕨类占优势地位, 工蕨类、石松类、真叶植物处于萌芽状态, 仅出现个别先驱型分子, 这一阶段可视为维管植物多样性演化的序幕。
志留纪早期的维管植物大化石记录颇有争议, 如华南的黔羽枝(Pinnatiramosus qianensis)。黔羽枝是发现于贵州凤冈县兰多维列统上部地层中的一种宏体植物, 它具有复杂的羽状分枝系统, 且具梯纹、网纹和圆形具缘纹孔管胞(耿宝印, 1986; Cai et al., 1996; Edwards et al., 2007; 王怿和徐洪河, 2009)。最近Wang等(2013)的研究认为, 黔羽枝不是志留纪植物, 而是二叠纪植物根系扎入到志留纪地层中并被保存下来的。
自志留纪温洛克世晚期(Homerian期, 约430 Ma)出现Cooksonia型植物大化石, 至早泥盆世洛赫考夫期末(约411 Ma), 这近20, Ma期间, 确凿的维管植物大化石记录在欧洲、澳大利亚、加拿大极区、中国、巴西、阿根廷、玻利维亚等地区均有分布, 但化石保存普遍比较破碎, 类型也较为单调, 其中最为丰富的是莱尼蕨类和工蕨类(Edwards and Wellman, 2001; Gensel, 2008; Hao and Xue, 2013a)。澳大利亚Victoria地区和加拿大极区Bathurst岛的罗德洛世地层中, 可见莱尼蕨类、工蕨类, 在前一地区可见石松类(Baragwanathia)以及可能的真叶植物(Hedeia)(Tims and Chambers, 1984; Kotyk and Basinger, 2002)。中国新疆北部的志留系普里道利统(Pridoli)乌吐布拉克组中产出Eocooksonia sphaerica (=Junggaria spinosa)、Filiformorama simplexa、Wutubulaka multidichotoma以及可能的石松类和工蕨类(王怿等, 2006; Wang et al., 2006, 2007; 王怿和徐洪河, 2009; 王祺和徐洪河, 2011)。Wutubulaka可能代表一类真叶植物的祖先类型(Kenrick et al., 2012)。华南确凿的志留纪维管植物为产自云南曲靖玉龙寺组顶部(普里道利世)的Zosterophyllum qujingense(Hao et al., 2007)。其他志留纪植物大化石分布于英格兰、威尔士、美国等地区, 以Cooksonia为主(Edwards and Wellman, 2001)。洛赫考夫期的植物大化石产地较多, 植物属、种比志留纪更为多样, 但仍以莱尼蕨类、工蕨类占主导, 个别产地见石松类(如Drepanophycus)和原始的真叶植物(Edwards and Wellman, 2001; Xue, 2012; Hao and Xue, 2013a)。这一阶段的维管植物分枝形式较为简单, 形态多样性(变异)主要体现在孢子囊的形状上(图 3)。孢子囊壁层结构、孢子特征等方面也可能存在很大差异, 但这些特征目前仅在少数属种中得以揭示(如Morris et al., 2011)。
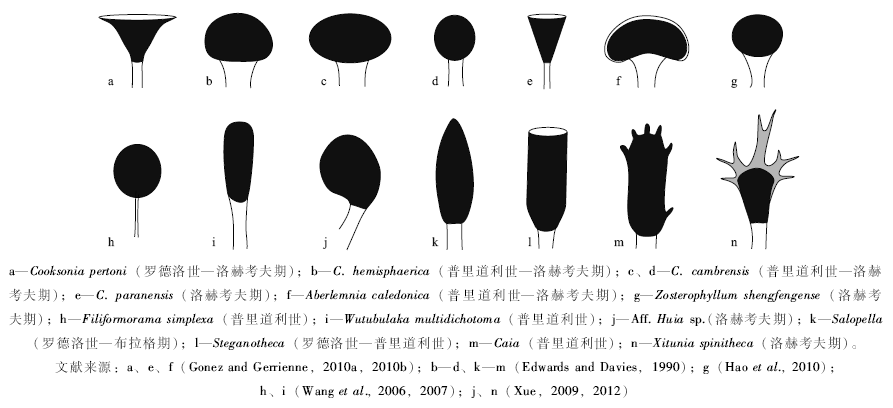 | 图3 志留纪— 洛赫考夫期代表植物的孢子囊形态Fig.3 Shape of sporangia of Silurian and Lochkovian representative plants |
