


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
四川盆地宣汉渡口和重庆北碚下三叠统海相碳酸盐碳同位素组成与演化
[黄可可 , 黄思静, 胡作维, 钟怡江, 李小宁]
, 黄思静, 胡作维, 钟怡江, 李小宁]
, 黄思静, 胡作维, 钟怡江, 李小宁]
|
|

第一作者简介: 黄可可,女, 1983年生,博士,成都理工大学沉积地质研究院讲师,从事沉积地质学的教学与科研工作。 E-mail: h-k-k@yeah.net。
通讯作者简介:黄思静,男, 1949年生,硕士,成都理工大学沉积地质研究院教授,博士生导师,从事沉积地质学的教学与科研工作。 E-mail: hsj@cdut.edu.cn。
早三叠世海水的碳同位素组成与演化是二叠纪—三叠纪生物群体灭绝事件之后全球生态萧条与重建时期地球碳循环的重要记录,为地学界长期高度关注。文中测试了四川盆地宣汉渡口和重庆北碚下三叠统及相邻地层 258个海相碳酸盐样品的碳、氧同位素组成,以便探讨早三叠世海水的碳同位素组成与演化。这些样品对海水碳同位素组成具有不同的保存性,飞仙关组及嘉陵江组第一、三段保存较好,而嘉陵江组第二、四段保存较差。基于氧同位素可以更为敏感地反映海相碳酸盐成岩蚀变性的原理,在碳、氧同位素相关性分析的基础上,以δ 18O<-7.5‰为剔除标准,筛选出近 200个有效数据建立了碳同位素演化曲线,该曲线与前人在贵州建立的同期曲线具有很好的可对比性。曲线显示出海水碳同位素组成的巨大波动,变化幅度达 8.55‰,包括 2个完整的上升—下降旋回,第 1个旋回由飞仙关组和嘉陵江组第一段组成,第 2个旋回由嘉陵江组第二段、第三段和部分第四段组成,同时还包括若干短周期的次级旋回。利用曲线形态与前人建立的同期曲线进行了岩石地层与年代地层单位的对比,飞仙关组可大致与 Induan阶对应,嘉一段可大致与 Smithian亚阶对应,嘉二段、嘉三段和嘉四段下部可大致 Spathian亚阶对应;绿豆岩作为下—中三叠统界线其位置可能偏高,下—中三叠统界线应该位于绿豆岩之下的嘉四段内部。早三叠世海水碳同位素组成高频率大幅度的波动指示了生物大灭绝后环境的不稳定性,其中菌藻类等微生物灾后过度繁盛,可能导致了有机碳在短时间内的快速埋藏和海水碳同位素的正漂移;而碳同位素大规模的负漂移可能与甲烷水合物释放带来的轻碳有关。研究认为,白云岩所在的地层段往往具有更高的δ 13C值,这可能说明微生物活动、白云岩形成和海水δ 13C值升高之间的有机联系。下三叠统的白云岩、尤其是嘉二段和嘉四段的白云岩可能是微生物成因的,嘉陵江组有机碳的生产和埋藏可能主要是与白云岩形成有关的微生物的贡献,具最高δ 13C值的嘉二段的烃类在一定程度上可能具有自生自储的特点。
The Early Triassic,as an interval following the mass extinction at the Permo-Triassic boundary,representing the ecosystem reconstruction and recovery after the end-Permian biotic crisis,has been garnered much attention in the geological fields. In order to approach carbon isotopic composition and evolution of Early Triassic seawater,we examined the δ13C value of 258 carbonate samples from the Early Triassic(including adjacent strata)from Dukou section in Xuanhan County and Beibei section in Chongqing City,Sichuan Basin. Most of the samples preserve the initial carbon isotopic composition of seawater,but the original signal in the second and fourth Members of Jialingjiang Formation is much more poorly preserved than that of Feixianguan Formation and first and third Members of Jialingjiang Formation. Based on the principle that the oxygen isotope is more sensitive to alternation during the diagenetic process,the relationship between δ13C and δ18O was analyzed,and 200 samples with the δ18O <-7.5‰ were selected to construct the carbon isotopic evolution curve of the Early Triassic. This isotopic evolution curve shows finely comparable patterns with the coeval curve originating from Guizhou Province. The curve presents larger fluctuations in δ13C value of Early Triassic seawater. There are two complete ascending and descending cycles. The cycle 1 consists of Feixianguan Formation,and the first Member of Jialingjiang Formation;while the cycle 2 consists of the second and third Members and part of the fourth Member of Jialingjiang Formation,as well as several sub-cycles. According to the pattern of the curve,the lithostratigraphic unit is correlated with chronostratigraphic unit from the coeval curve of Guizhou Province originating from literatures,and further shows that the Feixianguan Formation is in accordance with the Induan Stage,the first Member of Jialingjiang Formation is corresponding to the Smithian substage,the second and third Members and part of the fourth Member of Jialingjiang Formation would be correlated to Spathian substage. Thus,the green-bean rock,as the boundary of Middle and Lower Triassic,is probably distributed in the Middle Triassic. The fluctuations of Early Triassic carbon isotopic curve,both in frequency and amplitude,would result from the perturbations of ecologic environments. The thriving of bacteria and algae in ocean ecosystems results in high burial efficiency of organic carbon and positive excursion of δ13C value. The negative excursion of δ13C might be the consequences of the hydrates methane release. The carbon isotopic evolution shows the tendency of commonly higher δ13C value in dolomite beds than in limestone beds,suggesting the interrelation among activity of microorganism,formation of dolomite and elevation of δ13C of seawater. The formation of dolomites in the Early Triassic,especially in the second Member of Jialingjiang Formation,might be originated from activity of microorganism,also as the main source for organic carbon burial. The hydrocarbon in dolomites of the second Member of Jialingjiang Formation is characterized by “in situ reservoir” to some degrees.
