

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
北京西山下苇甸剖面寒武系徐庄组鲕粒研究
[邢延路 , 冯李强]
, 冯李强]
, 冯李强]
|
|

第一作者简介 邢延路,男,1991年生,中国地质大学(北京)硕士研究生,主要从事矿物学、岩石学和矿床学方面的研究。E-mail: jdbshsj@163.com。
对北京西山下苇甸剖面寒武系徐庄组鲕粒灰岩进行了观察和研究,在其中不仅发现了有核及无核同心—放射状鲕、单(或多)晶鲕、泥晶鲕和复合鲕,而且还发现了暗色颗粒包壳、凝聚颗粒和黄铁矿聚集体等特殊组构。暗色颗粒包壳的发现尚属首次,其可能是丝状蓝细菌菌体的残留物。凝聚颗粒由方解石或白云石构成,可能代表了微生物席内的含氧气泡。生物碎屑上的附着体可能是微生物群落的痕迹,在其内部发现有类似鲕粒的球状颗粒。复合鲕胶结松散,包壳不发育,表明其可能形成于相对稳定的水体环境。研究表明,徐庄组鲕粒可能形成于水动力条件相对较弱的浅海环境中,微生物在鲕粒的形成过程中发挥了较大的作用。
About the first author Xing Yanlu,born in 1991,is a candidate for master's degree in China University of Geosciences(Beijing). He is mainly engaged in mineralogy,petrology and ore geology. E-mail: jdbshsj@163.com.
This paper systematically studied the characteristics of oolitic limestone of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Western Hills of Beijing. Core-beard and core-missed concentric-radial ooids,monocrystalline/polycrystalline ooids,micritic ooids and compound ooids have been found in this study. At the same time,we also found some special textures,such as dark-colored cortices,compounded granules and pyrite agglomerates. Dark-colored cortices are found for the first time,which are probably the remains of filamentous cyanobacteria bodies. Compounded granules,which may represent the oxygen-containing bubbles in microbial mats,are composed of crystals of calcite or dolomite. An attachment is found on the surface of a bioclast and ooid-like particles are found inside the attachment. Compounded ooids,which are loosely compacted and lack of outside cortices,may indicate that they are generated in a relatively quiet shallow water environment. Our research shows that ooids in the Xuzhuang Formation were probably generated in a quiet shallow water environment and microbes played a key role in this process.
