

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
巨鲕的微生物成因:来自重庆石柱地区下寒武统的证据
[段雄, 时志强 , 金鑫, 范鸿, 姬国锋]
, 金鑫, 范鸿, 姬国锋]
, 金鑫, 范鸿, 姬国锋]
|
|

通讯作者简介 时志强,男,1972年生,教授,主要从事沉积学教学和研究。E-mail:Szqcdut@163.com。
第一作者简介 段雄,男,1988年生,硕士研究生,沉积学专业。
巨鲕的产出时代多见于新元古代和早三叠世,而寒武纪的巨鲕却鲜见报道。近期,笔者在重庆石柱地区下寒武统天河板组中发现厚约35 cm的透镜状巨鲕灰岩。在该层巨鲕灰岩中,巨鲕粒径一般为5~7 mm,最大可达9 mm,约占岩石组分的55%,与大量强烈重结晶的正常鲕粒伴生。钙质生物壳体较常见,颗粒间见亮晶方解石胶结物。研究区巨鲕最外圈纹层上常见蓝细菌化石,形成发育不完全的突起状“夭折的纹层”;巨鲕内的暗色纹层中亦见杂乱交错的、有一定重结晶的蓝细菌等微生物化石。故推测其为微生物成因形成的,且早寒武世频繁的强风暴背景和蓝细菌等微生物的积极参与是该层巨鲕形成的必备条件。
About the corresponding author Shi Zhiqiang,born in 1972, is a professor in Chengdu University of Technology. Now he is engaged in sedimentology. E-mail:Szqcdut@163.com.
About the first author Duan Xiong,born in 1988,is a master degree candidate in Chengdu University of Technology. Now he majors in sedimentology.
Giant ooids are commonly seen in the Precambrian and Lower Triassic,while they are hardly reported in the Cambrian. Recently,we found giant ooid limestone with about 35 cm thick in the Lower Cambrian Tianheban Formation at the Shiliu section in Shizhu area,Chongqing. The diameter of these giant ooids is generally 5-7 mm,with a maximum diameter of 9 mm. The giant ooids make up 55% of rock components. Meanwhile,the recrystallized normal ooids and calcareous shells are relatively common in the limestone. The cements of sparry calcites can be found between the grains. Abundant Cyanobacteria fossils found in the outermost laminae of a giant ooid,form a sort of protruberance-shaped laminae named as “abortive