Gray(1993)依据植物分散孢子化石和大化石记录, 将陆生植物的演化历史划分为3个阶段:始胚植物时代(Eoembryophytic epoch), 始维管植物时代(Eotracheophytic epoch)和真维管植物时代(Eutracheophytic epoch)(另见郝守刚等, 2002)。其中志留纪兰多维列世晚期至早泥盆世洛赫考夫期中期, 被认为是始维管植物时代; 洛赫考夫期晚期至二叠纪中期, 为真维管植物时代。但随着资料的积累, 这种划分方案显然需要改进。澳大利亚Victoria地区和加拿大极区Bathurst岛的罗德洛世植物群仅比最早的维管植物记录晚大约5, Ma, 其中的工蕨类、石松类和真叶植物可以和数十个百万年之后(如早泥盆世布拉格期)的类型相比拟, 这说明这些类群的起源时间非常早, 真维管植物时代至少应从罗德洛世开始。与维管植物古老的起源时间不同的是, 它们的多样性辐射却滞后地发生在布拉格期以来。
第2阶段是维管植物分支爆发式演化的主幕, 植物的形态、解剖特征在多个方面多样化, 特别是以生殖结构、叶性器官(如大型叶、孢子叶等)的演化分异最为显著。这一时期的植物群在世界范围内广泛分布, 以华南的坡松冲植物群和徐家冲植物群、澳大利亚的上Baragwanathia植物群、加拿大极区Bathurst岛植物群、英国威尔士Anglo-Welsh盆地植物群以及苏格兰莱尼燧石层植物群为代表(Hao and Xue, 2013a以及其中的文献)。在布拉格期, 植物大化石和分散孢子的多样性明显增加, 植物组织结构和器官急剧演化分异, 出现诸多可以与中晚泥盆世植物相比较的类群, 标志着现生维管植物的主要分支快速分化。中国云南的坡松冲植物群以其丰富的内涵、高度进化的营养器官和生殖器官组合特征给人们留下了深刻的印象, 为重构陆生植物多样性的起源和早期演化提供了极为珍贵的科学资料(郝守刚等, 2002; 郝守刚和王德明, 2003; 王怿等, 2006)。布拉格期植物群有以下几个特点:
1)莱尼蕨类、工蕨类仍然在布拉格期比较繁盛, 但两者表现出此消彼长的关系, 前者的多样性出现衰减, 而后者的多样性增加(Edwards and Davies, 1990; Hao and Xue, 2013a)。
2)虽然在前布拉格期已出现原始的石松类(如Baragwanathia、Drepanophycus), 但规则的叶序、具单一孢子囊的孢子叶、似叶球状结构等石松类的典型性状, 在布拉格期开始显现(如Zhenglia)(Hao et al., 2006)。
3)虽然真叶植物的个别分子在前布拉格期有记录, 但真叶植物的分支分化主要发生在布拉格期。Hao和Xue(2013a)的系统学分析表明, 布拉格期的真叶植物至少可以识别出5个支系(图 2):(Ⅰ )Psilophyton— 枝蕨纲伊瑞蕨目、瘤指蕨目支系(与真蕨类的起源有关); (Ⅱ )Estinnophyton— 楔叶纲支系; (Ⅲ )Polythecophyton(和Hedeia)— 前裸子植物纲无脉树目支系(与种子植物的起源有关); (Ⅳ )Celatheca— 种子蕨纲支系; (V)以Eophyllophyton为代表的始叶蕨纲支系(未发现直接后裔, 已灭绝)。这5个支系的分化主要体现在生殖结构的差异上(Hao and Xue, 2013b)。特别值得一提的是, 依据中晚泥盆世真叶植物解剖结构特征所建立的2个类群— — Radiatopses和Monilophytes— — 在早泥盆世布拉格期植物的生殖结构方面, 已发生了明显的分化, 分别以上述的支系(Ⅰ )、(Ⅱ )和支系(Ⅲ )、(Ⅳ )为代表。这为现生维管植物分子钟的研究(如Pryer et al., 2001)提供了新的时代标定点。