鲕粒多为直径小于 2 mm 的球状颗粒, 主要存在于碳酸盐岩中。目前, 关于鲕粒的成因有2种主流的认识:一种是机械及化学成因说(Duguid et al., 2010), 即认为鲕粒形成于较为动荡的水体环境中。另一种是微生物成因说, 认为微生物在碳酸盐沉淀及鲕粒同心纹层的形成过程中发挥了关键的作用。如Duguid 等(2010)通过对巴哈马滩鲕粒的观察和研究, 认为鲕粒的形成与微生物不存在直接的关系, 在方解石、文石沉淀及鲕粒同心纹层形成过程中, 化学过程起到了关键的作用。然而, Brehm 等(2006)通过实验室培养, 成功地利用他们所定义的“ 球状微生物群落” 人工合成了鲕粒, 进一步强调了鲕粒形成的微生物作用机制。此外, 通过对瑞典Lake Geneva地区淡水鲕的研究, 发现光合微生物所控制的微生物群落在鲕粒形成的早期诱导了方解石的沉淀(Plé e et al., 2008)。
在鲕粒的机械及化学成因模式下, 鲕粒的形成似乎离不开一个动荡的水体环境。然而, 若是微生物的作用占主导, 则较为动荡的环境条件可能不再是一个必要条件(Pacton et al., 2012), 这也就导致了2种成因说在解释鲕粒的形成条件上的较大矛盾。2种假说的差异, 代表了在地质历史时期地层中广泛发育的鲕粒所反映的沉积相及其代表的相应沉积环境不同。因此, 研究鲕粒的成因, 对更为准确地厘定其所反映的古沉积环境、划分沉积相都具有十分重大的意义。
基于以上认识, 笔者对北京西山下苇甸寒武系徐庄组顶部的厚层鲕粒灰岩进行了系统的野外观测和室内研究, 发现了很多鲕粒形成过程中的微生物作用信号。文中通过讨论在研究过程中所发现的特殊的鲕粒及组构, 试图为鲕粒的微生物成因研究提供更多的理论依据。
北京西山下苇甸寒武系剖面位于北京市门头沟区妙峰山镇(图 1), 是华北地台寒武系沉积现象较为丰富的地层剖面之一。20世纪80年代以来, 很多学者对该剖面进行了较为系统的沉积学研究(孟祥化等, 1986; 王英华等, 1989; 冯增昭等, 1990; 卢衍豪等, 1994; 梅冥相等, 1997, 2011; 高林志和乔秀夫, 2001; 张旭等, 2009; 朱传庆等, 2009)。该剖面寒武系出露较为完整, 从下至上依次为:第二统昌平组、馒头组; 第三统毛庄组、徐庄组、张夏组、崮山组; 芙蓉统长山组、凤山组。各组地层出露相对完好, 发育丰富的沉积现象。其中, 在徐庄组、张夏组及崮山组顶部均发育有大套厚层状鲕粒灰岩。前人利用该剖面对张夏组鲕粒灰岩及生物丘(王成述等, 1990; 郝云兰等, 2012)、崮山组叠层石生物丘及沉积组构(梅冥相等, 2011)等进行过较为详细的研究。但是, 尚未有学者对该剖面徐庄组鲕粒灰岩中鲕粒的特征及其他一些沉积组构进行过系统的研究。
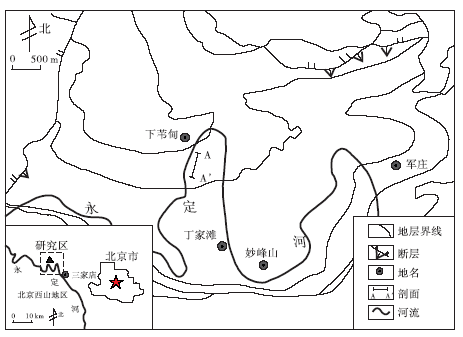 | 图1 北京下苇甸剖面位置(据郝云兰等, 2012, 有修改)Fig.1 Geographical location of Xiaweidian outcrop in Beijing(modified from Hao et al., 2012) |