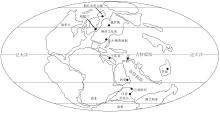
早三叠世是二叠纪末全球生物灭绝事件后地球生态修复与重建的关键时期, 记录该时间间隔全球碳循环的古海水碳同位素组成及变化长期以来被高度关注。然而, 三叠纪全球碳酸盐岩沉积盆地较小、碳酸盐沉积不发育(金振奎等, 2013), 早三叠世全球很多地方更是以巨厚的碎屑沉积为主(Korte et al., 2003), 这给该时间间隔期古海水信息的获取带来困难, 因而仅在全球范围内为数不多的地区建立了早三叠世碳同位素曲线(图 1)。在由Veizer等(1999)建立的为人们所广泛使用的显生宙碳同位素曲线中, 早三叠世时间段的分辨率明显较低, 这与人们对该时间间隔期重要地质事件的关注程度极不相称。
 | 图1 二叠纪— 三叠纪转折期全球古地理重建(黑点代表早三叠世海相碳酸盐碳同位素曲线研究位置; 底图据 Erwin, 1993, 有修改)Fig.1 Paleogeographic reconstruction during the Permo-Triassic transition(Black dots representing study areas of carbon isotopic curve of the Early Triassic marine carbonates; Base map modified from Erwin, 1993) |
已报道的早三叠世海相碳酸盐的碳同位素组成和演化趋势并不完全一致, 差异主要表现在δ 13C的数值大小和曲线的变化幅度上, 原因可能与样品的成岩蚀变性、岩性差别、地方性或区域性事件有关。已有的研究包括:Tong等(2002, 2007)和Zuo等(2006)对中国华南的安徽、江苏、浙江、湖北、广西以及贵州等地区多个剖面早三叠世碳酸盐碳同位素组成的系统研究, Korte等(2005)对欧洲和西亚碳酸盐碳同位素组成的研究(但其早三叠世数据缺乏整个Spathian亚阶和部分Smithian亚阶的样品), Horacek等(2007)对意大利北部碳酸盐碳同位素的研究, Payne等(2004, 2011)、Meyer等(2011a)对中国贵州多个早三叠世剖面碳同位素组成、演化趋势及控制因素的研究, 以及Huang等(2012)对中国上扬子地区重庆中梁山等剖面早三叠世碳酸盐碳同位素组成与演化趋势的研究等。
就样品类型而言, 用于早三叠世碳同位素组成研究的材料主要为全岩样品, 这与早三叠世腕足类生物(作为古海洋地球化学信息保存最好的材料之一)普遍缺乏有关; 另外, 一些研究(如Horacek et al., 2007)中使用了白云岩的数据, 但其从碳、氧同位素之间的相关性出发, 认为白云岩的碳同位素没有偏离海水, 只有氧同位素偏离了海水。然而, 一些新近的研究(Horita, 2014)以及文中后面的有关讨论表明, 白云岩可能比伴生的石灰岩更富13C, 因而有关机理仍然需要进一步研究。
虽然不同地区早三叠世海相碳酸盐的碳同位素组成和变化幅度并不完全一致, 但多数剖面数据都显示出碳同位素发生巨大波动, 最大值和最小值之间的差值达到10‰ , 甚至更高, 且多数研究剖面都检测到了很高的δ 13C值, 如贵州关刀剖面的δ 13C最大值达到6‰ , 该地区的另外2个剖面的δ 13C最大值达到了8‰ (Payne et al., 2004); 意大利北部早三叠世碳酸盐的δ 13C最大值接近6‰ (Horacek et al., 2007); 重庆中梁山等剖面早三叠世碳酸盐的δ 13C最大值达也达到了8‰ (Huang et al., 2012), 该值超过了显生宙其他时代海水的δ 13C最大值, 但该高δ 13C值持续时间不长, 这不同于二叠纪长时间的持续稳定的高δ 13C值(Huang and Zhou 1997; Veizer et al., 1999)。同时, 与早三叠世相邻的二叠纪大部分时间和中三叠世海水的碳同位素的变化都十分稳定, δ 13C值的变化幅度均很少超过3‰ (Huang and Zhou, 1997; Payne et al., 2004)。这显示出早三叠世海相碳酸盐碳同位素组成具有高频率、大幅度变化的特点。早三叠世全球范围大陆植被的减少、海洋钙质无脊椎动物和海相碳酸盐数量的减少、海洋系统中菌藻类的过度繁盛以及大洋周期性的分层可能是早三叠世海水具有极高的δ 13C值和同位素组成大幅度波动的重要原因(Payne et al., 2004; Horacek et al., 2007; Huang et al., 2012)。
国内已有报道的早三叠世海相碳酸盐的碳同位素研究仍然十分有限, 且样品密度难以刻划出一些短周期的变化。文中综合了四川省宣汉渡口与重庆北碚2个剖面共258个早三叠世(包括相邻的晚二叠世末和中三叠世初)海相碳酸盐的碳、氧同位素数据, 在讨论碳同位素组成对海水代表性及对数据筛选的基础上, 建立了相应的演化曲线, 以研究早三叠世海水的碳同位素组成特征、变化规律及可能的控制因素。
文中涉及的下三叠统及相邻地层的样品采自四川盆地东北部的四川宣汉渡口剖面和该盆地东部的重庆北碚剖面。构造位置上, 重庆北碚剖面属于北东向的华蓥山构造带, 宣汉渡口剖面属于大巴山北西向构造带与华蓥山北东向构造带交汇处(图 2)。
