
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
海相遗迹学的成就、问题与展望
[范若颖 , 龚一鸣]
, 龚一鸣]
, 龚一鸣]
|
|

通讯作者简介 龚一鸣,男,1958年生,中国地质大学(武汉)教授、博士生导师,主要从事遗迹化石、泥盆系和地球生物学研究。E-mail: ymgong@cug.edu.cn。
第一作者简介 范若颖,女,1990年生,中国地质大学(武汉)博士研究生,专业为古生物学与地层学,主要从事遗迹化石、沉积学研究。E-mail: cugfry@163.com。
海相遗迹学经历了博物学者时代、藻类时代、大辩论时代、黎明时代,目前正处于现代遗迹学时代。海相遗迹学发展历程中的里程碑事件包括:博物学者对遗迹化石的初步研究、遗迹化石真实属性的确立、德国森根堡海洋研究所的建立、遗迹化石的行为习性分类,以及遗迹相、遗迹组构等重要概念的提出和发展。在系统回顾总结海相遗迹学成就和问题的基础上,着重展示了海相遗迹学发展的 3个方向:遗迹化石三维形态重建与形态发生模拟,综合遗迹分类学以及微观手段的运用。海相遗迹学在诠释遗迹化石所反映的生物行为习性以及不同尺度生物—环境相互作用与协同演化等方面具有重要意义。
About the corresponding author Gong Yiming,born in 1958,is a professor and Ph.D. supervisor of China University of Geosciences(Wuhan),and is mainly engaged in ichnology,the Devonian and geobiology. E-mail: ymgong@cug.edu.cn.
About the first author Fan Ruoying,born in 1990,is a Ph.D. candidate of China University of Geosciences(Wuhan),and majors in palaeontology and stratigraphy. Now she is mainly engaged in ichnology and sedimentology. E-mail: cugfry@163.com.
Marine ichnology mainly discusses marine invertebrate trace fossils and embraces a long history of more than 200 years,which dates back as far as the Renaissance-the Age of Naturalists. After the Age of Fucoids,Period of Controversy,and the Development of the Modern Approach,marine ichnology is currently in the Modern Era. Key events in the history of marine ichnology include:The preliminary study of marine ichnofossils by naturalists such as Leonardo da Vinci,clarification of “fucoids”as animal traces through experiments by A.G. Nathorst,ethological classification of trace fossils by Adolf Seilacher,establishment of Senckenberg am Meer in Germany by Rudolf Richter,and the proposition and practice of the ichnofacies and ichnofabrics concepts. However,with the vast development of computer and life sciences,along with much progress in experimental tools,a period of interdisciplinary ichnological studies dictate. This review acknowledges the rich legacies of ichnology in the past and highlights three promising areas where marine ichnology can go:(1)three-dimensional morphological reconstruction and behavioral simulation(morphogenesis)of traces fossils;(2)comprehensive ichnotaxonomy incorporating multi-scale geometrical description,such as topology and fractal geometry;and(3)application of geochemical and geobiological analyses. Intensive studies on these three topics will render important messages to the interpretation of fossil behaviors,their palaeobiology,as well as organism-environment interaction and co-evolution on various scales.
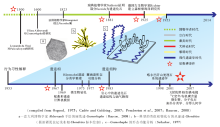
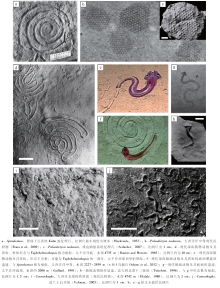
