



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国东部晚白垩世恐龙化石集群埋藏特征及国内外对比
[王能盛1  , 旷红伟
, 旷红伟1 , 柳永清1 , 彭楠1 , 许欢1 , 章朋2 , 汪明伟3 , 王宝红3 , 安伟1 ]
, 旷红伟, 柳永清, 彭楠|
|

通讯作者简介 旷红伟,女,1969年生,教授,主要从事沉积学和油气地质与勘探研究。E-mail: kuanghw@126.com。柳永清,男,1960年生,研究员,主要从事沉积地质学、沉积古地理学和古生态学研究。E-mail: liuyongqing@cags.ac.cn。通讯地址:北京市西城区百万庄大街26号;邮编:100037。
第一作者简介 王能盛,男,1991年生,硕士研究生,沉积学与能源地质学专业。通讯地址:北京市西城区百万庄大街26号;邮编:100037。E-mail: wangnengsheng0103@126.com。
运用埋藏学与沉积地质学的研究方法与手段,对中国东部晚白垩世具有恐龙化石集群埋藏盆地中恐龙骨骼化石的埋藏地层层序与年代、埋藏骨骼化石特征、骨骼化石埋藏类型及其埋藏成因等进行了分析与总结,并将其产出时代、埋藏相、埋藏特征与国外进行了对比研究。研究认为:中国东部与国外相比,虽然化石产出的具体时期有所差异,但从恐龙种群、埋藏相及沉积地层学等方面均具有相似性,反映了晚白垩世全球古环境、古生态及古气候的相似性。( 1)中国晚白垩世恐龙骨骼化石埋藏事件可分为早期与晚期 2个阶段:早期的 Cenomanian期与晚期的 Santonian-Maastrichtian期。而国外晚白垩世恐龙骨骼化石埋藏事件只发生在晚期,在美洲及蒙古国发生在 Campanian晚期,在欧洲则为 Campanian-Maastrichtian期的过渡时期及 Maastrichtian期。( 2)中国埋藏骨骼化石种属以鸟脚类鸭嘴龙科与兽脚类恐龙为主,它们埋藏于紫红色地层中,埋藏环境主要为冲积扇与河流环境;美洲以鸭嘴龙科为主,欧洲与蒙古国的种群各地不相同,都埋藏于紫红色岩石地层中,埋藏环境主要为河流—泛滥平原。骨骼在不同沉积相组合地层中埋藏学特征各异,与古地理、古生态及古环境密切相关。( 3)结合盆地中的化石埋藏学特征与盆地演化特征,总结了恐龙化石埋藏类型主要有原地埋藏与异地埋藏,盆地古地理格局变动、火山活动、古温度升高及古地震等事件的综合作用是可能的埋藏成因,并提出了恐龙化石的埋藏模式。此研究为揭示中国东部乃至全球晚白垩世的古地理、古气候及古生态环境及恐龙灭绝原因的探讨提供了新的材料与证据。
, Kuang Hongwei, Liu Yongqing, Peng NanAbout the corresponding authors Kuang Hongwei,born in 1969,is a professor. She is mainly engaged in sedimentology and petroleum geology and exploration research.E-mail: kuanghw@126.com.Liu Yongqing,born in 1960,is a research professor. He is mainly engaged in sedimentary geology,sedimentary palaeogeography and palaeoecology. E-mail: liuyongqing @cags.ac.cn.
About the first author Wang Nengsheng,born in 1991,is a master degree candidate. He majors in sedimentology and energy geology. E-mail: wangnengsheng0103@126.com.
This paper analyzes the Late Cretaceous dinosaur bones according to stratigraphic successions,dinosaur species,articulation and association,diversity,the quantity and quality of preservation and the arrangement of dinosaur fossils from the eastern China by method of taphonomy and sedimentology, and make a comparison between demestic and abroad from the points of occurrence age, and taphonomic facies and characteristics. The studies suggest that: Although the concrete preservation times are different,the species, taphonomic facies and characteristics are similar,which reflecting the similarity of palaeoenvironment, palaeoecology, and palaeoclimate during the Late Cretaceous in a global scale. (1)The burial process of dinosaur bones in China during the Late Cretaceous can be divided into two stages: the early stage of Cenomanian and the late stage of Santonian to Maastrichtian. Foreign burial events appeared during the late Campanian in America and Mongolia,and the Campanian-Maastrichtian transitional period and Maastrichtian in Europe. (2)In China,bones were buried mainly in purple rocks of alluvial fan facies and fluvial facies,and the dinosaur species are mainly Ornithopod hadrosaurs and theropods. Compared with dinosaur mass extinction abroad, they were mainly preserved in lacustrine and flood plain. Hadrosaurs and ceratopsids were flourishing in America and Mongolia, while dinosaurs in Europe and Mongolia are different. The characteristics are different in different successions,which are closely related with palaeogeography,palaeoenvironment and palaeoclimate. (3)By combination of taphonomic and basin evolution characteristics,it concluded that burial types are autochthonous and allochthonous,and the probable burial comprehensive factors are: the change of palaeogeography,volcanism,high temperature etc.,then a taphonomic model were built. It is significant for further study of bone taphonomy and palaeogeography,palaeoenvironment and palaeoclimate,and proposes many new materials and evidence for the research of palaeogeography,palaeoenvironment and palaeoclimate.