4)与前布拉格期植物显著不同的是, 布拉格期植物的侧生器官高度分异, 特别是真叶植物的大型叶(及相应的同源器官)。Hao和Xue(2013a, 2013b)将早期真叶植物的侧生营养器官, 称为“ 枝叶复合体” (branch-leaf complexes), 用以指示这些侧生分枝系统兼具枝、叶的双重属性, 它们的缩合、扁化和蹼化导致真正大型叶的出现。在布拉格期的5个真叶植物谱系中, 至少可见3种类型的枝叶复合体, 分别为Psilophyton-型(支系Ⅰ )、Estinnophyton-型(支系Ⅱ )和Celatheca-型(支系Ⅳ 、Ⅴ )。另外, 在始叶蕨支系(V)中, 同时兼具Celatheca-型的枝叶复合体和真正的大型叶。这些枝叶复合体各自独特的分枝方式, 预示着后期大型叶多样化的叶脉样式。大型叶的多次起源已经显现在布拉格期的几个支系中。
5)随着叶性器官的出现, 植物体孢子囊与叶性器官的组合关系(即生殖器官的组构)发生了巨大变化。前布拉格期的植物, 其孢子囊裸露、顶生或侧生、没有任何包被结构(如莱尼蕨类、工蕨类), 而在布拉格期植物中, 出现多样化的叶性结构伴生或包被孢子囊的现象, 如石松植物中的Zhenglia, 真叶植物中的Celatheca、Eophyllophyton, 以及诸多系统位置不太明确的植物如Adoketophyton、Bracteophyton、Stachyophyton和Dibracophyton。Adoketophyton的孢子囊扁圆形、椭圆形, 被扇形孢子叶包被(Li and Edwards, 1992; Hao et al., 2003b); Bracteophyton的生殖单元由1~2个苞片包被1个长卵形的孢子囊构成(Wang and Hao, 2004); Stachyophyton的孢子囊呈长卵形, 着生于顶端分叉的苞片上(耿宝印, 1983; 王怿和蔡重阳, 1996); Dibracophyton的孢子囊呈长椭圆形, 被上、下2枚苞片包被(Hao et al., 2012)。另一个有意思的发现是, 上述植物中Celatheca、Adoketophyton、Stachyophyton和Dibracophyton缺乏真正的营养叶(茎轴光滑或仅具枝叶复合体), Eophyllophyton的生殖叶化石也远远多于营养叶化石(Hao and Xue, 2013b)。孢子囊和叶性结构(苞片或孢子叶)的紧密联系, 是植物体躯体构型(body plan)的一种创新, 这说明孢子囊和叶性结构的发育相关性在布拉格期已成为普遍现象, 显示了植物的生殖器官在不同类群中的趋同演化现象。Hao和Xue(2013a, 2013b)将这种趋同演化与早泥盆世高大气二氧化碳浓度、高温等环境因素对植物生理学的影响联系了起来。这些包被孢子囊的叶性结构营光合作用和营养双重功能, 生殖叶的演化要早于营养叶, 说明植物生殖部分面临的选择压力更为强烈。
