
{kind=link}
{kind=link}
光合作用的起源:一个引人入胜的重大科学命题
[梅冥相 , 高金汉]
, 高金汉]
, 高金汉]
|
|

第一作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作。E-mail: meimingxiang@263.net。
光合作用的起源是一个非常古老的事件,对这个事件的证据,包括基本生物学过程的开启和发展之类的问题,如地球早期光合作用生物的属性以及光合作用生物如何获取光合作用装置等,可能已经消失在时间的长河之中;因此,光合作用起源就成为一个引人入胜的重大科学命题。尽管如此,地质学、生物地球化学、比较生物化学和分子进化分析,为光合作用起源及其复杂的进化历史,提供一些新认识和新线索,主要涉及到以下3个方面:(1)光合作用生命的起源;(2)光合作用装置的起源;(3)光合作用催化剂的起源。追索科学家们对这一重大科学命题的持续研究与艰苦努力,以及所取得的一些重要而且富有智慧的认识,将为今后的深入研究提供一些重要的思考途径和研究线索。同时,追逐光合作用起源的研究进展,对于深入了解早期地球复杂的圈层耦合过程也具有重要意义。这些作用过程主要包括:(1)从不生氧光合作用到生氧光合作用的转变;(2)大气圈与生物圈之间复杂的相互作用和协同进化;(3)生氧光合作用起源与进化所造成的、从一个缺氧的大气圈到今天含氧大气圈的复杂演变过程;(4)大气圈和水圈的渐进氧化作用对地球表面环境以及生命的起源和发育所造成的一个长时间影响;(5)早期地球表层古地理面貌的成型等。更为重要的是,对光合作用起源的地质学尤其是沉积学思考所得出的一些重要认识,尽管不是结论,但是拓宽了沉积学的研究范畴,开阔了沉积学家的研究视眼,同时也成为一个多学科协同作战的范例。
About the first author Mei Mingxiang,born in 1965, graduated and obtained Ph.D. degree from China University of Geosciences(Beijing) in 1993. Now he is a professor at School of Earth Sciences and Resources,China University of Geosciences(Beijing),and is engaged in sedimentology and stratigraphy. E-mail: meimingxiang@263.net.
The origin of photosynthesis is such an ancient event that definitive evidence as to how this essential biological process began and developed may be lost forever,such as the nature of the earliest photosynthetic organisms as well as the problem dealing with how photosynthetic organisms obtain their photosynthetic apparatus;thus,studies on the origin of photosynthesis became an important and enchanting scientific theme. Nevertheless,some evidence can be assembled from a variety of sources,including geology,biogeochemistry,comparative biochemistry and molecular evolution analysis,which begin to give some insights into the complex origin and evolutionary history of photosynthesis that chiefly deal with several aspects as follows:(1)the origin of photosynthetic life;(2)the origin of photosynthetic apparatus;and(3)the origin of photosynthetic catalysts. It is very meaningful to pursuit and to summarize some important cognitions enriched-wisdom resulted from both the lasting research and hardworking effort by scientists,which can provide important studying clues and thinking approaches for the further researches of this fascinating scientific. Furthermore,it is also meaningful to trace the advancement on the study of the origin of photosynthesis for the further understanding of the sophisticated coupling process among earth spheres in the early time,which are as follows:(1)the transformation from the anoxygenic to the oxygenic photosynthesis;(2)a complex interaction and its synergetic evolution between the biosphere and the atmosphere;(3)a sophisticated evolutionary process from a anoxic to an oxic atmosphere generated by the origin and evolution of the oxygenic photosynthesis;(4)a major long-term effect on Earth-surface environments and the development of life that is benefited from the progressive oxygenation of the atmosphere and hydrosphere;and(5)the molding of the palaeogeographical version of the early Earth and so on. More importantly,some geological especially for sedimentological thinking on the origin of the photosynthesis and some important cognitions,though are not conclusions,have broaden the studying domain of sedimentology and widen the field of view of sedimentologists. Ultimately,these researches become a scientific paradigm that is applied with multi-subjects.