下苇甸剖面徐庄组厚70~80 m, 与下部毛庄组及上部张夏组均为平行不整合接触。徐庄组下部主要为一套潮坪相的细粒碎屑岩— 泥岩沉积, 夹有白云岩透镜体及透镜层; 向上为1套厚层鲕粒灰岩, 厚约40 m, 反映了高能的鲕粒滩相沉积(冯增昭等, 1990; 梅冥相等, 2011)。总体上, 鲕粒灰岩呈浅灰色— 灰色, 粒径多为0.3~0.7 mm, 部分颗粒粒径可达1 mm; 颗粒含量可达50%以上, 分布相对均匀。
由于徐庄组鲕粒灰岩所显示出的高能鲕粒滩相, 是在华北地台上早古生代以来首次出现的高能量的滩, 且是以陆源沉积为主导的坪环境向以碳酸盐沉积为主导的滩环境演化的第1个滩(冯增昭等, 1990), 而到了张夏期, 华北地台则成了一个广阔的碳酸盐岩台地(冯增昭等, 1990; 朱传庆等, 2009)。因此, 徐庄组鲕粒滩在沉积环境演变的过程中发挥了承上启下的作用, 故对其中以鲕粒为代表的沉积组构的研究, 对深入了解这种演变背后的环境意义具有不可替代的作用。
对徐庄组顶部鲕粒灰岩以2~3 m为间隔取样, 进行了手标本及镜下特征的详细观察和记录, 并将鲕粒灰岩等间距划分为5个层段(图 2)。总体上, 徐庄组内鲕粒类型丰富, 主要包括具核同心— 放射状鲕、无核心同心— 放射状鲕、单(或多)晶鲕、泥晶鲕以及复合鲕粒等。
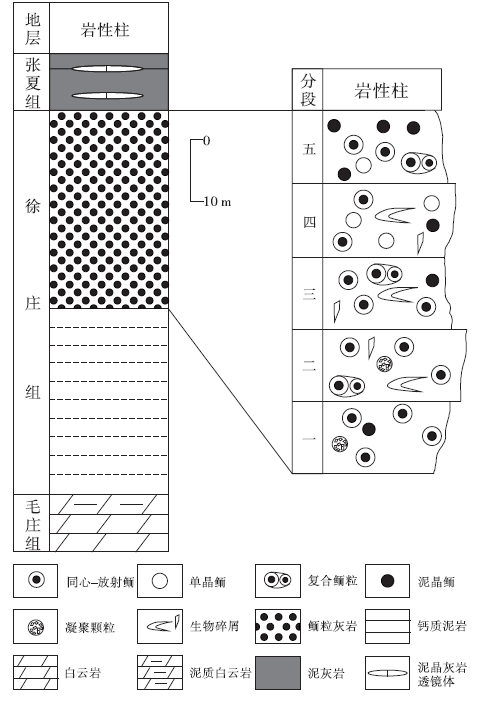 | 图2 北京下苇甸剖面寒武系徐庄组地层特征及主要鲕粒类型Fig.2 Characteristics of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Beijing and main types of ooids |
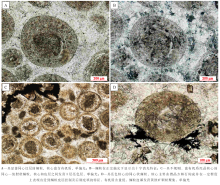
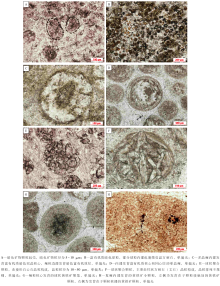
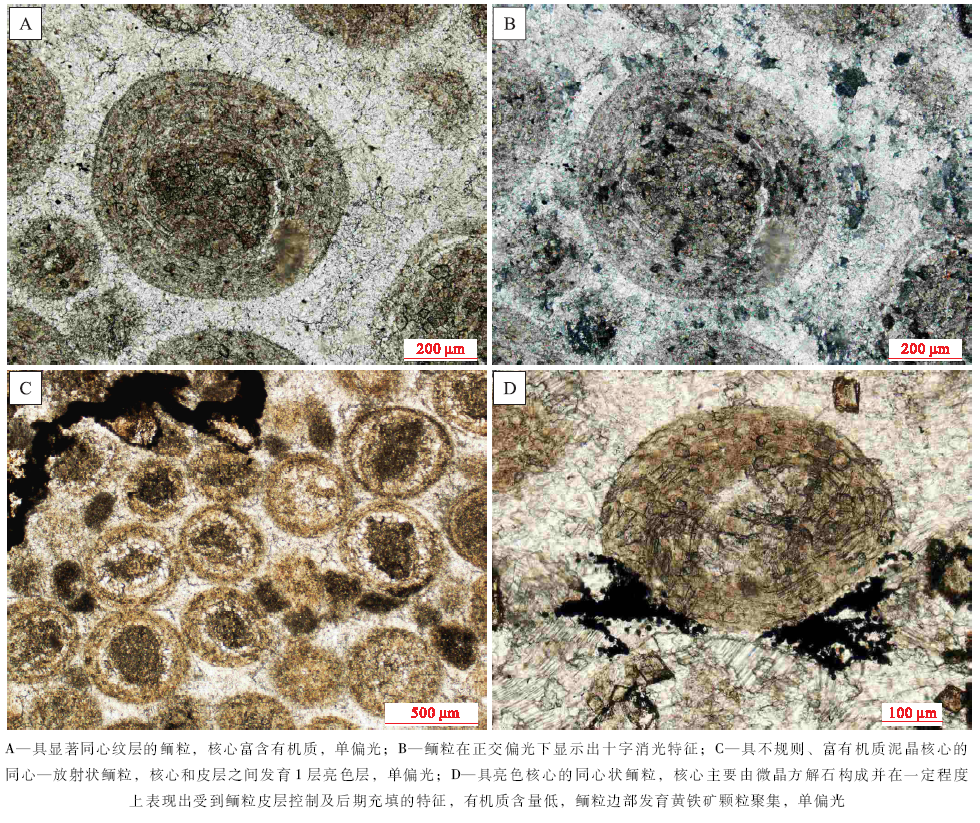
具核同心— 放射状鲕在徐庄组鲕粒灰岩中发育最为广泛, 在各个层段均有发现, 其中, 在第一、四和五段发育较多。总体上, 具核同心— 放射状鲕的粒径多为300~600 μ m, 形状规则, 多呈圆— 椭圆状, 分布不均匀, 局部成群密集分布。鲕粒的核心特征变化明显, 根据成分可划分为泥晶方解石核心(图 3-C)、亮晶方解石核心(图 3-D)、白云石晶粒聚集核心及生物骨屑核心等。此外, 核心的直径变化较大, 为50~300 μ m。相应地, 皮层厚度也变化较大, 但其成分变化不大, 主要为有机质含量较高的泥晶或微晶方解石。由于受到后期成岩作用影响, 大部分鲕粒的同心纹层遭到了不同程度的破坏, 变得模糊不清。但是, 亦发现有由白云石晶粒规则排列所表现出的同心纹层结构(图 3-A), 其可能是受后期白云岩化作用改造所致, 反映了原始鲕粒同心纹层的存在。
 | 图3 北京下苇甸剖面寒武系徐庄组具核同心— 放射状鲕粒特征Fig.3 Characteristics of concentric-radial ooids with obvious cores of the Cambrian Xuzhuang Formation at |
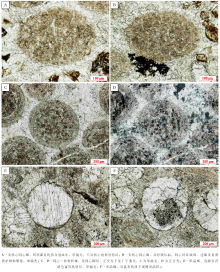
无核同心— 放射状鲕以其不发育明显核心而与有核鲕粒区分开来。该种鲕粒发育相对广泛, 在徐庄组第二、三段发育较为集中。鲕粒的粒径多为300~500 μ m, 形状为较规则的球— 椭球状(图 4-A, 4-B, 4-C, 4-D), 多数仍依稀可见由微晶方解石构成的圈层结构。该种鲕粒的成分均一, 内外变化不明显, 具有相对较高的有机质含量(图 4-B, 4-C)。部分鲕粒的外表面发育有黄铁矿聚集体(图 4-B)。无核同心— 放射状鲕的外表面平滑完整(图 4-A, 4-C), 这与现代巴哈马滩鲕粒较为相似, 表现为浮游鲕粒的典型特征(Gerdes et al., 1994)。梅冥相等(2011)曾在该剖面寒武系崮山组鲕粒灰岩中发现了类似的无核心鲕粒, 认为这种鲕粒属于微生物席内形成的底栖鲕粒。