遗迹化石(trace fossil或ichnofossil)或生物遗迹(biogenic/organic trace, 德文lebensspur, 法文trace d' activité )指生物的生命活动在沉积物或坚硬底质中形成的结构构造(Hä ntzschel, 1975)。作为一种自然实体, 遗迹化石最早在文艺复兴时期即有文字记载(Baucon, 2008; 图1-a), 但直到19世纪初才始得广泛研究。无脊椎动物遗迹学与脊椎动物遗迹学相互独立发展。脊椎动物的遗迹主要是足迹, 起初就被认为是动物行走留下的痕迹。海相遗迹学主要研究海相无脊椎动物遗迹。欧洲白垩纪— 新近纪复理石相遗迹化石的研究拉开了海相无脊椎动物遗迹学的序幕, 然而它们最早被认为是藻类化石。典型的复理石相遗迹化石多沿水平层面分布, 具有分枝、弯曲、重复的形态, 与植物十分相像(图 1-b)。法国植物学家Brongniart(1823)建立了Fucoides属, 用于这些“ 藻类化石” 的系统分类。19世纪末陆续有学者质疑这些化石(fucoids)的“ 藻类” 属性(见Pemberton et al., 2007)。最早用实验证明fucoids中大部分为遗迹化石的是瑞典植物学家Nathorst(1886)。此后的近20年里, 学术界展开了对fucoids本质的激烈辩论, 直至20世纪初, 人们才普遍接受“ fucoids” 实为动物遗迹(Osgood, 1975)。
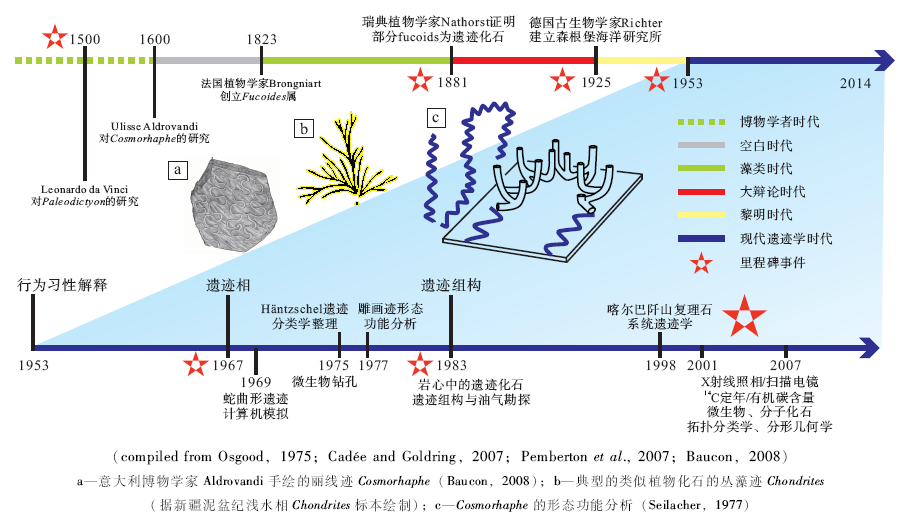 | 图1 海相遗迹学研究历史略图(参考Osgood, 1975; Cadé e and Goldring, 2007; Pemberton et al., 2007; Baucon, 2008等编制)Fig.1 Historical scheme of the development of marine ichnology (compiled from Osgood, 1975; Cadé e and Goldring, 2007; Pemberton et al., 2007; Baucon, 2008) |
遗迹学早期研究中心在欧洲和北美。欧洲中心又集中在意大利、法国和德国, 分别为16— 17世纪, 18— 19世纪和19— 20世纪西方科学中心。17— 18世纪, 英国为世界的科学中心, 但这一时期的遗迹化石研究几乎为零, 成为遗迹学史上的研究空白(图 1)。1929年, 德国古生物学家Rudolf Richter在瓦登海附近建立了森根堡海洋研究所(Senckenberg am Meer)。这是第1个专门研究现代沉积环境的海洋研究所, 现代生物遗迹研究在建立之初便是其重要研究方向之一, 被称为无脊椎动物遗迹学的摇篮(Cadé e and Goldring, 2007)。自此遗迹学研究开始步入正轨。北美遗迹学研究可分为加拿大学派(主要由加拿大地质调查局的地质学家组成)和辛辛那提学派(主要由古生物爱好者组成)。欧洲和北美在遗迹学早期阶段各自独立发展(Pemberton et al., 2007)。
先后有不少遗迹学者对遗迹学的发展阶段作了总结。Osgood(1975)将遗迹学发展历史分为3大阶段:藻类时代(1823-1881), 大辩论时代(1881-1925)和现代遗迹学手段初创(文中称之为黎明时代)(1925-1953)。Pemberton 等(2007)将1953年至现在的这一阶段称为“ 现代遗迹学时代” 。Baucon(2008)将文艺复兴时期博物学者(如达· 芬奇)对遗迹化石的初步研究也纳入遗迹学发展历程中, 建立了“ 博物学者时代” (图 1)。
20世纪中后叶开始, 世界范围内深海和大洋钻探计划的实施, 提供了大量第四纪沉积物岩心样品, 遗迹化石与海洋环境、第四纪气候变化的关系得以窥探(图 1)。海底箱式取样、海底摄像也使研究现代海底(特别是深海)的生物扰动作用成为可能。作为一门日益成熟的学科, 遗迹学中各种定量、微观手段广为发展, 得益于数学理论、计算机技术以及先进实验设备的运用。进入21世纪, 遗迹学开始向多极化发展, 世界各地发展起众多遗迹学研究团队, 遗迹学家之间广泛交流并尝试联合海洋科学、地球化学、地球生物学等领域进行跨学科研究。
国内海相遗迹化石系统的研究始于20世纪80年代, 起步较晚, 多集中于遗迹化石的分类鉴定以及简单的古环境和古生态分析, 综合性、理论性的研究成果较少(龚一鸣等, 2009)。这些基础的分类学资料和环境分析是进一步深入研究的重要素材, 有待于新一代遗迹学者的开发和利用(如杨式溥等, 2004)。龚一鸣等(2009)对国内遗迹学研究现状作了较为全面的论述, 此处不加赘述。文中主要关注国外海相遗迹学的动态, 大致可以反映较为完整的海相遗迹学的研究现状。由于深水相遗迹化石形态分异度高、保存程度好, 且几乎包含了所有浅水类型, 因此深水相遗迹化石的研究对于海相遗迹学具有重要的价值, 是文中重点关注的内容。
遗迹化石最常用的分类单元为遗迹属和遗迹种, 遗迹科、遗迹亚属、遗迹亚种和变种则较为少见, 遗迹科以上分类单元基本不存在。这些遗迹学分类单元仅代表遗迹化石形态的不同类别, 而不具有生物分类位置或亲缘关系的意义。采用双名法命名遗迹化石既可以吸收以往大量文献的分类学资料, 也便于遗迹化石名的修订、传播与应用(见Bertling et al., 2006)。遗迹化石不仅反映了造迹生物的形态、行为特点, 同时也是特定沉积、埋藏环境的产物。遗迹化石的命名同时考虑了这两方面的特征。遗迹化石最为显著的形态、行为特征用于区分遗迹属, 而较不重要的形态、行为特征(包括潜穴直径、埋藏状况引起的形态差异等)为次要鉴定特征, 用于区分遗迹种, 有时根据细节特征可进一步划分遗迹亚种或变种。前人还将若干遗迹属归为更高的分类单元(遗迹类群或遗迹科), 如动藻迹类群(Zoophycos Group)(Uchman, 1995)、雕画迹(Graphoglyptids, 遗迹科级别)(Seilacher, 2007)等。
遗迹化石为生物成因的沉积构造, 据其在赋存沉积物中的产出状态(toponomy)可分为若干类型。广泛使用的包括Seilacher(1964)和Martinsson(1970)的分类方案。其中Seilacher(1964)对遗迹化石与地层的实际产出关系以及可能的成因类型作了详细的分析。Chamberlain(1971)在Seilacher(1964)的基础上, 给出了遗迹化石的保存状态和成因类型图解, 基本可以解释所有可能的遗迹化石与地层的产出关系。遗迹化石的保存通常有赖于沉积物中的岩性差异。
欧洲白垩纪— 新近纪复理石相遗迹化石得到较为详实的研究, 种类丰富, 可作为形态分类的良好素材。Uchman(1998)通过对波兰南部喀尔巴阡山复理石相遗迹化石的系统研究, 在Ksi