光合作用是地球上最为重要而且古老的生物作用过程, 可将光能转变为生物所需要的化学能, 能进行光合作用的生物被称为光养生物(phototroph)。实际上, 生物学家多年的研究已经高度认识到“ 光合作用是光养生物从光中获取能量, 产生三磷酸腺苷(ATP)、并消耗ATP和利用还原型辅酶(NADH)还原二氧化碳的复杂反应过程, 从而完成生命的光合营养途径” (Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Nisbet et al., 1995; 徐桂荣等, 2005; Madigan et al., 2006; Nisbet et al., 2007)。ATP是一种由腺苷衍生的核苷酸(C10H16N5O13P3), 通过ATP的水解作用生成的二磷酸腺苷, 可为细胞进行各类生化过程提供大量能量, 包括肌肉收缩及糖分释放; NADH是一种还原形式的辅酶(AND; C21H27N7O14P2), 这种辅酶存在于大多数活着的细胞中, 而且在各种新陈代谢过程中, 还可以作为氧化剂或还原剂交替使用。
光氧生物以光作为能量来源、运用二氧化碳和水合成碳氢化合物、并释放氧气而且将其作为副产品, 这就是生氧光合作用(oxygenic photosynthesis)。生氧光合作用过程, 发生在具有某种叶绿素的生物中, 这个过程包括:(1)光解作用(Photolysis), 生物体内的叶绿素和其他类型的色素, 利用光将水分解并释放出氧气, 完成以下反应, 即2H2O→ 4H+O2; (2)固碳作用, 把氢与二氧化碳化合成碳水化合物, 完成以下反应, 即CO2+2H2→ (CH2O)+H2O。综合起来, 这就是著名的光合作用方程:H2O+CO2→ (CH2O)+O2。绿色植物、藻类、蓝细菌等生物, 利用水作为一种较弱的电子授体并作为还原动力, 氧化水分子并释放出氧气, 而且氧气作为副产品; 需要注意的是, 生氧光合作用生物还原动力的产生以及能量保存均需要光, 从而区别于不生氧光合作用(anoxygenic photosynthesis)。
不产生氧气的光合作用被称为不生氧光合作用, 不生氧光合作用发生在具有某种细菌叶绿素的生物中, 不生氧光养生物从类似于硫化氢之类的物质中获得电子授体和还原动力, 并产生NADH, 最终完成生命的光和营养途径(Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Nisbet et al., 1995; 徐桂荣等, 2005; Madigan et al., 2006)。不生氧光合作用的最古老的形式, 运用还原性的硫化物作为固定二氧化碳的电子授体(Olson and Pierson, 1986; Olson and Blankenship, 2004)。
光合作用装置的主要单元包括反应中心、触角复合体、电子授体的复合体和固碳作用装置(Weise and Koegh, 1977; Greenwood and Edward, 1979; Olson and Pierson, 1986; Brock et al., 1994; Olson and Blankenship, 2004; Madigan et al., 2006)。对于光合作用来讲, 关键的催化剂是二磷酸核酮糖羧化酶, 常常被简化定义为Rubisco; Rubisco的特异性(对二氧化碳的偏爱超过氧气), 会促使生命从环境中的二氧化碳获取碳(Nisbet et al., 2007)。
化石证据表明, 蓝细菌存在于2.5 Ga至2.6 Ga之前(Olson and Blankenship, 2004), 或者出现在更加古老的时代(Nisbet et al., 2007; Buick, 2008; Guo et al., 2009; Holland, 2009; Farquhar et al., 2010; Kah and Bartley, 2011; Schopf, 2011; Czaja et al., 2012; Crowe et al., 2013; Riding et al., 2014); 更为重要的是, 在蓝细菌出现之前, 肯定存在各种形式的不生氧光合作用细菌(Olson and Blankenship, 2004)。
鉴于上述诸方面, 追逐和研究光合作用起源, 主要涉及到以下3个方面的问题(Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Olson and Blankenship, 2004; Nisbet et al., 2007; Buick, 2008; Holland, 2009; Farquhar et al., 2010; Kah and Bartley, 2011; Schopf, 2011; Czaja et al., 2012; Crowe et al., 2013; Riding et al., 2014): (1)光合作用生命的起源; (2)光合作用装置的起源; (3)光合作用催化剂的起源。对光合作用起源的地质学约束(Nisbet et al., 2007; Buick, 2008; Holland, 2009; Farquhar et al., 2010; Schopf, 2011; Riding et al., 2014), 为生物学角度研究光合作用起源提供时间约束方面的基本材料。
光合作用的起源, 是地球上最为重要而且最为古老的生物作用事件(Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; 徐桂荣等, 2005; Madigan et al., 2006), 这一点已经取得共识(Nisbet et al., 2007; Buick, 2008; Holland, 2009; Farquhar et al., 2010; Schopf, 2011; Riding et al., 2014); 但是, 光合作用起源的证据, 即这个基本的生物学过程是如何开始而且如何发展的问题, 可能消失在时间的长河之中(Olson and Blankenship, 2004)。让人感到欣慰的是, 一些光合作用起源的证据还是可以从各种来源中收集到, 包括地质学、生物地球化学、比较生物化学和分子进化分析, 这些均能对光合作用的复杂起源和进化历史提供一些新认识及新线索, 尤其是对光合作用生命起源的了解。
