






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
安徽南部下、中奥陶统紫台组碳酸盐岩微相与海平面变化
[栾晓聪1, 2  , 刘建波
, 刘建波3 , 詹仁斌1 , 吴荣昌1 ]
, 刘建波|
|

第一作者简介 栾晓聪,男,1990年生,2013年7月本科毕业于南京大学地球科学与工程学院,现为中国科学院南京地质古生物研究所硕士研究生,研究方向为华南奥陶纪碳酸盐岩沉积学。E-mail: luan1990@126.com。
下、中奥陶统紫台组形成于华南奥陶纪生物大辐射的关键时期,其中蕴藏了一些与大辐射有关的重要信息。笔者通过对安徽南部石台丁香剖面、池州洪家剖面的紫台组进行野外实测、采样、室内切片和镜下观察与分析,共识别出9个碳酸盐岩微相,即:MF-1钙质泥岩、MF-2泥质泥晶灰岩、MF-3泥晶灰岩、MF-4腹足类—介形虫生屑泥晶灰岩、MF-5海绵骨针含生屑泥晶灰岩、MF-6含生屑泥质泥晶灰岩、MF-7生屑泥质泥晶灰岩、MF-8生屑泥晶灰岩和MF-9泥晶生屑灰岩。这些微相主要形成于深潮下带及相邻环境中。研究表明:皖南紫台组沉积时期水深波动幅度不大,且自早至晚沉积水深逐渐加大;丁香剖面由于更接近台地边缘,总体沉积水深要大于洪家剖面。在2条剖面的紫台组中识别出至少5次比较显著的海平面上升事件,它们与上扬子区海侵事件具有良好的对应关系,并可以与澳大利亚、北欧、北美、华北等板块进行对比。由此推断,区域性相对海平面变化、甚至全球性海平面变化控制着下扬子区下、中奥陶统紫台组的沉积及其变化。
About the first author Luan Xiaocong,born in 1990,got a bachelor degree in Nanjing University in June 2013,and is currently a master degree candidate at Nanjing Institute of Geology and Palaeontology,Chinese Academy of Sciences. Now he majors in carbonate sedimentology of the Ordovician in South China. E-mail: luan1990@126.com.
The Lower to Middle Ordovician Zitai Formation,a unique lithologic unit formed during the critical period of the Great Ordovician Biodiversification Event(GOBE),contains some important information about this major biotic event. Based on detailed investigation and sampling in the field and observation under the microscope in the lab,nine carbonate microfacies were distinguished in the Zitai Formation at Dingxiang section of Shitai County and Hongjia section of Chizhou County,southern Anhui Province,East China,named as the MF-1 calcareous shale,the MF-2 argillaceous mudstone,the MF-3 mudstone,the MF-4 gastropod-ostracod wackestone,the MF-5 calthrop-bearing wackestone,the MF-6 argillaceous bioclast-bearing wackestone,the MF-7 argillaceous bioclastic wackestone,the MF-8 bioclastic wackestone and the MF-9 bioclastic packstone. These microfacies were deposited in deep subtidal and adjacent zones with low amplitude of changes in water