化石埋藏学在20世纪70年代以前被看作是古生态学的一部分, 之后则与古生物学、古生态环境学及沉积学等学科建立了密切关系, 成为了一门独立的学科。化石埋藏学注重化石在埋藏、保存和破坏等过程中的研究, 发现化石从生物圈到岩石圈的变化, 解决古生态环境、古地理环境及地层时代等方面的问题(Lawrenee, 1968; Behrensmeyer and Kidwell, 1985; Kidwell, 1986)。
恐龙化石作为埋藏学研究的重要对象, 在中国分布广泛。据统计在黑龙江、内蒙古、辽宁、吉林、云南、四川、重庆、贵阳、浙江、河南、山西、新疆、甘肃、山东、广东等26个省自治区都相继发现侏罗纪— 白垩纪的恐龙化石(董枝明, 2010); 虽然中国恐龙化石数量丰富, 但是以恐龙骨骼化石为对象的埋藏学研究却明显落后美洲与欧洲(Brinkman et al., 1990; Varricchio and Homer, 1993; Therrien, 2005; Calvo and Kellner, 2006; Therrien et al., 2009; Fiorillo et al., 2010; Godefroit and Motchurova, 2010; Grigorescu, 2010; Horner et al., 2011; Egerton et al., 2013)。美欧科学家对恐龙骨骼化石埋藏学已经有深入和普遍的研究与成果, 通过对骨骼化石埋藏地层层序与年代、埋藏骨骼化石特征(种群、骨骼改造特征与完整性、富集程度、保存数量和质量、骨骼的排列方式与行为方式、骨骼内部结构与地球化学特征等)、骨骼化石埋藏类型及其埋藏成因等进行研究, 恢复同期古生态环境、古地理及古气候(Behrensmeyer and Kidwell, 1985; Eberth, 1990; Anthony, 1991; Behrensmeyer and Hook, 1992; Behrensmeyer and Chapman, 1993; Behrensmeyer et al., 2000)。
中国晚白垩世恐龙骨骼化石主要分布在内蒙古二连浩特、阿拉善左旗地区, 山西天镇县, 河南汝阳县、栾川县等, 浙江天台县、仙居市、金华市、衢州市、丽水市、永康市等, 山东莱阳市、诸城市, 黑龙江嘉荫市。在上述地区开展的恐龙骨骼化石研究主要集中在恐龙分类学、恐龙化石沉积地层及特殊成因的埋藏解释上(康铁笙等, 1996; 庞其清等, 1996; 庞其清和程政武, 2000; 张晓红等, 2001; 吕君昌等, 2006; 赵海滨等, 2006; 赵喜进等, 2007; 何碧竹等, 2011; 姜杨等, 2011)。例如有关研究认为, 山东胶莱盆地早白垩世出现的强地震事件(殷秀兰和杨天南, 2005; 田洪水等, 2007; 何碧竹等, 2012)是当时古地理环境改变和恐龙生态环境激烈动荡的重要因素; 晚白垩世强地震事件依然频繁, 使古地表高位的风化产物松动, 加剧剥蚀, 并产生快速的洪流与泥石流等负荷地质作用(何碧竹等, 2011)导致恐龙化石的快速埋藏。因此古地震事件作为一种特殊成因解释了恐龙化石的快速埋藏。近几年有学者开始对恐龙骨骼化石的埋藏特征包括赋存特征、埋藏层年代、埋藏相及骨骼内部特征和地球化学特征作了研究(柳永清等, 2010, 2011; 旷红伟等, 2013a, 2013b, 2014), 对古气候、古地理及古生态环境的揭示起到了重要作用。
作者以山东胶莱盆地大量的野外实际资料为重点, 运用埋藏学与沉积地质学的研究方法与手段, 根据中国晚白垩世恐龙骨骼化石盆地的岩石地层、年代地层与恐龙骨骼化石生物地层, 对恐龙骨骼化石集群埋藏的主要沉积盆地即内蒙古二连盆地(Currie and Eberth, 1993; 张晓红等, 2001; Xu et al., 2007), 山西天镇大同盆地(庞其清等, 1996, 2000), 河南汝阳盆地(吕君昌等, 2006; Lü et al., 2006, 2007a, 2007b, 2009; Zhang, 2009), 浙江天台盆地(姜杨等, 2011; 俞方明等, 2012), 山东胶莱盆地(杨钟健, 1958; 胡承志等, 2001; Butler and Zhao, 2009; 柳永清等, 2010, 2011; 季燕南, 2010; Xu et al., 2010; Hone et al., 2011; 旷红伟等, 2013a, 2013b, 2014), 黑龙江嘉荫盆地(李萍, 2004, 2005; 赵海滨等, 2006)进行对比研究, 综述了恐龙化石埋藏学特征与沉积地层学特征, 揭示晚白垩世古地理、古生态与古环境, 以期较全面地认识动物群与古地理、古生态环境之间的响应。
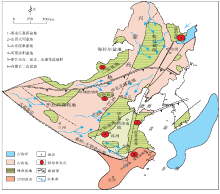
晚白垩世早期(K21)(Cenomanian-Santonian)东亚大陆边界受到太平洋板块的作用, 古地理面貌由早白垩世早期的“ 东高西低” 转变为“ 西高东低” , 沉积盆地主要分布在大兴安岭— 太行山— 华鉴山一线以东地区, 包括松辽盆地、胶莱盆地、苏北盆地、渤海湾盆地等裂谷盆地系, 该期盆地沉积基本都以冲积扇相— 河流相为主, 同时发育冲积平原、小范围浅湖沉积及干旱气候条件下的石膏和沙漠沉积; 在晚白垩世晚期(K22)(Campanian-Maastrichtian)古地理面貌的主要特点是全国广泛分布小型凹陷盆地, 持续的太平洋板块俯冲影响导致中国东南赣州— 杭州断裂带以东全面隆升成山, 出现冲积相— 河流相— 滨浅湖相沉积, 干旱— 半干旱气候条件下的石膏与钙质结核普遍发育(王成善等, 2009; 曹珂, 2009, 2010)。晚白垩世, 中国恐龙骨骼化石集群埋藏在中东部地区受多期次构造活动改造的中小型盆地中, 如:内蒙古二连盆地, 河南汝阳盆地, 浙江天台盆地, 山西天镇大同盆地, 鲁东胶莱盆地和黑龙江嘉荫盆地等(图 1)。