大约在40多年以前, Olson(1970)就认为, 祖先式的“ 光合作用者” 可能已经运用光来驱动那些有机化合物的同化作用, 残留在化学进化作用的生命起源之前的有机化合物之中。自从20世纪70年代至今, 因为对早期地球H2、CH4和NH3的含量水平估计, 得到一定程度的下降(Kastings and Brown, 1998), 所以, 生命起源的有机汤假说(the organic soup hypothesis; 有机化合物被认为合成于还原性质的无机化合物的假说), 没有得到完全认可(Olson and Blankenship, 2004)。在有机汤合适浓度缺乏的条件下, 似乎祖先性的光合作用生物运用外来的还原剂如硫化氢来固定二氧化碳; 也就是说, 在能够进行生氧光合作用的蓝细菌出现之前, 肯定存在各种形式的不生氧光合作用细菌(Olson and Blankenship, 2004); 更进一步讲, 在早期地球的某一个时间, 发生了从不生氧光合作用到生氧光合作用的重大转变。尽管很多细节问题还不清楚, 但是, 科学家们还是取得了以下共识:生氧光合作用曾经一度在生命历史中进化, 而且所有今天的生氧光合作用均发生在蓝细菌以及在真核生物细胞(蓝细菌的后代)的叶绿体之中(Blankenship et al., 2007; Farquhar et al., 2010; Schopf, 2011; Olson et al., 2013; Riding et al., 2014)。
对于光合作用生命的祖先, 存在3个主要的线索:化学标志物、叠层石化石体和古代生物的显微化石(Olson and Blankenship, 2004; Nisbet et al., 2007; Buick, 2008; Holland, 2009; Farquhar et al., 2010; Schopf, 2011; Riding et al., 2014)。
在化学标志物之中, 沉积有机碳(干酪根)中的13C/12C值表明, 生物成因的二氧化碳的固定作用, 可以追溯到3.5~3.8 Ga(Schidlowski, 1988)。有机碳是亏损13C的, 然而, 无机碳则不是。碳的这2种形式之间的差异, 表现为样本之中的13C的相对亏损。地质记录对有机碳表现出一个连续的-27‰ ± 7‰ 的值, 对于无机碳则为+0.4‰ ± 2.6‰ ; 这个记录与RuBP羧化酶所主导的生物二氧化碳固定作用(Schidlowski, 1988; Olson and Blankenship, 2004)相一致, 但是, 与其他的固碳作用机制也相吻合, 所以说, 有机碳的同位素记录还是不能肯定二氧化碳固定作用究竟是否是光合作用成因的。就像Mojzsis 等(1996)曾经认为的那样, 来自于格陵兰的岩石中的3.8 Ga的古老的碳质包裹物与自养微生物的二氧化碳固定作用相吻合; 但是, 对这些结构的生物成因和起源的解释, 又受到一些学者的质疑(Olson and Blankenship, 2004)。也就是说, 要寻求明确的光合作用生命起源的地球化学证据, 还有待于今后的深入研究, Eigenbrode 等(2008)对2720~2560 Ma的生氧光合作用蓝细菌的碳水生命标志物的确定和研究, 代表了一个成功的实例。
叠层石, 是由交互式的造席生物和沉积物所形成的纹层组成的纹层状沉积构造(Reid et al., 2000, 2011; Riding, 2000, 2011; Gerdes, 2010; 梅冥相, 2011, 2014), 是微生物群落对沉积颗粒的捕获、黏结和沉淀作用的连续性过程的增生物(Awramik, 1977), 紧接着发生了胶结作用(Knoll, 2008); 这3个增生作用机制之中, 在沉积记录中最具有说服力的是沉淀作用, 不管这种沉淀作用是生物成因的还是非生物成因的。现代叠层石总是包含有丝状光合作用细菌如蓝细菌, 从而表现为蓝细菌生物席的建造物, 但是, 现代蓝细菌微生物席可能与凝块石状的微生物岩相关、而不是与太古宙的叠层石相联, 原因是太古宙叠层石的微结构确实不支持一个作为蓝细菌生物席的建造物的结论(Chacó nb et al., 2011)。由于大多数古代叠层石的生物成因常常受到质疑, 所以说, 与可能的光合作用生命的早期形式相关的叠层石证据还得不到确定(Olson and Blankenship, 2004; Riding, 2008, 2011)。
蓝细菌是否是太古宙叠层石的建造者还存在着争论, 但是它们对地球历史的地球生物学影响是独特的而且还是关键性的(Chacó nb et al., 2011):在前寒武纪早期的某些点上, 它们释放出有毒的、但是在现在确实是维持生命所必需的氧气, 从而永久性地改变了地球生物进化的途径; 再者, 澳大利亚西部3.43 Ga的叠层石生物礁的研究表明, 这些古老的叠层石还是应该解释为微生物成因(Allwood et al., 2006; Awramik, 2006), 甚至是蓝细菌生物席的建造物(Altermann et al., 2006)。因此, 尽管还难以确定以蓝细菌为代表的光合作用生命起源的确切时间点, 但是, 对太古宙叠层石的研究, 还是为研究光合作用生命起源提供了重要线索。
Allwood 等(2006)对澳大利亚西部Pilbara 克拉通3.43 Ga的叠层石的研究, 认为是一个包含着可能的生物成因纹层状构造(叠层石)的沉积岩建造(图 1)。他们对形成在特定古环境中的叠层石进行特定模态分析, 从而拒绝了这些叠层石形成作用的非生物假说; 最后, 他们还证明了叠层石的多样性、复杂性及其与产出环境的联系, 得出了这些叠层石属于微生物成因的结论, 而且这个结论还得到了Awramik(2006)的充分肯定。更为重要的是, 上述认识和结论不但得到了Altermann(2008)的强烈推崇, 即太古宙叠层石中多样化的生物沉积构造的构架与形式, 难以用纯粹的物理或化学沉积物堆积作用进行解释, 从而成为无可辩驳的微生物成因; 更为重要的是, 还得到了Bosak 等(2009)以及Petroff 等(2010)的印证和支持, 也与Riding 等(2014)关于“ 太古宙氧气绿洲” 的研究所得出的结论相一致。
 | 图1 澳大利亚西部Pilbara克拉通3.43 Ga的锥状叠层石(据Awramik, 2006)Fig.1 Image showing conical stromatolites of 3.43 Ga from Pilbara Craton in western part of Australia(after Awramik, 2006) |
Bosak等(2009)认为, 锥状叠层石是早到3.5 Ga的水生环境之中的光合作用和趋光性微生物存在的坚实标志。他们论述到, 只是趋光性本身还难以解释很多太古代、早元古代和中元古代锥状叠层石脊部的破损、弯曲和卷曲的纹层; 但是, 在现代锥状叠层石聚合体尖端, 所观察到的蓝细菌氧气生产作用, 以及这种作用产生的扭曲纹层、亚毫米级别到毫米级别的被缠住的气泡, 以及这些相似大小的化石气泡和扭曲纹层, 似乎只是出现在一些锥状叠层石的脊部带, 这些叠层石老到2.7 Ga或者更古老。这就意味着, 蓝细菌不但建造了元古代早期的锥状叠层石, 这类叠层石生长可能包含着生氧光合作用进化的时间进程, 而且超前于大气圈在2.4 Ga左右的初始氧化作用的地球化学记录(Canfield, 2005; Holland, 2006)。
Petroff 等(2010)基于生物物理学原理, 建立一个定量关系来探索锥状叠层石的几何形态及其与生物物理过程之间的成因关系; 而且还注意到, 所有现代的锥状叠层石以及很多至少形成在2.8 Ga的锥状叠层石, 均表现出一个在相邻构造之间的特征性空间间距, 这个突出的特征代表着锥状叠层石的组织化作用, 被考虑为锥状叠层石之间调节性竞争的扩散作用过程的结果。他们还进一步认为, 很多太古代的小型锥状叠层石, 还是应该识别为地球最早期光合作用群落演化的标志; 锥状叠层石与一些现代微生物席之间的定性化的相似性, 表明了古代锥状叠层石的光合作用成因。