depths. Taking into account of some other evidences that we observed in the field,we proposed that depositional environment of the Zitai Formation at Dingxiang section was deeper than that at Hongjia section. Five transgressive events in the Zitai Formation were recognized at both sections. All these transgressions can be precisely correlated between the studied sections and the sections on the Yangtze Platform,as well as on some other palaeoplates. Thus the changes in deposition of the Zitai Formation in the Lower Yangtze region were controlled by regional and even global eustatic sea-level fluctuations.
奥陶纪生物辐射(Ordovician Radiation)(Sepkoski, 1979, 1981)指的是发生在奥陶纪特别是早、中奥陶世的海洋生物多样性急速增加的生物演化事件, 后被称为奥陶纪生物多样化事件(Great Ordovician Biodiversification Event; Webby, 2004)。它与寒武纪生命大爆发最重要的区别在于奥陶纪生物辐射主要表现在目及其以下较低级别的分类阶元上。与国际研究基本同步, 华南奥陶纪生物大辐射的研究已有10余年的历史, 所取得的阶段成果已经引起国际同行的广泛关注(Zhan et al., 2005, 2006; 周志毅等, 2006; Zhang et al., 2007; Liu and Zhan, 2009; Zhang et al., 2010; Yan et al., 2011; Wu et al., 2012), 这些工作所涉及的研究区域主要集中在华南扬子台地、特别是上扬子台地中部。
在奥陶纪生物大辐射第1次峰值前后, 扬子台地边缘地区广泛发育有一套特殊岩相的地层, 即紫台组(中国科学院南京地质古生物研究所, 1974)。紫台组以紫红色瘤状泥质灰岩为特征, 从上扬子区到下扬子区东西向延伸数千千米(吴荣昌等, 2007), 与在上扬子区广泛分布的大湾组和湄潭组大致同期, 形成于早奥陶世弗洛晚期到中奥陶世达瑞威尔早期(Chen et al., 1995; 汪啸风等, 1996; 董卫平, 1997; 高振家等, 2000; 张允白等, 2002; 陈朋飞和詹仁斌, 2006)。与湄潭组和大湾组相比, 有关紫台组的古生物学、地层学和沉积学研究至今仍比较薄弱。紫台组中壳相化石丰富, 对其中的头足动物鹦鹉螺和节肢动物三叶虫已有一些专门研究(如朱忠德等, 1995; 张允白, 2003; Turvey and Zhou, 2004; Zhou et al., 2011), 牙形刺化石研究亦取得一些成果(安太庠, 1987; 吴荣昌等, 2007; 吴荣昌和王志浩, 2008; Wu et al., 2012), 但对于奥陶纪生物大辐射在这一特殊古地理区域的表现尚缺乏系统分析(吴荣昌等, 2007)。有关紫台组沉积学的研究散见于相关奥陶系地层学、岩相古地理(周名魁等, 1993; 冯增昭等, 2001)、层序地层学与海平面变化(汪啸风等, 1996; 苏文博, 2001)的讨论中, 而对于紫台组的沉积学特征、海平面变化研究和沉积环境恢复尚未系统开展, 对于其中紫红色沉积的成因尚缺少专门探讨(胡修棉等, 2006)。
在良好的生物地层学框架内, 本研究对安徽南部池州和石台的紫台组进行了细致的野外和室内沉积学研究(图 1), 采用定量微相分析方法探讨紫台组的沉积环境及其变化规律, 恢复高频海平面变化曲线, 并与扬子区其他地区和其他古板块海平面变化事件进行对比, 初步探讨紫台组沉积的成因。本研究可为进一步深化奥陶纪生物大辐射研究、特别是在海洋生物多样性演变与古环境变迁的协同关系探讨方面提供重要的科学依据。