对遗迹学研究影响更为深远的是行为习性分类。Seilacher(1953)首次将遗迹化石所反映的动物行为习性归为5大类:(1)爬行迹(Repichnia), 代表简单的运动; (2)牧食迹(Pascichnia), 代表动物在底质表面边运动边取食的行为; (3)觅食迹(Fodinichnia), 代表动物在沉积物内部营造潜穴用于觅食; (4)停息迹(Cubichnia), 代表动物将自身埋藏在泥沙中, 以暂时躲避天敌、环境对自身的伤害或在底质上停歇休息; (5)居住迹(Domichnia), 代表动物生活在相对永久的潜穴中, 以获取潜穴内部空间或外部的氧气和食物为生。随着遗迹化石研究的不断深入, 更多的行为习性类别被提出, 如逃逸迹(Fugichnia)、耕作迹(Agrichnia)、捕食迹(Praedichnia)(Ekdale, 1985)、化学生物迹(Chemichnia)(Seilacher, 1990)和诱捕迹(Irretichnia)(Lehane and Ekdale, 2013)。
遗迹沉积学一词在国内仅有零星使用(如龚一鸣和Droser, 2002), 在国外则主要源自Savrda(1993), 用作“ 遗迹学和沉积学” 的简称。真正系统的综合遗迹学和沉积学古环境分析方法, 直到近几年才发展起来(如Heard and Pickering, 2008; Angulo and Buatois, 2012), 但并没有一个明晰的定义。遗迹沉积学是介于遗迹学与沉积学之间的交叉学科, 是指通过对生物遗迹产出状态、埋藏、形态和行为习性及其寄主沉积的无机特征的综合分析, 精细识别沉积环境参数(如沉积速率、水动力、底质粘度、古氧相和盐度等)和类型(如海相、过渡相和陆相)的地质学分支学科。遗迹相和遗迹组构是遗迹沉积学中的两个核心概念, 而形态功能分析、定量遗迹学、新遗迹学等均是其重要分析手段。
2.1.1 遗迹相
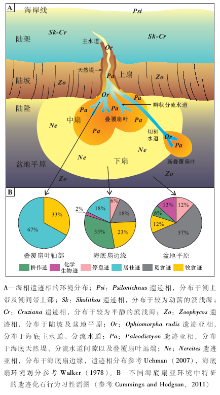
遗迹相是指特定环境所具有的相对稳定的、在时间和空间上具有复现性的生物遗迹组合(Seilacher, 1967)。从最初Seilacher(1967)的5种遗迹相发展至今, 许多新的遗迹相被发现, 现在被广泛认可的遗迹相共有11种, 其分布于海洋、河湖以及土壤等环境。其中Scoyenia, Mermia, Coprinisphaera遗迹相属于陆相遗迹相, Psilonichnus, Skolithos, Cruziana, Zoophycos和Nereites遗迹相属于海相遗迹相, 其余的Glossifungites, Teredolites和Trypanites遗迹相是以底质命名的, 分别对应于固底、木底和硬底(MacEachern et al., 2007)。
遗迹相概念的本质是对应于一组复杂、相互作用的环境因素的遗迹组合特征(Buatois and Má ngano, 2011), 也即遗迹相与具体、复杂的环境因素有关, 如水动力、沉积速率、氧含量、营养物质含量等, 沉积环境大类或水深并不是其直接决定因素(Ekdale, 1988; Frey et al., 1990)。由于风暴或浊流作用的影响, 在某段时间内底质的水动力条件会发生较大的变化, 此时沉积物中相应层段的不同遗迹相代表的并不是海水深度的突然变化, 而是水动力、盐度以及沉积速率等环境条件的突变。一般而言, 各种环境因素, 例如水动力、氧含量、营养条件、盐度、浊流等随海水深度发生有规律的变化。因此, 不同遗迹相(代表不同生态类型的底栖生物群落)通常对应于不同水深的海底环境(图 2-A)。
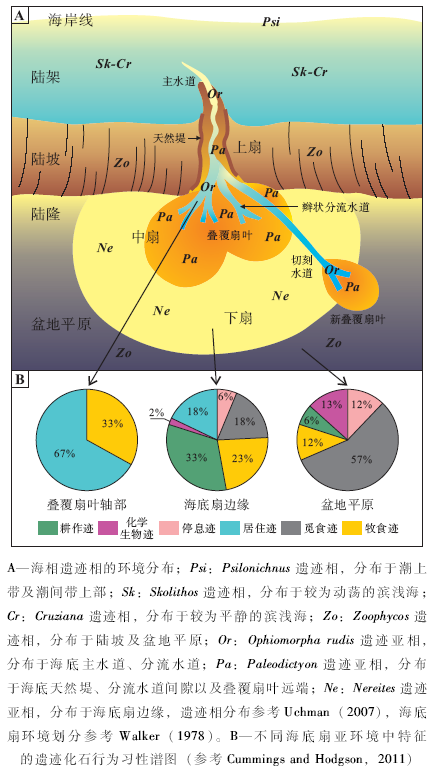 | 图2 海相遗迹相以及海底扇相关遗迹亚相的环境分布与遗迹化石行为习性谱图Fig.2 Environmental distribution of marine ichnofacies, with an emphasis on submarine fan related ichnosubfacies and ethological diagrams |
在5种海相遗迹相中, 只有Nereites遗迹相主要分布于陆隆— 盆地平原环境, 是与海水深度耦合最好的遗迹相。Nereites遗迹相具有一些深水相独有的遗迹化石类型, 反映了深海环境、深海底栖生物及其行为习性的特殊性。Nereites遗迹相特征的遗迹化石类群是雕画迹(Graphoglyptids), 代表黑暗、低能、贫营养的半深海— 深海软底质环境(Wetzel and Uchman, 1998)。在半深海— 深海浊积岩序列中, 雕画迹通常表现为平行层面分布的放射状、蛇曲状、螺旋状或网状等较为规则的形态(Uchman, 2003)。雕画迹通常个体较小、分异度较高、具有较窄的生态位, 为适应长期平稳的半远洋— 远洋沉积环境的平衡遗迹群落(K-选择)(Ekdale, 1985)。
Nereites遗迹相可分为若干遗迹亚相, 对粗略识别海底扇亚环境具有一定指示意义。Seilacher(1974)将Nereites遗迹相分为Paleodictyon遗迹亚相和Nereites遗迹亚相, 分别代表富砂和富泥的远端浊流环境。Uchman(2001)建立了Ophiomorpha rudis遗迹亚相, 用于表征海底扇水道和叠覆扇叶轴部厚层砂岩的生物遗迹特征。遗迹相是指导性的抽象模式, 一经建立就较为稳固。因此后续研究主要在于识别特殊遗迹相(如深水相中出现典型的浅水型遗迹相, 或是浅水相中出现典型的深海型遗迹相), 进一步探究地质历史中遗迹相的演化、变迁等方面(如Crimes and Fedonkin, 1994; Uchman, 2007)。
行为习性谱图是遗迹相研究中常用的半定量手段。行为习性谱图将不同沉积亚环境的遗迹化石按照行为习性类别进行多样性统计, 得出不同行为习性类别遗迹属的百分比, 多以饼图呈现(图 2-B); 或将遗迹化石分异度、浊流前/后遗迹化石分异度、雕画迹分异度按不同沉积亚环境进行统计, 多以柱状图呈现。行为习性谱图简单明了, 可用于刻画不同海底扇亚环境的生物— 环境特征(如Heard and Pickering, 2008; Cummings and Hodgson, 2011)。然而对于海底扇亚环境与遗迹化石的耦合关系研究方面, 缺乏对沉积环境的定量描述和刻画, 例如沉积速率、氧含量、有机质含量等, 这需要多学科综合研究, 特别是地球化学、沉积学和地球生物学等手段的运用。
2.1.2 遗迹组构
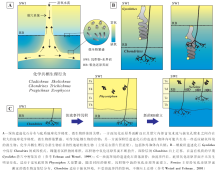
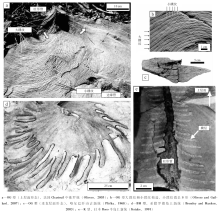