 | 图4 北京下苇甸剖面寒武系徐庄组无核心同心— 放射状鲕及单(多)晶鲕特征Fig.4 Characteristics of concentric-radial ooids without cores and single(multiple) crystle(s) ooids of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Beijing |
这种颗粒主要发育在徐庄组鲕粒灰岩的第四段, 在其他几个层段零星出露。单(或多)晶鲕最典型特征表现为其由一块或数块方解石晶体构成, 粒径多为300~400 μ m; 形态规则, 呈球— 椭球状; 未见有明显的变形或破碎。相当一部分颗粒的边部发育有暗色的富有机质环边(图 4-E), 可能是原生鲕粒被溶蚀后形成的富有机质残余。虽然受后期成岩作用的影响, 单(或多)晶鲕的原生结构几乎被破坏殆尽, 但仍然有少部分鲕粒的核心被保存下来(图 4-F)。单(或多)晶鲕往往被认为是原生鲕粒在后期成岩改造过程中遭受溶蚀、后为方解石充填而成, 因此并不是真正的鲕粒, 但可在一定程度上反映原生鲕粒的发育特征。
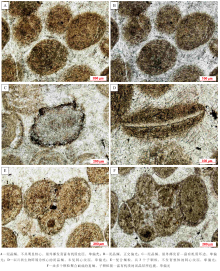
徐庄组鲕粒灰岩内部各层段均见泥晶鲕零星分布, 以第五段发育相对更为集中。泥晶鲕的粒径为250~350 μ m, 多小于其他类型鲕粒; 往往不具明显的核心, 同时也不见明显的同心或放射状结构(图 5-A, 5-B, 5-C); 形态不甚规则, 呈球状、椭球状或不规则状; 有机质含量相对较高, 同时, 一些鲕粒的最外部则发育有富有机质的泥晶套或暗色环边(图 5-A, 5-C)。一些泥晶鲕内部包裹片状的生物碎屑, 但这种鲕粒同样不见同心纹层(图 5-D)。与高能水体中以异化碳酸盐颗粒为核心生长的鲕粒所不同的是, 在徐庄组鲕粒灰岩中发育的泥晶鲕往往垂直于生物碎屑的主平面向两侧增生, 因此并不能简单地认为其包裹生物碎屑的增生体是以机械运动的方式生长的。此外, 徐庄组泥晶鲕外表面往往凹凸不平或是边界较为模糊, 显示出其可能经历了溶蚀作用。
 | 图5 北京下苇甸剖面寒武系徐庄组泥晶鲕及复合鲕粒特征Fig.5 Characteristics of micritic ooids and compound ooids of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Beijing |
复合鲕见于徐庄组鲕粒灰岩的第二、三和五段, 其中以第三段发育最多, 总体上数量稀少。复合鲕主要由2~3个子鲕粒聚合而成, 部分可具3个以上子颗粒(图 5-E, 5-F); 其粒径受子颗粒的数量和大小的限制, 变化较大。大多数复合鲕的子颗粒与相应层段的主要鲕粒类型一致, 且大小也处于同一变化范围。值得注意的是, 大部分复合鲕的外部都发育一个暗色的、薄层、富有机质的泥晶包壳(图 5-F), 这种泥晶包壳与一些学者所发现的复鲕的同心包壳结构(梅冥相等, 2011; Woods, 2013)明显不同。在一些复合鲕内部, 子颗粒的接触部位也常见黄铁矿聚集体发育(图 6-H)。
 | 图6 北京下苇甸剖面寒武系徐庄组特殊组构Fig.6 Characteristics of special textures of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Beijing |