6)解剖结构方面, 布拉格期植物也体现出其复杂性以及与后续中晚泥盆世植物的延续性。由外始式或心始式初生木质部束构成的原生中柱在布拉格期普遍出现, 如Stachyophyton、Psilophyton、Eophyllophyton和 Adoketophyton等。据Gerrienne等(2011)的研究, 法国晚布拉格期至最早埃姆斯期地层中的一种类似于Psilophyton的植物已具有最早的次生木质部, 说明布拉格期植物已具备维管形成层和次生生长习性, 这为中、晚泥盆世森林的形成奠定了基础。前布拉格期的管胞类型仅有C-型、Ⅰ -型和G-型, 而布拉格期新出现了S-型和P-型管胞(Edwards, 2003)。Wang等(2003)基于曲靖徐家冲组Hsü a deflexa解剖结构的研究, 认为G-型管胞次生壁化学分异至少在布拉格期已经出现。特别值得一提的是, 在坡松冲植物群中的Yunia中, 茎轴的原生中柱中有2个由薄壁细胞组成的区域, 周围为原生木质部管胞, 这种结构被解释为“ 原始的髓” , 与真蕨类植物髓的起源有关(Hao and Beck, 1991)。
综上所述, 前布拉格期与布拉格期植物的差异是综合的、多方面的, 涉及到植物的营养器官、生殖器官和解剖结构等, 并最终反映在植物谱系的多样性上。此外, 布拉格期的一些植物株高可能达到米级, 如Celatheca和Polythecophyton, 而前布拉格期的植物一般比较纤弱、矮小, 如Hao 等(2010)发现的1株完整保存的洛赫考夫期工蕨植物(Zosterophyllum shengfengense), 其根、茎总长度仅约12 cm。
早期维管植物的地理分布格局一直是人们关注的问题, 但对于古植物地理区的具体划分存在不同意见。Edwards(1990)认为澳大利亚下Baragwanathia植物群(罗德洛世)肯定与劳俄地区的拟莱尼蕨组合不同, 但对这两个地区是否代表2个古植物地理区存有疑问。Raymond 等(2006)依据志留纪罗德洛世、普利多利世的植物大化石资料, 将全球该期植物群划分为北劳俄区(加拿大极区Bathurst岛)、南劳俄— 西北冈瓦纳区(英国、波多里亚、玻利维亚)、哈萨克斯坦区和东北冈瓦纳区(澳大利亚)4个植物古地理区。Hao等(2007)、Xue(2012)的研究表明, 在志留纪晚期— 早泥盆世洛赫考夫期, 工蕨类在以华南和加拿大极区为代表的低纬度地区占主导地位, 而在中、高纬度地区(劳俄大陆南部、冈瓦纳大陆西部), 占优势地位的是拟莱尼蕨类。
依据植物大化石属的分布, Raymond等(1985)将早泥盆世布拉格期— 埃姆斯期的维管植物群划分为澳大利亚、南冈瓦纳、赤道— 中纬度3个古植物地理区, 其中赤道— 中纬度区由西伯利亚、北美、欧洲、哈萨克斯坦和中国5个亚区组成。之后, Raymond(1987)又进一步研究了植物属种和性状的分布, 将早泥盆世植物群划分为澳大利亚、哈萨克斯坦— 北冈瓦纳、赤道— 中纬度区, 后者包含西伯利亚— 北劳俄、南劳俄和中国等3个亚区。Raymond等(1985)的上述划分, 主要是根据植物分类单元、性状在特定地区的有无编码形成数据矩阵, 再进行多变量定量分析(如聚类分析、对应分析), 来获得不同地区植物群之间相似性的定量估计。Edwards(1990)认为在早泥盆世洛赫考夫期, 可识别出具有独特植物组合的4个区域, 即劳俄、哈萨克斯坦、西伯利亚和澳大利亚, 但她对这些区域是否代表古植物地理区持保留意见, 认为仍有待更多化石资料的发现。然而, Edwards和Berry(1991)的综述认为早泥盆世时期, 中国、澳大利亚、西伯利亚、哈萨克斯坦和劳俄各自作为独立的古植物地理区似已是共识, 争议在于对于劳俄区的进一步细分。Hao和Gensel(1998)依据澳大利亚上Baragwanathia植物群、华南坡松冲植物群的相似性, 提出早泥盆世布拉格期“ 东北冈瓦纳古植物地理区” 的概念, 该区包括华南和澳大利亚2个亚区。