遗迹相是具有预测性的生物— 环境概念模型, 然而对于精细刻画生物— 底质的相互作用则明显不足。从这点来看, 遗迹组构是对遗迹相概念的良好补充。遗迹组构指地层中所有生物扰动或侵蚀作用形成的生物— 沉积构造(Ekdale and Bromley, 1983; Buatois and Má ngano, 2011)。遗迹组构中最为重要的概念是阶层(tiering), 即生物遗迹(或生物群落)在沉积物不同深度分布所构成的群落结构, 反映了沉积物中垂向上生境的分层特征(Ausich and Bottjer, 1982, 图3)。遗迹群团(或译为遗迹社群)(ichno-guild)则是指特定阶层所具有的遗迹化石组合特征。一方面, 底质在垂向上具有物理、化学和生物等多种分层性。在较为连续的沉积条件下, 表栖和内栖生物会随着沉积物的堆积, 不断向上迁移和演替。另一方面, 风暴、浊流或火山灰等事件沉积会使底质生态环境发生突变(Bromley and Ekdale, 1986), 并可能保留有“ 冻结阶层” (frozen tier), 对认识复杂阶层型式具有重要作用(Taylor et al., 2003)。生物扰动指数(Bioturbation index, BI)和遗迹组构组成图是遗迹组构基本的描述参数和研究方法(参见Taylor and Goldring, 1993)。
 | 图3 深海遗迹组构与底质氧化还原界面和其他地球化学条件变化的密切关联Fig.3 Sketch showing the close relationships of deep-sea ichnofabrics with redox boundary fluctuations and other geochemical gradients |
深海遗迹组构主要指浊积岩序列中的遗迹组构。对于一般的浊积岩序列, 遗迹化石主要见于鲍马序列Tc-Te段, 为浊流后机会遗迹群落, 典型遗迹化石包括Phycosiphon和Nereites, 并被适应缺氧环境、从下部向上迁移的Chondrites交切(Wetzel and Uchman, 2001; Uchman, 2007, 图3-C)。深海环境特有的雕画迹遗迹群落主要发育于鲍马序列的Te段, 通常在浊积砂岩底面呈半浮雕或全浮雕形式保存, 在沉积物纵切面上较难观察。大洋钻探岩心是深海遗迹组构的重要研究素材(Chamberlain, 1975)。Callow 和 McIlroy(2011)对显生宙浊积岩遗迹组构统计发现, 只有为数不多的遗迹属是浊积岩遗迹组构的常见分子, 包括Chondrites, Planolites, Scolicia, Zoophycos, Ophiomorpha, Phycosiphon, Thalassinoides和Nereites。Knaust(2009)通过对挪威海晚白垩世海底扇岩心的遗迹组构分析, 提出7种遗迹组构。其中4种为具有复现性的遗迹组构, 代表分布范围较广泛的沉积亚环境; 其余3种为局域性的遗迹组构, 代表时空分布较局限的特殊环境。Callow等(2013)对墨西哥晚白垩世的一套海底水道— 天然堤地层进行了露头遗迹组构研究, 总结了5个遗迹组构大类, 共12种遗迹组构。深海最为常见的遗迹组构有以下9种:Ophiomorpha遗迹组构、Planolites遗迹组构、Scolicia遗迹组构、Palaeophycus-Planolites遗迹组构、Phycosiphoniform遗迹组构、Thalassinoides遗迹组构、Chondrites遗迹组构、Nereites遗迹组构、Diplocraterion(或aff. Ilmenichnus)遗迹组构(de Gibert and Martinell, 1998; Knaust, 2009; Bednarz and McIlroy, 2009; Callow et al., 2013)。岩心遗迹组构研究虽然取样区域小, 难以观察到完整的遗迹形态, 通常只能鉴定到属, 但具有连续记录的特点, 对识别不同遗迹组构的演替关系以及沉积环境的变迁具有重要价值。目前深海遗迹组构分析主要用于鉴别海底扇不同亚环境, 是海底扇沉积系统储集层质量预测和潜在油气运输屏障识别的重要手段(Knaust, 2009)。
对于深海遗迹组构的深入研究, 有助于加深对深海沉积物浅表地球化学、微生物群落特征、宏体生物群落演替乃至第四纪气候变化的认识。遗迹化石与底质的氧化还原环境具有一定的耦合关系。Wetzel(2002)对中国南海赋存Nereites的现代沉积物研究发现, Nereites通常沿沉积物的氧化还原分界面附近分布, 其造迹生物以该界面附近丰富的微生物为食(图 3-A)。Uchman和Wetzel(1999)在瑞士白垩纪复理石中发现一种螺旋状的、变异形态的Chondrites, 认为是Chondrites的造迹生物占据之前类似Gyrolithes的螺旋状潜穴空间而形成(图 3-B)。这从另一方面证明了深海遗迹组构或阶层的存在。Uchman(2010)在波兰喀尔巴阡山复理石中发现一种放射型遗迹Skolichnus, 其阶层比同一岩层中的Chondrites还要深, 表明其较Chondrites更为适应缺氧环境, 为典型的化学生物迹。Wetzel和Uchman(2013)报道了一个新遗迹种Cladichnus parallelum, 由上部的竖直管和下部平行排列的水平分枝构成, 为中— 深阶层遗迹。该遗迹适应于缺氧环境, 其水平管的多层分布特征可能与底质地球化学性质的变化(氧化还原界面的波动)有关。富氧的管道与其周围缺氧的孔隙水之间存在较高的地球化学梯度, 管道表面微生物活动十分活跃, 可能起到重要的降解有机质的作用, 其本身也是重要的食物来源(图 3-A)。对现代深海沉积物中含Zoophycos遗迹组构进行详细的X射线照相及碳同位素测年, 可探明Zoophycos造迹生物的行为生态属性, 并在恢复第四纪的古气候、海洋沉积环境演变等方面具有重要应用(Lö wemark and Werner, 2001; Lö wemark and Schä fer, 2003; Lö wemark et al., 2004)。然而对于底栖生物与底质地球化学环境的耦合关系目前仍处于初步研究阶段, 其机理仍不甚明确。
遗迹化石的形态功能分析主要探讨遗迹化石的形态发生方式及其生物学目的。Simpson(1956)对Chondrites造迹生物的趋避性进行了详细讨论。Raup和Seilacher(1969)对典型的蛇曲状复理石相遗迹化石进行了计算机模拟, 证明蛇曲状遗迹可由若干简单行为要素来表征。Seilacher(1974, 1977)对复理石相遗迹化石, 特别是雕画迹作了详细的形态功能分析。此前, Seilacher(1962)已对雕画迹的沉积学、埋藏学特征作了详细的讨论, 认识到雕画迹是沉积物浅表发育的空心潜穴, 通常遭受浊流的剥蚀与铸型而保存于浊积砂岩底面。据此, Seilacher(1977)针对不同形态大类(连续蛇曲、单分枝蛇曲、二分枝蛇曲、放射形遗迹、不规则遗迹和规则网状)提出若干基本的造迹模式, 并对这些基本的造迹模式进行微调而得到其演化序列。作者设想的原始空心潜穴系统, 其精细和规则的形态无不使人称奇, 但至今未能得到证实, 仅停留在假说阶段。Seilacher将形态功能分析的思想贯穿于其遗迹学研究中, 对几乎所有遗迹化石都进行了形态功能分析, 这集中体现在Seilacher的遗迹学著作《Trace Fossil Analysis》中(Seilacher, 2007)。计算机模拟方法的运用使得遗迹化石的形态功能分析逐渐向定量化发展(详见2.3.2)。