 | 图2 四川盆地宣汉渡口和重庆北碚剖面位置Fig.2 Location of Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
研究剖面下三叠统由飞仙关组和嘉陵江组构成(图2)。飞仙关组位于上二叠统含燧石结核生物灰岩之上, 两者之间以同期火山碎屑成因的黏土岩为界。该界线为古生界与中生界的界线, 是一个被国际认可的界线, 黏土岩中锆石SHRIMP的U-Pb同位素年龄为252.6± 0.2, Ma(Mundil et al., 2004)。中国西南地区下— 中三叠统均以夹于碳酸盐岩(或蒸发岩)中的火山碎屑岩(已普遍蚀变成水云母黏土岩, 俗称绿豆岩)为界(图 3), 由于火山喷发作用的等时性, 该界线已成为华南下— 中三叠统对比的重要标志。在某些近期成果(谢韬等, 2013)中, 罗平生物群(安尼期)下部位于下— 中三叠统界线附近的绿豆岩的年龄数据为246.6± 1.4, Ma。因此, 本研究涉及的样品主要介于2套火山碎屑沉积之间, 年龄值位于252.6± 0.2— 246.6± 1.4, Ma之间。
 | 图3 四川盆地东部重庆北碚剖面下三叠统嘉陵江组和中三叠统雷口坡组的界线Fig.3 Boundary between the Lower Triassic Jialingjiang Formation and Middle Triassic Leikoupo Formation at Beibei section in Chongqing City, eastern Sichuan Basin |
北碚剖面的飞仙关组陆源碎屑含量较高, 并具有一定的四分性, 自下而上为:第一段为杂色泥质微晶灰岩、钙质泥岩和粉砂岩; 第二段为紫红色钙质泥岩夹泥质微晶灰岩; 第三段为灰岩、泥岩、泥质微晶灰岩夹鲕粒灰岩; 第四段为紫色、黄绿色钙质泥岩与泥质微晶灰岩。飞仙关组岩性段的可对比性较差, 如在不同的地点, 飞仙关组的鲕粒灰岩可分布于不同的岩性段。宣汉渡口剖面的飞仙关组中不仅陆源碎屑含量显著较少, 而且也缺乏明显的四分性, 除紫红色钙质泥岩以外, 主要由灰岩构成, 包括鲕粒灰岩、生物碎屑灰岩和微晶灰岩等, 顶部和底部发生白云岩化。考虑到这类灰岩样品更有利于进行氧、碳同位素分析, 因此文中飞仙关组的样品采集选择了渡口剖面。需要在此进一步说明的是, 在1︰200000地质图中, 渡口剖面的这套地层被称为大冶组, 但考虑到近年来与文中研究有关的大多数文献仍然将其称为飞仙关组, 因而文中仍然使用飞仙关组这一岩石地层单位名称。
研究涉及的2个剖面中, 嘉陵江组都有良好的四分性和可对比性:第一段和第三段主要为微晶灰岩夹透镜状生物屑灰岩、内碎屑灰岩和鲕粒灰岩等; 第二段和第四段主要为蒸发盐岩和白云岩、地表发育盐溶角砾岩和次生灰岩, 在地表剖面中也很容易被区分和识别。但宣汉渡口剖面嘉陵江组上部因地层发生褶皱而使得采样困难, 且作为下— 中三叠统界线的绿豆岩也难于被识别, 因而嘉陵江组上部(第三段和第四段)和雷口坡组底部样品的采集选择了绿豆岩发育良好且出露清楚的北碚剖面(图 3), 这使得下— 中三叠统界线附近的样品具有更好的可靠性。
2个剖面共采集样品258个:185个样品采自宣汉渡口剖面, 涉及地层包括二叠系顶部, 下三叠统飞仙关组和嘉陵江组第一段和第二段; 73个样品采自重庆北碚剖面, 涉及地层包括下三叠统嘉陵江组第三段和第四段以及中三叠统雷口坡组底部。
采自宣汉渡口剖面的185个样品采用微钻取样, 以获得岩石中的微晶部分而回避方解石脉和亮晶胶结物, 但由于微钻获得样品的数量较少、难以用常规化学分析方法进行CaO、MgO、Mn、Sr和Fe含量等分析, 因而这些样品仅做了氧、碳同位素分析。由于对采自重庆北碚剖面的73个样品要同时进行元素分析和同位素分析, 需要的样品数量较大, 因而采用手工选样, 首先对样品在宏观上进行了分拣, 尽可能地回避了方解石脉和晶洞中的方解石造成的影响(将另文讨论这些样品元素构成及同位素组成的关系, 文中仅涉及这些样品的碳、氧同位素组成)。
对大多数用于同位素分析的样品都进行了薄片鉴定和阴极发光检测, 以配合对样品的选择。样品经挑选后粉碎至200目进行氧、碳同位素分析(宣汉渡口剖面的样品), 或缩分后分别用作CaO、MgO、Mn、Sr和Fe含量分析以及碳、氧同位素分析(重庆北碚剖面的样品)。
碳、氧同位素分析由中国科学院南京地质古生物研究所完成, 采用Finigan MAT 253气体同位素质谱仪, 分析结果相对于PDB标准, 以δ 值表示, 采用GBW-04405标样, δ 13C和 δ 18O 测定值标准偏差分别小于0.04‰ 和0.08‰ (PDB)。阴极发光分析由成都理工大学油气藏地质及开发工程国家重点实验室完成, 使用剑桥仪器公司CL8200MK5阴极发光仪(配以Leica偏光显微镜), 典型测试条件为束电压10 kV, 束电流300 μ A。
2个采样剖面的下三叠统及相邻地层均是在开挖公路时被揭露, 已经历了数十年的暴露, 同时大气水的作用深度可达地下百米甚至更深(黄思静, 2010), 在南方潮湿气候条件下更是如此, 说明大气水以及埋藏成岩过程中的孔隙水等非海相流体对碳酸盐原始海水碳氧同位素组成、尤其是氧同位素组成的影响是肯定存在的, 因而有必要针对样品的碳同位素组成对海水的代表性进行评估。