 | 图1 中国东部晚白垩世古地理概图及主要恐龙骨骼化石埋藏盆地(据王鸿祯等, 1985, 有修改)Fig.1 Sketch map of palaeogeographic and main burial basins of dinosaur fossils during the Late Cretaceous in eastern China (modified from Wang et al., 1985) |
晚白垩世早期(K21)(Cenomanian-Santonian)恐龙骨骼化石埋藏地有内蒙古二连盆地、河南汝阳盆地与浙江天台盆地。
1)内蒙古二连盆地属于断陷盆地, 下白垩统从下往上依次发育阿尔善组、腾格尔组和赛汉组, 以温湿气候的湖泊相— 河流相为主, 赛汉组顶部为冲积扇相— 河流相, 气候转为干旱。上白垩统二连组, 底部为河流相河床滞留砂砾岩沉积, 下部为河流相褐色砂岩, 上部则变为湖相灰色泥岩, 恐龙骨骼化石埋藏于二连组最底部河床滞留沉积砂砾岩中, 已鉴别出的恐龙种属有鸭嘴龙、似鸟龙、暴龙、蛇颈龙、翼龙、驰龙科、伤齿龙等(沈鸿章, 1988; Currie and Eberth, 1993; 梁官忠等, 2013), 时代为Cenomanian。早白垩世末期到晚白垩世初期气候干旱, 处于盆地冲积充填阶段, 晚期盆地进一步断陷接受沉积。
2)河南发育众多中生代断陷盆地, 汝阳盆地属其中之一, 下白垩统从下往上为九店组、陈宅沟组和蟒川组下段, 为滨浅湖相— 曲流相— 辫状河相, 九店组沉火山岩锆石SHRIMP U-Pb年龄为133~130 Ma(江小均等, 2010)。上白垩统主要为蟒川组中段, 其岩性以粉砂质泥岩为主, 化石主要埋藏在紫红色泥质粉砂岩和砂质泥岩中, 为河流相沉积, 时代为Cenomanian。汝阳盆地沉积岩性表现为由粗到细的变化, 在晚白垩世盆地西部主要为冲积粗碎屑沉积, 东部小范围为河流及湖泊细碎屑沉积, 此时盆地逐渐消亡(徐莉等, 2012)。潭头盆地发育上白垩统秋扒组, 岩性为棕色厚层泥岩— 粉砂岩, 为湖泊相沉积; 秋扒组粉砂质泥岩中埋藏巨型蜥脚类恐龙与驰龙科栾川龙化石(江小均等, 2010), 时代为Cenomanian。
3)浙江地区在中生代晚期受太平洋构造带板块运动作用影响, 火山活动频繁, 在浙东一带形成一系列大小不一的受多种构造线控制的断陷盆地, 天台盆地属其中之一。天台盆地下白垩统塘上组为灰紫色流纹质角砾凝灰岩, 上白垩统赖家组紫红色中薄层状粉砂质泥岩、泥质粉砂岩, 偶尔也夹带火山间歇喷发的玻屑凝灰质沉积, 湖相紫红色薄层状粉砂质泥岩为主化石埋藏层, 其中夹层晶屑凝灰岩的锆石U-Pb年龄分别为114.3 Ma和94 Ma(俞方明等, 2012)。上部赤城山组以紫红色中薄层状粉砂岩、砂砾岩为主, 剖面上部砂砾岩的砾石成分以火山岩为主, 顶部砂砾岩中含少量沉积岩砾石, 锆石U-Pb年龄值为94 Ma, 在泛滥平原相紫红色泥质粉砂岩中也埋藏化石。从年龄上看2个化石埋藏层都为Cenomanian。目前已发现恐龙化石有鸭嘴龙科、慢龙科与兽足类等(蔡正全和俞云文, 2001; 姜杨等, 2011)。天台盆地晚白垩世伴随火山活动的碎屑沉积, 盆地晚期是以冲洪积粗碎屑沉积为主, 局部为湖相沉积。而盆地群中的丽水盆地、永康盆地、金衢盆地及仙居盆地中, 上白垩统中埋藏骨骼类型及数量较少, 埋藏地层基本为湖相及泛滥平原相紫红色、灰紫色细碎屑沉积物, 时代上应归属为晚白垩世Santonian(汪庆华和斯小君, 2000; 罗来等, 2010; 姜杨等, 2011; 吴因业等, 2015)。
中国北方晚白垩世晚期(K22)(Campanian-Maastrichtian)骨骼化石埋藏盆地有山西天镇大同盆地与山东胶莱盆地及黑龙江嘉荫盆地。
1)山西天镇大同盆地上白垩统灰泉堡组埋藏有恐龙骨骼化石。上段湖相暗紫色含钙质结核的粉砂质泥岩中主要埋藏有蜥脚类和甲龙类, 并含有兽脚类和鸭嘴龙类, 化石丰富且保存完整; 下段底部为冲积扇相灰白色砂质砾岩, 保存较少骨骼, 所处时代大体在Campanian期(庞其清等, 1996; 庞其清和程政武, 2000)。晚白垩世该盆地大部分处于干旱气候下冲积扇粗碎屑充填阶段。
2)山东胶莱盆地白垩系陆相地层序列为莱阳群— 青山群— 王氏群(130~65 Ma)。下白垩统莱阳群以河流及湖泊相沉积物为主; 青山群为中酸性火山岩、火山碎屑岩间夹沉积岩; 上白垩统王氏群下部以冲积扇、泥石流和辫状河道相的沉积物和岩石组合为主, 中部则为滨浅湖相与河流相沉积物与岩石组合, 上部和顶部为冲积扇泥石流、辫状河和洪泛平原相沉积物和岩石组合, 局部夹玄武岩, 反映了晚白垩世末期古地理、古沉积环境已从早白垩世相对湿润和温热的冲积、湖泊环境转变为燥热、干旱的冲积环境。骨骼化石主要保存在诸城臧家庄、恐龙涧和库沟王氏群中上部, 埋藏的恐龙类型以角龙、暴龙、鸭嘴龙以及甲龙和虚骨龙为主, 赋存于辫状河相黄绿色含砾粗砂岩、粗砂岩和洪泛平原相粉砂岩中。莱阳地区王氏群上部洪泛平原微相中埋藏鸟脚类、甲龙类、肿头龙累、兽脚类等恐龙骨骼化石, 岩性为紫色钙质粉砂岩、细砂岩, 其所夹玄武岩Ar-Ar年龄为76.2~73.5 Ma(柳永清等, 2010, 2011; 旷红伟等, 2013a, 2013b, 2014), 所处时代为Campanian。胶莱盆地在晚白垩世时期伴随火山活动和强地震事件(朱日祥等, 2011; 何碧竹等, 2012), 处于盆地发育晚期, 湖泊急剧萎缩, 以冲积粗碎屑沉积充填为主。