复杂形态的遗迹化石(如Zoophycos、Paleodictyon)的形态功能分析一直是遗迹学界关注的焦点。形态复杂的遗迹化石代表了特殊、专门的生物行为习性, 常见于沉积环境较为稳定的深海环境。现代化实验技术手段如碳同位素测年、地球化学分析(有机碳、碳酸钙含量测试)的运用, 使得形态功能分析逐步精细化。如Lö wemark和Schä fer(2003)通过对Zoophycos蹼层与邻近沉积物的碳同位素测年以及蹼层化学成分与结构分析, 得出Zoophycos的造迹生物具有食表面沉积物和储存食物的行为习性。
定量遗迹学旨在从遗迹化石形貌的定量刻画与表征的结合上, 量化形态与功能的关系。此外, 定量遗迹学还在沉积学、储集层预测中具有一定应用(详见后文)。
2.3.1 遗迹化石形态定量刻画
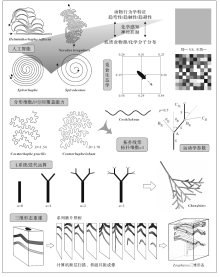
遗迹化石形态定量刻画得益于各种现代数学方法的运用, 包括拓扑几何学、分形几何学以及图论。
Gong和Si(2002)根据拓扑维数(Urysohnic dimension, Du), 将遗迹化石分为4个遗迹目(ichnoorder)。进一步根据非分点数(NP)、欧拉示性数(EC)、点的指数(PI)、闭环数(CL)等参数将其划分为22个遗迹科(ichnofamily)。作者将4种不同类型的遗迹化石(停息迹、爬行迹、被动充填潜穴、主动充填潜穴)分别抽象为0维、1维、2维和3维遗迹。这种拓扑遗迹分类方案具有清晰明了、系统性强的优点。此外, 运用拓扑思想分析遗迹化石形态即忽略遗迹化石长度、角度等度量特征, 舍弃了这些容易受压实、成岩和连续变形作用强烈影响的参数, 符合遗迹分类学属一级的分类原则(Bertling et al., 2006)。但拓扑几何学本质是:允许变形以达到同胚(homeomorphy)即求同的方法, 不适用于对遗迹化石形态的精细刻画(求异)。例如, 无论是螺旋状的Spirorhaphe、Spirodesmos, 还是弯曲和蛇曲状的Cosmorhaphe、Cochlichnus, 均为拓扑线形, 其拓扑维数为1(图 4)。
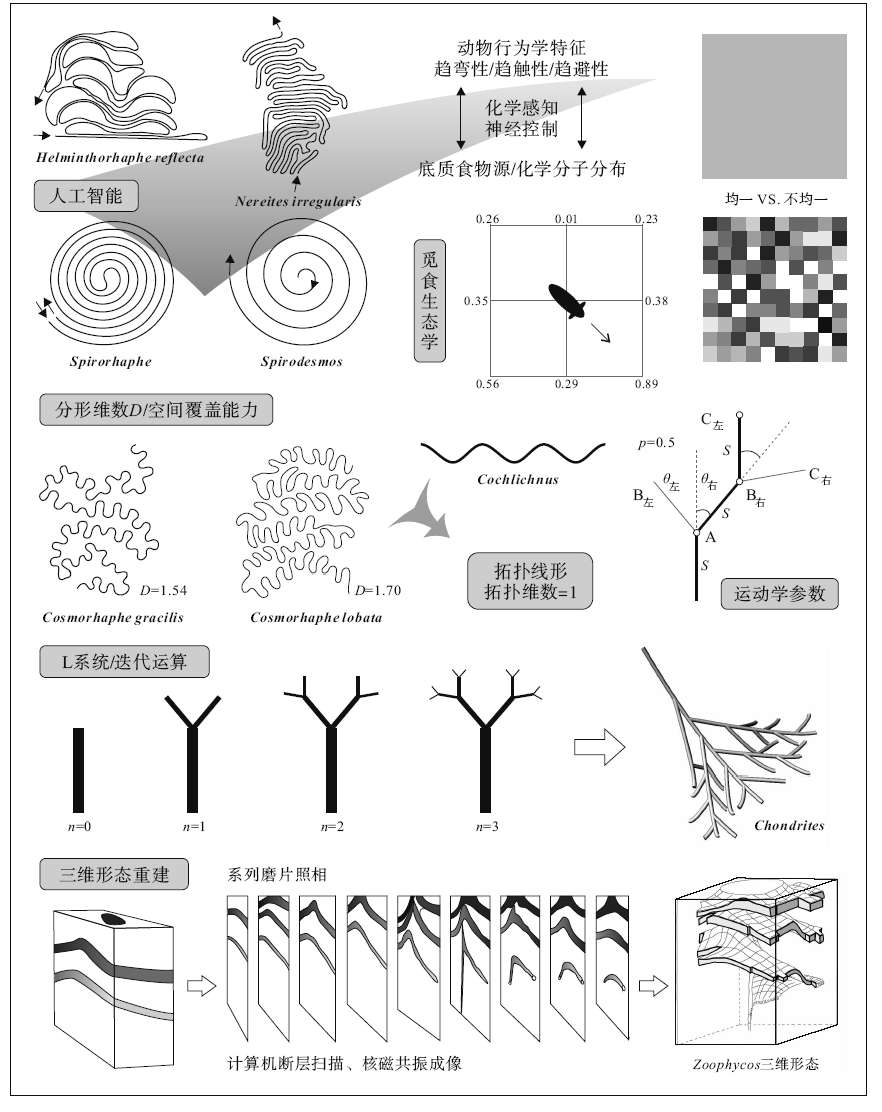 | 图4 定量遗迹学分支概览(参考Raup and Seilacher, 1969; Plotnick, 2003, 2007; Hofmann, 1990; Lö wemark and Schä fer, 2003编制)Fig.4 Sketches showing various branches of quantitative ichnology (compiled from Raup and Seilacher, 1969; Plotnick, 2003, 2007; Hofmann, 1990; Lö wemark and Schä fer, 2003) |
Baucon(2010)首先将分形几何学引入遗迹化石研究中。分形几何学是一门试图描述自然界中复杂现象的几何学分支, 由波兰裔法国数学家Mandelbrot创立(Mandelbrot, 1983)。分形物体的基本特点是具有自相似性和标度不变性, 即跨尺度的对称性(见Mandelbrot, 1989; 陈顒和陈凌, 2005)。分形几何学在物理、化学和地质勘探等领域中已有广泛的应用, 但在化石形态描述与形态功能分析上应用较少(如韩玉英, 1997)。分形几何学中最为重要的概念便是分形维数(简称分维)(fractal dimension), 可用于表征物体占据空间(包括直线、平面和体等)的能力, 分维值越大则对空间的覆盖能力越强。Baucon(2010)对Paleodictyon, Cosmorhaphe, Phymatoderma等遗迹化石进行了分维的计算, 得到结果分别为1.48, 1.56和1.64。Lehane和Ekdale(2013)在此基础之上, 对包括雕画迹在内的更多深海遗迹化石进行了分维计算, 所得Paleodictyon的分维与Baucon(2010)相似, 而Cosmorhaphe的分维则较之要小。虽然Lehane和Ekdale(2013)将雕画迹与觅食迹如Zoophycos的分维进行了比较, 他们并没有给出Zoophycos分维的计算方法。Zoophycos是一种具有三维空间形态的遗迹化石, 其分维显然不能运用计算平面分维的软件实现。运用分形几何学方法刻画遗迹化石形态才刚刚起步。分形维数是衡量遗迹化石空间占据能力的良好参数, 然而这必须基于准确的分维计算。遗迹化石分形维数的计算(如图像处理、置信区间、三维形态遗迹化石分维计算等)的理论基础还需进一步加强。由于分维仅仅是一个数学参数, 对于精细刻画遗迹化石的形态仍显不足。
除了运用拓扑几何学、分形几何学研究遗迹化石的形态, Honeycutt和Plotnick(2005)运用图论的方法对Paleodictyon的形态进行了研究, 并对其造迹可行性进行了讨论, 认为Paleodictyon并非生物潜穴。
2.3.2 遗迹化石三维形态重建
遗迹化石形态三维重建是近年发展起来的一个新方向。化石三维重建通常是基于实际素材的系列磨片(serial grinding)(图 4)。Naruse和Nifuku(2008)通过对发育Phycosiphon的大陆斜坡环境沉积岩进行系列磨片、光学照相, 对Phycosiphon进行了三维重建, 发现Phycosiphon潜穴因受重力作用而发生压扁现象, 其压扁面方向与水平层面的交角大致可以代表大陆斜坡的坡度。Bednarz和McIlroy(2009)也进行了类似的Phycosiphon三维重建工作, 并指出在岩心中识别Phycosiphon是较为困难的, 必须基于详细的三维遗迹化石形态分析, 因而建议在命名遗迹组构时用“ Phycosiphoniform” 代替“ Phycosiphon” 。X射线照相可以加强遗迹化石清晰度, 对岩心遗迹组构的精细化研究具有极大的推动(Lö wemark and Werner, 2001; Lö wemark et al., 2006; Lö wemark, 2012)。Lö wemark和Schä fer(2003)将含Zoophycos岩心进行等距离切片、X射线照相, 获得一系列X射线照片, 据此对Zoophycos进行了三维重建(图 4)。近年来计算机断层扫描(Computed Tomography)、核磁共振成像(MRI, Magnetic Resonance Imaging)等技术在古生物学中广为运用, 使得不破坏样品即可获得一系列样品内部切面图像用于三维重建(如Gingras et al., 2002)。