| 表1 四川盆地宣汉渡口和重庆北碚剖面下三叠统海相碳酸盐样品δ 13C 和 δ 18O 值汇总结果 Table1 Summary statistics results of δ 13C and δ 18O values from the Lower Triassic marine carbonate samples at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
表1给出了四川盆地宣汉渡口和重庆北碚剖面下三叠统(包括相邻的二叠系顶部和中三叠统底部)258个海相碳酸盐样品δ 13C 和 δ 18O 值按地区和层位的汇总结果。由于大多数样品都没有进行CaO、MgO、Mn、Sr和Fe含量分析, 因而难于从样品的Mn、Sr、Fe含量和Mn/Sr比等角度评估样品碳同位素组成对海水的代表性, 因而文中选择了从碳、氧同位素关系角度讨论这些样品碳同位素组成对海水的代表性。
海相碳酸盐是自然界中最为重要的无机碳库, 地壳中约有80%的碳存储于碳酸盐岩中(Kump et al., 1999)。与碳酸盐岩相比, 成岩流体中的碳是非常有限的, 成岩过程对碳来说总体上表现为岩石缓冲; 与碳不同的是, 氧主要存在于水中, 因而成岩过程对氧来说, 总体上表现为水缓冲。碳酸盐的氧同位素组成在成岩作用过程中非常容易变化(Horacek et al., 2007), 而碳同位素则变化十分有限。基于这样的原理, 人们经常用样品的氧、碳同位素之间的相关性来判断碳同位素对海水的代表性(Horacek et al., 2007), 如果二者之间存在相关性, 则表明成岩过程中氧同位素改变时, 发生了碳同位素的协同变化, 碳同位素对海水的代表性较差; 如果二者之间不存在相关性, 或相关性很差, 则表明成岩过程中氧同位素改变时, 碳同位素没有发生变化或变化很小, 碳同位素对海水的代表性较好。
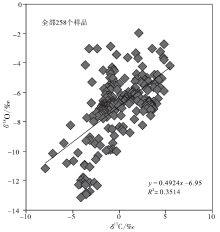
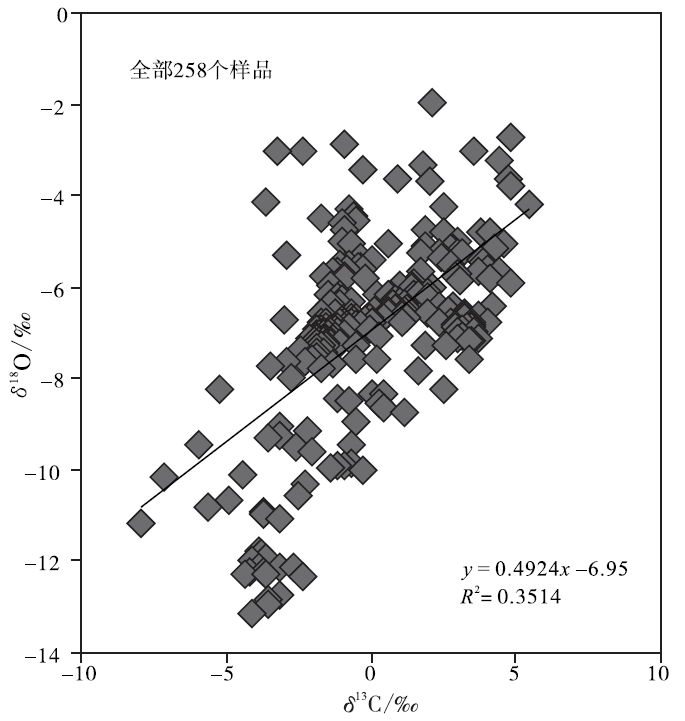 | 图4 四川盆地宣汉渡口和重庆北碚剖面全部258个碳酸盐样品的 δ 18O 和δ 13C值投点图Fig.4 Cross-plots of δ 18O values vs δ 13C values for the total 258, marine carbonate samples at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
图4是四川盆地宣汉渡口和重庆北碚剖面下三叠统全部258个海相碳酸盐样品的 δ 18O 和δ 13C值投点图, 该图显示这些样品的 δ 18O 和δ 13C值之间存在良好的相关性, 相关系数接近0.6, 说明就全部样品而言, 在海相碳酸盐岩的成岩过程中, 一部分样品的碳同位素对海水的代表性较差, 需要在建立演化曲线时予以剔除。
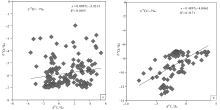
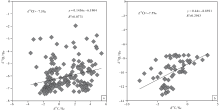
基于氧同位素可以更为敏感地反映海相碳酸盐成岩蚀变性的考虑, 文中按氧同位素组成对宣汉渡口和重庆北碚剖面全部258个样品进行了分组并考查其 δ 18O 和δ 13C值的相关性(图 5; 图6; 图7)。图5是关于 δ 18O> -7‰ 和δ 18O< -7‰ 样品的对照, 该图显示δ 18O< -7‰ 的样品其 δ 18O 和δ 13C值的相关性较好, 确定系数0.317; 而δ 18O> -7‰ 的样品其 δ 18O 和δ 13C值的相关性较差, 确定系数只有0.0305, 可以认为基本没有相关性。与之类似:图6是关于 δ 18O> -7.5‰ 和δ 18O< -7.5‰ 样品的对照, 图7是关于 δ 18O> -8‰ 和δ 18O< -8‰ 样品的对照, 显示δ 18O< -7.5‰ 的样品和 δ 18O < -8‰ 的样品其 δ 18O 和δ 13C值的相关性显著好于 δ 18O> -7.5‰ 和 δ 18O> -8‰ 的样品, 后者的 δ 18O 和δ 13C值基本上没有相关性。
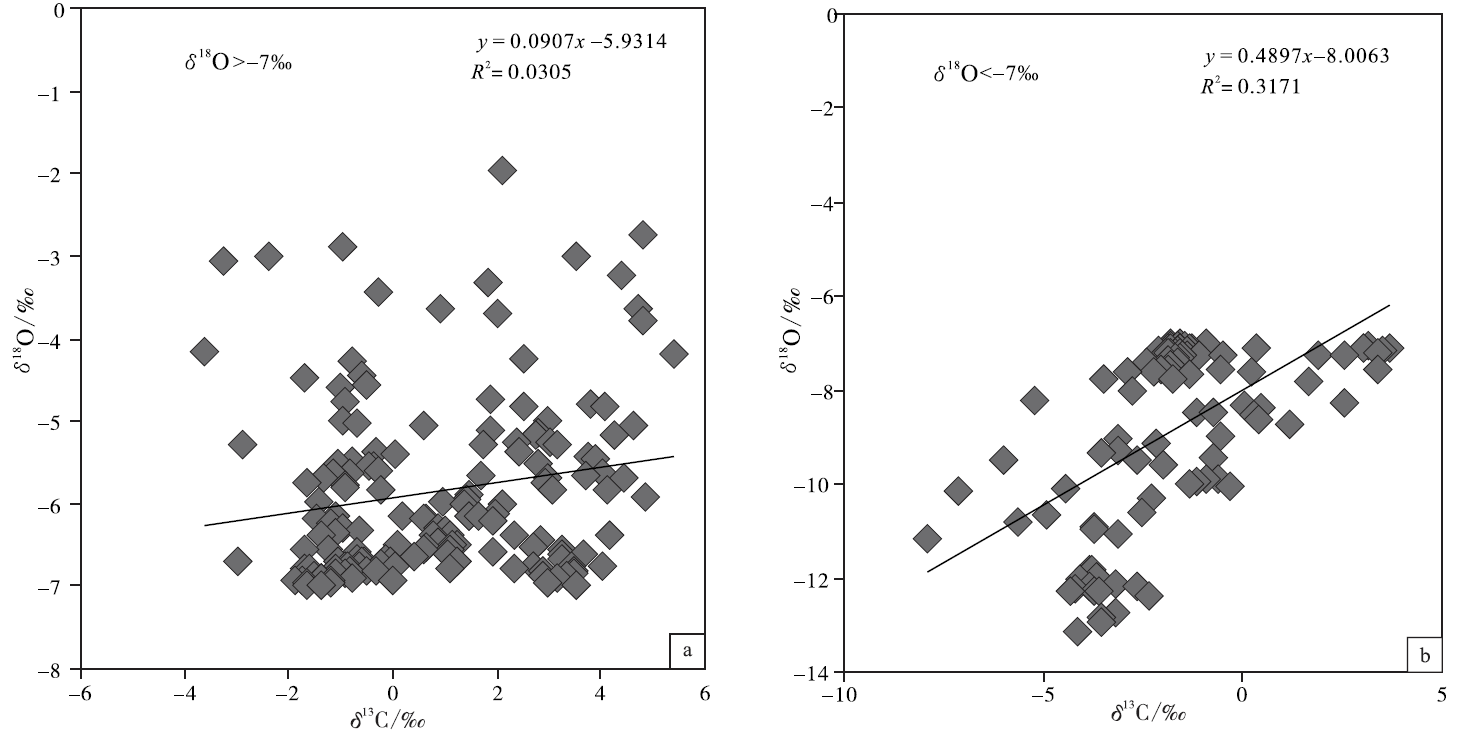 | 图5 四川盆地宣汉渡口和重庆北碚剖面海相碳酸盐样品 δ 18O 和δ 13C值投点图(a图数据来自于 δ 18O> -7‰ 的样品; b图数据来自于 δ 18O< -7‰ 的样品)Fig.5 Cross-plots of δ 18O values vs δ 13C values for marine carbonate samples with δ 18O> -7‰ (a) and δ 18O< -7‰ (b)at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
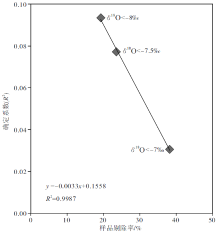