Riding 等(2014)基于加拿大Steep Rock一带大约2.8 Ga的浅海灰岩和深水富铁沉积物的稀土元素(REE)研究, 表明了广泛沉淀着浅水灰岩的海水是氧化的, 然而相邻的形成富铁沉积物的深水则是缺氧的。他们还进一步认为:(1)通过可溶解铁种类的氧化迁移, 氧气促进了灰岩的沉淀作用, 即从可溶解的二价铁(Fe2+)氧化成不可溶解的三价铁(Fe3+)的强氧化物, 而且估计到在海水中至少是10.25 μ Mol的氧气浓度, 这对于Steep Rock灰岩中的这个作用过程的完成是必须的, 说明了在太古宙海洋中可溶解Fe2+的丰富供应可能会阻碍灰岩建造的沉淀作用; (2)尽管不存在氧气来源的直接证据, 但是, 在Steep Rock灰岩中的多样化的叠层石和有机碳同位素, 表明了蓝细菌的存在, 从而支持了以下观点, 即在太古宙时期明显的氧气水平首先发育在营养物受到局限的浅海生境中, 而且这些促进灰岩沉淀作用的浅海生境在空间上是局限的、并且是瞬时性的; (3)如果太古宙海相灰岩总体上反映了在缺氧富铁海洋的边缘可溶解铁的氧化迁移, 那么早期的“ 氧气绿洲” 就要比以前所假定的清楚得多。
被接受为真实的古代蓝细微菌化石的时代大致为2.0 Ga(Schopf et al., 2007)。大于2.5 Ga的蓝细菌状生物的存在证据, 是发现于古代岩石中的化学生命标志物2-甲基荷烷(2-methylhopanoids; Summons et al. 1999); 但是, 只是根据生命标志物, 还不能够确定这些蓝细菌祖先是否真正进行着生氧光合作用。
Schopf 和 Walter(1983)以及Awramick 等(1983)在澳大利亚西部的太古宙地层中(2.8 Ga的Fortescue 群和3.5 Ga的Warrawoona 群), 识别出4种类型的丝状细菌化石, 而且认为其中的一些是光合作用的微生物。基于形态学特征以及地球化学的进一步研究, 较为肯定地认为这些光合自养生物(photoautotrophs), 可能曾经是古代造成氧气演变的蓝细菌(Schopf et al., 2007; Schopf, 2011)。来自于澳大利亚的发现以及深入研究得出的认识, 还得到了来自于南非2.5~2.6 Ga的蓝细菌化石的发现与研究的印证(Kazmierczak and Kremer, 2002; Altermann et al., 2006; Kazmierczak et al., 2009)。
Kazmierczak和Kremer(2002)展示了在南非获取的2.5~2.6 Ga的蓝细菌化石证据(图 2); 在此基础上, Altermann等(2006)进一步提出并认识到:(1)蓝细菌能够在其有机包裹物之中储存钙离子和镁离子, 从而在它们的鞘和细胞外聚合物质内沉淀碳酸盐, 因此, 这些作用机制就会驱动沉积碳酸盐的生产作用; (2)蓝细菌和异养细菌的相互作用, 对于地球历史中至少3.0 Ga的碳酸盐工厂的形成, 具有主要的贡献。Kazmierczak 等(2009)后来的进一步研究, 尤其与来自于土耳其Lake Van地区的现代微生物席中的蓝细菌矿化组构的对比(图 2), 认为:南非新太古代Nauga组的沉积碳酸盐岩中的硅化微生物席, 由那些与现代底栖繁殖的蓝细菌(Chroococcales 和 Pleurocapsales)相似的微生物群所构成; 它们的残余物, 由蒴果状的包裹物所组成, 还包括黏性的鞘, 以及具有Al-K-Mg-Fe硅酸盐混合物的钙碳酸盐矿化的细胞组合。围绕着单个细胞和形成菌落的细胞束状体的黏性鞘的蒴果状组织化作用, 以及矿化作用样式, 成为新太古代所描述的微生物群及其现代类比物的普遍特征。所以说, 蓝细菌不但通过影响地球的大气圈构成而成型在地球表面, 而且给地球带来了一种全新的岩石类型。
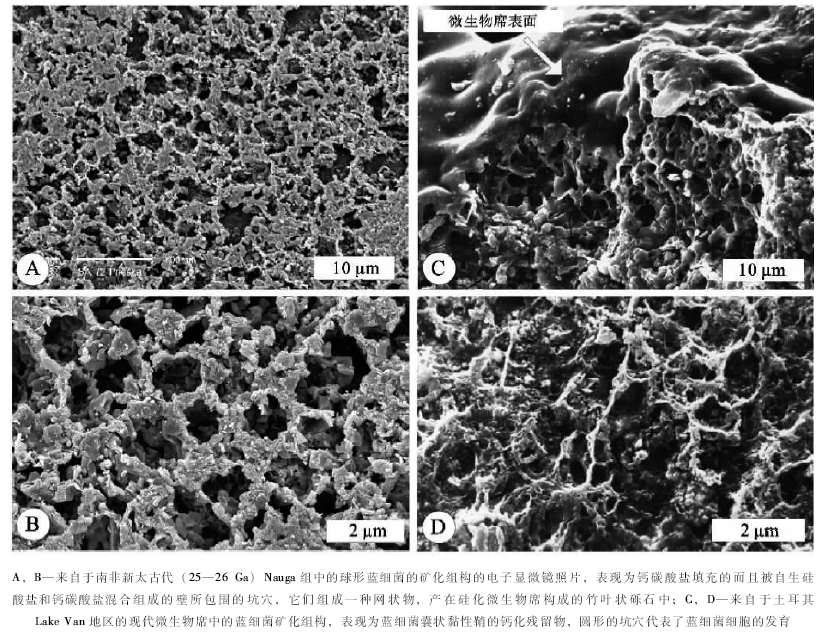 | 图2 现代与太古代球形蓝细菌聚合体的鞘和囊状物的对比(据Kazmierczak et al., 2009)Fig.2 Comparison of sheaths and capsules of coccoid cyanobacterial aggregates between those from the Neoarchean(A and B)and those of the modern(C and D)(after kazmierczak et al., 2009) |
综上所述, 可以肯定的是, 早期地球本质上是缺氧的, 很多标志表明了2.7, ~3.0 Ga的生氧光合作用的存在。大气圈和水圈的渐进氧化作用, 对地球表面环境以及生命的起源和发育, 具有一个长时间的影响, 而且这个氧化作用的开始主要归因于太古宙蓝细菌生氧光化作用的发展(Canfield, 2005; Holland, 2006, 2009)。在2.7~3.0 Ga前的生氧光合作用的存在(Canfield, 2005; Nisbet et al., 2007; Buick, 2008; Schwartzman et al., 2008; Schopf, 2011), 主要表现在以下几个方面(Riding et al., 2014):(1)在2920~2980 Ma的沉积物中, 铬同位素和氧化— 还原敏感的痕迹金属元素的存在(Crowe et al., 2013); (2)2760 Ma前的较负的碳同位素值, 说明了甲烷氧化作用的存在(Hayes, 1994); (3)2720 Ma的叠层石中磷酸盐化丝状体所表征的可能的丝状蓝细菌的存在(Buick, 1992);
(4)表明2720~2560 Ma的生氧光合作用蓝细菌的生命标志物(Eigenbrode et al., 2008); (5)2680 Ma的沉积物中, 可以对比的铁和钼的同位素构成(Czaja et al., 2012); (6)在2.7~2.5 Ga的条带状铁建造(BIF)中的菱铁矿, 及其具有的δ 56Fe负值是由于铁的氧化作用所致, 从而作为生氧光合作用进化的直接结果(Johnson et al., 2008)。多数早期的自由分子氧气(O2), 被认为通过与海水中可溶解铁的反应而消失, 这主要归因于深水背景中富铁和富硅的条带状铁建造的沉积作用(Canfield, 2005; Holland, 2006, 2009)。无论如何, 分子氧的实质性的量, 可能局部聚集在局限浅水环境中, 因为这样的环境有利于蓝细菌的生产作用(Kasting and Brown, 1998), 并且有效地孤立起来以至于氧气没有完全被马上消除。随着大气圈较低的氧气交换速率, 在这些“ 氧气绿洲(Oxygen oases; Fischer, 1965)” 中, 即使在缺氧的大气圈之下, 氧气含量水平可能达到0.016 atm(0.08倍的现代大气圈含量水平(PAL))(Kasting and Brown, 1998; Pavlov and Kasting, 2002; Olson et al., 2013)。更进一步讲, 早于2.7 Ga的叠层石和来源于明显缺乏还原能力的蒸发湖泊沉积物中的生命标志物, 强烈地说明了产生氧气的蓝细菌已经得到进化; 甚至在3.2 Ga前, 厚度较大而且分布广泛的干酪根殖的页岩与需氧的光合自养海相浮游生物相吻合, 以及来自于3.8 Ga的金属沉积物中的U-Pb测年, 也说明这个新陈代谢途径在地质记录开始后不久就已经产生(Buick, 2008)。因此, 在地球大气圈完全彻底被氧化之前含氧光合作用就已经进化的假说得到了多方面的支持。尽管这些认识和发现没有为光合作用生命的起源提供最后答案, 但是, 立足于多学科研究得出的这些富有智慧的认识, 为今后的深入研究奠定了坚实的基础。