2.3.3 遗迹化石形态发生模拟
遗迹化石形态发生模拟主要是利用计算机技术模拟遗迹化石形态发生过程。因对生物行为习性的量化方式不同, 所选择的模拟方法也不尽相同。Plotnick(2003)和Baucon(2010)运用L系统(Lindenmayer systems)模拟了生物遗迹的形态发生过程。L系统是一种计算机迭代程序, 最初用于模拟树木分枝的形态发生过程, 具有极高的逼真性(Prusinkiewicz and Lindenmayer, 1990)。在遗迹化石模拟中, 迭代语句则由“ 向前进” (F), “ 向右转” (+), “ 向左转” (-)等运动方式组成, 代表特定的生物行为。由于迭代语句可高度精细化, L系统在模拟复杂遗迹化石的形态发生上具有良好前景(如Chondrites、Paleodictyon)。Plotnick(2007)还运用觅食生态学理论, 模拟生物感知不均一分布食物源而进行运动、取食。Hofmann(1990)模拟了水平曲线形生物遗迹的形态发生过程, 主要参数为一定运动距离S的转动角度θ 。其中S可设为固定值1, 而转动的方向(或左或右)由随机数决定(图 4), 所得模拟轨迹具有随机特征。Raup和Seilacher(1969)深入分析了遗迹化石的行为习性特征, 归纳出蛇曲状生物遗迹的若干形态(行为)控制要素, 即趋弯性(strophotaxis)、趋触性(thigmotaxis)和趋避性(phobotaxis)。这些控制要素的特定调用方式可构成一个造迹模型, 计算机则能将这一模型用数学语言表达并加以物理实现(人工智能)。若所得结果与实际遗迹形态相符合, 则证明这一造迹模型是正确的。作者实现了一些蛇曲状生物遗迹的模拟, 所得结果与实际遗迹形态具有极高的相似性。因此动物遗迹的形态发生过程可以用由若干形态(行为)控制参数构成的程序模拟, 且对于同一程序, 其参数值的微小变化即可造成遗迹种甚至属的变化。
计算机能够将生物行为的基本控制要素以计算机命令的形式作用于假想的“ 造迹生物” 上, 因此计算机模拟可以很好地检验造迹模型的可行性。基本形态(行为)控制要素的确定对进一步分析造迹生物的感知能力具有重要启示。例如, 假如被证实的造迹模式中具有“ 趋触性” 这一形态要素, 则造迹生物具有不断靠近其先成遗迹的趋势。这反映了造迹生物的哪些感知特征呢?也许造迹生物对已扰动沉积物具有一定的感知能力(化学感知chemoreception), 因此可以沿着已扰动的沉积物运动。同样, 对于“ 趋避性” 也可以由化学感知来解释。而“ 趋弯性” 则可能与造迹生物的体长有关。但自Raup和Seilacher(1969)的开创性工作之后, 基本无人步其后尘。一方面Raup和Seilacher(1969)指出, 当遗迹形态复杂到一定程度, 计算机模拟就很难进行甚至无法进行。另一方面可能是遗迹学界对遗迹化石的定量和模拟方面考虑较少。
2.3.4 遗迹组构模拟
遗迹组构模拟主要目的是刻画生物扰动的分布特征, 而与以过程为导向的遗迹化石形态发生模拟相区别。Gingras等(1999)对Glossifungites遗迹相的生物扰动进行了计算机模拟, 并辅以实验测试和野外测量数据, 考察了生物扰动对沉积物渗透率的影响。de Croix等(2012)对Skolithos, Thalassinoides, Planolites, Zoophycos和Phycosiphon进行了计算机形态模拟, 并将其不同组合型式表征不同遗迹相(Skolithos, Cruziana和Zoophycos遗迹相)的生物扰动特征, 从而定量刻画出生物扰动对沉积物孔隙度和渗透率的影响。在地下含水和含油气层中, 沉积物的生物扰动特征对流体的运移和油气质量具有重要影响, 因此遗迹组构模拟有助于对不同环境油气运移、质量的定量判别(Gingras et al., 2002, 2004)。
新遗迹学(neoichnology)是研究现代生物遗迹的遗迹学分支(Hä ntzschel, 1975)。遗迹学根本性的革命是由新遗迹研究推动的— — 无论是Nathorst证明“ fucoids” 的动物遗迹属性, 还是森根堡海洋研究所对现代潮坪生物遗迹的研究, 都为遗迹学成为一门成熟学科奠定了基础。此外, 陆地上深海沉积岩露头的稀少与现代大洋广袤的沉积盆地形成鲜明对比。对现代深海的新遗迹学研究将弥补地层记录的局限性, 并为古代深海遗迹化石研究指明方向(Uchman, 2007; Wetzel, 2010)。早在20世纪60年代, 就有学者指出现代深海生物遗迹与遗迹化石的相似性(Bourne and Heezen, 1965)。Seilacher(1977)看到这项发现, 指出现代海底摄像或深潜器在深海遗迹学研究中的潜在作用。各类深海和大洋钻探计划的实施以及海底摄影的发展极大地推动了深海新遗迹学研究(Ekdale, 1980; Wetzel, 1983; Gaillard, 1991; Bett et al., 1995)。近年来海底摄影、摄像技术快速发展, 图像质量大幅提升。
然而, 目前海底摄像主要用于生物学研究, 专门针对生物遗迹的设计较少。例如Osborn等(2012)对深海半索动物门肠腮纲的分类学进行了论述, 其中从现代海底摄像视频中抓取了许多肠腮动物的照片, 可见肠腮动物与其粪粒共存, 粪粒整体形态与遗迹化石Spirodesmos十分相像(图 5-a, 5-f)。而此文的重点在动物分类学上, 未对这张插图所具有的新遗迹学内涵作详细讨论。目前的海底摄像只能观察沉积物表面的遗迹。如果能够对海底摄像装置进行一定改进, 例如具有喷口, 利用水流或气流将浅表沉积物揭开或利用环氧树脂铸型(Seilacher, 1977), 也许能够观察到更多的不同类型的生物遗迹。深海环境中这方面的实例直到最近才出现(Rona et al., 2009; Seike et al., 2012)。
 | 图5 现代与古代的深海生物遗迹形态对比:具有极高的相似性Fig.5 Remarkable similarities between modern and ancient deep-sea biogenic traces |