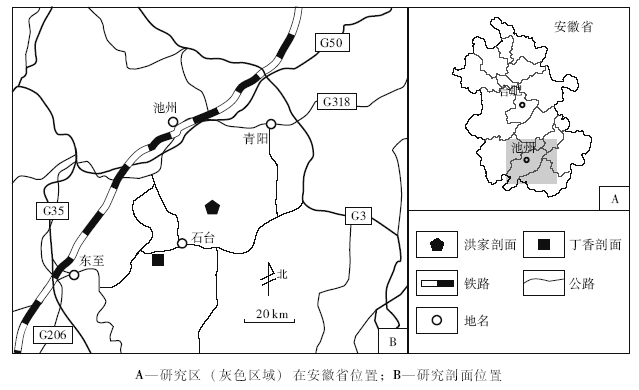 | 图1 安徽南部石台丁香剖面和池州洪家剖面位置Fig.1 Location of Dingxiang and Hongjia sections in Shitai and Chizhou respectively, southern Anhui Province |
早、中奥陶世, 华南板块处于较低纬度地区, 是冈瓦纳超级大陆边缘的一个地体(陈旭等, 2001)。当时的华南板块包括扬子台地、江南斜坡和珠江盆地3个部分(Chen et al., 1995; 冯增昭等, 2001)。扬子台地的西缘和西北缘(现今的定向)发育有一系列古陆(张允白等, 2002), 从西北往东南存在一个非常清晰的陆— 台— 台缘— 坡— 盆的古地理格局(图 2), 在华南广大地区发育的奥陶系也基本上呈现出南西— 北东向的带状分布(陈朋飞和詹仁斌, 2006), 且岩相、生物相具有明显的规律性。
 | 图2 扬子台地紫台组分布及厚度对比(古地理图修改自陈朋飞和詹仁斌, 2006; 吴荣昌等, 2007)Fig.2 Correlation of thickness and distribution of the Zitai Formation on Yangtze Platform (palaeogeographic map modified from Chen and Zhan, 2006; Wu et al., 2007) |
在早奥陶世晚期至中奥陶世早期, 扬子台地的不同区域分别发育有紫台组、大湾组、湄潭组等不同的岩组, 但产出相似的笔石和壳相动物群, 属同期异相沉积。紫台组由穆恩之等于1974年创立, 命名地点在贵州印江合水镇紫台村(GPS:N28° 03'09″, E108° 33'57.2″)。紫台组典型岩性可分为3部分, 下部为土黄色、黄绿色夹紫红色的泥页岩, 中部为紫红色瘤状泥质灰岩, 上部为土黄色砂岩, 且以中部的紫红色、灰绿色瘤状泥质灰岩为特色(吴荣昌等, 2007)。紫台组地理分布广泛, 在黔东、黔东北、渝东南、湘西北、鄂西南、鄂南、鄂东南、赣北及皖南等地都有发育(图 2)。下扬子区的紫台组厚度较上扬子区小, 且以碳酸盐岩为主, 未见紫台组上部典型的砂岩、粉砂岩沉积(图 2)。
紫台组通常化石丰富, 类群多样, 包括三叶虫、棘皮类、鹦鹉螺、腕足类、苔藓虫、牙形刺等。如湖北松滋刘家场张山堰剖面的紫台组下部, 就产有三叶虫Megalaspides、Illaernus、Nileus、Eucalymene、Carolinites等; 腕足类Sinorthis、Nereidella、Martellia、Yangtzeella等(吴荣昌等, 2007)。紫台组中牙形刺化石也非常丰富, 除下伏红花园组的延续分子外, 还出现新生类群, 如Oepikodus evae等(安太庠, 1987; 朱忠德等, 1995; 吴荣昌和王志浩, 2008)。紫台组中的鹦鹉螺化石比其同期大湾组更加丰富, 如紫台组中Bathmoceras的种级分异度较大湾组高(张允白, 2003)。此外, 紫台组底部产出少量笔石化石(穆恩之等, 1979)。
有关紫台组年代地层学的工作尚需进一步加强。据已有资料, 扬子区紫台组顶、底界应普遍存在穿时现象(吴荣昌等, 2007), 其底界往往较湄潭组和大湾组底界高, 可位于下奥陶统弗洛阶中上部的Didymograptus(Corymbograptus)deflexus笔石带内(穆恩之等, 1979), 或者Oepikodus evae牙形刺带底部(廖翰卿等, 2013)或略低(吴荣昌, 2011)。该组顶界可能位于中奥陶统达瑞威尔阶下部的Lenodus variabilis牙形刺带上下(吴荣昌, 2011)。
研究区位于安徽省南部, 处于扬子台地东南缘呈带状分布的紫台组东端, 属于下扬子区西部。与上扬子区相比, 研究区内紫台组厚度较小, 泥质含量较低, 底栖壳相化石较为少见, 且几乎全部由紫红色瘤状泥质灰岩组成, 缺少命名剖面中紫台组上部的土黄色砂岩段。研究区内紫台组与下伏红花园组和上覆牯牛潭组均为整合接触关系, 岩性呈渐变过渡。
笔者实测了皖南石台丁香和池州洪家2条紫台组剖面(图 1), 进行了野外观察和描记, 采集牙形刺和岩石学样品, 室内切制薄片, 分层进行紫台组的薄片观察, 定量统计了基质、泥质、颗粒的百分含量, 并识别不同沉积组构, 描述、记录、归纳生屑颗粒的种类、磨圆、分选等特征, 进而归纳、分析紫台组的古环境特征及其变迁。
2.2.1 丁香剖面
丁香剖面位于安徽省石台县丁香镇东侧(GPS:N30° 17'48.7″, E117° 21'2.4″)。笔者2012年考察时, 该剖面虽然完整, 但局部仍有覆盖, 故地层学资料主要参考吴荣昌(2011)(图 3)。