光合作用装置的主要单元是光合作用反应中心、触角复合体、电子授体的复合体和固碳作用装置; 对于所有的生物来说, 光合作用装置不具有相同的进化历史, 以至于光合作用装置被看作是一个镶嵌体, 这个镶嵌体由很多具有自己独特的进化历史的次级结构所组成(Olson and Blankenship, 2004; Olson et al., 2013)。就像上文所涉及到的光合作用生命的起源一样, 光合作用装置的起源也是一个极为复杂的科学命题, 下文将简要介绍这些方面的中研究进展。
生物学家的研究已经揭示出, 光合作用仅仅发生在具有某种叶绿素(生氧光合生物)或细菌叶绿素(不生氧光合生物)的生物中, 也就是说, 叶绿素和细菌叶绿素是光合作用进行的重要媒介, 从而成为光合作用的中心色素(Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Nisbet, 1995; Madigan et al., 2006; Olson et al., 2013)。在原核生物中, 没有叶绿体, 以叶绿素为主的光合作用色素结合在内膜系统之中, 即光合作用膜内, 而且光合作用膜就是原核光合作用生物(如蓝细菌)进行光合作用的场所; 在真核光合生物(如高级藻类和植物)中, 光合作用与特殊的细胞器(叶绿体)存在着成因关联, 叶绿素附着在叶绿体的片层膜结构上。在光合作用膜内, 叶绿素和细菌叶绿素与蛋白质结合形成50~300个分子的特殊复合体, 在其中只有少数成为光合作用的反应中心, 在反应中心中的色素分子直接参与光能向ATP的转化, 从而为生物的细胞活动获取能量。所以说, 光合作用装置的起源首先涉及到的就是叶绿素的起源。
复杂的有机光合作用剂(Photosensitizers)的进化, 例如叶绿素, 借助于Granick在1957年提出的假说而合理化, 即生物合成途径囊括了它们的起源和进化(Mauzerall, 1992)。叶绿素是光合作用生物的绿色物质, 由一种卟啉化合物所组成, 是典型的光合作用剂, 通常存在于分离体中, 并且仅仅在有光线和活细胞含有铁的时候才能存在, 常常包括叶绿素a和叶绿素b的混合物, 还含有不同量的其他色素(如胡萝卜素和叶黄素); 叶绿素a是高等植物、大部分藻类和蓝细菌的主要叶绿素。在原核生物中, 生氧光合生物的蓝细菌含有叶绿素a, 不生氧光合作用生物(如紫色细菌和绿色细菌)可以含有任意种类的细菌叶绿素, 细菌叶绿素是指不生氧光合作用的光养生物中的叶绿素。Granick假说回答了叶绿素的生物合成途径是如何通过随机突变而进化的问题; 这个途径是随着每一个步骤实现一个有用的功能而迅速建立的, 最终被下一个步骤得到改进的用途而被替代。
沿着Granick假说所提供的思考途径, Olson 和 Pierson(1987a, 1987b)建立了一个假想, 即来自于叶绿素a 和细菌叶绿素 a的生物合成途径在反应中心(光合作用复合物)中的起源和进化历史。他们提出, 原始的叶绿素在叶绿素 a存在之前的一些时代, 在一个原始的反应中心中可能已经功能化; 而且细菌叶绿素 a的生物合成途径表明, 在细菌叶绿素 a存在之前, 叶绿素 a就已经出现在进化历史之中。Xiong 等(1998, 2000)、Xiong和Bauer(2002a, 2002b)对几个叶绿素合成基因的分子系统发生学的研究表明, 原细菌世系(细菌叶绿素a)可能要比日心细菌世系(细菌叶绿素 g)或蓝细菌世系(叶绿素 a)更加根深蒂固。
所以说, 相对于叶绿素 a和细菌叶绿素 a在进化历史之中出现的序次来说, Granick假说可能是错误的; 但是, 并非证明了该假说在所有方面均不正确。对于在光合作用进化历程中过渡性的色素如原卟啉IX和原叶绿素的生物合成, Granick假说具有正确的一面。较为肯定的是, 原始卟啉IX合成作用的途径, 可能在与丰富而且有效的电子传播辅助因子的进化压力相响应的过程中起源并进化, 如细胞色素b中的血红素(Olson and Blankenship, 2004)。
光合作用的起源与进化, 主要是光养生物内的光化反应中心(photochemical reaction center; RC)的起源与进化。这个被简称为RC的反应中心, 实质上是一种光合作用复合物, 含有叶绿素(或细菌叶绿素)以及其他的组成, 在这种复合物中发生光合作用电子流最初的电子转移反应(Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Olson and Blankenship, 2004; 徐桂荣等, 2005; Madigan et al., 2006; Olson et al., 2013)。
关于反应中心和光合作用的起源, 存在2个学派的思想(Olson and Blankenship, 2004)。第1个学派设想到, 反应中心与光合作用的起源开始于生命起源之前(Mauzerall, 1992; Hartman, 1998); 然而, 第2个学派认为, 反应中心和光合作用的起源与进化的历程, 要晚于细菌(真菌)的厌氧呼吸作用, 而且认为反应中心起源于细菌中的细胞色素b, 这些细菌包含着DNA、电子授体的蛋白质和ATP(腺苷三磷酸)和酶(Meyer et al., 1996; Nitschke et al., 1998)。第1个学派设想到光合作用的起源和生命的起源是紧密相关的; 第2个学派则把光合作用的起源看成要晚于所有生命的共同祖先出现之后很久, 根据这个观点, 光合作用起源于细菌之中, 而且要晚于与古菌分开的时间。
基于上述2个学派的认识, 科学家们还进一步提出了2个模型(选择损耗模型与融合性模型)来解释反应中心的起源。2个模型均提出, 反应中心随后进化于原生细菌、绿色的(非硫的)丝状细菌、蓝细菌、日心细菌、绿色的硫细菌(Xiong et al., 1998, 2000; Xiong and Bauer, 2002a, 2002b; Olson and Blankenship, 2004)。
在选择损耗模型(Selective loss model)之中, 将反应中心的起源和进化与叶绿素合成基因(Xiong et al., 2000)和细胞色素b的起源与进化(Schü tz et al., 2000)综合起来考虑。叶绿素合成基因进化的分叉指令(branching order)是:原细菌→ 绿细菌→ 日心细菌+蓝细菌。另一方面, Gupta(2003)根据非光合作用蛋白质对光合作用生物的分叉指令, 又提出了一个不同的变化序列:日心细菌→ 绿色丝状细菌→ 蓝细菌→ 绿色硫细菌→ 原细菌。这2个分叉指令是否能够调和在一起还有待研究。总的来讲, 该模型认为, 所有的光合作用体系的最近的普遍祖先(Olson and Pierson, 1987a, 1987b), 均被认为包括了RC1和RC2。而且进一步认为, 在原细菌和绿色的丝状细菌中的反应中心的起源与进化形成于RC1的损耗, 然而反应中心在日心细菌和绿色硫细菌中的进化则形成于RC2的损耗; 但是, RC1和RC2均会保留在蓝细菌之中。