3)黑龙江嘉荫盆地下白垩统分为宁远村组与淘淇河组。宁远村组由陆相火山碎屑岩建造和火山熔岩堆积组成, 岩性以流纹质熔岩和碎屑岩为主; 淘淇河组为砾岩、砂砾岩、砂岩等冲(洪)积相地层。上白垩统为永安村组、太平林场组和渔亮子组:永安村组和太平林场组为湖相沉积体系, 以灰色泥岩、泥质粉砂岩和细粒砂岩为主(李萍, 2004)。渔亮子组主要为冲积扇相沉积体系, 由紫红色砂砾岩、砾岩、含砾砂岩组成。恐龙骨骼化石埋藏于渔亮子组的中下段冲积扇相粗碎屑沉积中, 发现的恐龙属种有鸭嘴龙科、暴龙类、虚骨龙科、似鸟龙、栉龙亚科等, 所处时代为Maastrichtian(李萍, 2005; 杨建国等, 2008)。嘉荫盆地晚白垩世气候温暖干旱, 初期以细碎屑沉积为主, 晚期则为粗碎屑沉积, 盆地逐渐萎缩消亡。
从上述各个埋藏骨骼化石地层层序和沉积物看, 晚白垩世早期埋藏相以河流相— 滨浅湖相为主, 以半还原— 半氧化色岩层为特点, 晚期则以冲积相— 河流相河道亚相及泛滥平原亚相为主, 以氧化色岩层为特点(图 2), 骨骼化石在各层中埋藏种群与数量各异; 从所有盆地骨骼化石埋藏地层的时代来看, 在Turonian与Coniacian 这2期没有发生恐龙化石埋藏事件。上述特征表明, 同期盆地的化石埋藏环境, 反映的古气候与古环境具有相似性, 晚白垩世早期盆地以湿热— 半干旱的古气候为主, 晚期以半干旱— 干旱炎热的古气候为主; 而各盆地古地理与古生态的差异对骨骼化石的埋藏特征有直接的影响。
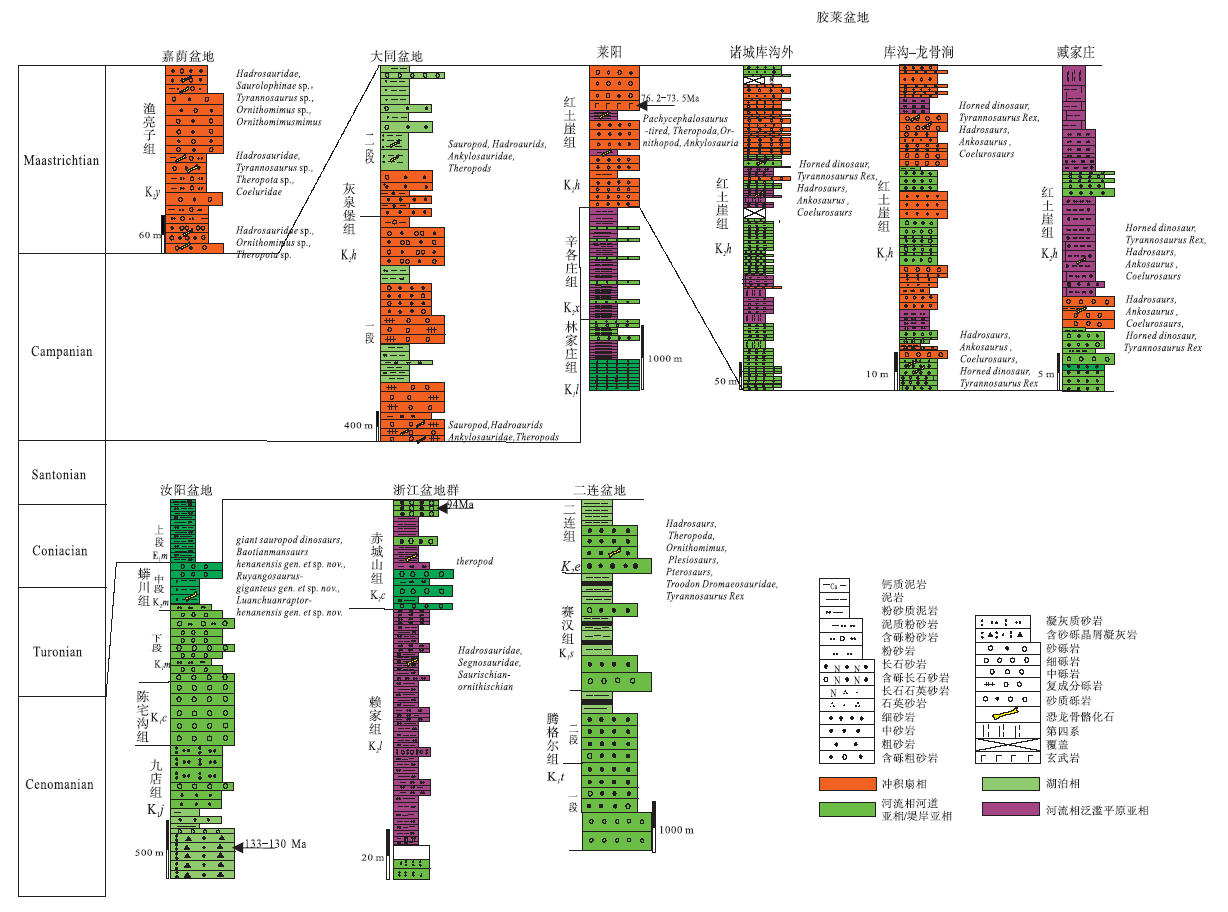 | 图2 中国东部晚白垩世恐龙骨骼化石埋藏地层综合图Fig.2 Comprehensive stratigraphic column of buried dinosaur fossils of the Late Cretaceous in eastern China |
恐龙骨骼化石埋藏特征与盆地古地理、古环境及古生态密切相关, 这些特征包括埋藏相、埋藏种群及生长阶段、骨骼大小、骨骼完整性、富集规模及程度、保存数量和质量、骨骼的排列方式等方面(Speyer and Brett, 1988; Behrensmeyer and Hook 1992; Behrensmeyer and Chapman, 1993; Behrensmeyer et al., 2000)。作者结合盆地沉积、构造演化及生态环境特征, 主要通过埋藏相、埋藏种群、骨骼完整性、保存数量和质量、骨骼排列方式等, 来探讨骨骼埋藏类型及恐龙死亡、埋藏原因。
化石在不同埋藏地层中保存特征不同, 而同一埋藏层内不同的化石种类保存特征也不相同, 根据化石的保存特征可划分出不同的埋藏单元, 不同保存状态的埋藏单元在时空上的分布就是埋藏相。化石的埋藏特征是特定沉积环境下的产物, 据地层中不同的化石埋藏组合, 即不同的埋藏相特征, 分析判断当时的沉积环境(Speyer and Brett, 1988)。
从恐龙骨骼化石在各盆地埋藏岩性及埋藏相(图 3)可以看出, 骨骼化石中埋藏相有冲积扇相、河流相泛滥平原亚相、河道亚相或堤岸亚相、湖泊相等, 通过骨骼化石主埋藏相的总结可以探讨恐龙埋藏特征与原因等。
 | 图3 山东胶莱盆地骨骼化石埋藏相Fig.3 Taphonomic facies of bones in Jiaolai Basin of Shandong Province |
山东胶莱盆地诸城地区与黑龙江嘉荫盆地恐龙骨骼化石主要埋藏相为冲积扇相。胶莱盆地诸城地区王氏群中上部泥石流相中埋藏有高密集度无关联的恐龙化石, 也零星可见极少数的半关联小个体恐龙化石, 而在次埋藏相辫状河流亚相与洪泛平原亚相中仅保存少量的漂浮状骨骼“ 砾石” 或零星骨骼化石(图 3-F, 3-G, 3-H, 3-Ⅰ , 3-J, 3-K); 嘉荫盆地渔亮子组中下段冲积扇相中含丰富骨骼化石, 所产化石关联性较好, 现已组装完成多个完整恐龙个体。
山东胶莱盆地莱阳地区与内蒙古二连盆地恐龙化石骨骼主埋藏相为河流相泛滥平原亚相。山东胶莱盆地莱阳地区王氏群上部洪泛平原微相中, 恐龙骨骼化石较为分散, 完全关联或半关联者少见, 大部分骨骼都表现为孤立状, 见不同程度的磨蚀或破损(图 3-A, 3-B, 3-C, 3-D, 3-E); 内蒙古二连盆地洪泛平原相中骨骼化石种群多, 关联性差, 埋藏位置靠近粗颗粒的河道沉积。
浙江天台盆地与山西天镇大同盆地恐龙化石骨骼主埋藏相为湖泊相。浙江天台盆地主要埋藏在紫红色薄层状粉砂质泥岩, 夹中薄层状泥质粉砂岩透镜体的湖泊相中(图 4-A, 4-B, 4-C); 山西天镇灰泉堡组上段湖泊相与下段洪积扇相中埋藏恐龙骨骼化石, 主埋藏湖泊相中的化石丰富且保存完整, 而在冲积扇中骨骼较少。
 | 图4 浙江天台盆地与河南汝阳盆地骨骼化石埋藏相Fig.4 Taphonomic facies of bones in Tiantai Basin of Zhejiang Province and Ruyang Basin of Henan Province |
河南汝阳盆地骨骼化石埋藏在紫红色泥质粉砂岩和砂质泥岩夹灰绿色砂质泥岩中(图4-D, 4-E), 该埋藏相为曲流河相。
可以看出, 盆地的主埋藏相多为快速搬运并埋藏的冲积扇相与适合恐龙生存、水动力条件相对较弱的湖泊相、河流相泛滥平原亚相, 而河流相河道环境水动力较强, 易搬运骨骼, 故骨骼一般被河水搬运至其他环境后沉积埋藏或在河道单元末端埋藏。主埋藏相的不同只能说明恐龙死亡后骨骼化石沉积及埋藏环境的异同, 而不能代表恐龙当时完整的生存环境及整个生态系统, 只有结合骨骼化石的自身特点及盆地沉积-构造演化等才能还原完整的恐龙生存、死亡再到埋藏的一系列过程。