在徐庄组鲕粒灰岩二段的上部, 可见一些十分特殊的由暗绿色颗粒构成的环形包壳结构(图 6-A)。这些包壳结构的直径比该层段中的正常鲕粒小, 为150~250 μ m, 内部充填暗色云雾状物质及泥晶方解石。构成包壳的暗绿色颗粒显微镜下几乎不透明, 粒径为5~10 μ m, 总体形状规则, 呈近圆状、椭圆状或短链状, 疑似丝状蓝细菌的残余物。梅冥相等(2011)曾在该剖面的寒武系崮山组中, 发现了类似的疑似丝状蓝细菌残余, 并把其定义为葛万氏菌(Girvanella)。对比发现, 二者在形态特征、大小及成分上有较高的相似性。另外, 在包壳状结构发育位置附近, 可见大量的有机质残余和暗色球粒(图 6-B)。这些形状和大小相对规则并以富有机质为显著特征的颗粒可能是丝状蓝细菌藻丝体的残片, 虽然数量十分稀少, 但是它们的发现却为揭示中寒武世北京西山地区由潮坪相向首个滩相环境转变, 以及可能存在微生物群落繁盛的底栖环境的沉积特征起到了不可替代的作用。
对于如此规则的包壳结构, 尤其是明显富含有机质的球粒对泥晶核心形成包裹, “ 滚雪球” 式的形成方式很难解释。而Gerdes 等(1994)通过对微生物席内底栖鲕粒的研究, 认为丝状蓝细菌在鲕粒纹层的形成过程中可能发挥了巨大的作用。这一研究结果亦被Brehm 等(2006)通过实验室微生物培养所证实, 即丝状蓝细菌的菌体可以包裹核心生长, 并通过其生命活动来改变附近微环境的物理化学条件, 如水体的pH值以及微区域内的CO2梯度, 从而诱导文石的沉淀。从这个角度上看, 研究区徐庄组鲕粒灰岩二段上部这种暗色颗粒形成的包壳结构可能就是正在形成中的鲕粒, 而呈球状或短链状的暗色颗粒可能正是蓝细菌菌体的残余。
如前所述, 在单晶鲕最外部大都发育1层暗色环边(图 4-E), 其可能由黄铁矿颗粒组成, 并可能含较多的有机组分。但是, 发育这种黑色包层的鲕粒其内部往往较为洁净, 故推测交代或重结晶作用可能是从内向外进行的, 从而使原始鲕粒中的暗色矿物及有机质向外迁移。当这种作用较为强烈时, 鲕粒内部的黄铁矿及有机组分等便会被排斥到结构的最外层, 从而形成暗色环边。
此外, 单(或多)晶鲕外部边界很多还是保留了较为平滑的外表面, 也未发生强烈的变形或是破碎, 这表明鲕粒所遭受的改造应该是在一种相对稳定的环境中发生的。因此, 需要解释的问题变为这些颗粒的原始鲕粒究竟具有什么样的特征, 而使其率先被改造。Gerdes 等(1994)在研究 Lanzarote 地区微生物席内的碳酸盐颗粒的时候, 将其中的一种由近乎透明的纤维状胶结物包裹富有机质球状核心所构成的球状颗粒命名为corcoids。这种颗粒并没有明显的同心纹层; 有机质在其核心高度富集(Gerdes et al., 1994)。考虑到 corcoids中的纤维状胶结物所含的杂质少, 同时相对于泥晶方解石来说, 其结晶程度更高, 因此在后期的成岩改造过程中, 相对于其他类型的碳酸盐颗粒, 其可能更容易发生重结晶从而形成大颗粒方解石晶体。corcoids结构特征与在徐庄组鲕粒灰岩中发现的一些单(或多)晶鲕相似, 主要体现在高度富集有机质的核心及近乎透明的包裹核心的皮壳(图 6-C), 故徐庄组中发育的单(或多)晶鲕可能正是这种 corcoids 的残余结构。值得一提的是, Gerdes等(1994)认为corcoids是富有机质的球状核心在微生物群落内的胶状物质中独立发育的产物, 他们认为 Flü gel 等(1982)提出的在缺乏扰动的环境中的“ 建造性” 微晶方解石的增生, 能够很好地解释这种颗粒的形成。