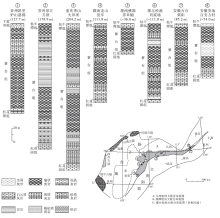
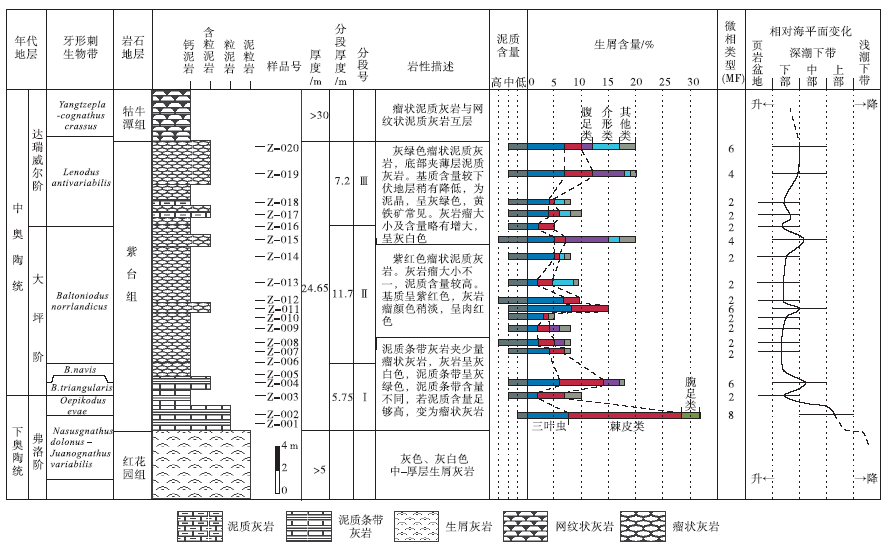 | 图3 安徽南部石台丁香剖面紫台组地层学和碳酸盐岩微相特征(生物地层资料据吴荣昌, 2011, 有修改)Fig.3 Summary of stratigraphic and carbonate microfacies data of the Zitai Formation at Dingxiang section of Shitai County, southern Anhui Province(biostratigraphic data modified from Wu, 2011) |
牯牛潭组(> 2 m)
灰色瘤状与网纹状泥质条带灰岩互层 2 m
— — — — — 整合接触 — — — — — —
紫台组(24.65 m)
第Ⅲ 段(7.2 m)
8.灰绿色瘤状泥质灰岩 5.2 m
7.紫红色薄层泥质灰岩 0.5 m
6.灰绿色瘤状泥质灰岩 1.5 m
第Ⅱ 段(11.7 m)
5.紫红色瘤状泥质灰岩、泥灰岩 11.7 m
第Ⅰ 段(5.75 m)
4.灰色瘤状泥灰岩, 瘤较大, 泥质含量较小 1.4 m
3.泥质条带灰岩, 泥质含量较大 1 m
2.灰色泥质条带灰岩, 泥质含量明显增加, 顶部有瘤状灰岩 1 m
1.灰色泥质条带灰岩 2.35 m
— — — — — — 整合接触 — — — — — —
红花园组(> 5 m)
灰色、灰白色中、厚层生屑灰岩 5.82 m
丁香剖面紫台组厚24.65 m, 根据岩性特征分为3段(图 3):第Ⅰ 段厚5.75 m, 主要为灰色泥质条带灰岩, 夹少量瘤状灰岩; 第Ⅱ 段厚11.7 m, 泥质含量增加, 以瘤状泥质灰岩为主, 特征为颜色呈紫红色, 其中基质为深紫红色, 灰岩瘤呈稍淡的紫红色; 第Ⅲ 段厚7.2 m, 依然为瘤状泥质灰岩, 但泥质含量较第Ⅱ 段略有降低, 颜色以灰绿色为主, 夹少量紫红色沉积。
丁香剖面紫台组中可识别出数个牙形刺生物带, 由下至上分别是(图 3):Nasusgnathus dolonus-Juanognathus variabilis 亚带(属Serratognathus diversus 带)、Oepikodus evae 带、Baltoniodus triangulars带、B. navis 带、B. norrlandicus 带、Lenodus antivariabilis 带(图 3)。紫台组底界位于Nasusgnathus dolonus-Juanognathus variabilis 亚带, 相当于Serratognathus diversus 带内; 顶界位于L. antivariabilis 带内。下、中奥陶统的界线位于第Ⅱ 段上部(修改自吴荣昌, 2011)。
N. dolonus-J. variabilis 亚带(属S. diversus 带)和O. evae 带的属种数以及新生率、灭绝率都较高, 而在B. triangularis带显示出明显的下降。另外牙形刺生物相分析显示, N. dolonus-J. variabilis 亚带属于较浅水生物相, O. evae 带和B. triangularis带属于深水生物相, B. navis 带、B. norrlandicus 带、L. antivariabilis 带代表水深波动较大, 但总体属于较浅水和深水之间的生物相。
2.2.2 洪家剖面
洪家剖面位于安徽省池州市贵池区棠溪乡048乡道附近(GPS:N 30° 16'29.5″, E 117° 39'23.1″), 距石台县城19.9 km, 距石台丁香剖面东偏北27 km。2012年, 笔者等实地考察该剖面, 并对紫台组进行了测量和样品采集。该剖面中紫台组发育完整, 出露基本连续, 只有上部一小部分被农田覆盖。文中研究的材料采自紫台组的下、中部(图 4)。
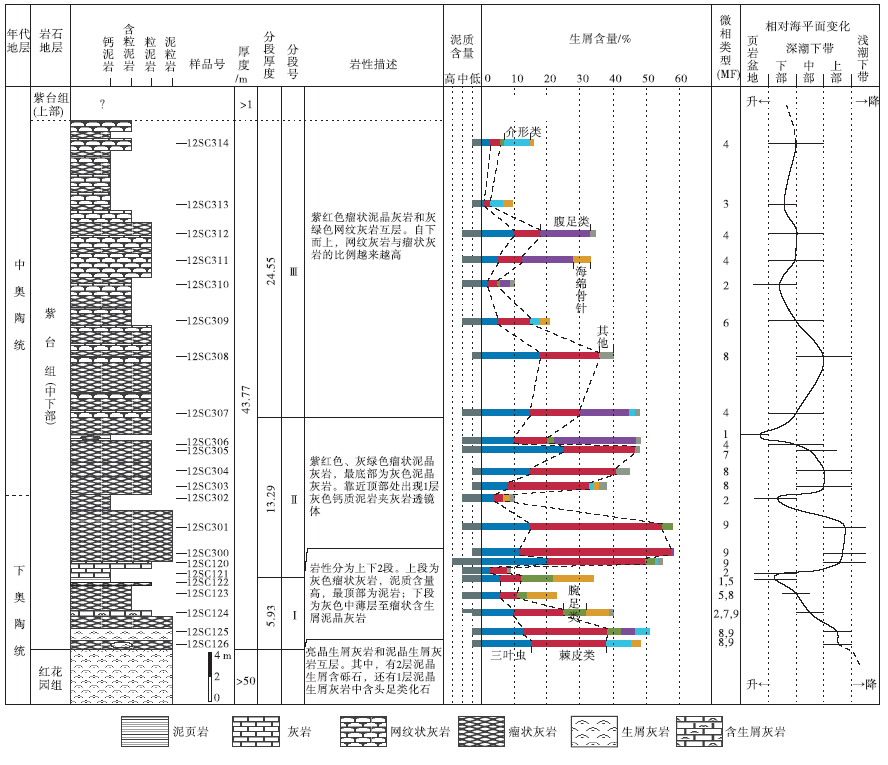 | 图4 安徽南部池州洪家剖面紫台组地层学及碳酸盐岩微相特征Fig.4 Summary of stratigraphic and carbonate microfacies data of the Zitai Formation at Hongjia section of Chizhou County, southern Anhui Province |