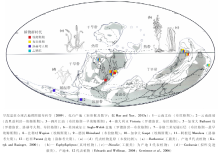
基于近年来的新资料(图 4), 在东北冈瓦纳区, 云南处于古赤道附近, 植物化石产出层位以曲靖的下西山村组、西屯组、徐家冲组和文山的坡松冲组为代表。化石组合中最为丰富的是工蕨类, 且以孢子囊螺旋排列的类型为主; 拟肾囊蕨类仅少量分布(Gonez et al., 2012; Xu et al., 2014); 最具特色的是, 东北冈瓦纳区的真叶植物在布拉格期已显示了高度的多样性和分异度。在劳俄区, 加拿大极区位于古赤道附近, 所以与华南显示了一定的相似性, 工蕨类植物自罗德洛世至布拉格期都占主导地位。西欧处于中纬度地区, 工蕨类和莱尼蕨类同时发育。北美(加拿大魁北克、新不伦瑞克)产有丰富的埃姆斯期植物组合, 具拟肾囊蕨类、工蕨类、石松类和真叶植物, 和华南布拉格期植物群有一定相似性。劳俄区的植物群中, 孢子囊成列排列的工蕨类非常丰富, 这与东北冈瓦纳区有显著不同。另外, 华南丰富的、较为进化的真叶植物如Polythecophyton、Celatheca和Eophyllophyton等出现于布拉格期, 而可比较的真叶植物在劳俄区较少见、或出现于埃姆斯期甚至更晚。
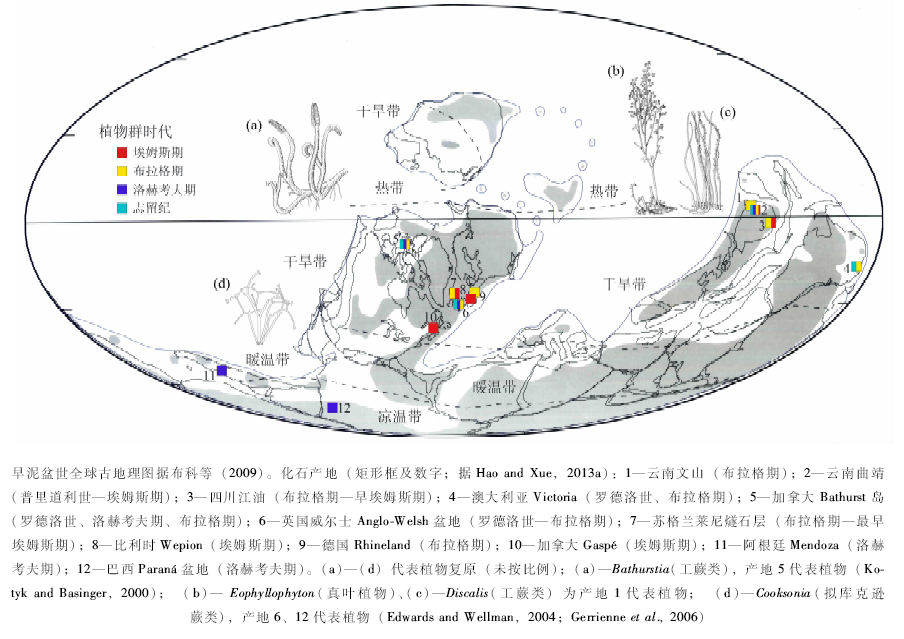 | 图4 志留纪— 早泥盆世维管植物大化石重点产地及代表性植物Fig.4 Selected localities of Silurian-Early Devonian vascular plant megafossils and representative plants |
上述关于早期维管植物古地理分区的假说和认识, 尽管得到诸多属种的定性比较、相似性系数等定量古地理学分析的支持, 但仍有待于更多化石资料的检验。未来的研究需从化石属种鉴定的可靠性、植物群时代的确切性、不同植物群之间的时序对比以及沉积相区对植物群组合的影响等多个方面进行深入探讨。除此之外, 目前尚没有分支生物地理的相关研究, 即在谱系关系框架下, 对早期维管植物的地理分布与辐射进行讨论。下文依据最新的系统发育框架和早期维管植物的属种数据库, 谨做一些初步的推断:
1)羊角蕨纲(Horneophytopsida)、拟库克逊蕨类(Cooksonia各种)、莱尼蕨目(如Rhynia、Aglaophyton)位于维管植物的最基干位置, 主要分布于西欧、北美、阿根廷等地区, 但它们在低纬度的华南地区、加拿大极区缺失(或非常罕见)。这说明上述类群的分支演化可能主要发生在南劳俄和西北冈瓦纳等中、高纬度地区。拟库克逊蕨类的最早出现时间为温洛克世, 是维管植物的最早记录。
2)最早的工蕨类植物产自加拿大极区(古赤道附近)的罗德洛世地层。自罗德洛世至布拉格期, 工蕨类在低纬度地区植物群中一直占据优势地位, 似乎表明低纬度地区是工蕨类的起源和辐射中心。工蕨类内部的2个目(工蕨目和戈斯林蕨目)都体现了跨大洋分布的特点, 在欧洲、北美和华南都有代表分子, 但优势类群不尽相同, 它们的分支演化和地理分布显示出错综复杂的镶嵌关系。