laminae”,with which the microbial cause of giant ooids is clearly revealed. The recrystallized inner dark laminae of giant ooids similarly contain messy girvanell fossils. So it is thought that the active involvement of microorganisms and frequent strong storms are the necessary conditions for the formation of giant ooids.
巨鲕(giant ooids)这一术语最早由Sumner和Grotzinger(1993)正式提出, 其为海相成因, 结构形态上类似于正常鲕粒, 但粒径大于2 mm, 甚至达厘米级。虽然巨型鲕粒一直吸引着广大地质工作者进行研究, 但对其细致的成因研究仍较为薄弱。巨鲕多集中于新元古代海洋中(Trower and Grotzinger, 2010), 常与正常鲕粒共生, 在中国华南地区早三叠世沉积中也见报道(梅冥相, 2008, 2012; 李飞等, 2010), 但对寒武纪巨鲕却鲜有报道(代明月等, 2014)。巨鲕的成因机理以及产出环境较为复杂:Sumner和Grotzinger(1993)对前寒武纪巨鲕的研究表明, 低速率的核心供给、高速率的生长及高强度的水体搅动有利于巨鲕的形成; Woods(2014)对早三叠世古海洋条件分析后指出, 强烈的水体流动和CO2释放激发的波浪是巨鲕形成的必备条件; Lehemann等(2012)根据对华南南盘江盆地巨鲕的研究指出, 水体化学性质变化而表现出来的高Ca/Mg值是形成巨鲕的最根本因素; 梅冥相(2012)在湖北利川下三叠统大冶组也发现了具有“ 生物活力” 的类似叠层石结构的巨鲕, 暗示巨鲕的形成可能与微生物活动有关。
笔者在重庆石柱野外工作时发现, 在石流剖面下寒武统天河板组近顶部沉积了一套厚约35 cm的透镜状巨鲕灰岩, 该套巨鲕灰岩总体上重结晶作用及白云化作用较弱, 镜下可见清楚的纹层, 且在不规则的暗层中见丰富的蓝细菌类微生物化石, 这为了解早寒武世巨鲕的成因提供了可靠的依据。
早寒武世构造活动及初期的大规模海侵作用造成了上扬子地区震旦纪碳酸盐岩台地的消亡(王长生和龚黎明, 1998), 之后的碎屑物质填平补齐作用使得重庆石柱地区沉积了以陆源沉积为主的下寒武统水井坨组、石牌组。石牌组之上沉积的是天河板组, 岩性主要为灰岩和页岩, 并可进一步分为2段。
本次研究的石流剖面位于重庆石柱县老厂坪背斜东翼龙泉溪一带, 地理位置为北纬29° 45'48.41″、东经108° 18'11.18″(图 1)。该剖面中天河板组一段厚约72 m, 岩性主要为灰色至浅灰色生物灰岩、深灰色厚层条带状含泥质灰岩。天河板组二段以黄绿色、灰绿色页岩为主, 上部夹灰色泥质灰岩及巨鲕灰岩、鲕粒灰岩, 顶部为一套厚约1.5 m的粉砂质页岩, 与上覆石龙洞组呈整合接触(图 2)。巨鲕灰岩透镜体的产出层位为天河板组和石龙洞组界线之下约2 m处, 在该层岩石中可见粒径大小不一的巨鲕颗粒(图 2-A, 2-B)。巨鲕灰岩之中夹厚约10 cm的泥质灰岩, 之下为厚约10 cm的微晶灰岩和20 cm的鲕粒灰岩, 之上为厚约30 cm的纹层状微晶灰岩(图 2)。