紫台组(43.77 m)(未到顶)
第Ⅲ 段(24.55 m)
26.灰绿色网纹状灰岩夹灰绿色瘤状灰岩 4.8 m
25.灰绿色网纹状灰岩夹灰绿色瘤状灰岩, 夹紫红色, 网纹状不密集 4.4 m
24.灰绿色瘤状灰岩 0.5 m
23.紫红色瘤状灰岩 0.2 m
22.灰绿色网纹状灰岩夹少量瘤状灰岩 5 m
21.瘤状泥晶灰岩夹少量网纹状灰岩 4 m
20.瘤状泥晶灰岩与网纹状灰岩互层 4.8 m
19.紫红色瘤状泥晶灰岩 0.7 m
18.灰绿色网纹状灰岩 0.15 m
第Ⅱ 段(13.29 m)
17.紫红色瘤状泥晶灰岩 1.6 m
16.灰色钙质泥岩夹瘤状泥晶灰岩透镜体 0.04 m
15.紫红色瘤状泥晶灰岩夹灰绿色瘤状泥晶灰岩 0.9 m
14.灰绿色瘤状泥晶灰岩夹紫红色瘤状泥晶灰岩, 紫红色泥质含亮晶 2 m
13.紫红色瘤状泥晶灰岩, 生屑含量增加 0.3 m
12.灰绿色瘤状泥晶灰岩夹少量紫红色瘤状泥晶灰岩 1.6 m
11.灰绿色瘤状泥晶灰岩夹较多紫红色瘤状泥晶灰岩 1.95 m
10.紫红色瘤状泥晶灰岩和灰绿色瘤状泥晶灰岩互层 1.6 m
9.灰色含生屑瘤状灰岩 1.6 m
8.紫红色含生屑泥晶灰岩 0.1 m
7.浅灰色泥晶灰岩, 含少量生屑, 夹紫红色泥质泥晶灰岩条带 1.6 m
第Ⅰ 段(5.93 m)
6.黄灰色泥岩, 底部夹瘤状泥晶灰岩 0.58 m
5.灰色瘤状灰岩, 泥质含量高 2.15 m
4.灰色中层含生屑泥晶灰岩, 含三叶虫化石 0.8 m
3.灰色瘤状— 薄层含生屑泥晶灰岩 1.2 m
2.灰色中层含生屑泥晶灰岩, 含少量瘤状灰岩 1 m
1.灰色瘤状含泥质泥晶灰岩夹生屑泥晶灰岩透镜体, 含风暴岩 0.2 m
— — — — — — 整合接触 — — — — — —
红花园组(> 50 m)
亮晶生屑灰岩和泥晶生屑灰岩互层, 含头足类化石 2.7 m
中层含砾生屑泥晶灰岩夹含生屑泥晶灰岩 0.7 m
洪家剖面紫台组实测部分厚43.77 m, 依据岩性特征分为3段(图 4):第Ⅰ 段厚5.93 m, 下部为灰色中薄层至瘤状含生屑泥晶灰岩, 上部为灰色瘤状泥质灰岩, 最顶部发育厚约58 cm的黄灰色泥岩。第Ⅱ 段厚13.29 m, 岩性为紫红色瘤状泥质灰岩和灰绿色瘤状泥质灰岩互层。第Ⅲ 段厚24.55 m, 以灰绿色网纹状灰岩出现为标志, 并与紫红色瘤状泥质灰岩互层, 且向上网纹状灰岩比例逐渐增高。
碳酸盐岩微相分析是碳酸盐岩沉积学研究的重要手段。笔者通过对丁香剖面、洪家剖面共计48片岩石薄片进行微相研究, 其中丁香剖面的紫台组薄片共17片, 编号由下至上为Z-002至Z-004, Z-007至Z-020(图3); 洪家剖面的紫台组薄片共31片, 编号由下至上为12SC126至12SC120, 12SC300至12SC314(图4)。
总体来说, 紫台组灰岩的颗粒种类并不丰富且组分成熟度不高, 古环境变化不很剧烈, 这给传统的微相划分带来了一定的困难。因此, 笔者在前人研究的基础上(刘建波, 2006), 使用点计数法(温俊君和刘建波, 2009)对紫台组进行定量微相分析, 结合所发育颗粒(主要为生屑)和基质类型、形态代表的古环境指示意义(王英华等, 1990; Flü gel, 2010)以及泥质成分含量等沉积特征, 在研究区共识别出9种碳酸盐岩微相(图 5)。
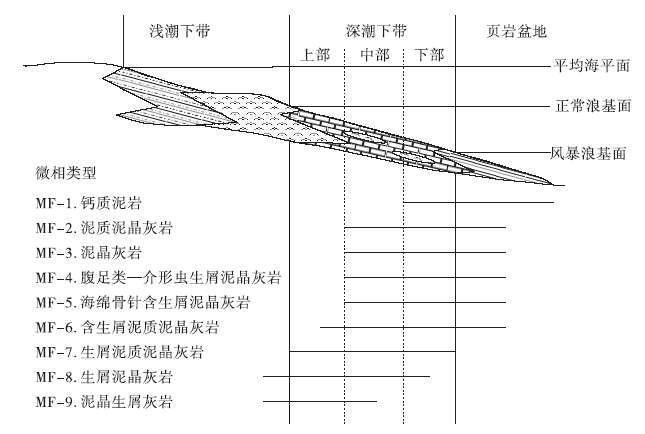 | 图5 紫台组碳酸盐岩微相在碳酸盐岩缓坡上的分布模式Fig.5 Distribution pattern of microfacies of the Zitai Formation on carbonate ramp |
沉积特征:泥质含量大于50%的层位, 纹层发育, 常发育页理, 野外易辨认。化石稀少, 偶见笔石碎片, 未见生物扰动构造。出现在紫台组下部。
环境解释:纹层发育、底栖化石缺少、碳酸盐沉积速率低等特征, 都说明该微相形成于安静贫氧的深水环境。推断其古环境为深潮下带下部至页岩盆地, 多见于页岩盆地。
 | 图6 安徽南部石台丁香和池州洪家剖面野外沉积学照片Fig.6 Field photographs of the Zitai Formation at Dingxiang and Hongjia sections in Shitai and Chizhou respectively, southern Anhui Province |