如果以δ 18O< -7‰ 为剔除标准, 则全部258个样品中有99个将被剔除, 剔除率为38.37%; 如果以δ 18O< -7.5‰ 为剔除标准, 则全部258个样品中有61个将被剔除, 剔除率为23.64%; 如果以δ 18O< -8‰ 为剔除标准, 则全部258个样品中有50个将被剔除, 剔除率为19.38%。可以看出来, 设置的条件越苛刻, 剔除的样品数量越多, 剔除率越高, 则剔除后剩余样品的 δ 18O 和δ 13C值的相关性越差(图 8), 说明剔除后剩余样品的成岩蚀变性较弱, 对海水的代表性也相对较好。
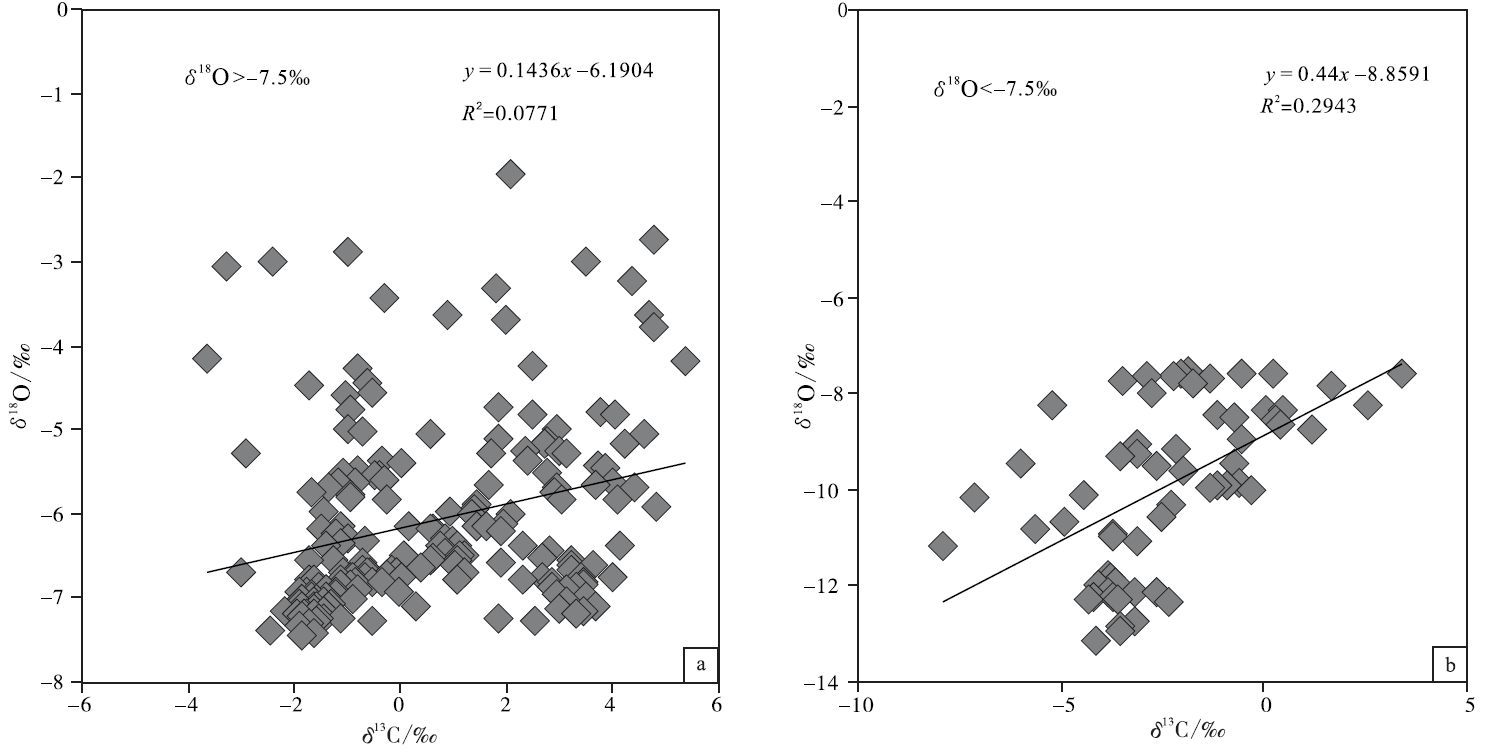 | 图6 四川盆地宣汉渡口和重庆北碚剖面海相碳酸盐样品 δ 18O 和δ 13C值投点图(a图数据来自于δ 18O> -7.5‰ 的样品; b图数据来自于δ 18O< -7.5‰ 的样品)Fig.6 Cross-plots of δ 18O values vs δ 13C values for marine carbonate samples with δ 18O> -7.5‰ (a)and δ 18O< -7.5‰ (b) at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
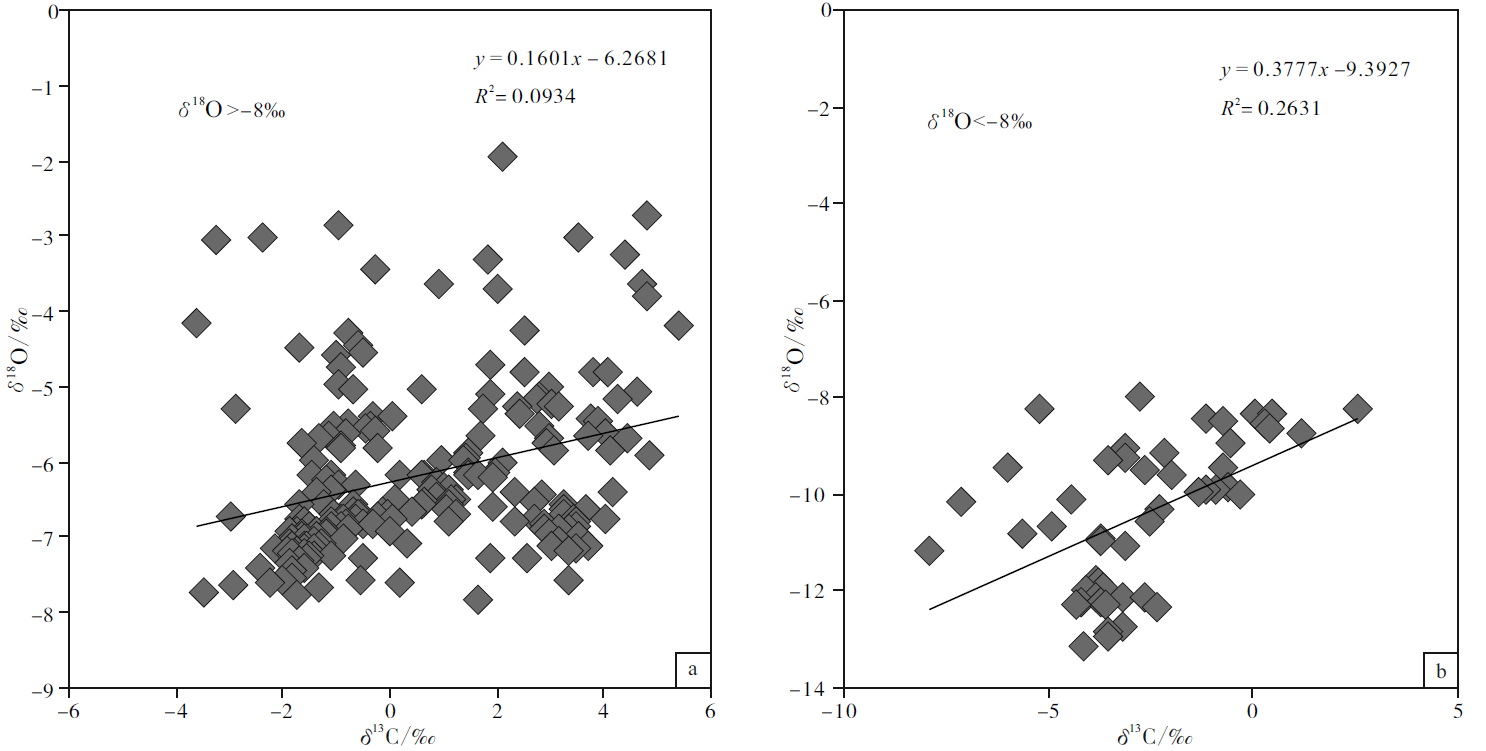 | 图7 四川盆地宣汉渡口和重庆北碚剖面海相碳酸盐样品 δ 18O 和δ 13C值投点图(a图数据来自于δ 18O> -8‰ 的样品; b图数据来自于δ 18O< -8‰ 的样品)Fig.7 Cross-plots of δ 18O values vs δ 13C values for marine carbonate samples with δ 18O> -8‰ (a)and δ 18O< -8‰ (b)at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
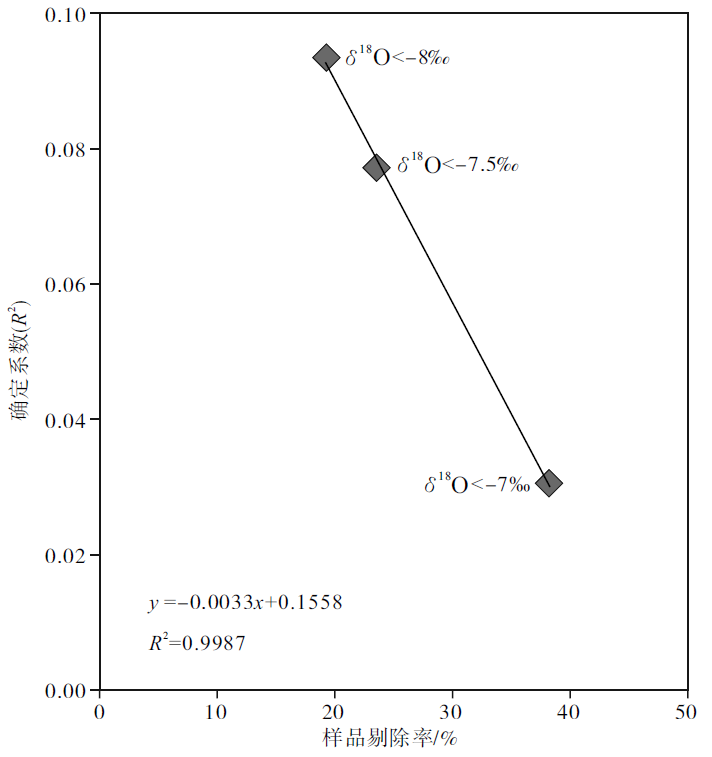 | 图8 四川盆地宣汉渡口和重庆北碚剖面海相碳酸盐样品的剔除率和剔除后剩余样品的 δ 18O 和δ 13C值的确定系数投点图Fig.8 Cross-plots of winkled ratio vs δ 18O/δ 13C determination coefficient of remind marine carbonate samples after winkling from Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
在最终完成的样品碳同位素组成与地层累积厚度投点图中, 笔者使用了全部数据, 但在完成碳同位素演化曲线的过程时, 所有的δ 18O< -7.5‰ 的样品(显然包括了δ 18O< -8‰ 的样品)都没有予以考虑, 这61个样品都可以被认为是被剔除的样品, 占全部258个样品的23.64%(表 2)。如上述讨论, 建立演化曲线时不考虑这些样品的理由是因为它们具有较低的氧同位素组成, 受到了 δ 18O 值较低的非海相流体的影响, 因而其碳同位素组成也可能不同程度地受到了这些非海相流体影响。实际上, 也可以从碳同位素演化曲线(图 9-a)中看出, 这些样品的大多数、尤其是其中的 δ 18O< -8‰ 的样品都显著偏离了主流演化曲线。在完成碳同位素演化曲线的过程中, 文中使用了一部分 δ 18O值介于-7.5‰ ~-7‰ 之间的样品, 这些样品主要分布在嘉陵江组第一段顶部, 但可以从演化曲线(图 9-a)中看出, 这些样品中的绝大多数并没有显著偏离主流演化曲线。
从表2可以看出, 被剔除的δ 18O< -7.5‰ 的61个样品主要分布在嘉陵江组第二段和第四段, 尤其是嘉陵江组第四段, 该段地层中被剔除的样品数达到了31个, 占嘉四段全部41个样品的75.61%, 而嘉陵江组第二段被剔除的样品数也达到了15个,
| 表2 四川盆地宣汉渡口和重庆北碚剖面下三叠统按不同的 δ 18O 值所需要剔除的样品数和剔除率 Table2 Numbers of winkled samples and winkled ratio based on δ 18O values of the Lower Triassic samples at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin |
占嘉二段全部52个样品的28.85%, 其余各地层组段需要被剔除的样品数量都很少, 这显示除嘉二段和嘉四段外, 2个剖面中的碳酸盐岩样品对海水的碳同位素组成总体上都保存较好。嘉二段和嘉四段具有如此高的样品剔除率的原因显然与这2个地层段主要由蒸发盐— 白云岩构成有关, 在地表剖面中, 这2个地层段基本上都是由盐溶角砾岩和次生灰岩构成, 而这2种岩石都是近地表条件下由于蒸发盐的溶解和夹层碳酸盐岩的破碎、崩塌而形成的, 与去膏化作用有关的次生方解石在这些岩石中占了相当大的比例(如岩溶角砾岩)、甚至构成了这些岩石的主体(如次生灰岩)。南方潮湿气候条件的表生成岩环境明显受大气水、富含有机碳源CO2的地下水和细菌参与的有机碳氧化环境的影响, 这使得富含次生方解石的嘉二段和嘉四段近地表环境的碳酸盐岩普遍具有较低的碳、氧同位素组成。从表2可以看出, δ 18O< -8‰ 的样品基本上都分布在嘉陵江组第二段和第四段, δ 18O< -10‰ 的27个样品中有24个都集中在嘉陵江组第四段, 有2个分布在嘉陵江组第二段。