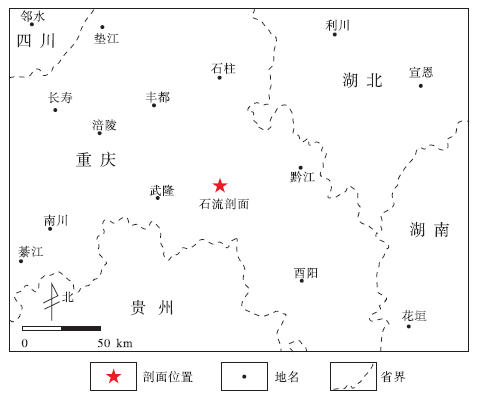 | 图1 重庆石柱石流剖面地理位置Fig.1 Location of Shiliu section in Shizhu area, Chongqing |
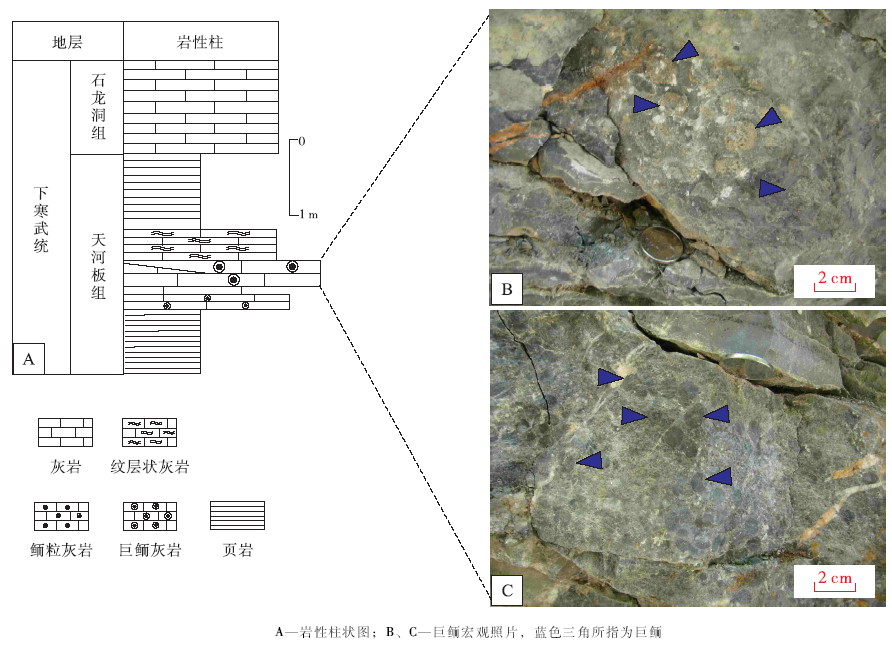 | 图2 重庆石柱石流剖面下寒武统上部岩性柱状图及巨鲕照片Fig.2 Lithological column and photographs of giant ooids of the upper part of Lower Cambrian at Shiliu section in Shizhu area, Chongqing |
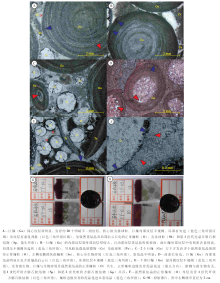
在石柱石流剖面, 下寒武统天河板组近顶部巨鲕灰岩中的巨鲕, 粒径最大可达9 mm, 大部分集中在5~7 mm。从手标本上看, 巨鲕约占岩石组分的50%~55%(图 3-G, 3-H), 多呈深灰色(图 2-B), 少量被铁质交代而呈浅褐色(图 2-A)。巨鲕灰岩层呈透镜状, 最厚为35 cm, 横向延伸可达5 m。
 | 图3 重庆石柱石流剖面巨鲕及相关组分特征Fig.3 Photographs showing characteristics of giant ooids and related composition at Shiliu section in Shizhu area, Chongqing |
显微镜下观察巨鲕时, 可见规则的、明暗相间的同心纹层, 且暗层明显厚于亮层。同心纹层数量不等, 最多可达20余层, 局部发生紊乱而呈现为突起状, 且此现象在巨鲕的外层较为普遍(图 3-A, 3-B), 推测与微生物活动有关。巨鲕核心可以分为2类:一类核心是生物碎屑, 如棘皮类等生物碎屑, 但具有这种核心的巨鲕在岩石中含量不高; 另一类核心是由微生物作用形成的藻球粒, 粒径2~4 mm, 是形成巨鲕的主要核心, 后期的重结晶和交代作用使巨鲕核部轮廓和内部结构模糊不清, 其主要成分为微晶方解石。巨鲕整体均发生不同程度的重结晶化, 以靠近核心处较为严重, 向巨鲕边缘逐渐变弱, 部分巨鲕内部可见较多的白云石晶体(图 3-D)。
巨鲕灰岩中常见正常鲕粒, 约占碳酸盐颗粒的25%左右, 粒径呈双峰态分布, 较大者约为1.5~1.8 mm, 较小者为0.6~0.9 mm(图 3-E)。所有鲕粒均为先被溶解再被重结晶方解石充填形成的铸型鲕, 其内部纹层不可见, 但圆形的外部轮廓非常明显。部分鲕粒发生破碎, 可见较多的微晶白云石(图 3-C)。鲕粒边缘常见泥晶化边(图 3-C, 3-E)。巨鲕灰岩中亦见大量的三叶虫化石碎屑、球粒、似球粒以及椭圆状似鲕粒与巨鲕共生(图 3-C, 3-E)。椭圆状似鲕粒(图 3-C)大小约为1, mm× 2, mm, 重结晶弱, 核心为生物化石, 外部圈层不规则, 见微生物, 应与巨鲕成因类似。
所有颗粒间均发育灰泥杂基, 而灰泥杂基一般受到微生物的改造, 局部色暗, 部分重结晶(图 3-A, 3-B)。巨鲕灰岩中的胶结物可分为2期:第1期为刀刃状亮晶方解石, 大致沿颗粒边缘垂直分布(图 3-F); 第2期为刀刃状方解石胶结物与灰泥杂基间的粒状亮晶方解石。
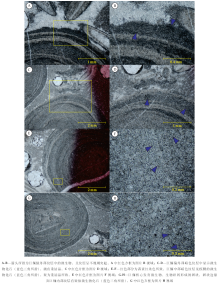