沉积特征:总体特征是生屑含量低, 泥质含量中等至高。生屑含量小于10%, 其中三叶虫和棘皮类较常见, 偶见完整的腹足类、腕足类和介形类等。各类生屑破碎程度低, 磨圆、分选均很差。另外, 泥质中常有与层面近平行的压溶线和破碎的生屑。瘤状构造发育, 少数具网纹状构造。本微相出现在2条研究剖面中, 丁香剖面尤为发育。
环境解释:该微相生屑含量低, 缺乏亮晶, 应沉积于正常浪基面之下。泥质含量较高, 但低于MF-1, 指示相对深水的沉积环境, 多见于深潮下带下部至页岩盆地上部, 少数可达深潮下带中部。
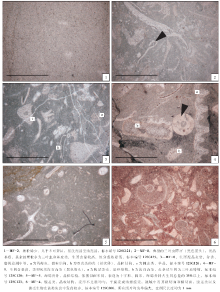
 | 图7 安徽南部池州洪家剖面紫台组碳酸盐岩微相显微照片Fig.7 Photos of thin section showing carbonate microfacies of the Zitai Formation at Hongjia section of Chizhou County, southern Anhui Province |
 | 图8 安徽南部石台丁香和池州洪家剖面紫台组碳酸盐岩微相显微照片Fig.8 Photos of thin section showing carbonate microfacies of the Zitai Formation at Dingxiang and Hongjia sections in Shitai and Chizhou respectively, southern Anhui Province |
沉积特征:MF-3泥晶灰岩与MF-2类似, 区别在于泥质含量低、小于10%。生屑发育状况大体同MF-2, 分选、磨圆差, 既有完整的生屑颗粒, 也有细小生屑颗粒因重结晶而难以辨认。薄层构造, 通常具网纹状构造。
环境解释:该微相与MF-2的区别仅在于泥质含量稍低, 所以推测该微相的沉积水体比MF-2稍浅, 为深潮下带中部至页岩盆地, 但多见于深潮下带下部。
沉积特征:生屑含量为15%~48%, 其中薄壳生屑(绝大部分为介形虫壳体)和腹足类占到35%, 泥质含量不稳定, 或低或高。泥晶基质支撑, 网纹状构造和瘤状构造均发育。
环境解释:本微相泥晶基质支撑, 指示低能环境。腹足类、薄壳生屑等生物占主导, 从这两类生物的古生态(王英华等, 1990)判断, 该微相代表了一种以浮游、游泳生物为主、缺少底栖生物的古环境, 推断为深潮下带中部至下部的环境, 个别可至页岩盆地上部。
沉积特征:生屑含量为22%~33%, 海绵骨针占生屑总量的35%以上, 其他生屑以三叶虫和棘皮类较为常见。泥质含量中等。具薄层构造。
环境解释:三叶虫和棘皮类等底栖生物骨骼化石指示正常海洋环境, 而丰富的海绵骨针通常保存于较深水环境(王英华等, 1990; Flü gel, 2010)。该微相沉积于深潮下带中部至页岩盆地上部, 但多见于深潮下带下部。
沉积特征:生屑含量为10%~23%, 泥质含量中等至高。生屑类型主要包括三叶虫、棘皮类、腹足类、海绵骨针等, 磨圆、分选差。以瘤状构造为主。
环境解释:综合其生屑特点以及泥质含量, 推测该微相指示了深潮下带中部偏上至页岩盆地上部的环境, 多见于深潮下带中、下部。
沉积特征:生屑含量为25%~48%, 泥质含量中等至高。常见的生屑有三叶虫、棘皮类、腕足类、腹足类、海绵骨针等, 生屑磨圆、分选差, 瘤状构造发育, 少量为薄层状。
环境解释:结合其泥质含量以及基质组成, 推测其沉积环境为深潮下带, 但多形成于深潮下带中部偏上。
丁香和洪家剖面处于下扬子区台地边缘区, 相距不足30 km。2个剖面岩性特征总体相近, 沉积序列和岩性变化基本一致:两者在岩性上都可以分为3段, 第Ⅰ 段和第Ⅱ 段的划分均是基于紫红色瘤状灰岩、泥灰岩的出现, 泥质含量也都是Ⅰ 、Ⅲ 段较低, Ⅱ 段较高(图 3, 图4)。根据牙形刺化石带和岩性特征, 可以将两地的紫台组进行比较精确的对比(图 9)。
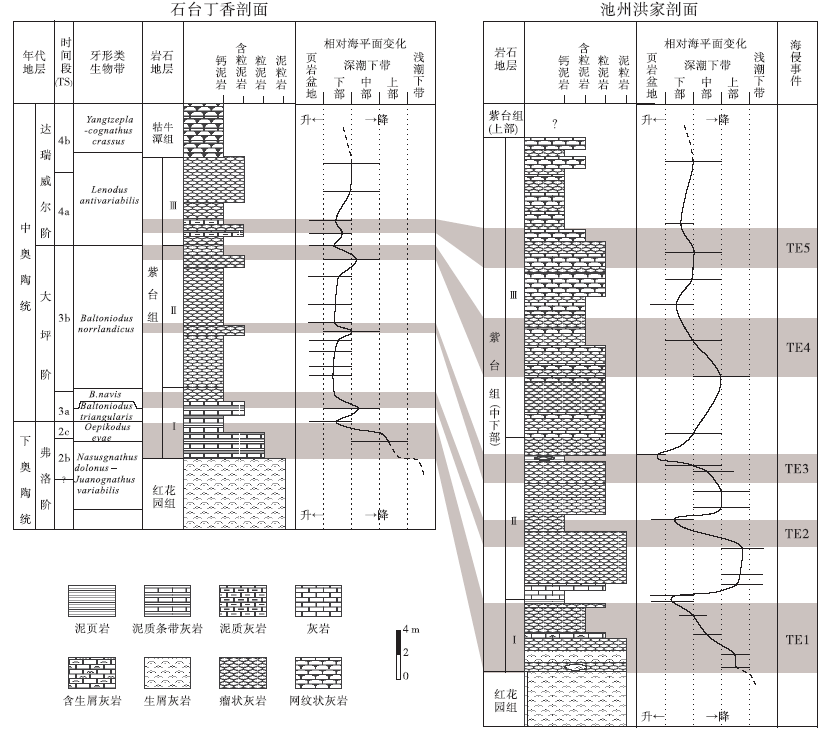 | 图9 安徽南部石台丁香剖面和池州洪家剖面紫台组形成期间海平面变化对比(TS是time slice的缩写, 指Webby等(2004)所建立的奥陶纪时段; TE是transgressive event的缩写, 指本研究识别出的相对海平面上升事件)Fig.9 Correlation of sea-level changes during the depositional period of Zitai Formation between Dingxiang section of Shitai and Hongjia section of Chizhou, southern Anhui Province(TS:Time slices of Webby et al.(2004); TE:Transgressive event identified in this study) |