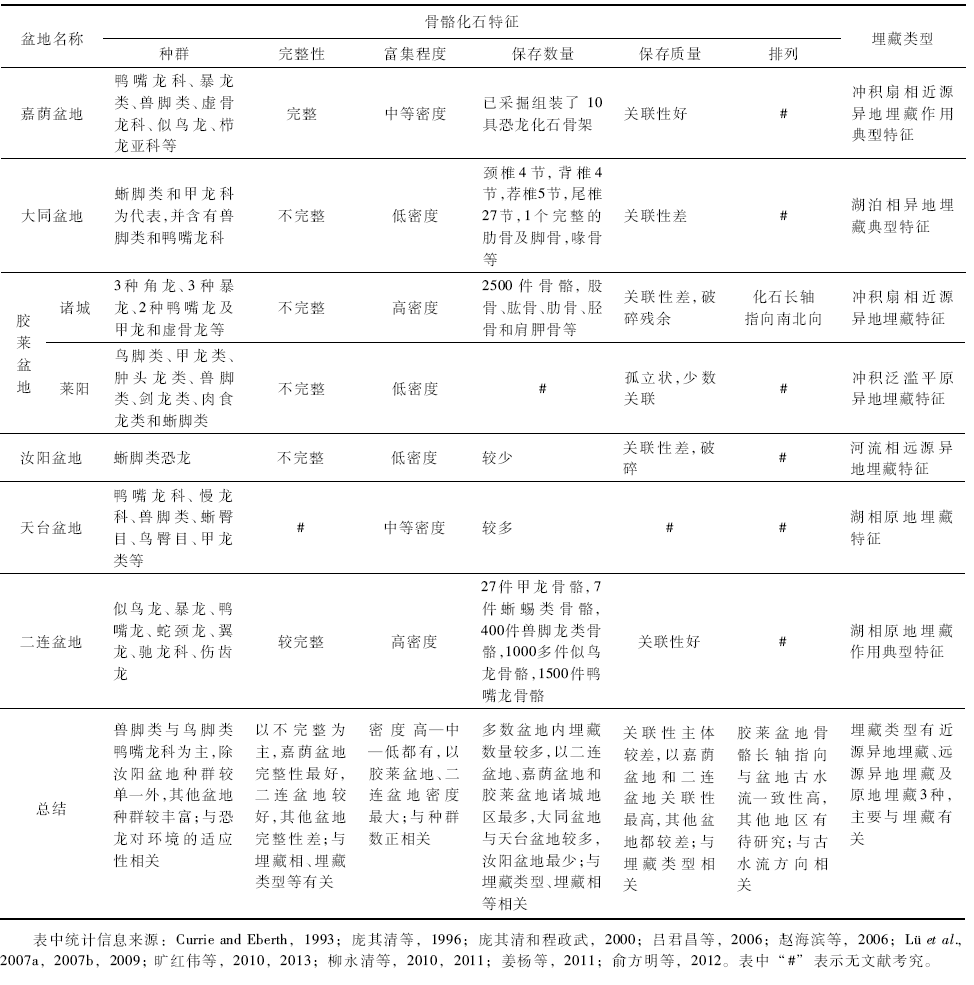
骨骼化石特征研究是化石埋藏学研究的重要组成部分, 是对埋藏种群及生长阶段、骨骼大小、骨骼改造特征与完整性、富集程度及规模、保存数量和质量、骨骼的排列方式等方面进行描述(Voorhies, 1969; Dodson, 1973; MacDonald and Jefferson, 1985; Turnbull and Martill, 1988; Rogers, 1990; Hunt, 1990; Smith and Kitching, 1997; Eberth et al., 2000; Fiorillo et al., 2000; Rogers, 2005), 总结中国晚白垩世恐龙骨骼化石盆地的化石特征(表 1), 由于资料数据的局限性, 主要是统计出埋藏骨骼种群、骨骼完整性与富集程度、保存数量与质量及骨骼排列等宏观特征。
| 表1 各盆地埋藏相中骨骼化石特征总结 Table 1 Summary of taphonomic characteristics of dinosaur fossils in different basins |
根据集群埋藏的骨骼化石在地层中的分布, 可以统计出骨骼数量、可分辨的骨骼化石所属恐龙种群与可确定最少恐龙个体数, 当恐龙种群数为2~5时, 称为低密度埋藏层; 种群数超过10, 则为高密度埋藏层; 种群数为6~9则为中等密度埋藏层(Turnbull and Martill, 1988; Rogers, 1990; Varricchio and Horner, 1993)。富集程度是个相对的概念, 与研究者对埋藏地层、埋藏相或埋藏体系与埋藏过程的认识有关; 通过对晚白垩世盆地骨骼化石在系统发育分支系统上“ 类” 级别上的对比(表 2)可以看出:兽脚类与鸟脚类恐龙在晚白垩各沉积盆地与各阶段都有分布, 其中的鸭嘴龙科骨骼埋藏数量在各盆地中最多, 例如恐龙种群最多的山东胶莱盆地(Campanian), 鸭嘴龙科在95%以上, 内蒙古二连盆地(Cenomanian)埋藏的绝大多数恐龙都是鸭嘴龙科(Currie and Eberth, 1993)。可见, 兽脚类与鸟脚类中的鸭嘴龙科在中国整个晚白垩世盆地中基本都有分布, 而集群埋藏骨骼规模与数量上鸟脚类鸭嘴龙科占有较大比例, 这一点可以说明鸭嘴龙科恐龙对晚白垩世古气候、古环境具有较强的适应能力。集群埋藏层骨骼化石密度可以指示古生态系统及古生物群对形成骨骼埋藏层的影响, 同时进一步了解埋藏盆地的生态系统(Eberth et al., 2000)。
| 表2 中国晚白垩世盆地产出恐龙骨骼化石种类对比 Table 2 Correlation of different kinds of dinosaur bones in basins of the Late Cretaceous in China |
完整恐龙个体在埋藏过程中会受到不同程度的地质作用(机械磨损、化学溶解等), 在埋藏层中骨骼总数量与可确定最少恐龙个体骨骼总数量的比值表示骨骼的完整性(Hunt, 1990; Fiorillo et al., 2000), 完整性判断与研究者对数量统计标准相关, 常有不同的结论。骨骼完整性特征被认为与水流对经过不同地质作用骨骼的分选或选择性搬运过程相关, 所以骨骼完整性可以反映地质作用的差异及对骨骼大小的影响(Dodson, 1973; Smith and Kitching, 1997)。如胶莱盆地冲积扇相埋藏作用, 骨骼受到泥石流的搬运作用, 表现出了不完整特征; 二连盆地湖泊相埋藏作用, 骨骼受到湖水的搬运, 表现出了较完整的特征。
骨骼化石的保存质量是恐龙个体经历埋藏过程后, 各骨骼关联性、骨骼风化程度如破碎、裂缝等特征。骨骼的关联性用生物个体埋藏后分离骨骼数与埋藏前铰接骨骼数的比例来表示, 分为不关联(孤立状)、半关联、关联及其中之二或三者都存在的混杂关联(Smith and Kitching, 1997; Rogers, 2005)。关联性是宏观观察的结果, 通过观察骨骼之间的关节连接或分离, 大致得出关联性好、较好、较差或差, 关联性的不同反映了骨骼化石不同的埋藏历史。生物的个体发育与生物种类在一定程度上也会引起关联性的差异。如嘉荫盆地冲积扇相埋藏作用, 骨骼关联性好, 胶莱盆地泛滥平原环境的埋藏作用, 骨骼呈孤立状、基本不关联。
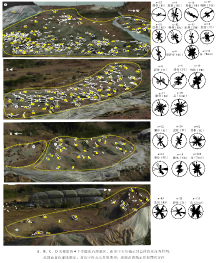
骨骼排列是对埋藏地层中较大骨骼长轴各向排列的统计, 骨骼排列与古水流搬运能力、水流方向、水深及骨骼自身大小、形状等有关(Voorhies, 1969; MacDonald and Jefferson, 1985)。通过对晚白垩世早期(Campanian)山东胶莱盆地诸城地区恐龙骨骼化石的观察, 发现诸城库沟— 恐龙涧地区骨骼化石排列上杂乱无章, 但通过较大骨骼的长轴方向测量、统计与玫瑰图显示了恐龙涧几个主要化石层的骨骼排列方向, 主要为南北向(图 5)。通过叠瓦状砾石方位的测量指示古流向总体向南、少量朝北(旷红伟等, 2013b), 骨骼排列方位与古水流方向的高度一致性表明骨骼与围岩经历了相同的搬运过程, 可以看出:大规模集群埋藏的骨骼排列可以佐证盆地古地理的演化并为深入研究提供科学依据。其他盆地骨骼化石排列则缺少具体的统计和研究, 可以确定的是集群埋藏经过搬运异地埋藏的骨骼排列方向与盆地古水流向具有较高的相关性。