在天河板组巨鲕中, 一般在暗色纹层中见微生物化石, 在多个巨鲕的最外部纹层多发育不规则突起状暗色层(图 3-A, 3-B, 3-E; 图4-A), 放大观察可知其为蓝细菌类, 微生物特征明显, 呈管束状、弯曲状交织在一起(图 4-B), 管体之间被灰泥填充, 管体长度一般为150~180 μ m、直径一般为7~10 μ m。笔者认为巨鲕最外缘和次外缘的由蓝细菌组成的不规则突起状暗色层是一种“ 夭折的纹层” , 越往核心暗色纹层相对而言保存的越比较完整, 为富有机质暗色层的初始状态, 或为风暴轻微改造富微生物纹层后的特征, 也不排除巨鲕沉积后蓝细菌在巨鲕边缘生长的可能性。
 | 图4 重庆石柱石流剖面巨鲕边缘及内部微生物微观特征Fig.4 Microphotographs showing microorganisms of giant ooids at Shiliu section in Shizhu area, Chongqing |
巨鲕内部暗色层中的微生物一般发生重结晶作用, 致其形态特征模糊, 很难辨认(图 4-C, 4-D, 4-E, 4-F, 4-G, 4-H), 总体上呈现越向核部化石重结晶越强烈的特征。在巨鲕内部, 微生物化石排列无定向性, 不易辨认。在较外部的纹层内, 管束状蓝细菌类微生物集合体的特征依稀可辨(图 4-C, 4-D), 且巨鲕的多数突起状不规则纹层基本都与蓝细菌有关(图 4-C, 4-D, 4-E, 4-F), 但在更靠近巨鲕内层时, 微生物化石重结晶较为强烈, 能模糊分辨出重结晶后呈蠕虫状的微生物集合体(图 4-E, 4-F, 4-G, 4-H)。从巨鲕含藻暗色纹层重结晶的程度可以看出, 发生重结晶的时间顺序应该是由内而外, 在巨鲕形成过程中即发生着钙化作用, 这可能和当时碳酸钙过饱和的古海洋条件有关; 巨鲕内部和外部的蓝细菌类微生物具有不同的生长特征, 内部纹层的管束状集合体的特征不明显, 显示其可能为不同种属的微生物。
早寒武世, 上扬子地区为典型的碳酸盐缓坡沉积环境(王剑, 1990; 蒲心纯等, 1993), 位于南半球低纬度地区(南纬约17° )(刘育燕等, 1993; 殷鸿福等, 1999), 而对现代大西洋海域的研究发现, 现代风暴只发生在赤道附近5° ~45° 之间(Marsaglia and Klein, 1983), 因此研究区在早寒武世位于风暴作用区。另外, 根据Berner(2006)的显生宙以来全球大气中氧气和二氧化碳浓度模式, 早寒武世晚期(距今约520 Ma)大气中的CO2浓度已达到整个寒武纪期间的峰值, 而大气中高CO2浓度造成的全球温度升高不仅可导致全球海平面的升高, 而且海洋上温暖的水蒸气提供的强大能量也是催生频繁风暴的重要“ 元凶” , 故早寒武世晚期大气中过高的CO2浓度为研究区频繁的风暴作用发生提供了有利条件。同时, 研究区的碳酸盐浅水缓坡有助于风暴盛行, 频繁的风暴作用致使研究区附近的下寒武统碳酸盐岩中风暴砾屑层广泛分布, 如金振奎等(1995)在黔东、湘西等地均发现了下寒武统风暴沉积; 马志鑫等(2012)也有黔东镇远地区下寒武统中发现风暴沉积的报道。综上可知:研究区早寒武世晚期碳酸盐缓坡沉积背景以及大气中过高的CO2浓度为频繁的风暴提供了有利条件。
巨鲕的成因机理以及其产出环境较为复杂, 如Sumner和Grotzinger(1993)认为前寒武纪巨鲕的形成条件是低速率的核心供给、高速率的生长、高强度的水体搅动等; Woods(2014)认为强烈的水体流动和CO2释放激发的波浪是早三叠世巨鲕形成的必备条件; 梅冥相(2012)根据湖北利川下三叠统大冶组巨鲕有类似叠层石结构, 暗示其可能与微生物活动有关。笔者对石柱石流剖面天河板组发现的巨鲕研究证实, 微生物与巨鲕形成存在必然的关系(图 4)。在综合考虑微生物作用、正常水体能量以及强风暴等因素的情况下, 笔者提出了石柱石流剖面天河板组巨鲕发育的模式(图 5)。