凝集颗粒是一种由方解石(文石)或白云石的晶粒聚合而成的球状或类球状颗粒(图 6-E, 6-F), 但在同一颗粒中, 以上2种晶粒未见同时出现。虽然这2种凝聚颗粒总体发育较少, 但是却呈现出局部富集的特征, 尤其是由白云石晶粒组成的凝聚颗粒, 甚至可以构成一些同心— 放射状鲕粒的核心并集中出现。
由白云石晶粒组成的凝聚颗粒总体上形态规则, 粒径多为250~500 μ m; 其中白云石晶粒多为自形— 半自形晶, 粒径多为10~50 μ m, 且大多数晶粒的边部都见有机质的富集(图 6-E)。由于白云石晶粒可能是后期成岩作用改造的产物, 故其原始结构可能是一种与鲕粒不同的、由一些富有机质的碎片物质相互粘合而成的球状颗粒。
由方解石晶粒组成的凝聚颗粒具有更加平滑的外表面, 其中的方解石(文石)晶粒呈柱状, 杂乱排列, 粒度大小不均匀, 且晶粒之间的空隙为富有机质泥晶所充填(图 6-F)。由于柱状晶粒排列松散、杂乱, 像是被限制而发育在一个球状空间内, 故推断这些呈柱状的方解石(文石)晶粒可能是在球状结构已经形成之后而在其内部生长起来的。虽然这种方解石(文石)聚合颗粒外边界平滑, 却存在明显的对这种球状结构加以限制的外壳(图 6-F), 因此, 这种球状结构很可能形成自一种球状空腔。Chafetz 等(1991)曾在热泉微生物群落内发现了一种存在于碳酸盐内的气泡, 他们认为这种气泡形成于自养细菌光合作用所释放出的氧气。由于气泡内外的 CO2分压的差异, 会导致文石沿着气泡的外边界发生沉淀。Gerdes 等(1994)也认为这种作用可以诱使文石沿着气泡沉淀, 从而使鲕粒皮层的形成成为可能。笔者认为, 方解石(文石)柱状晶体的存在可以很好地解释气泡内部的变化。由于异养生物的存在, 呼吸作用使得气泡内的 O2逐渐消耗, 并生成 CO2, 导致气泡内的压力降低。气泡内部的空腔为矿物的沉淀提供了相对宽裕的生长空间, 方解石(文石)晶体便可以较自由地生长, 最终形成一种由杂乱排列的柱状晶体所填充的结构。有机体死亡后充填在晶粒间的空隙中, 形成富有机质的填充物。
研究区徐庄组部分鲕粒内部发育有由黄铁矿晶粒聚集成的暗色核心(图 6-G), 而该暗色核心外部的同心纹层多由泥晶及菱形白云石晶粒构成。这些黄铁矿颗粒可构成形状规则的同心层或是球状结构, 其与Brehm等(2006)在实验室培养出的微生物菌落形成的球状颗粒极为相似。
在一些复合鲕中, 除了可以见到黄铁矿小颗粒在复合鲕子颗粒的中心富集, 还可以见到黄铁矿聚集体沿子颗粒的接触部位发育(图 6-H)。这些沿着鲕粒边界发育的黄铁矿颗粒反映了在该部位有硫酸盐还原细菌的发育, 这可能对鲕粒聚合形成复鲕起到了粘结作用。因此, 在徐庄组复鲕的形成过程中, 微生物作用可能在鲕粒的粘合过程中发挥了更为主要的作用。
在徐庄组鲕粒灰岩中, 笔者发现一些生物碎屑的空腔中包裹了球状的碳酸盐颗粒(图 7-A), 这些颗粒的粒径与该层段鲕粒相比较小, 发育有不完善的同心纹层。同时, 在生物碎屑的边部见附着体(图 7-A, 7-B)。从形态特征上来看, 该附着体的外边界平滑, 且发育1层富有机质的泥晶环边; 在其与生物碎屑的接触部位, 可见有机质的明显富集; 二者的接触部位呈弧状平滑过渡。在附着体内部, 发育有4个呈椭球状的富有机质颗粒, 这些颗粒的粒径十分接近, 结构特征也基本一致。在徐庄组中发现的附着体与 Harrison 等(2005)描述的微生物膜(biofilm)极为相似。其内部的球状颗粒虽然较小, 但具有鲕粒的典型特征。笔者认为, 这一结构很可能就是一个附着在底栖生物外壳或生物碎屑上的小型微生物群落。其内部发育的呈球状的暗色颗粒虽然未见清晰的同心纹层, 但是其形态特征及矿物组成与该层段鲕粒相似, 因此很有可能就是微生物作用下形成的鲕粒的初始结构。