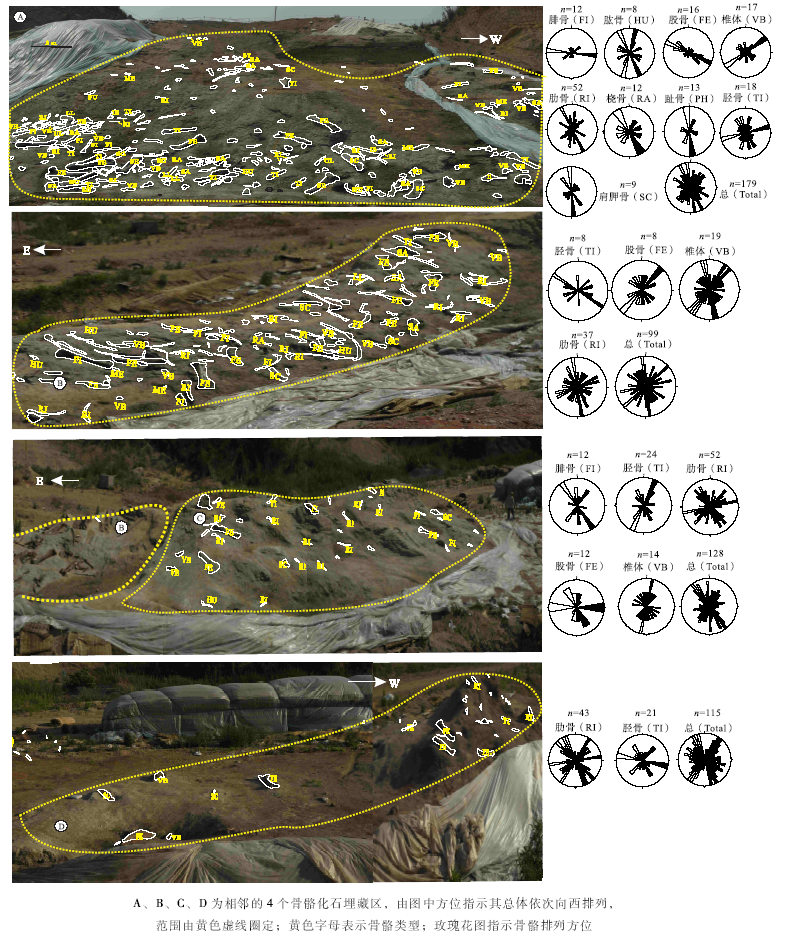 | 图5 山东省恐龙涧恐龙骨骼化石类型及其埋藏排列方向Fig.5 Types of dinosaur bones fossils and their orientation in Konglongjian site in Shandong Province |
骨骼化石特征与盆地古地理、古生态环境有密切的关系, 恐龙种群的相似性说明了中国东部晚白垩世古生物群、古生态的相似性, 骨骼化石完整性、关联度等特征的差异性说明了化石经历了不同的埋藏过程, 经历了不同的地质作用, 也即盆地古地理环境的演化存在差异性。
在生物化石埋藏研究中, 埋藏类型系指生物遗体因不同的原因和不同的方式被掩埋起来, 并在长期的地质作用下得以保留的全过程。埋藏类型分为原地埋藏与异地埋藏, 原地埋藏指生物个体死亡之后、在尚未遭到自然营力的搬运或其他介质的搬运之前, 就被沉积物迅速地掩埋起来而逐渐形成的化石; 异地埋藏指生物的个体或群体死亡后因受到各种自然营力的作用, 搬运到其他的沉积环境中堆积并埋藏, 最终形成的化石(Voorhies, 1969)。笔者结合埋藏相、骨骼宏观特征等判断骨骼化石的埋藏类型及过程。
浙江天台盆地、山西天镇大同盆地有相同的埋藏相, 但其埋藏类型不同。从图2可知, 浙江天台盆地骨骼化石埋藏在湖相的赖家组与泛滥平原相的赤城山组, 而埋藏层靠近下伏河流相含砾砂岩、砂岩沉积, 保存的骨骼完整性较差、零星分布, 表明骨骼化石在河流长距离搬运过程中, 较强的水动力以及砂质的摩擦, 骨骼化石容易遭受风化, 因此能完整地被搬运到湖泊或泛滥平原的骨骼化石大为减少, 多以关联性低、完整性差为特征, 属于异地埋藏类型; 山西天镇大同盆地恐龙骨骼化石丰富, 保存完好, 骨骼排列杂乱, 但关联度高, 产出大型完整的恐龙化石, 属于原地埋藏。
山东胶莱盆地和黑龙江嘉荫盆地骨骼化石有相同的主埋藏相, 埋藏类型也相同。冲积扇泥石流亚相具有沉积物密度大、摩擦作用强、水动力强、搬运能力大、沉积速度快等特点, 其中骨骼化石为快速、近距离埋藏。例如山东胶莱盆地冲积扇沉积中化石关联性与完整性受到了破坏, 基本为无关联的大小不一的骨骼, 高密度堆积, 也见零星、极少数的半关联小个体恐龙化石, 骨骼中裂缝、破损等痕迹较为发育(图 6-A, 6-B, 6-C), 反映骨骼化石在泥石流的搬运下, 砂砾石作用于骨骼的强摩擦力与高黏度的水动力使骨骼相互碰撞更频繁, 使得骨骼受到高风化程度、强破坏作用, 并快速密集埋藏, 与之构成埋藏序列的辫状河道亚相与泛滥平原亚相中存少量的漂浮状骨骼“ 砾石” 或零星骨骼化石, 多数骨骼(99%)所表现的破碎残余状则是由于泥石流更长距离的搬运, 使骨骼数量减小、破碎程度升高(图 6-D)。
 | 图6 山东胶莱盆地诸城市库沟区恐龙骨骼化石冲积扇相异地埋藏类型及特征Fig.6 Taphonomic types and charactersitics of dinosaur bone fossils in Kugou of Zhucheng City, Jiaolai Basin of Shandong Province |
由此可见, 埋藏类型反映的是骨骼化石的搬运过程, 是骨骼特征的直接影响因素, 也反映了盆地的古地理环境。盆地边缘高地冲(洪)积环境能快速或短距离搬运埋藏沉积物, 冲积扇砂砾石对骨骼的强摩擦及干旱气候的暴露风化使骨骼断面具高丰度的裂面等, 若为快速原地埋藏, 骨骼的完整性、关联性及密度等都较高, 如嘉荫盆地恐龙化石的集群埋藏; 若为短距离的异地埋藏, 则具有较高的密度, 但完整性和关联性较差, 如诸城恐龙化石的集群埋藏。盆地中部河流环境具有较强水动力及搬运能力的特点, 河流砂质沉积物对骨骼有较强摩擦, 从而骨骼间碰撞频率高且有分异作用, 一般完整性差、密度低及关联性差, 如汝阳盆地。盆地末端湖泊和泛滥平原环境水动力与搬运能力弱, 同时粉砂质泥质沉积物对骨骼摩擦作用弱, 骨骼碰撞频率低, 若恐龙生存在水分和植被都很充分的湖泊及泛滥平原环境下, 由于某种原因死亡后原地埋藏在该环境中, 其骨骼化石完整性、关联性及密度等都很高, 破碎及破裂程度很低, 主要以抬升暴露风化为主; 若是由于河水的泛滥作用异地搬运埋藏在湖泊与泛滥平原中, 则完整性、关联性及密度等都很低。
到目前为止, 以恐龙为主的中生代古生物灭绝有很多种假说, 并且从各自的角度提出理论依据(戎嘉余和詹仁斌, 2006; 戎嘉余和黄冰, 2014), 例如灾变论假说有天体撞击、气候骤变、食物链中断、有毒物质或放射性物质污染了食物、地球磁场异常、巨大热量使生物化为灰烬等等, 围绕假定的灾变结果发现了很多地质现象, 如在世界范围内发现了铱异常现象, 找到了可能与撞击有关的冲击层和冲击变质矿物等(Alvarez et al., 1980; Hildebrznd, 1990; Sharptom et al., 1992; Jehanno, 1992; Schulte et al., 2010); 相反的观点是用渐变论或地内作用来解释, 如病态论、竞争论、强火山作用等(阎锡海, 1984; Officer and Drake, 1987; Keller, 2012; Keller et al., 2012)。
恐龙生活在地球上的时间跨度很大, 从侏罗纪— 白垩纪出现连续或间断性的集群死亡。作者总结晚白垩世含骨骼化石盆地古地理、古环境、古气候, 即从盆地沉积演化、构造演化、生态环境演化出发, 探讨其死亡埋藏成因。