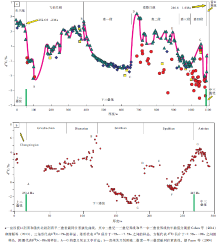
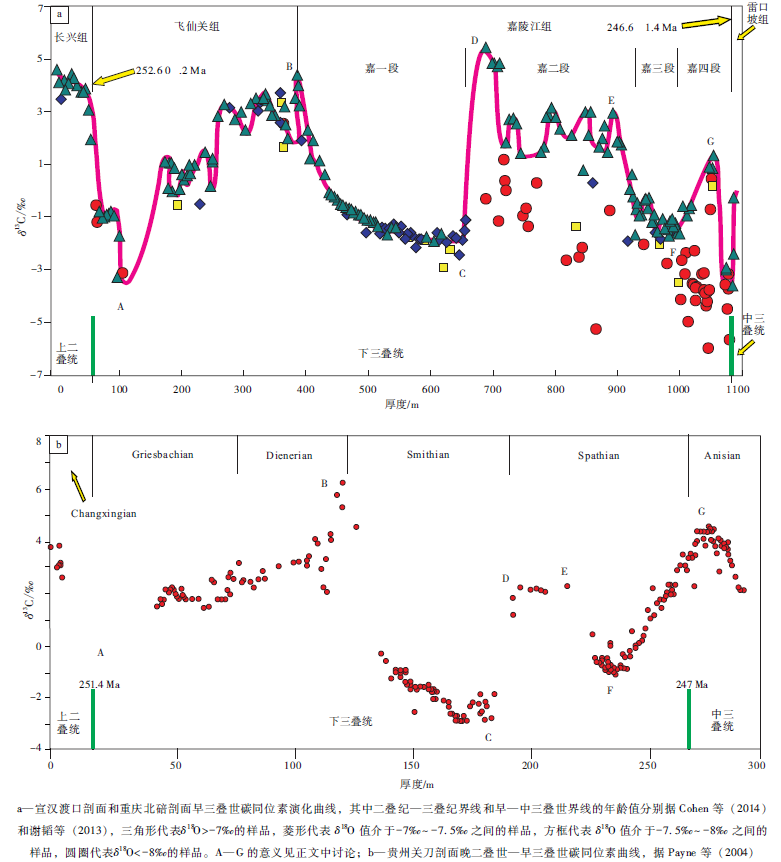 | 图9 四川盆地宣汉渡口和重庆北碚剖面早三叠世碳同位素演化曲线及其与贵州关刀剖面同时代曲线的对比Fig.9 Comparison of δ 13C isotope curve of the Early Triassic at Dukou section in Xuanhan County and Beibei section in Chongqing City, Sichuan Basin(a)and coeval carbon isotope evolution curve at Guandao section, Guizhou Province(b) |
图9-a是笔者建立的早三叠世(含相邻的晚二叠世末和中三叠世初)的碳同位素演化曲线, 该曲线中的上二叠统顶部、下三叠统飞仙关组、嘉陵江组第一段和第二段的数据来自宣汉渡口剖面; 嘉陵江组第三段和第四段的数据以及雷口坡组底部的数据来自重庆北碚剖面。该碳同位素曲线主要依据δ 18O> -7‰ 的样品的碳同位素数据, 仅在嘉一段顶部使用了 δ 18O 值介于-7‰ ~-7.5‰ 之间的样品的碳同位素数据, 没有使用δ 18O< -7.5‰ (显然包括了δ 18O< -8‰ )的样品数据。曲线的早三叠世部分正好介于2个有放射性同位素年龄值的火山碎屑岩之间, 因而曲线主体部分的年龄时限介于这2个年龄值(252.6± 0.2— 246.6± 1.4, Ma)之间, 时间间隔约为6, Ma, 但该时间间隔的长度超过了地质年代表(Cohen et al., 2014)所给出的早三叠世4.97, Ma的时间长度。后面笔者将进一步讨论这2个火山碎屑岩作为二叠纪— 三叠纪界线和早— 中三叠世界线的合理性。图9-b给出了综合的贵州晚二叠世— 早三叠世碳同位素曲线(Payne et al., 2004), 以便和文中建立的曲线(图9-a)进行对比(包括2个地区同时代碳同位素曲线形态的对比以及相应的岩石地层单位和年代地层单位的对比)。
中国华南的二叠系— 三叠系界线是一个被国际上认可的、且容易识别的直观界线, 前人(Claoué -Long et al., 1991; Mundil et al., 2004)曾多次测试过该界线黏土层锆石的U-Pb同位素年龄, 较新的数值为252.6± 0.2, Ma(Mundil et al., 2004), 图9-b的贵州关刀剖面也给出了一个类似的年龄值(251.4, Ma)。另外, 二叠系— 三叠系界线附近的碳酸盐岩还具有人们所熟知的δ 13C极小值, 而该极小值也存在于全球范围内其他剖面的二叠系— 三叠系界线附近(陈锦石等, 1984; Holser and Magarita 1987; Magarita et al., 1988; 黄思静, 1994; Kozur, 2003)。宣汉渡口剖面的二叠系— 三叠系界线附近也检测到了这一极小值(-3.125‰ )。尽管贵州关刀剖面二叠系— 三叠系界线附近尚没有检测到这一碳同位素的极小值(Payne et al., 2004), 但在其他的贵州二叠系— 三叠系界线附近也检测到了这一极小值。与Payne等(2004)综合的碳同位素曲线类似, 该极小值位于下三叠统底部, 二叠系— 三叠系界线在δ 13C值发生急剧变化的2个样品(δ 13C值大致由2‰ 降至-1‰ , 降幅约3‰ )之间。海相碳酸盐的δ 13C极小值代表了地质界线附近生物的灭绝与更替, 由于界线附近发生生物的群体灭绝, 故生物对有机碳的生产量会大幅度降低, 如果同时伴随有机碳的氧化, 则当有机碳的氧化量大于埋藏量时, 被氧化的有机碳进入大气中并溶解于海水, 最终造成同期海相碳酸盐δ 13C值的降低。当不适应环境的老种属灭绝、适应环境的新种属出现, 其对有机碳的生产量逐渐增加, 有机碳的埋藏量相应增加, 碳酸盐δ 13C值将止跌回升。因而海相碳酸盐的δ 13C极小值代表了适应环境的新种属生物产生的有机碳在数量上已能取代灭绝的老种属, 并与氧化有机碳达到平衡的转折点, 它代表了地质历史中生物的灭绝更替转折点。