许多学者如Mathis(1990)、Blankenship(1992)、Meyer 等(1996)、Xiong 等(1998)以及 Xiong 和 Bauer(2002a, 2002b), 在一个基本方面均不同意Olson 和 Pierson(1987a, 1987b)的认识, 即“ 所有的光合作用体系的最近的普遍祖先, 均被认为包括了RC1和RC2” 。他们认为RC1和RC2均独立进化于不同的生物, 然后通过基因的融合作用组合在一种生物之中, 从而形成具有2个光合体系序列的光合作用的蓝细菌世系, 从而提出了融合性模型(fusion model)。融合作用假说的最近的版本, 已经被Xiong 和 Bauer(2002a, 2002b)总结出。在它们的框架中, RC2(紫色细菌/原细菌)从一个细胞色素b-状的蛋白质中进化而来。这个RC2然后沿着3个不同的途径进化:第1个途径, RC2(紫色细菌)进化成RC2(绿色丝状细菌); 第2个途径, RC2(紫色细菌)发育成RC2(蓝细菌); 第3个途径, RC1(日心细菌则经历了一个RC2(紫色细菌)和祖先式的CP47/CP43蛋白质的基因融合作用。从RC1(日心细菌)到RC1(绿色硫细菌)和另外一个RC1(蓝细菌), 存在一个进化途径。进化的结果是:蓝细菌从“ 日心细菌” 中获得RC1, 而且从“ 原生细菌” 中获得RC2; 但是, 驱动着不同进化途径的进化压力还没有特征化。所以说, 在融合性模型之中, 最近的而且最普遍的祖先被认为来自于2个线索, 一个包括了RC1, 另一个包括了RC2。RC1线索导致了日心细菌和绿色硫细菌的反应中心, 而且RC2线索则产生了原细菌和绿色丝状细菌的反应中心; 该模型进一步认为, 蓝细菌的反应中心是包含RC1的生物与包含RC2的生物的基因融合作用结果。
但是, 2个针对光化反应中心(RC)的模型或假说, 尽管存在着一些分歧, 但是, 均得出一个重要结论, 即RC蛋白质的进化历史不同于色素合成酶的进化历史; 这个结论表明, 很多光合作用装置的构成起源于不同的途径。这个情况已经反映在光合作用原核生物的基因组之中, 在其中大规模的横向基因转化事件, 可能已经成型了这些生物的进化历史(Raymond et al., 2002, 2003; Olson and Blankenship, 2004)。各种光合作用触角(或天线), 即光获取复合体(如LH1和LH2触角复合体、绿色体、叶绿体、Fenna-Matthews-Olson(FMO)蛋白质、藻胆体、CP43与 CP47核心触角等等), 它们的起源和进化历史似乎完全是独立的; 也就是说, 光合作用装置的不同部分的起源, 遵循着多样的途径, 而且在2种不同类型的生物之间、或者甚至在相同生物的不同部分之间, 均存在着不同的起源与进化途径; 因此, 不存在一个可以遵循的单线小路, 去追逐光合作用装置复杂的起源与进化历史(Olson and Blankenship, 2004)。在光合作用装置复杂的起源与进化历史进程中, 所发生的一个重大变化是从不生氧光合作用到生氧光合作用的转变, 这个重大的转变可能发生在蓝细菌学会了如何使用水作为电子授体而进行二氧化碳还原并固定碳的时候; 在此之前, 过氧化氢可能作为过渡性的电子授体, 而且铁离子可能是还原动力的源泉(Olson and Blankenship, 2004)。
不生氧光合作用是最古老的光合作用形式, 运用还原性的硫化物作为固定二氧化碳的电子授体(Olson and Pierson, 1986); 随着环境逐渐变得更加氧化, 运用即使很微弱的电子授体的进化压力将选择第2种光合作用体系(PSII), 这要比只是被第1种光合体系(PSI)能够使用的电子授体要有效得多。但是, 当条带状含铁建造的沉积作用在大约1.85 Ga结束之后(Slack and Cannon, 2009), 向海洋供应的二价铁离子可能是亏损的, 以至于在一定程度上首先是双氧水(H2O2)、然后是水逐渐替代二价铁离子, 从而成为光合作用的电子授体(Blankenship and Hartman, 1998); 一些学者(McKay and Hartman, 1991; Bader, 1994; Samuilov, 1997)甚至强调, 双氧水是早期第2种光合作用体系(PSII)的电子授体。所以说, 从不生氧光合作用向生养光合作用的转变, 还是一个富有争议的主题; 但是, 生物通过光合作用产生氧气的能力, 毫无疑问发生在2.4 Ga左右的古元古代大气圈巨型氧化作用事件(Canfield, 2005)之前(Olson and Blankenship, 2004), 甚至还可能早到3.5 Ga之前(Buick, 2008; Bosak et al., 2009; Riding et al., 2014)。
叶绿体的起源, 很早就被Schimper 和 Meyer在1880年提出, 而且在上个世纪早期被Mereschkowsky进一步发展(Blankenship, 2007); 但是, 经过Margulis(1970)的努力, 在1970年前后这个理论才被总体接受。叶绿体内共生起源的强烈证据, 来自于叶绿体和蓝细菌染色体组的比较。在几乎所有的基因测序能够进行的情况下, 在系统进化树之中的叶绿体基因束, 具有来源于蓝细菌的同源基因。将真核光合作用微生物(和叶绿体)置于2.9 Ga左右(Han and Runnegar, 1992; Knoll, 1992, 2008)是较为合适的。是否所有种类的叶绿体均来自于一个单一的原始内共生事件, 或者相反, 产生于多重事件之中, 一直是长期讨论与争论的主题。大多数合理的证据表明, 一个单一的原始内共生事件, 足以解释叶绿体的分布; 但是, 关于这一点, 证据还是断断续续的, 因为几乎不可能将一个单一的原始内共生事件(endosymbiosis)、与几个独立于空间的事件区分开来。原始内共生事件不同于次生的内共生作用, 在内共生作用之中, 真核藻类可以融合成另外的细胞, 而且这样的进化演变已经发生在真核藻类的不同类型的多重内共生作用之中(Palmer and Delwiche, 1996)。来自于蓝细菌的叶绿体的进化, 已经导致了许多重要的生物结构的变化, 但是基本的需氧光合作用的化学特征则没有改变。
光合作用的副产品是氧气, 光合作用最重要的机能是固定二氧化碳并生成细胞物质而支撑光养生物的自养生长, 生物学家已经揭示出几个重要的光合作用生物化学机制, 其中, 获得广泛认可、并以其发现者Melvin Calvin命名的光合作用生物化学机制就是著名的卡尔文循环(Calvin cycle; Weise and Koegh, 1977; Greenwood and Edward, 1979; Brock et al., 1994; Nisbet, 1995; 徐桂荣等, 2005; Madigan et al., 2006; Nisbet et al., 2007)。卡尔文循环是一个消耗能量的过程, 通过该过程将二氧化碳转化成糖。多数光养生物通过卡尔文循环来固定二氧化碳, 在这个循环过程之中, 二磷酸核酮糖羧化酶(简称为:rubisco)是关键的催化剂, 还被誉称为“ 著名的大气圈麦克斯韦守护神(the Maxwell' s demon of the atmosphere)” (Nisbet et al., 2007)。Rubisco的特异性(对二氧化碳的偏爱胜过氧气), 会促使生命从环境中的二氧化碳之中获取碳。Rubisco是大气圈的主要建造者, 它设计了生命的房屋, 在泥中和在一个上部的含氧居住地中, 具有一个较低的缺氧基础, 而且决定了表面温度, 通过它的碳温室效应气体的管理来完成细胞物质的产生; 一旦基本的设计被构建而且温度被控制, 无机化学定律就会在生物所设置的框架内活动。Rubisco通过与其他类型酶的共同作用, 尤其是通过氮气的产生以及对氮气的控制而设置了大气圈的压力, 这些复杂的作用机制均紧密地与光合作用相联系(Joshi and Tabita, 1996)。共存在4种形式的Rubisco(Hanson and Tabita, 2001; Ashida et al., 2003, 2005; Nisbet et al., 2007)。