遗迹化石概念模型包括形态与行为两个方面(图 6)。利用系列磨片照相或其他非破坏性成像技术以及计算机图形学获得遗迹化石的三维立体形态, 是基于实际素材所得的客观三维形态, 不同于经验推测的概念模型。由于遗迹化石的形态如实反映了造迹生物特定的行为习性, 故客观呈现遗迹化石的形态是一切后续行为习性分析的基础。遗迹化石三维形态重建可用于检验已有的形态概念模型, 是对传统形态学描述的深化和拓展。文献中主要对Phycosiphon、Zoophycos等遗迹化石进行了初步形态重建。但对其他大多数遗迹属及不同遗迹种的系统三维重建工作尚无。因此, 后续工作主要在于系统的遗迹化石三维重建, 并据此深入挖掘其形态功能与生态环境意义。
 | 图6 海相遗迹学研究新方向概念图Fig.6 Conceptual diagram showing new paths of marine ichnology |
通过遗迹化石的形态功能分析, 通常可以设想一个造迹模式, 即行为概念模型。通过计算机形态发生模拟, 可以动态追踪生物遗迹的造迹过程, 从而对行为概念模型进行修正。这是一个从成因、过程入手的研究方法, 具有独特的生物学内涵, 在研究相关生物学控制因素及生物行为的起源、演化方面具有广阔前景。前人主要关注的是二维平面上曲线形遗迹化石的形态发生模拟(Raup and Seilacher, 1969), 而对具有复杂三维形态遗迹化石的模拟较少。因此, 后续工作主要在于复杂遗迹化石(如Zoophycos)的形态发生模拟。
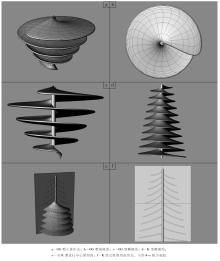
文中对Zoophycos的形态发生模拟进行了初步尝试。Zoophycos分布时间长(寒武纪— 现代), 形态类型多样。笔者从文献中归纳出3种较为典型的Zoophycos形态, 分别称之为OG(Olivero-Gaillard)型、K(Kotake)型和BH(Bromley-Hanken)型。此处的形态类型相当于遗迹化石的“ 形态概念模型” 。虽未进行三维形态重建, 但根据保存完好的遗迹化石与不同切面形态, 其三维形态特征应该是较为准确的。
OG型对应于最为典型的Zoophycos brianteus的形态(图 7-a, 7-b, 7-c)。在层面上表现为近圆形蹼层, 中央为柱形管, 从柱形管向外辐射J形或反J形的大蹼纹, 大蹼纹之间可见与大蹼纹呈角度相交的、近平行排列的略具S形的小蹼纹, 近圆形蹼层的周缘为边缘管。在垂直层面方向, 可见若干蹼层平行排列, 蹼层切面中可见新月形主动充填构造。Olivero和Gaillard(2007)对Z. brianteus的造迹模式进行了细致的讨论, 认为这是一种食泥动物的摄食潜穴。大部分OG型Zoophycos自下而上蹼层直径逐渐增大, 可能反映了造迹生物个体的增大, 由此推测其造迹生物是自下而上建造潜穴的。也有很少的OG型Zoophycos是自上而下建造潜穴的。大蹼纹的绕旋方向可能为右旋或左旋, 两者各占50%。
 | 图7 3种典型的Zoophycos形态Fig.7 Three typical Zoophycos morphologies |
K型Zoophycos与OG型相比, 个体相对较小, 而蹼层数量却大大增加(图 7-e)。OG型通常保存1~3个蹼层, 最多不超过5个, 而K型Zoophycos蹼层数量通常大于10。K型单个蹼层半径在1~3 cm之间, 而OG型蹼层半径通常为10~15 cm, 大者可达90 cm。K型Zoophycos通常自上而下蹼层直径逐渐增加, 被认为是自上而下建造的。K型Zoophycos以Kotake(1991)在日本新近纪地层中发现的标本最为典型, 因此命名为K型。在深海岩心中, 也可看到具有多层结构的K型Zoophycos(Rodrí guez-Tovar et al., 2011)。
BH型Zoophycos的典型特征是近圆形蹼层周围发育不规则舌状构造(图 7-d), 通常每隔若干个正常的类似OG型的大蹼纹就发育一个舌状构造。BH型Zoophycos以Bromley和Hanken(2003)的研究最为细致, 由此命名。BH型的形态具有一定的特殊性, 舌状体分布不规则, 且很可能与底质有机质分布及特殊的化学生物迹生态习性有关, 因此在以造迹生物运动方式为主要模拟对象的形态发生模拟中较难实现。文中主要对前两类Zoophycos进行计算机模拟。
从Zoophycos平行层面和垂直层面的形态可知, Zoophycos是具有柱形管、边缘管的多层蹼状构造。蹼层的水平轮廓为近圆形。在纵切面上显示主动充填形成的蹼状构造, 且相对的围绕柱形管的蹼层在垂向上有一个小的间隔, 而与其他的蹼层相距较远。因此, 笔者认为造迹生物的侧向运动很可能类似于螺旋线, Zoophycos是造迹生物在沉积物的不同深度螺旋线形侧向移动形成的。建模的过程也符合这一造迹模式。首先建立一条螺旋线(即边缘管), 并建立螺旋线的轴线(即柱形管), 这两条线是造迹生物侧向运动的两条“ 路径” 。造迹生物在侧向运动时, 分别以边缘管和柱形管为两端。其次, 需要建立两条曲线, 分别代表造迹生物的初始状态和终止状态。最后我们使用一种形成曲面的工具“ 扫掠” , 以初始状态和终止状态的两条曲线作为断面曲线, 螺旋线和轴线作为路径, 从而生成一个螺旋形曲面。OG型Zoophycos的最下部, 可能代表幼年造迹生物的特殊行为方式, 蹼层较为直立, 在模型中作了相应修改(图 8-c)。运用与OG型相似的建模方式, 模拟了K型Zoophycos。K型的蹼层数可达10, 而且是自上而下建造(图 8-d)。可以看到, 造迹模式中的侧向运动, 就类似于计算机模拟中的“ 扫掠” 。生成的三维模型, 从不同的角度观察及其切面形态都与笔者的“ 形态概念模型” 十分相像(图 8)。这也证明, 前人提出的Zoophycos造迹模式是可行的。因为, 只有在数学上是合理的、在计算机上能实现的对象, 才有可能在现实世界中存在。
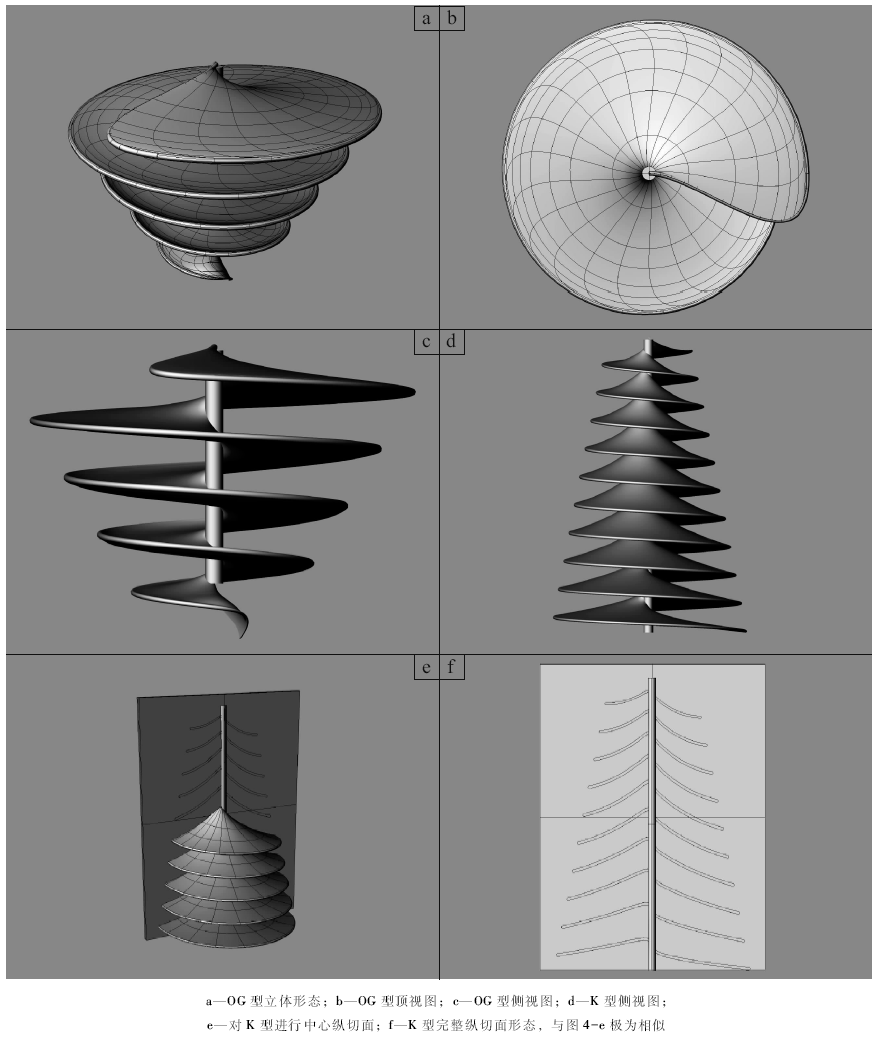 | 图8 典型Zoophycos的形态发生模拟Fig.8 Results of three-dimentional morphological simulation of Zoophycos |
计算机模拟表明OG型与K型Zoophycos的造迹方式是相似的, 均是蠕虫状动物在柱形管四周沉积物中摄食, 并作螺线形侧向移动所形成的多层蹼状构造。有趣的是, OG型与K型的分布时代有所差别, OG型通常分布于前侏罗纪地层中, 而K型则从白垩纪开始才广泛出现(Kotake, 2014)。OG型十分古老, 早古生代其原型就已出现, 在晚古生代则发育完善, 具有典型的柱形管和边缘管, 可见大蹼纹和小蹼纹。OG型与K型的个体大小差别反映了造迹生物类型的差异, 而造迹方向趋向反映了底质有机质分布特征等生态环境差异。但两者形态结构和形态发生上的相似性说明其造迹生物采取了相近的运动方式。