 | 图7 北京下苇甸剖面寒武系徐庄组生物碎屑及其上的附着体Fig.7 Bioclast and special attachment of the Cambrian Xuzhuang Formation at Xiaweidian outcrop in Beijing |
微生物膜一般是由多种微生物所构成的一个共生综合体, 这些微生物通过各自的生命活动, 对其他微生物的生命活动产生影响, 并对水介质的物理化学条件起到了局部的控制作用。Brehm 等(2006)研究并分析了微生物作用诱导方解石沉淀的机制, 认为球状的微生物席或生物膜(biofilm)体系有利于微生物生命活动的进行, 这主要体现在以下几个方面:(1)保护膜内的微生物免遭外部机械作用的干扰及破坏; (2)促进微生物的增殖; (3)促进光、矿物质及一些其他资源的吸收和利用。硫酸盐还原细菌等异养微生物通过呼吸作用, 消耗由蓝细菌等自养微生物所合成的有机物(表 1, Eq.1), 并释放 CO2, 同时还原SO42-并生成 H2S, 这一过程会对其周围水体的pH值及 CO2分压起到很大的改变(表 1, Eq.2, Eq.3), 从而促进了方解石的沉淀。在鲕粒内部大量发现的黄铁矿晶粒正是这种作用发生过的有力例证, 在徐庄组中发现的附着体很可能就是一个附着在底栖生物外壳或生物碎屑上的小型微生物群落。
| 表1 微生物膜内微生物共生体系的生物— 化学模型 Table1 A bio-chemical model of symbiotic systems in microbe biofilm |
核心往往被认为是鲕粒形成的必要条件之一。高能动荡的水体环境可以促使同心纹层包裹核心逐步增生, 从而形成鲕粒。前已述及, 在徐庄组鲕粒灰岩中发育一种较为特殊的有核同心— 环状鲕粒(图 3-D), 其核心是由含有机质很少的微亮晶方解石构成, 因而在显微镜下表现为一个明亮的被暗色的富有机质鲕粒皮层包裹了的核心。但是, 这一所谓的“ 核心” 为较为纯净的微亮晶方解石, 更像是后期充填的产物。如果这个核心是在鲕粒皮层形成之后被方解石填充形成的话, 那么鲕粒皮层的形成方式便不能用一个动荡水体环境下的机械— 化学作用模式来解释。这一看似矛盾的现象利用前人对微生物群落的研究成果得到了较为有效的解释。Theobold(1852)曾在微生物席内首次发现了被碳酸盐所包裹的气泡, Chafetz 等(1991)也在热泉石灰华中发现了类似的气泡。他们一致认为这些被碳酸盐所包裹的气泡是光合细菌所释放的氧气形成的。Gerdes 等(1994)在对 Lanzarote 地区微生物席内的碳酸盐颗粒开展了系统的研究后, 对他们的这种论断表示了肯定。由于气泡内富集氧气, 因此 CO2的分压则相对较小, 这就造成了在该气泡附近的微区域内具有一个较大的 CO2梯度, 由于气泡边界处的水体缺少 CO2, 从而大大提高了该部位 CaCO3的饱和程度, 这会导致文石沿着该气泡边界晶出, 这一过程逐渐向外部发展, 则形成了鲕粒皮层。在整个过程中, 气泡作为核心存在。此外, 由方解石晶粒所构成的凝聚颗粒也可能是气泡被填充的产物, 柱状方解石晶粒则是晶体较自由生长的有力证据。由此可见, 微生物席内由微生物生命活动所产生的气泡的存在似乎为解释鲕粒的成核提供了一个新途径。
徐庄组泥晶鲕往往并没有明显的核心, 外表面不平滑, 与 Gerdes 等(1994)所描述的底栖鲕粒的典型特点相吻合。在徐庄组内部, 泥晶鲕往往与其他类型鲕粒相伴产出, 甚至有些鲕粒的核心表现出了与泥晶鲕相似的特征。由于泥晶鲕总体上发育较少, 因此不排除其可能是由水流的搬运携带而来, 若是这样的话, 泥晶鲕的存在部位则不能反映其原始发育环境的特点。因此, 需要结合与泥晶鲕相伴出现的其他鲕粒的特征来综合探讨其形成环境。在泥晶鲕集中发育的部位, 有机质含量整体较高, 并见有一些暗色的球状颗粒, 这可能反映了底栖环境下一个由微生物作用所控制的鲕粒产出“ 工厂” 。