Rubisco的形式Ⅰ , 是今天空气中的碳与有机质中的碳的对话者和交换者, 它常常被蓝细菌所利用。一些蓝细菌使用形式IA 的Rubisco, 另外一些蓝细菌则使用形式IB, 就像绿藻和高等植物那样; 一些原生细菌则使用形式Ⅱ C, 其他的藻类则使用形式Ⅰ D(Badger and Price, 2003; Badger et al., 2005)。
形式Ⅱ 的Rubisco出现在很多原生细菌(Proteobacteria)中, 包括嗜氧的和厌氧的原生细菌, 尽管一些嗜氧和厌氧细菌在光合氧气均存在的情况下生长得很快, 但是却不具有Rubisco(Swingley et al., 2007); 形式Ⅱ 的Rubisco, 还发生在鞭毛虫之中(Morse et al., 1995; Morden and Sherwood, 2002)。
产甲烷的古菌含有形式Ⅲ 的Rubisco, 它们是厌氧性生物; 应该注意到的是, 形式Ⅲ 的Rubisco, 似乎没有在产甲烷生物中起到主要作用。
蛋白质状的 Rubisco形式Ⅳ (RLP), 在很多厌氧生物中发现, 包括杆状菌和绿色硫细菌。RLP不是严格的Rubisco, 因为它们不具有碳酸酵素和加氧酶的功能。
Tcherkez 等(2006)的研究表明, 在现代需氧光合作用生物中, 形式Ⅰ 的 Rubisco的特异性几乎接近于完美(Gutteridge and Pierce, 2006), 而且这个优化性可能发生在古老的地质时代。然而, 酶的微细调整和转变, 在漫长的地质历史长河中肯定得到了改善, 甚至在数百万年内(地质历史的瞬间); 蓝细菌已经存在了数十亿年, 自然选择压力表明, 进化优化作用可能发生得很早。因此, 形式I 的Rubisco的优化, 可能发生在很早的时间, 尽管还没有获得可靠的证据, 但是, 光合作用催化剂的起源和进化的复杂历史, 已经在以下几个方面获得了许多富有智慧的认识。
首先, 生氧光合作用可能起源在2.9 Ga左右。使用形式Ⅱ 的 Rubisco的不生氧光合作用微生物(如Rhodospirillum rubrum)捕获光的机制, 可能在形式Ⅰ 的Rubisco的生氧光合作用之前, 就已经起源并进化(Nisbet et al., 1995)。因此, 如果形式Ⅰ 的Rubisco证据出现在太古宙记录之中, 那么, 不生氧光合作用可以追索到太古宙早期。进一步讲, 自从不生氧光合作用之后, 特殊的生氧光合作用(形式Ⅰ 的Rubisco所催化的光合作用)形成在2.9 Ga左右, 这得到了地球上最古老的冰川作用支持(Nisbet et al., 1995; Young, 2002, 2004; Williams, 2008); 较高的生产率和二氧化碳的降低, 允许了海相碳酸盐生物礁的发育, 也证实了这一点(Nisbet et al., 1995, 2007; Altermann et al., 2006; Altermann, 2008; Kazmierczak et al., 2009; Riding et al., 2014)。Nisbet 等(2007)还进一步证实到, 从那个时候起, 就已经意味着羧化酶(carboxylase)和氧合酶(oxygenase)作用之间的平衡, 形式Ⅰ 的Rubisco的特异性(它的所谓的“ 无效性” )已经控制了大气圈二氧化碳和温室效应, 而且支撑着空气中的自由氧与二氧化碳的不成比例。
第2方面, 形式Ⅰ 的 Rubisco的起源和进化历史, 必须与生氧光合作用紧密联系。在地球早期的缺氧历史进程中, 形式Ⅲ 的 Rubisco和RLP可能已经多样化, 而且可能生根在化能无机营养生物(chemolithotroph)中。大气圈氧气的地质记录是富有争议的, 尽管局部富集氧气的条件, 可能存在于3.7 Ga前(Rosing and Frei, 2004; Riding et al., 2014), 而且损耗在空间中的氢气曾经是氧化作用动力的来源(Catling et al., 2001), 但是, 独立于质量的硫同位素分馏作用(MIF)所获得的强有力的证据表明, 早期太古代的大气圈是缺氧的(Farquhar et al., 2010)。这些研究所取得的认识, 也进一步说明了生氧光合作用发生在2.9 Ga之前。
上述2个方面的认识, 也得到了Blank 和Sá Nchez-Baracaldo(2010)针对蓝细菌形态学和生态学革新的时间进程研究的支持。他们的研究, 阐释了以下几个问题:(1)不太严格的生物钟分析表明, 早期的蓝细菌世系一直到2.4 Ga均被限制在淡水体系之中, 这发生在进入海岸微咸水和海水环境之前; (2)随着生境空间的增加和营养物可用率的增加, 以及随后的进入深海的有机碳沉积作用, 可能产生了较大规模的进入生物圈的脉冲, 这可能解释了氧气上升为什么在2.4 Ga左右如此快速; (3)快速的大气圈氧化作用可能消除了甲烷驱动的温室效应, 同时造成二氧化碳分压的降低, 导致了“ 雪球地球” 条件的诞生; (4)与较厚的、纹层状微生物席相关的属性(大型的细胞直径、丝状生长、鞘游动性和固氮作用), 一直到2.32 Ga之后, 都没有见到, 这些属性的出现叠加在2.2~2.1 Ga之间的全球碳同位素异常之上, 代表着蓝细菌最为显著的形态学和生态学革新。因此, 复杂的纹层状微生物群落在海水环境中的出现, 可能产生了一个巨大的生物地球化学循环的重新排序, 最终产生了这个蓝细菌最为显著的形态学和生态学革新; 而且这个重大生物事件, 与发生在2.2~1.8 Ga全球性超事件(梅冥相, 2010; Eriksson et al., 2010)存在着某种程度的关联, 这一次全球性超事件包括: (1)2.3~2.4 Ga的低纬度“ 雪球地球” 冰川作用(Kirschvink et al., 2000; Kirschvink and Kopp, 2008); (2)2.32 Ga之前的大气圈快速增氧作用(Bekker et al., 2004); (3)硫同位素的独立于质量的衰减作用(Farquhar et al., 2000, 2010); (4)2.06与2.20 Ga 之间的Lomagundi-Jatuli碳同位素异常事件(Kirschvink et al., 2000; Kirschvink and Kopp, 2008); (5)首次有意义的石油和磷块岩在2.0 Ga左右的世界性堆积作用(Melezhik et al., 2005)。
同时, 太古宙氮循环与生物的固氮作用的研究(Garvin et al., 2009; Godfrey and Falkowski, 2009; Ettwig et al., 2010), 也印证了形式Ⅰ 的 Rubisco的起源和进化历史必须与生氧光合作用紧密联系, 而且运用形式Ⅱ 和Ⅲ 的Rubisco以及RLP的生物已经在生氧光合作用生命起源之前, 可能已经多样化。氮循环对生物圈提供了基本的营养物, 但是, 其现代形式的古老遗迹还不清楚。基于澳大利亚的2.5 Ga的Mount McRae页岩中的富有机质页岩的岩心的研究, 发现氮同位素值从+1.0‰ 变到+7.5‰ , 然后又回到+2.5‰ ; 这些变化很明显记录了一个从高度缺氧的氮循环变到含氧氮循环的偏离, 同时意味着硝化固氮微生物和去硝化微生物, 已经在太古宙晚期之前得到进化, 而且出现在氧气第一次聚集在大气圈之前(Garvin et al., 2009)。基于南非Campbellrand-Malmani台地的受到轻微改变的页岩中萃取出的干酪根的氮同位素记录, 在古太古宙与2670 Ma之间, 发现干酪根的δ 15N大致上升了2‰ , Godfrey 和Falkowski(2009)进一步解释到, 这个上升可以作为表面海洋中的脱氮作用以及与固氮作用之间的耦合作用开始的标志, 这要求自由氧气的存在。已经知道3种产生氧气的生物学途径:光合作用、氯酸盐的呼吸作用和还原性氧种类的去毒作用; 因为氧化氮总是出现在早期地球上, 在开启生氧光合作用起源和进化之前, 氧气可能用来进行微生物新陈代谢作用, 在早期地球的缺氧大气圈下, 脱氮作用是通过亚硝酸盐的氧气生产作用而进行的, 这个过程产生的氧气随后就被运用于呼吸作用, 这就是第4种途径:“ 通过生氧细菌产生的亚硝酸盐驱动的厌氧甲烷氧化作用” (Ettwig et al., 2010)。