3)石松类植物最早出现于罗德洛世(澳大利亚), 普利多利世、洛赫考夫期仅非常零星地分布(新疆、德国), 至布拉格期在华南获得很大发展(坡松冲植物群中4属5种、徐家冲植物群中至少1属1种), 且出现诸多革新性状(孢子叶、似叶球状结构、规则叶序)。因此, 现有证据表明, 石松类的基干类群和真正的石松类都起源于东北冈瓦纳区, 且最早在该区辐射。
4)真叶植物中, Psilophyton— 枝蕨纲支系、Estinnophyton— 楔叶纲支系、Polythecophyton— 前裸子植物纲支系、Celatheca— 种子蕨纲支系均最早出现于云南的布拉格期坡松冲植物群中; 始叶蕨纲支系虽未发现直接后裔, 但其典型的大型叶是地球上最古老的大型叶记录。尤为重要的是, 真叶植物的最重要分化事件, 即Radiatopses和Monilophytes的分化, 在坡松冲植物群中已经显现, Radiatopses演化出现代的种子植物, 而Monilophytes演化出现代的真蕨类、楔叶类等。因此, 布拉格期植物群事实上奠定了后续真叶植物群的演化基础, 真叶植物基干类群的分支演化最早发生在东北冈瓦纳区, 特别是华南亚区。这被认为与华南长期处于古赤道附近的低纬度地区有关(Hao and Xue, 2013a)。
Jablonski(1993)通过研究中生代以来的海洋动物, 认为热带海洋是演化革新的重要源泉。Jablonski等(2006)进一步提出“ 走出热带” 的宏观演化模式(” out of the tropics” model), 认为热带是生物多样性的摇篮和博物馆, 属种倾向于在热带起源, 并向两级扩散。现代生态学的研究表明, 植物、无脊椎动物和脊椎动物的属种、高级分类单元、革新性状在热带地区的频度要高出很多(Hillebrand, 2004), 这可能是由于热带地区高新生率、低灭绝率所致, 特别是由于前者(Jablonski, 1993)。早期维管植物的研究表明, 至少工蕨类、石松类和真叶植物是在低纬度地区最早发生分化并快速辐射的; 维管植物谱系在布拉格期的爆发式演化, 主要是热带植物群革新演化的贡献。也就是说, 早期维管植物中的一些类群的分支演化和地理辐射可能也遵循“ 走出热带” 模式; 当然, 并不排除另一些类群以非热带地区为分化中心的可能性。这还需更多实证研究予以检验。
1)志留纪— 早泥盆世是早期陆生维管植物谱系分化的关键时期。近年来基于分支系统学分析, 对这一时段的多囊蕨类、莱尼蕨类、工蕨类、石松类、真叶植物等若干类群的系统发育关系的研究更为深入, 但尚未取得一致认识。主要基于华南资料, Hao和Xue(2013a)对早期维管植物提出了新的谱系关系假说和新的分类方案。被归入“ 前维管植物” 的Aglaophyton、Horneophyton、Nothia等植物的输水细胞的属性仍有争议, 多囊蕨类这一分支的概念还有待深入研究。莱尼蕨纲(并系或多系)下分莱尼蕨目、拟库克逊蕨类(cooksonioids)、拟肾囊蕨类(renalioids)和其他有疑问的植物。羊角蕨纲、拟库克逊蕨类、莱尼蕨目位于维管植物的最基干位置。工蕨纲下分工蕨目(Zosterophyllales)和戈斯林蕨目(Gosslingiales)。工蕨纲和石松纲可能各自起源于拟肾囊蕨类。真叶植物的谱系分化发生在早泥盆世布拉格期, 这一时期可识别出5大支系:Psilophyton— 枝蕨纲支系、Estinnophyton— 楔叶纲支系、Polythecophyton(和Hedeia)— 前裸子植物纲支系、Celatheca— 种子蕨纲支系、以Eophyllophyton为代表的始叶蕨纲支系。