中国西南地区下— 中三叠统界限以火山碎屑成因的水云母黏土岩(俗称“ 绿豆岩” )为界。火山喷发作用所具有的等时性使该界线成为西南地区下— 中三叠统界限对比的直观标志, 近期成果如Lehrmann 等(2006)获得的下— 中三叠统界线年龄应介于247.32± 0.08, Ma和247.13± 0.12, Ma之间, 谢韬等(2013)报道的罗平生物群(安尼期)下部位于下— 中三叠统界线附近的绿豆岩的年龄数据为246.6± 1.4, Ma。由于北碚剖面的绿豆岩发育良好, 因而绿豆岩附近样品的碳同位素组成应代表了西南地区下— 中三叠统界线附近的碳同位素组成。图9-a中绿豆岩附近样品存在δ 13C值的负偏移, 但贵州关刀剖面的该界线附近的负偏移出现在中三叠统Anisian阶内部, 给出的放射性年龄值为247, Ma(用牙形石带来限制中— 晚三叠世的时间尺度), 并且早期的报道证实Spathian-Anisian界限存在碳同位素的正漂移(Atudorei and Baud, 1997), 因而中国西南地区的绿豆岩应该位于中三叠世地层中, 绿豆岩作为下— 中三叠统界线的古生物地层学的合理性可能还需要继续研究, 这也包括对更多剖面进行的放射性年龄研究以及界线附近样品的稳定同位素研究等。
在全球范围内, 二叠纪大部分时间段的δ 13C值具有高的且稳定的演化趋势(Huang and Zhou 1997; Veizer et al., 1999), 图9-a记录了厚度约60, m的二叠系顶部碳酸盐岩的δ 13C值, 代表了从二叠纪δ 13C高值到二叠纪— 三叠纪界线附近低值的变化, 其后是整个早三叠世海水δ 13C值的大幅度、高频率的波动。从海水碳同位素组成的角度来说, 飞仙关组沉积时间代表了一个有效的生态修复期, 有机碳埋藏速度持续稳定增加(由于消失的海洋物种尚未复苏, 因而菌、藻类等微生物在有机碳的生长与埋藏中起至关重要的作用), 并大于其氧化量, 使得该时间间隔中海水的碳同位素组成总体上表现为低幅波动的持续稳定升高, δ 13C值从二叠系— 三叠系界线附近的约-3‰ 升至飞仙关组— 嘉陵江组界线处的4.4‰ (图 9-a中的A— B段), 上升幅度超过7‰ 。对照Payne等(2004)的贵州关刀晚二叠世— 早三叠世碳同位素曲线可以看出, 二叠纪— 三叠纪界线之后的海水δ 13C值的升高持续到Induan/Olenekian期的界线处(图 9-b中的B点), 因而飞仙关组可大致与Induan阶所对应, 按2014的地质年代表(Cohen et al., 2014), 该生态修复时期持续时间约为0.97, Ma。
图9-a中的B— C段代表了海水碳同位素组成的稳定持续降低时期, δ 13C值从飞仙关组顶部(与贵州关刀晚二叠世— 早三叠世曲线(Payne et al., 2004)的Induan/Olenekian期的界线对应)的4.4‰ 降至嘉陵江组第一段顶部的-1.8‰ , 下降幅度超过6‰ 。由于嘉陵江组第一段发育大套的微晶灰岩, 样品采集容易, 基本上没有需要剔除的样品(图 9-a), 因而获得的碳同位素组成对海水的代表性较好。在贵州关刀晚二叠世— 早三叠世碳同位素曲线(Payne et al., 2004)中也可以清楚地识别出与之对应的δ 13C值的下降段(图 9-b中的B— C段), 该下降段基本上分布于Smithian亚阶, 因而嘉陵江组第一段可大致与Smithian亚阶对应。Smithian亚期中海水δ 13C值的持续降低原因并不十分清楚, 一般的解释是有机碳埋藏量的减少和氧化量的相对增加, 或与甲烷的释放有关(Erwin, 1993), 故这种短时间内海水碳同位素组成的频繁变化可能与生物缓慢复苏过程中碳循环的不稳定性有关。
图9-a中的C— D段是一个短时间内海水碳同位素组成强烈正漂移区间, δ 13C值从嘉一段顶部的-1.8‰ 上升到嘉二段底部的5.4‰ , 上升幅度超过7‰ , 基本上没有检测到在碳同位素组成上的过渡样品。贵州关刀剖面也存在类似情况(Payne et al., 2004), 即δ 13C值从-3‰ 升至2‰ 以上, 上升幅度也超过5‰ , 也较为缺乏碳同位素组成上的过渡样品, 人们至今还找不到完全合理的机制来解释如此短时间内海水碳同位素组成发生如此强烈正漂移。
图9-a中的D— E段代表了作为白云岩— 蒸发盐段低幅振荡的持续的高δ 13C值区间, 海水δ 13C值主要分布在1‰ ~3‰ 的范围内。该时间间隔期海水的高δ 13C值可能显示了嘉二段白云岩为微生物成因, 这些白云岩可能是甲烷形成过程中的副产物。作为灰岩段的嘉三段具有与嘉一段类似的碳同位素组成, δ 13C值从嘉二段的1‰ ~3‰ 的高值降至0‰ 附近, 再缓慢降至-2‰ (图 9-a中的F点)。到嘉四段的白云岩— 蒸发盐段, δ 13C值再重新升至约1‰ 的较高值(图 9-a中的G点)。因此, 嘉二段、嘉三段和嘉四段下部基本构成一个不对称的“ V” 字形, 该“ V” 字形区间可大致与贵州关刀剖面Spathian亚阶对应(Payne et al., 2004)(图 9-b中的D— F区间和F— G区间的大部分)。嘉四段上部样品, 包括G点所在的峰值附近以及作为下— 中三叠统界线的绿豆岩都已落在贵州关刀剖面中三叠统的Anisian阶(Payne et al., 2004), 因而将绿豆岩作为下— 中三叠统界线其位置可能偏高。嘉四段白云岩的成因与嘉二段类似, 属于微生物成因的准同生白云岩, 但嘉四段微生物的生产能力不如嘉二段, 嘉二段的天然气在一定程度上可能是自生自储的。