一个普遍的事实是, 在很多浅海、高能且CO2过饱和的海洋环境中缺乏鲕粒的存在(Pacton et al., 2012), 因此, 在鲕粒形成的初期, 还必然存在一些其他的作用因素为鲕粒的成核创造条件。徐庄组中的特殊组构及鲕粒的发现, 为微生物在鲕粒的成核过程中可能发挥的巨大作用提供了有力的例证。因此, 笔者认为, 微生物作用可以诱导鲕粒成核, 同时, 在形成鲕粒同心纹层的过程中, 具有一定刚性的异化颗粒的存在不再是必要条件。
如果地质历史时期的鲕粒可由微生物作用大量形成, 那么, 鲕粒的出现所具有的环境指示意义将会与机械— 化学作用模式下给出的环境条件解释有一定的出入(Pacton et al., 2012)。就鲕粒的“ 滚雪球” 成因模式而论, 鲕粒的形成似乎离不开一个较为动荡的水体环境(Davis et al., 1978; 梅冥相等, 2011)。动荡的水流有利于CO2脱气作用的进行, 从而造成海水的碳酸盐超饱和, 水体中的 CaCO3便可以围绕核心析出(梅冥相等, 2011; Woods, 2013)。但对于微生物作用所主导的鲕粒的形成过程, 这种较为动荡的水体环境的认识似乎不能完全成立, 故徐庄组中鲕粒的发育所反映的可能并不是一个典型的高能滩相环境。原因如下:
1)由于微生物如丝状蓝细菌的生存需要一个相对稳定的水体环境, 且出于光合作用的需要, 这些微生物群落应发育于透光性相对较好的地区, 这就要求水体不能太深同时应相对清澈, 不宜有大量悬浮物。
2)徐庄组为数不多的复合鲕粒(图 5-E, 5-F), 其外边界形态复杂, 包裹层厚度也很小, 子颗粒的接触并不紧密(图 5-F)。这就产生了这样一个问题, 它们是否能够经受较强的水流冲击以及其他碳酸盐颗粒的碰撞。答案显然是否定的。首先, 就其形成来看, 有机质似乎对于单一鲕粒的相互粘合起到了重要的作用。其次, 这些由单个鲕粒聚合而成的复鲕, 虽然其结合的并不紧密, 但是依然保存了下来, 这不仅表明其形成环境是相对平静的, 甚至说其在形成之后也并未经过水流的强烈改造或是搬运。
3)泥晶鲕作为徐庄组内的一种主要的鲕粒类型, 其核心的缺失很难用滚动来对其形成加以说明。徐庄组泥晶鲕由富有机质的泥晶所构成, 除了整体呈一个较规则的球体外, 并不存在其他特殊的内部结构。故泥晶鲕很可能是由微生物直接作用而成。当然, 具体的作用方式尚有待探讨。
文中着重对北京西山下苇甸寒武系徐庄组上部厚层鲕粒灰岩中的鲕粒进行了系统的研究和分析, 并对其中的有核同心— 放射状鲕、无核同心— 放射状鲕、单(或多)晶鲕、泥晶鲕以及复合鲕粒进行了较为详细的描述。同时, 还发现了一些较为特殊的组构及现象, 包括:(1)由暗色矿物颗粒构成的单层包壳结构及其附近的暗色球粒; (2)单(或多)晶鲕边部的富有机质残余; (3)方解石(文石)及白云石晶粒凝聚颗粒; (4)鲕粒内部的黄铁矿聚集体; (5)具有特殊结构的复合鲕粒; (6)生物碎屑上的附着体。这些组构的发现, 与国内外一些学者在野外及实验室研究过程中所观察到的一些微生物作用产物有着极大的相似性。就目前的研究结果来看, 徐庄组鲕粒的形成可能与微生物作用有着巨大的联系, 徐庄组中鲕粒的发育所反映的可能并不是一个典型的高能滩相环境。
致谢 研究过程中得到了梅冥相教授的悉心指导和帮助; 王楚楚、冯定素等参与了野外工作; 显微照相得到了杨立强教授的大力支持。两位审稿专家对本文的修改给出了十分宝贵的意见。冯增昭教授对本文的进一步修改给予了作者极大的鼓舞和指导。文章的完成离不开以上各位的帮助, 作者在此一并致以衷心感谢!
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|