受区域性地壳伸展作用的影响, 胶莱盆地在早白垩世不断扩大, 盆地内气候湿润, 植被繁茂, 适宜各种动植物生存, 以恐龙为代表的大型脊椎动物群栖息在河湖边缘。早白垩世中晚期由于软流圈上侵, 强烈频繁的火山喷发致使环境恶化, 同时岩石圈减薄和破坏(张岳桥等, 2004; 朱日祥和郑天愉, 2009; 朱日祥等, 2011)导致了强地震事件多发(殷秀兰和杨天南, 2005; 田洪水等, 2007; 何碧竹等, 2011, 2012), 生态环境发生了极大变化。晚白垩世继承了早白垩世晚期的环境和气候特点, 古温度达到了地质历史时期最高峰, 气候炎热、干旱, 水系枯萎, 植被减少, 恐龙等大型脊椎动物开始不断集群死亡, 并为强古地震事件引起的突发洪水、泥石流搬运并埋藏, 最终在白垩纪末期走向了灭绝(柳永清等, 2011; 旷红伟等, 2014)。
根据孢粉分析, 结合沉积物特征, 晚白垩世的嘉荫恐龙群生活在一个气候温暖、湿润, 水分、食物充足的环境中。但晚白垩世晚期(渔亮子组沉积时期), 气候逐渐干旱, 气温升高, 生态环境恶化, 植物骤减, 由气候变化引发的多种因素联合作用使嘉荫恐龙群在晚白垩世末期全部灭绝, 并被泥石流沉积物快速埋藏(赵海滨等, 2006)。
浙江地区白垩纪各盆地总体属于山间湖盆环境, 晚白垩世早期, 盆地气候干旱温热, 水分充足, 生态环境较适宜动植物生存, 恐龙动物群集中分布在湖泊、平原中; 在晚白垩世中期有过间歇性火山活动热事件, 形成干燥炎热的山间湖盆环境; 晚白垩世晚期, 有数次区域性强弱不一的火山活动, 盆地周围发生强烈抬升, 盆内急剧下降, 恐龙的灭绝与此时的山洪暴发、河流的泛滥有关, 大量恐龙死亡后被异地搬运埋藏至赖家组的湖相沉积中(姜杨等, 2011; 俞方明等, 2012)。
山西天镇大同盆地恐龙化石为原地埋藏的, 其主化石层磷灰石裂变径迹年龄(73 Ma)接近锆石裂变径迹年龄(73~78 Ma), 其下部磷灰石裂变径迹年龄(60~70 Ma)较锆石裂变径迹年龄年轻约10 Ma, 而磷灰石裂变径迹的封闭温度为200~250 ℃。这表明在坎潘阶之前源区经历过温度超过200~250 ℃的热作用, 随后发生冷却、搬运和沉积。这些热作用有可能对晚白垩世以来的生态环境和气候产生重要影响(康铁笙等, 1996), 恐龙的死亡埋藏可能与该处的热作用及构造运动有关。
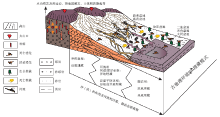
上述盆地演化特征与骨骼化石的埋藏学特征总结, 得出晚白垩世恐龙埋藏模式图。晚白垩世盆地演化导致恐龙出现死亡和埋藏的因素包括:构造火山喷发与构造深部热运动、盆地萎缩高山隆起的盆山效应、频发性古地震及冲洪积作用、水系的干涸、古气温的极度升高、食物供应的急剧减少等, 恐龙无法适应与承受不断恶化的环境, 从而出现了间歇性的死亡与埋藏(图 7)。继而在适宜恐龙生存的湖泊及泛滥平原环境中, 其大部分死亡后是埋藏于原地的, 而死亡后经河流搬运的骨骼异地埋藏在曲流河或者进入湖泊中埋藏, 部分由于河水泛滥被河水搬运至泛滥平原中, 而阵发的冲洪积作用能快速地埋藏正在迁徙的恐龙, 近源堆积在冲洪积扇中。
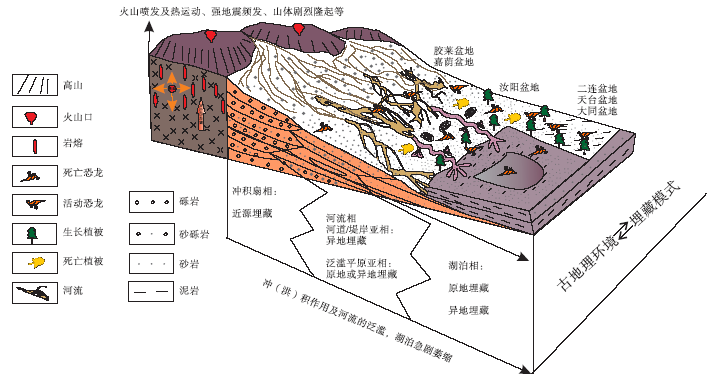 | 图7 中国东部晚白垩世恐龙死亡埋藏模式Fig.7 Taphonomic patterns of the Late Cretaceous dinosaurs in eastern China |
晚白垩世, 中国东部受到太平洋板块俯冲作用的影响, 先期的原型盆地及同期小型断拗型盆地受到持续的改造而最终成为残余型盆地。残余盆地演化历史多变且复杂, 盆地古地理格局变动较大, 盆地边缘高山隆起、内部萎缩效应十分显著, 加之伴随强构造断裂、古地震及火山活动等, 盆地古地理、古气候条件发生较大变化。上述一系列相连贯的事件是中国东部古地理环境改变及动物衰亡与埋藏的诱因, 也是恐龙死亡及埋藏的主导因素。
晚白垩世, 全球范围内包括蒙古、美洲与欧洲地区都出现了恐龙集群死亡与埋藏事件。
1)与中国北方相邻的蒙古戈壁沙漠地区, Campanian时期的Djadochta组、Barun Goyot组与Nemegt组, 都为河湖相红层沉积地层中原地埋藏完整、风化程度低的骨骼化石。恐龙种群有迅猛龙、原角龙、甲龙、似鸟龙、窃蛋龙, 可能是沙暴和酸雨气候条件下的死亡(Saneyoshi et al., 2011; Nied zwiedzki et al., 2012; Shaena et al., 2013)。
2)美国Montana地区 Upper Two Medicine组与Hell Creek 组中, 保存有大规模高密度的恐龙骨骼化石, 赋存地层为湖泊— 河漫滩相钙质泥岩、砂泥岩, 大部分为三角龙, 其次为埃德蒙顿龙、太西龙、似鸟龙和甲龙、霸王龙, 为集群原地埋藏的结果。同样该地区Judith River组中, 冲积平原砂和泥砂中异地埋藏有角龙为主及冠龙、似鸟龙(Varricchio and Horner, 1993; Horner1 et al., 2011); Big Bend area of Texas地区, Aguja 组在冲洪积河道之上的紫红色泥质砂岩中保存大量鸭嘴龙(占到60%)和少量普通角龙及甲龙, 还有霸王龙、奔龙和似鸟龙, 可能是旱灾和洪灾作用异地埋藏作用, 以上地层都为Campanian期(Sankey, 2010); Northern Alaska地区Prince Creek 组中, 富含有机质的粉砂岩和泥岩及细粒砂岩的岩性地层中异地埋藏骨骼化石, 大范围分布在废弃河道、决口扇、天然堤、湖泊和其他低能河漫滩相中, 有3层高密度分布骨层, 兽脚龙的大小骨骼, 肿头龙、角龙、凌齿龙和鸭嘴龙骨骼平行或横向排布, 大部分骨骼无关联, 地层时代为Campanian-Maastrichtian期(Fiorillo et al., 2000)。