早在1927年, 地质学家Macgregor就曾经指出, 空气的历史是前寒武纪地质学最大的谜。空气是生物建造物, 在其主要的气体构成中, 分子氧和二氧化碳是通过生氧光合作用和呼吸作用而循环的。自从生命出现在我们星球上以来, 生氧光合作用是最为重要的生物革新。对生物量和能量的生产, 生氧光合作用(以及呼吸作用)提供了一个很有效的途径和方式, 而且对生命在地球上的多样化起着关键作用(Farquhar et al., 2000, 2010), 包括前寒武纪末期的后生动物起源、骨骼生物的多样化以及若干方面的生态成功(Butterfield, 2011; Erwin et al., 2011), 乃至在早古生代陆生有胚植物在大陆上的繁盛以及陆生植被的起源和进化(Tal and Paola, 2007; Davies and Gibling, 2010a, 2010b; 梅冥相和刘少峰, 2013)。
地球大气圈分子氧的较高浓度, 被认为是最壮观而且在地质学范畴中是重要的生命信号。早期的地球大气圈是贫乏氧气的, 在生物圈的氧化作用和生物地球化学循环中, 蓝细菌起着一个基本而且重要的作用(Blank and Sá Nchez-Baracaldo, 2010), 氧气在大气圈中的聚集作用大致开始于3.0~2.5 Ga前生氧光合作用起源与进化之后(Payne et al., 2011), 2.9 Ga地球上最古老的冰川作用(Nisbet et al., 1995; Young, 2002, 2004; Williams, 2008)、2.2~1.8 Ga全球性超事件(梅冥相, 2010; Eriksson et al., 2010), 正是地球大气圈起源与进化之后的重要表现。在追逐光合作用的起源与进化过程之中, 迄今为止, 许多细节问题还难以确定, 如:(1)光合作用生命起源的时代标定(Farquhar et al., 2000, 2010; Olson and Blankenship, 2004; Blank and Sá Nchez-Baracaldo, 2010; Payne et al., 2011); (2)光合作用装置及其不同部分所遵循的独立于多样化起源的途径(Olson and Blankenship, 2004); (3)光合作用催化剂的谜一样的起源(Nisbet et al., 1995, 2007)等。在探索光合作用起源这一重大的科学命题的过程中, 除了生物学家的艰辛努力之外, 地质学家尤其是沉积学家们的探索与努力, 为今后的研究提供了许多富有智慧的线索和思考途径, 成为我们学习的光辉榜样。
Buick(2008)认为, 大气圈很明显地经历了一个自从2.4 Ga前的“ 巨型氧化作用事件” 以来的氧化过程, 他对来自于较古老沉积岩中的地质和地球化学信息的探索, 得出的一些重要认识, 为今后的深入研究提供了许多可借鉴的成功经验, 如:(1)在2.45 Ga砂岩中的石油流体包裹体, 来源于产生并需要分子氧的生物; (2)稍微较古老的(2.5 Ga)干酪根页岩中的钼和铼的丰度、以及硫同位素的分类体系, 记录了一个大气圈氧气的过渡型脉动; (3)早于2.7 Ga的叠层石, 以及来源于明显缺乏还原能力的蒸发湖泊沉积物中的生命标志物, 强烈地说明了产生氧气的蓝细菌已经得到进化; (4)甚至在3.2 Ga, 厚度较大而且分布广泛的干酪根质的页岩, 与生氧光合自养海相浮游生物存在着关联; (5)来自于3.8 Ga的金属沉积物中的U-Pb测年也说明, 光合作用这个新陈代谢途径在地质记录开始后不久就已经产生。
许多学者对叠层石形成的生物信号, 尤其是光合作用生命起源的追逐与研究(Kazmierczak and Altermann, 2002; Allwood et al., 2006; Altermann et al., 2006; Awramik, 2006; Schopf et al., 2007; Kazmierczak et al., 2009; Schopf, 2011), 对探索光合作用起源与进化, 提供了许多重要的科学信息, 而且拓展了沉积学的研究思路和研究内容。Bosak 等(2009)提出的重要理念(即锥状叠层石是在早到3.5 Ga的水生环境之中光合作用和趋光性微生物存在的坚实标志), 与Petroff 等(2010)基于生物物理学原理来探索锥状叠层石的几何形态及其与生物物理过程之间的成因关系的探索与研究, 具有异曲同工之妙。Riding 等(2014)对太古宙灰岩的研究与思考, 认为它们可能是氧化作用的代表物, 还有可能是蓝细菌产生的氧气对铁的迁移结果; 所提出的太古宙“ 氧气绿洲” 的甄别, 也是一个开阔了沉积学和古地理学研究思路的成功范例。
徐桂荣等(2005)在《生物与环境的协同演化》的重要著作中, 对光合作用和光合作用生命的起源与进化, 进行了高度的回顾与总结; 从殷鸿福等(2004)的《生物地质学》到谢树成等(2011)的《地球生物学》, 不但代表了从古生物学到地球生物学的跨越, 也为沉积学家去探索光合作用起源与进化之类的重大科学命题, 提供了一些重要的思考途径; 冯增昭(2013)主编的沉积学巨著《中国沉积学》(第二版)囊括了沉积学的方方面面, 为沉积学研究提供了许多基本概念和研究思路, 浓缩了许多中国沉积学家的智慧和新成果, 为今后从沉积学去探索光合作用起源与进化之类的重大科学命题, 奠定了坚实的沉积学基础。
光合作用起源这一个重大的科学命题, 科学家们的探索与研究已经持续了100多年, 取得了许多令人瞩目的成功, 但是, 就像Olson 和 Blankenship(2004)的精辟的回顾与总结那样, “ 已经发生的而且更加复杂的非线性历史, 正在接受挑战并引起极大的兴趣去揭开, 这将会占据科学家至少另外一个30年的时间” 。把光合作用的关键催化剂(Rubisco)誉称为“ 麦克斯韦守护神” , 以及从太古宙灰岩的组构和地球化学特征去甄别“ 太古宙氧气绿洲” , 概念和方法的本身, 就意味着光合作用起源这一重大科学命题的复杂性。因此, 文中对这一个大科学命题的追逐, 目的在于激起同行的兴趣而起到抛砖引玉的作用。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|
| 99 |
|
| 100 |
|
| 101 |
|
| 102 |
|
| 103 |
|
| 104 |
|
| 105 |
|
| 106 |
|
| 107 |
|
| 108 |
|
| 109 |
|
| 110 |
|
| 111 |
|
| 112 |
|
| 113 |
|
| 114 |
|
| 115 |
|
| 116 |
|
| 117 |
|
| 118 |
|
| 119 |
|
| 120 |
|
| 121 |
|
| 122 |
|