依据中晚泥盆世真叶植物解剖结构特征所建立的2个类群— — Radiatopses和Monilophytes— — 在早泥盆世布拉格期植物的生殖结构方面, 已发生了明显的分化。对上述类群性状特征的补充和完善, 揭示不同类群的时空分布模式, 探讨一些关键革新性状的起源等方面, 是未来的研究重点。对真叶植物基部类群的研究尤为关键。
2)定量的多样性动态统计和植物形态学、解剖学的比较研究表明, 志留纪— 早泥盆世的维管植物演化可划分为2幕:前布拉格期序幕阶段和布拉格期爆发阶段。第1幕的植物类型以莱尼蕨类植物占优势地位, 工蕨类、石松类、真叶植物处于萌芽状态, 仅出现个别先驱型分子。第2幕是维管植物分支爆发式演化的主幕, 植物的形态、解剖特征在多个方面发生多样化。莱尼蕨类、工蕨类在布拉格期仍比较繁盛, 但两者表现出此消彼长的关系。规则的叶序、具单一孢子囊的孢子叶、似叶球状结构等石松类的典型性状, 在布拉格期开始显现。真叶植物的分支分化发生在布拉格期。布拉格期植物的侧生器官高度分异, 特别是真叶植物的大型叶(及相应的同源器官)。在布拉格期的5个真叶植物支系中, 至少可见3种类型的枝叶复合体, 它们各自独特的分枝方式, 预示着后期大型叶多样化的叶脉样式。大型叶的多次起源已经显现在布拉格期的真叶植物支系中。孢子囊和叶性结构的发育相关性在布拉格期已成为普遍现象, 孢子囊和叶性结构(苞片或孢子叶)的紧密联系, 是植物体躯体构型(body plan)的一种创新。解剖结构方面, 布拉格期植物也体现出其复杂性以及与后续中晚泥盆世植物的延续性。
3)古植物地理方面, 在东北冈瓦纳区的华南亚区, 化石组合中最为丰富的是工蕨类, 且以孢子囊螺旋排列的类型为主; 最具特色的是, 华南亚区的真叶植物在布拉格期已显示了高度的多样性和分异度, 而可比较的真叶植物在劳俄区较少见、或出现于埃姆斯期或者稍晚。在劳俄区的加拿大极区, 工蕨类植物自罗德洛世至布拉格期都占主导地位; 而劳俄区的西欧地区, 工蕨类和莱尼蕨类同时发育; 北美产丰富的埃姆斯期植物组合, 具拟肾囊蕨类、工蕨类、石松类和真叶植物, 和华南布拉格期植物群有一定相似性。劳俄区的植物群中, 孢子囊成列排列的工蕨类非常丰富, 这与东北冈瓦纳区有显著不同。
4)羊角蕨纲、拟库克逊蕨类、莱尼蕨目(顶生孢子囊的类型)主要分布于西欧、北美、阿根廷等地区, 这些类群的分支演化可能主要发生在南劳俄和西北冈瓦纳等中、高纬度地区。但是, 与上述类群不同的是, 工蕨类、石松类和真叶植物是在低纬度地区最早发生分化并快速辐射的; 维管植物分支在布拉格期的爆发式演化, 主要是热带植物群革新演化的贡献。也就是说, 早期维管植物中一些类群的分支演化和地理辐射可能遵循“ 走出热带” 模式。
致谢 感谢审稿专家对本文提出的建设性意见和建议。感谢中国科学院植物研究所的王祺博士对本文初稿的修改和有益讨论。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|
| 99 |
|
| 100 |
|
| 101 |
|
| 102 |
|
| 103 |
|
| 104 |
|
| 105 |
|
| 106 |
|
| 107 |
|
| 108 |
|
| 109 |
|
| 110 |
|
| 111 |
|
| 112 |
|