传统的遗迹化石分类鉴定主要基于欧式几何学特征。拓扑几何学、分形几何学都可对遗迹化石形态进行定量刻画, 是遗迹化石分类鉴定的重要辅助手段。综合多种几何学手段、从不同尺度对遗迹化石进行形态描述和刻画是遗迹分类学未来的发展方向之一。拓扑几何学侧重宏观、整体、大类(属以上, 如目、科等)的定量刻画和表征; 欧式几何学侧重属种的定量刻画和表征; 分形几何学则主要针对部分具有自相似性属种的表征以及对遗迹化石空间充填能力的评估(图 6)。
对每一个遗迹属/种, 均有一系列的参数限定其形态特征。首先是拓扑结构, 以欧拉示性数、非分点数、点的指数等拓扑几何学参数表示。虽然拓扑几何学主要用于属以上遗迹化石的大类划分, 但遗迹属内部也有细节拓扑结构的差异性, 可能反映不同的造迹生物或对特殊环境的行为适应。其次是分形几何学参数, 主要包括遗迹化石的自相似性描述与分维计算。例如Cosmorhaphe是典型的具有自相似性的遗迹化石, 具二级蛇曲, 运用分形几何学术语则可描述为:具二级自相似性(或二级标度不变性)(见图4)。分维的计算则对任何遗迹化石均可进行, 特别是牧食迹和觅食迹类, 用于刻画遗迹化石的空间充填能力, 从而对造迹生物的摄食方式、底质有机质分布特征作一窥探。欧式几何学参数主要包括角度、长度(长短、粗细、厚薄)和曲率等, 主要用于属种级别的定量描述, 可作为表征遗迹化石个体发育(针对其造迹生物)、形态演化的基本参数。运用以上几何学方法的前提是对遗迹化石完整欧式几何学形态的掌握(通过三维形态重建)。
前文已对Zoophycos的三维形态和造迹模式作了较为详细的讨论, 在此基础上可对其几何形态进行综合描述。Zoophycos具有在造迹过程中保持中空的柱形管和边缘管以及主动充填形成的蹼层(实心), 且在完整的Zoophycos中蹼层为一螺旋面体。因此Zoophycos的拓扑结构是由3部分组成的:边缘管和柱形管可分别简化为一螺旋线和该螺旋线的轴线, 而蹼层可看作一螺旋面。Zoophycos具有三维立体形态, 对其分形维数的计算需要建立在三维重建基础之上, 目前还难以进行。欧式几何学参数如蹼层直径、数量及与水平面的夹角、柱形管与边缘管的大小等度量均可作为属种层次精细研究的素材。
微观手段主要是从元素或同位素、分子、微生物等不同微观尺度着手, 运用先进实验技术对遗迹化石进行测试分析, 对于精细刻画生物行为习性及其与环境的耦合关系具有重要作用。前文已述, Lö wemark和Schä fer(2003)曾对Zoophycos的蹼层以及蹼层外沉积物分别进行14C定年以及有机碳、碳酸钙含量测定, 确定二者新老关系和物质成分特征, 据此对Zoophycos的造迹模式、行为习性加以讨论。遗迹化石中的生物大分子研究, 单一实例为Rona等(2009)对含Paleodictyon沉积物DNA测序, 鉴定其网格状结构中物质来源为深海底栖有孔虫, 进而对Paleodictyon的属性进行了详细的讨论。Gong等(2007, 2008)通过扫描电镜观察和生物标志化合物测试发现了Zoophycos蹼层中的微生物化石和分子化石, 是将地球生物学方法运用到遗迹化石中的典型实例。Seilacher(1977)提出了雕画迹作为深海动物诱捕和培植微生物的“ 花园” 行为习性模式。Seilacher(1990)指出某些遗迹化石(如Chondrites)的造迹生物具有与微生物化学共栖(chemosymbiosis)的生理行为特征(图 3-A)。Bromley和Hanken(2003)对希腊罗德岛深海沉积岩中发现的一种大型叶片状Zoophycos进行了形态功能分析, 也指出微生物可能与造迹生物有化学共栖关系。从理论上说, 微生物占了生物界生物量的大部分, 无处不在, 其与宏体生物之间必定存在一定的相互依存关系, 但在地质历史时期较少提及。遗迹化石为研究微生物与宏体生物的相互作用打开了一扇窗。微生物不仅作为共生体存在, 其本身也是重要的造迹生物。微生物钻孔与环境因素(如光、温度、盐度、无机营养等)密切相关, 可作为环境指示标志(Glaub et al., 2007)。微观手段能够提供大量前所未有的信息, 使得不可见之物逐渐走入科学家的视野。微观手段所获得的信息通常定量性较好, 且从食物链和生态系的底层着眼, 对诠释生物与生物以及生物与环境之间的相互作用和协同演化具有重要价值。
生物与环境相互作用是通过无机环境、微生物以及宏体生物等不同物质层次实现的。这三者相互牵连, 其中一个发生变化, 必将在其他方面留下踪迹。遗迹化石(针对宏体遗迹化石)主要反映了生物的运动行为和生理行为方式。运动行为方式主要受神经系统控制, 而神经系统的起源和发育与环境密切相关。趋弯性、趋触性和趋避性等运动行为特性与造迹生物的感知能力具有密切的联系(见2.3.3)。生物能够感知无机化学分子的浓度以及生物间传递的信息素。深海遗迹形态复杂而规则, 其造迹生物的感知方式是怎样的?信息素能否在地质记录中保存下来, 从而对生物的化学感知和神经系统控制作用作一窥探?生理行为方式则是环境与生物发生相互作用的渠道, 适应环境的生理行为方式会被保留, 不适宜环境的生理行为方式会被自然淘汰, 从而促使生物行为、形态、神经系统与环境的协同演化。宏体生物与微生物的共生必定是适应环境的结果, 其原因及其对环境的影响又是怎样的?这些都是当今和未来值得探索的新方向。
海相无脊椎动物遗迹学的研究历史久远, 然而很长一段时间内其真实属性未知。对现代生物遗迹的观察促使遗迹学的革新, 使得生物遗迹或遗迹化石真正成为具有清晰物理边界和严格科学定义的研究对象。继Seilacher创立遗迹相并对遗迹化石进行系统的形态功能分析, 遗迹学扬起了新时代的风帆。20世纪后半叶以来, 遗迹学在沉积学中得到了极为广泛的应用, 已成为古环境、古生态分析和油气勘探领域不可或缺的一部分。同时, 针对遗迹化石本身, 其命名分类及形态功能分析亦有诸多进展。各种现代化分析测试手段(地球化学, 地球生物学、分子生物学方法等)以及定量方法(如拓扑几何学、分形几何学以及三维形态重建、计算机形态模拟等)的发展使遗迹学逐渐向一门交叉学科和成熟学科迈进。遗迹化石作为古代生物— 环境相互作用的重要记录, 必将在地球生物学和深时全球变化研究中扮演重要的角色— — 这也是遗迹学的终极追求。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|
| 99 |
|
| 100 |
|