与前人已建立的曲线类似, 宣汉渡口和重庆北碚剖面早三叠世碳同位素曲线的主要特点是其代表了早三叠世这一很短时间内(分别按252.2, Ma和247.1, Ma作为二叠纪— 三叠纪和早— 中三叠世界线的年龄值(Cohen et al., 2014), 该时间跨度只有5.1, Ma)全球海水碳同位素组成的巨大波动, 碳同位素组成从二叠系— 三叠系界线附近飞仙关组底部(大致对应Griesbachian亚阶下部)的-3.125‰ 的δ 13C最小值, 变化至嘉二段底部(大致对应Spathian亚阶下部)的5.421‰ , 变化幅度为8.55‰ 。该曲线包括2个完整的上升— 下降旋回, 第1个旋回由飞仙关组(大致对应Induan阶)和嘉陵江组第一段(大致对应Smithian亚阶)组成; 第2个旋回由嘉陵江组第二段、第三段和第四段部分地层(大致对应Spathian亚阶)组成, 同时还包括若干短周期的次级旋回。碳同位素组成的这种高频率、大幅度的变化只发生在早三叠世, 与之相邻的二叠纪和中三叠世大部分时间内海水碳同位素变化都十分稳定, δ 13C值的变化幅度很少超过3‰ 。前人(Retallack, 1999; Krull and Retallack, 2000; Dickens, 2003; Payne et al., 2004, 2011; Horacek et al., 2007; Meyer et al., 2011a, 2011b; Huang et al., 2012)对此已有过很多讨论, 最主要的解释是生物群体灭绝后的生态复苏过程中环境的不稳定性对全球碳循环和海水碳同位素组成的影响。由于早三叠世海水碳同位素信息获取的相对困难性, 因此一些推论仍然需要更多数据的积累和更高分辨率的演化曲线来证实。归纳前人的研究, 早三叠世海水的高δ 13C值和碳同位素组成的大幅度变化可能主要与2个原因有关:(1)菌、藻类等微生物的过度繁盛。生物群体灭绝后全球大量海洋物种消失, 存活的菌、藻类无法被食物链高端的物种摄取而过度繁盛, 碳同位素的强烈波动和δ 13C高值可能与菌、藻类阵发性的繁盛有关。(2)碳同位素大规模的负漂移可能与甲烷水合物释放带来的轻碳有关。另外, 要进一步强调的是, 在宣汉渡口和重庆北碚剖面中, 白云岩所在的地层段往往具有更高的δ 13C值, 而一些灰岩段则具有较低的δ 13C值, 这种情况在嘉陵江组尤其明显, 这可能说明微生物活动、白云岩形成和海水δ 13C值升高之间存在有机联系。换句话说, 下三叠统的白云岩, 尤其是嘉二段和嘉四段的白云岩, 可能是微生物成因的, 嘉陵江组有机碳的生产与埋藏可能主要是与白云岩形成有关的微生物的贡献, 具最高δ 13C值的嘉二段的烃类在一定程度上可能具有自生自储的特点。
1)宣汉渡口和重庆北碚剖面下三叠统海相碳酸盐岩对海水碳同位素组成具有不同的保存性, 主要作为灰岩段的飞仙关组、嘉陵江组第一段和第三段保存较好, 而主要作为蒸发盐— 白云岩段的嘉陵江组第二段和第四段保存较差。
2)基于氧同位素可以更为敏感地反映海相碳酸盐成岩蚀变性的考虑, 主要以 δ 18O < -7.5‰ 为剔除标准, 对全部258个样进行了筛选, 剔除的样品主要分布于嘉陵江组第二段和第四段, 在此基础上, 利用近200个有效样品建立了下三叠统碳酸盐的碳同位素演化曲线, 该曲线与前人在贵州建立的同期曲线具有较好的可对比性。
3)利用碳同位素曲线形态, 与前人建立的同期曲线进行了岩石地层单位与年代地层单位的对比结果表明, 飞仙关组可大致与Induan阶对应, 嘉一段可大致与Smithian亚阶对应, 嘉二段、嘉三段和嘉四段下部可大致与Spathian亚阶对应, 绿豆岩作为下— 中三叠统界线其位置可能偏高, 下— 中三叠统界线可能位于嘉四段内部。
4)宣汉渡口和重庆北碚剖面早三叠世碳同位素曲线的主要特点是其代表了早三叠世这一很短时间段内海水碳同位素组成的巨大波动, 变化幅度达8.55‰ , 包括2个完整的上升— 下降旋回, 第1个旋回由飞仙关组(大致对应Induan阶)和嘉一段(大致对应Smithian亚阶)组成; 第2个旋回由嘉二段、嘉三段和部分的嘉四段(大致对应Spathian亚阶)组成, 同时还包括若干短周期的次级旋回。
5)结合前人研究成果, 认为早三叠世海水碳同位素组成具有高频率、大幅度变化的主要原因是生物大灭绝后的延迟复苏过程中环境的不稳定性对碳循环的影响, 海洋物种大量消失造成灾变中存活下来的菌、藻类等微生物无法被充分摄取而过度繁盛, 并成为有机碳的主要原始创造者, 碳同位素大规模的负漂移可能与甲烷水合物释放带来的轻碳有关。
6)在宣汉渡口和重庆北碚剖面, 白云岩所在的地层段往往具有更高的δ 13C值, 这可能说明微生物活动、白云岩形成和海水δ 13C值升高之间存在有机联系, 下三叠统的白云岩、尤其是嘉二段和嘉四段的白云岩可能是微生物成因的, 嘉陵江组有机碳的生产与埋藏可能主要是与白云岩形成有关的微生物的贡献, 具最高δ 13C值的嘉二段的烃类在一定程度上可能具有自生自储的特点。
致谢 沉积地质研究院研究生袁桃、胡博、黄树光、李江勇、罗文、胡颖和陈永梅等参加野外剖面测量、样品采集工作以及室内的薄片分析、阴极发光分析和样品的加工处理工作, 中国科学院南京地质古生物研究所陈小明和刘静高级工程师在百忙中为本项目进行同位素分析并在样品处理方面给予多方指导, 匿名审稿人对论文提出了建设性的修改意见, 在此表示衷心感谢。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|