丁香和洪家剖面紫台组岩性比较单调, 以紫红色灰岩为特征(图 6-3, 6-4), 以泥晶基质支撑结构为主, 以瘤状构造发育为特点, 矿物成分以方解石为主、白云石少见、泥质成分较丰富。颗粒类型基本为生屑, 包括三叶虫、棘皮类、腹足类、腕足类、海绵骨针、介形虫以及苔藓虫等。其中三叶虫和棘皮类最为常见(图 3, 图4), 三叶虫碎屑总体比较完整, 基本没有分选和磨圆; 棘皮类多为海胆纲和海百合纲, 大多较为破碎, 但在生屑含量高的层位也易见1~2 mm的海百合茎、海百合骨板等。其他种类的生屑较少, 分选、磨圆较差。
生屑特征分析对于碳酸盐岩沉积学研究非常重要(刘建波等, 2013)。紫台组中优势生屑往往以近原地沉积为主。营底栖游移和底栖游泳生活的三叶虫、营底栖游移和底栖固着生活的棘皮类在紫台组沉积中占有优势地位, 说明当时海洋水体的含氧量较高, 这与奥陶纪海水中氧气含量显著增加的论断(Wilde, 1987)相吻合。大气中氧含量的增加有可能与奥陶纪生物大辐射事件有着密不可分的联系(如Berner, 2006)。而扬子台地边缘区海洋底层水体氧气含量高无疑与紫台组海相红层的形成有着密切的联系(胡修棉等, 2006; Kiipli et al., 2008, 2010; 吴荣昌, 2011)。
通过分析, 2个研究剖面中的紫台组可区分出9个碳酸盐岩微相, 所有微相主要形成于深潮下带(介于正常浪基面和风暴浪基面之间), 个别微相可形成于浅潮下带下部较高能环境或页岩盆地环境(图 5)。其中, 洪家剖面紫台组中MF-1至MF-9都有发育(图 4), 而丁香剖面紫台组只发育有4种微相(MF-2、MF-4、MF-6和MF-8)(图 3)。丁香剖面紫台组中发育最多的是MF-2泥晶泥质灰岩(图 3), 生屑颗粒含量低。洪家剖面发育最多的微相包括MF-4、MF-8和MF-9(图 4), 生屑含量相对较高, 并在底部层位出现生屑颗粒支撑结构和泥— 亮晶混合基质结构(图 7-4), 这在丁香剖面紫台组中从未见及。丁香剖面紫台组上部依然以紫红色瘤状灰岩为主, 而洪家剖面上部则以网纹状灰岩为主, 泥质含量明显偏低。由此推断, 两地的碳酸盐沉积速率和/或细粒陆源碎屑供给量存在一定的差异。丁香剖面更靠近下扬子区台地边缘区, 紫台组沉积时期水体深度总体上比洪家剖面要大一些, 从而使该地区带壳底栖生物偏少, 碳酸盐沉积速率偏低, 沉积物中泥质含量相对较高, 瘤状构造发育。
丁香剖面和洪家剖面紫台组与下伏地层红花园组之间沉积特征差异较大。研究区内红花园组与上扬子区(廖翰卿等, 2013)类似, 以浅水碳酸盐岩台地沉积为主, 顶部为沉积于浅潮下带的亮晶生屑灰岩(图 3, 图4)。紫台组底部为泥质条带泥晶灰岩或瘤状泥晶灰岩, 沉积于深潮下带(图 9)。这个沉积相快速变化现象是扬子区早奥陶世碳酸盐岩台地淹没事件在本地区的表现。
碳酸盐岩台地淹没(drowning of carbonate platform)指浅水碳酸盐台地突然被深水沉积覆盖(Schlager, 1981)。扬子台地早奥陶世碳酸盐岩台地淹没事件在整个台地广泛分布(冯增昭等, 2001; 苏文博, 2001; 刘建波, 2006; 廖翰卿等, 2013)。该事件主要是由于海平面快速上升(刘建波, 2006)和来自于台地西部剥蚀区的陆源碎屑供给增多(廖翰卿等, 2013)造成的。由于在早奥陶世下扬子区陆源碎屑缺乏, 也无重要的剥蚀区(冯增昭等, 2001), 故研究区内红花园组顶部的淹没事件更多受控于弗洛晚期(TS.2b-2c)发生的相对海平面的快速上升(TE1, 图9)。沉积水深的快速增大降低了紫台组碳酸盐的沉积速率, 从而深水瘤状灰岩发育, 类似成因的瘤状灰岩也发育于湘西北上奥陶统(葛祥英等, 2013)。根据牙形刺生物地层学资料(吴荣昌, 2011, 廖翰卿等, 2013), 此次海平面上升事件(TE1)发生于早奥陶世弗洛中、晚期, 与在上扬子区TS.2b晚期(刘建波, 2006)、澳大利亚的后ECEE海侵(Young and Laurie, 1996)、北欧的Evae淹没事件(Nielsen, 2004)以及华北地块(Liu, 1998)发生的海侵事件同步, 具有全球性。