3)加拿大Alberta地区Judith River组, 在凹槽斜层理砂岩层的河道中保持较完整的关联较好的鸭嘴龙、角龙、甲龙和暴龙骨骼, 其中50%的鸭嘴龙、27%的角龙、14.6%甲龙和8.3%的暴龙, 在边滩沉积中埋藏有零星骨骼, 同时期Wapiti 组的粉砂岩、细砂岩和泥岩的地层序列即低能量沼泽冲积中保存鸭嘴龙为主, 其他恐龙比较少见, 包括暴龙、蜥鸟盗龙、理查德伊斯特龙、近爪牙龙和奔龙, 地层时代为Campanian期(Wood et al., 1988; Brinkman, 1990)。
4)阿根廷Santa Cruz Province地区在Campanian-Maastrichtian期的冲积河道致密中粒砂岩中埋藏有禽龙的部分铰接骨骼和大部分碎骨; Bajodel Añ elo地区the RioColorado Subgroup的红色砂岩有少量化石(Calvo and Kellner, 2006)。
5)西班牙Province of Cuenca地区在Campanian-Maastrichtian期埋藏的恐龙包括雷龙(96%), 兽脚龙(3%)和1%的似鸟龙, 呈现完整与高关联度形态, 埋藏相为泛滥平原相中红色泥质岩, 为快速原地埋藏(Egerton et al., 2013)。
6)保加利亚Cherven Bryag西南部地区的Kajlâ ka组, 主要发育Maastrichtian晚期滨海灰岩, 埋藏破碎程度很高的似鸟龙骨骼化石(Godefroit and Motchurova, 2010)。
7)罗马尼亚Haţ eg Basin中 Sâ npetru Formation红色泥岩中埋藏有小型兽脚龙、翼龙、类伤齿龙, 为河流— 冲积平原相, 处于Maastrichtian早期(Therrien, 2005; Therrien et al., 2009)。
通过对国外晚白垩世骨骼化石的总结研究表明, 从种群类别上, 中国东部在晚白垩世大范围的恐龙集群埋藏种类主要以鸟脚类鸭嘴龙科与兽脚类为主, 而美洲以鸭嘴龙科为主, 其次是角龙类。欧洲与蒙古国的种群各地不相同, 鸭嘴龙科分布时期与地区跨度大, 表明了该类恐龙发育上的某种环境的适应性。从时代分布上, 中国东部埋藏事件可分为2阶段4期次:晚白垩世早期阶段包括Cenomanian(内蒙二连盆地, 河南汝阳盆地, 浙江天台盆地、金衢盆地)和Santonian(浙江丽水盆地等), 晚白垩世晚期阶段包括Campanian(山西天镇, 山东莱阳和诸城)和Maastrichtian(黑龙江嘉荫地区); 而美洲与欧洲地区恐龙集群埋藏事件则在晚白垩世晚期Campanian与Maastrichtian出现, 分布时期有差异。可见, 中国在晚白垩世恐龙的分布时期上更加广泛, 鸭嘴龙科分布时间跨度最大; 国外分布局限在晚期, 但全球的古气候都以半干旱— 干旱为主, 盆地的沉积岩性以氧化色为特征, 河流相— 泛滥平原相及湖泊相是恐龙化石的主埋藏相, 故从种群、分布时代及埋藏层特征等显示了晚白垩世全球古环境与古生态的相似性。
| 表3 亚洲其他国家与欧洲及美洲晚白垩世恐龙骨骼化石埋藏特征对比 Table 3 Comparision of brief taphonomic characteristics of dinosaur fossils among Asia, Europe and America |
以山东胶莱盆地恐龙骨骼化石为重点研究对象, 运用埋藏学与沉积地质学的研究方法与手段, 对中国晚白垩世主要集群埋藏恐龙骨骼化石盆地进行岩石地层、时代地层与恐龙骨骼化石生物地层的对比, 较全面地研究与探讨了埋藏骨骼化石特征、埋藏类型及埋藏成因。
1)中国晚白垩世恐龙集群埋藏点主要位于内蒙古二连盆地, 山西天镇大同盆地, 河南汝阳盆地, 浙江天台盆地, 山东胶莱盆地, 黑龙江嘉荫盆地等; 数量多、埋藏规模大, 为研究晚白垩世恐龙化石埋藏学提供了大量基础资料和研究成果。
2)中国东部晚白垩世埋藏盆地主要为残余盆地, 埋藏地层多为氧化色, 埋藏相以冲积扇、河流相泛滥平原亚相及湖泊相为主。埋藏地层序列反应的盆地沉积演化过程:温暖湿润湖泊环境向半干旱环境湖泊萎缩, 河流— 三角洲环境发育到干旱环境河流、冲积扇发育。晚白垩世晚期盆地周边隆升强烈, 温度极高, 气候极干旱, 恐龙生存与埋藏环境发生了明显的变化。这些古地理、古气候及古生态环境变化对恐龙的死亡及埋藏学特征产生了直接影响。
3)通过对恐龙骨骼化石埋藏特征(分类学特征、骨骼改造特征与完整性、富集程度、保存数量和质量、骨骼的排列方式等)与盆地演化特征的研究表明, 骨骼化石在埋藏密度与完整性、埋藏数量与质量、排列方式等受到盆地埋藏环境及埋藏过程的直接影响, 主要的埋藏环境包括冲积扇相、河流相及湖泊相, 埋藏过程包括原地埋藏与异地埋藏, 而骨骼埋藏特征反映了不同的埋藏环境与埋藏过程。
4)通过对比蒙古、美国、加拿大、阿根廷、西班牙、保加利亚及罗马尼亚等国晚白垩世恐龙化石埋藏时期、主要种群、埋藏相、骨骼特征及埋藏类型可知: 虽然分布时期有差异, 但恐龙种群、埋藏相及沉积地层学等方面显示了晚白垩世全球为鸭嘴龙科为主的古生物群系统, 古气候条件为半干旱— 干旱, 盆地沉积环境的古地理格局变化剧烈。中国东部乃至全球晚白垩世恐龙化石集群埋藏特征的研究, 对于恢复恐龙化石埋藏成岩过程, 追溯恐龙的死亡原因和埋藏类型, 重建盆地演化的古地理、古生态环境及古气候具有重要意义。
致谢 感谢审稿专家的评审意见和有益建议以及编辑们负责、细致、辛苦的工作。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|