 | 图5 重庆石柱石流剖面下寒武统巨鲕演化示意图Fig.5 Sketch map showing evolution of giant ooids of the Lower Cambrian at Shiliu section in Shizhu area, Chongqing |
首先, 在巨鲕发育的初期, 微生物会依附在异地搬运过来的生物碎屑或者藻球粒上, 并以此为核心生长(图 4-G)。生物碎屑或者藻球粒里的矿物组分可以为微生物的生长发育提供充足的营养成分。镜下未能发现以正常鲕粒为核心的巨鲕, 而从微生物的发育特征(巨鲕及填隙物中均有微生物发育)分析, 鲕粒与巨鲕是不同环境的产物, 鲕粒是异地形成后与生物碎屑一起被搬运来, 然后与巨鲕沉积在同一地方。其次, 微生物在其生长发育的过程中, 在风暴期或者高潮期会不断捕获被风暴水流或潮汐水流带来的碳酸盐颗粒, 比如灰泥(Liu and Zhang, 2012)。此外, 微生物能分泌一种细胞外多糖物质, 这种细胞外多糖物质也能捕获周围的微晶碳酸盐颗粒(Liu and Zhang, 2012)。微生物和这些捕获的灰泥或微晶颗粒交织在一起组成了巨鲕的暗色纹层。再次, 当时大气中CO2浓度过高, 加之频繁的风暴作用会致使海水CO2过饱和, 这为微生物的光合作用消耗了周围水体中大量的CO2提供了有利条件, 而且微生物的光合作用还会导致水体pH升高, 从而在碱性的海水环境下造成了微生物表面高速率的CaCO3快速沉淀(许靖华, 1980)。CaCO3的高速沉淀, 会在暗色纹层的外部形成一层以亮晶方解石为主的亮色纹层。如此反复, 随着巨鲕的不断生长, 明暗相间的纹层数量也会不断增多。
需要指出的是, 以生物碎屑和藻球粒这两种核心发育的巨鲕出现明暗纹层的顺序不同, 前者先发育亮层, 而后者先发育暗层。出现这种现象的原因在于微生物在依附在生物碎屑的过程中并未形成同心状纹层, 但是此时周围水体的化学性质已经改变, 这时便会先沉积CaCO3形成亮层, 而以藻球粒为核心的巨鲕是直接沉积一圈暗色纹层。当巨鲕发育到一定程度, 正常的水体能量已达到不能卷起并搅动其的临界值, 这时巨鲕会沉积到海底, 但是微生物的活动并未停止, 而是在巨鲕表面继续进行活动, 即在风暴平静时期, 巨鲕在微生物的活动下发育形成完整的暗色纹层, 而强风暴的来临会提供一个超越正常水体能量临界值的水体环境, 这样巨鲕被卷起、搅动, 使得巨鲕表面含微生物的暗色纹层会由于颗粒之间的相互碰撞而发生磨损, 以致巨鲕最外部的暗色纹层残缺不全, 即形成所述的“ 夭折的纹层” ; 强风暴盛行时期, “ 夭折的纹层” 将被进一步磨损而形成外圈的暗层(图 4-C, 4-D); 具有“ 夭折的纹层” 的巨鲕如果遭遇环境水体能量变低、风暴不再影响, 则可沉积保存下来。
由此可见, 巨鲕的形成与3个方面有着必然的联系:频繁的风暴作用提供的较强水动力条件; 海洋中CO2浓度过饱和的海水化学性质; 微生物的生理作用对巨鲕圈层的直接以及间接作用。同时, 这3个方面也并不是孤立存在的, 都是相互影响相互作用才导致巨鲕的最终形成。
重庆石柱地区天河板组近顶部发现的巨鲕灰岩, 对研究巨鲕的成因机理有重要意义:巨鲕的核心可以是生物碎屑, 也可能是藻球粒, 在不同的核心的基础上发育的巨鲕出现明暗纹层的顺序不同。巨鲕是在缓坡环境下受到微生物作用而形成的, 并受到强风暴的影响, 故巨鲕的形成是微生物的积极参与、极端的古气候背景和有利的古地理条件综合作用的结果。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|