除了TE1这次明显的相对海平面上升事件外, 在丁香和洪家剖面紫台组中还可以识别出4次海平面上升事件, 并可以相互对比(TE2-TE5, 图9)。丁香剖面紫台组第Ⅰ 段主要发育泥质条带灰岩, 顶部发育少量灰色瘤状灰岩, 泥质含量低到中等(图 8-3)。该段上部的微相类型从MF-6变为MF-2, 沉积环境由深潮下带中部变为深潮下带下部(图 3), 指示在TS.3a晚期有1次相对海平面升高事件(图 9中TE2)。TE2期间, 牙形刺生物相从较浅水Paroistodus相变化为较深水Periodon-Protopanderodus相(吴荣昌, 2011), 同样指示海平面的上升事件。该次事件在洪家剖面表现为紫台组第Ⅱ 段出现紫红色瘤状灰岩, 并与灰绿色瘤状灰岩互层, 泥质含量低到高不等, 生屑含量较高(图 4)。这次海侵事件在上扬子区则表现为深色页岩沉积增多现象(刘建波, 2006), 也在北美被识别出来(Ross and Ross, 1992)。
丁香剖面紫台组第Ⅱ 段开始出现紫红色瘤状灰岩, 泥质含量高到中等, 灰质瘤较小。中部生屑含量从不足10%增加到15%, 生屑类型以三叶虫和棘皮类为主(MF-6)(图 3)。之上生屑含量降低至小于10%, 并开始出现较深水生屑(MF-2), 指示出现了海平面上升事件(TE3, TS.3b中期)。洪家剖面紫台组第Ⅱ 段中部出现一次从MF-9到MF-2的微相变化, 并在之上出现钙质泥岩(MF-1)(图 4, 图6-1), 可能指示同期海平面变化事件(图 9)。该事件在上扬子区宜昌陈家河剖面表现为大湾组“ 中灰岩” 之上灰绿色泥岩沉积的增多, 在重庆城口大槽剖面表现为厚坪组浅潮下带亮晶生屑灰岩的消失(刘建波, 2006)。而且TE3可以与在北欧(Nielsen, 2004)、北美(Ross and Ross, 1992)和澳大利亚(Young and Laurie, 1996)等地识别出的海侵事件进行对比(刘建波, 2006)。
丁香剖面紫台组第Ⅱ 段顶部依然以紫红色瘤状灰岩为特征, 再次出现生屑含量增高的层位(> 20%), 生屑类型以腹足类为主, 三叶虫次之(MF-4)(图 3)。之上生屑含量降低至5%左右(MF-2), 代表海平面的上升事件(TE4, TS.3b末期)。该剖面第Ⅲ 段下部开始出现灰绿色瘤状灰岩, 泥质含量低到中等, 瘤体相对第Ⅱ 段变大, 下部虽然以MF-2为主, 但根据生屑含量和类型变化, 推断在TS.4a早期发生过一次小规模的海平面上升事件(TE5)。在洪家剖面, 紫台组第Ⅲ 段中下部以紫红色瘤状泥晶灰岩和灰绿色网纹状灰岩互层为特征, 生屑含量依旧大于40%(MF-8)(图 4), 向上生屑明显减少, 这可能与TE4海平面上升事件有关。紫台组第Ⅲ 段上部微相类型从MF-4变为MF-3, 生屑含量减少了20%以上, 沉积环境从深潮下带中、下部变为深潮下带下部, 与丁香剖面识别出的TS.4a早期的TE5相当(图 9)。TE4和TE5海平面上升事件之前在上扬子区的宜昌陈家河剖面和城口大槽剖面同样被识别出(刘建波, 2006)。在陈家河剖面, 2次海平面上升事件均表现为深色页岩沉积的发育。在对三叶虫生物相(如Turvey and Zhou, 2004)、牙形刺生物相(吴荣昌, 2011)的研究中均在紫台组上部识别出该时期海平面上升事件。这2次事件还可以与在北欧多地识别出的达瑞威尔期起始时期的碳酸盐岩台地“ 淹没事件” (Nielsen, 2004)进行对比。
华南下、中奥陶统紫台组以紫红色瘤状泥质灰岩为特征, 分布于扬子台地东南缘。本研究通过对安徽南部紫台组进行沉积学研究, 获得如下新的认识:
1)通过野外工作和室内定量碳酸盐岩微相分析, 在皖南紫台组中识别出9种碳酸盐岩微相, 其主要形成于台地边缘深潮下带中、下部及相邻环境中。
2)皖南紫台组沉积时水深波动幅度不大, 且自下而上沉积水深逐渐加大; 丁香剖面由于更接近台地边缘, 总体沉积水深要大于洪家剖面。
3)在2条研究剖面的紫台组中识别出至少5次比较显著的海平面上升事件, 分别发生在弗洛晚期TS.2b-2c(TE1)、大坪期TS.3a晚期(TE2)、TS.3b中期(TE3)、TS.3b末期(TE4)和达瑞威尔期TS.4a早期(TE5)。
4)下扬子区台地边缘海平面上升事件与上扬子区海侵事件具有良好的对应关系, 并可以与澳大利亚、北欧、北美、华北等板块进行对比, 说明区域性相对海平面变化、甚至全球性海平面变化控制着紫台组的沉积及其变化。
致谢 王光旭、孙永超、闫振、景宇轩在野外工作中提供帮助, 周传明、王宗哲、卓二军提供技术支持并与作者进行有益讨论, 审稿人给出宝贵意见, 笔者谨此一并致以诚挚谢意!本研究为IGCP591项目的成果。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|