


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
现代叠层石的多样化构成:认识古代叠层石形成的关键和窗口*
[梅冥相 , 孟庆芬]
, 孟庆芬]
, 孟庆芬]
|
|

第一作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作。 E-mail: meimingxiang@263.net。
自从 Kalkowsky在 1908年构筑了叠层石的术语之后,叠层石一直是地质学家采用不同方法研究和思考的主题,而且一直被当作证明地球早期生命历史的代表物而得到深入调查。叠层石确实为地球早期生命历史提供了间接而且复杂的证据,所以,现代叠层石确实代表着明显的生物信号而成为研究的焦点。最为引人注目的是,现代叠层石的多样化构成,确实表明了蓝细菌生物席建造了叠层石,而且进一步表明了微生物席转化成叠层石不是一个直接的作用过程。那些反映现代叠层石多样化构成的典型实例包括:( 1)南极 Untersee地区的湖泊相锥状泥质叠层石;( 2)新西兰 North群岛被称为煎锅湖的热水湖泊中以及美国黄石国家公园热泉中的硅质叠层石;( 3)巴哈马台地、澳大利亚鲨鱼湾以及巴西东南部海湾碳酸盐沉积物构成的叠层石。由于蓝细菌微生物席是否代表了古代叠层石的形态学前体总是存在争议,而且在生命的图像中叠层石一直是一个迷惑的关键片段,因此,现代叠层石的多样化构成,将成为认识古代叠层石形成的关键和窗口。立足于前人的研究成果,追踪和总结现代叠层石的多样化构成,以及它们所代表的沉积作用和微生物新陈代谢活动丰富而复杂的信息,将不但丰富微生物沉积学的研究内容,还将拓宽沉积相分析的基本内容,对深入了解叠层石复杂的沉积学特征和生物学属性具有重要的科学意义。
About the first author Mei Mingxiang,born in 1965, graduated from China University of Geosciences in 1993,and obtained Ph.D. degree. Now he is a professor at School of Earth Sciences and Resource,China University of Geosciences(Beijing),and is engaged in sedimentology and stratigraphy.
Ever since Kalkowsky created a term of stromatolite in 1908,research on stromatolites has continuously been expanded from different methodological perspectives;a century later stromatolites continue to be actively investigated to document the early history of life on Earth. Stromatolites provide an indirect and complex evidence of early life on Earth,and today they represent a diversifying signature of life activities in which much of the modern research is focusing. More enchantingly,modern stromatolites that are marked by the composition diversity not only does demonstrate that cyanobacterial mats are constructing stromatolites but also reflect that the translation from cyanobacterial mats to stromatolites is not a direct process. These typical examples reflecting the composition diversity of constructing stromatolites are:(1)large conical stromatolites that are made up of muddy sediments in Lake Untersee,Antarctica;(2)siliceous stromatolites that are formed in thermal lake at Frying Pan Lake in North Island of New Zealand as well as at the Yellowstone National Park in USA;(3)carbonate stromatolites in Bahamas,Rio de Janeiro in Brazil and Shark Bay of Australia. Because of both that stromatolites continue to be a puzzling and crucial piece in the map of life and that whether cyanobacterial mats may be the potential morphological precursors for ancient stromatolites is a continue debating problem,the composition diversity of modern stromatolites becomes a key and window for the further understanding of the formation of ancient stromatolites. On basis of lots researching fruits by many scientists,tracing these fruitful and starting researches on the composition diversity of modern stromatolites that could delegate rich and sophisticate information of sedimentation and microbial metabolism are meaningful to further understanding the sedimentilogical feature and the biological nature of stromalites;moreover,this work can enrich the researching content of mat sedimentology and broaden the studying domain of facies analysis.
叠层石被广泛认为是纹层状的、早期石化的、自生成因的微生物构造, 发育在淡水、海水、热水和蒸发环境中, 生长在沉积物与水体的界面上(Awramik and Sprinkle, 1999; Riding, 2000, 2011)。除了广泛的环境分布以外, 叠层石还代表着横跨了35亿年历史的地质记录; 超过一个世纪的研究表明, 叠层石的多样化组构和复杂演变历史的许多细节问题, 还没有得到充分的了解(Grotzinger and Knoll 1999; Riding and Awramik 2000; Riding, 2000, 2011; 曹瑞骥和袁训来, 2006)。自从Kalkowsky(1908)构筑了叠层石的术语之后, 叠层石至少存在着2个范畴的定义: (1)非基因成因的定义, Semikhatov等(1979)将叠层石定义为“ 粘附式的、纹层状的、石化了的沉积生长构造, 这种构造从一个受到局限的初生面或一个生长点开始堆积” 。但是, 这个定义, 留下一个微生物是否参与了叠层石形成的问题, 尤其是那些大于32亿年的古老叠层石(Lowe, 1994; Grotzinger and Rothman, 1996; Pope et al., 2000; Awramik, 2006; Altermann, 2008; McLoughlin et al., 2008; Chacó nb et al., 2011)。(2)微生物调节作用产物的定义, 这是Walter(1976)所倡导的定义, 即“ 叠层石是通过沉积物捕获、粘结或沉淀作用所产生的有机沉积构造, 而且作为微生物新陈代谢和微生物生长的结果” 。
地质学家对现代叠层石研究的初衷和原动力, 是为了了解那些控制地球5/6的生命历史进程, 因为在现代的动物和植物世界, 完全为叠层石所主导的大陆和海洋景观是非常罕见的(Reid et al., 2011)。实际上, 叠层石2个范畴的定义, 就已经表明了蓝细菌是否是太古宙叠层石建造者的问题, 这个问题一直争论到今天; 但是, 今天建造叠层石的蓝细菌生物席, 对地球历史的地球生物学影响是独特的而且还是关键性的, 因为在前寒武纪的某个时间点上, 蓝细菌释放出有毒的、但是在现在却是维持生命所必需的氧气, 而且永久性地改变了地球生物进化的途径(Chacó nb et al., 2011)。总的来讲, 叠层石是微生物席的建造物已成为共识; 作为地球上最为古老的“ 宏观化石” (Chacó nb et al., 2011)和地球历史中第1个进化的“ 海相生物群” (Awramik and Sprinkle, 1999), 叠层石就成为了解过去复杂多变的碳酸盐岩世界的重要线索, 从而成为碳酸盐岩微生物席沉积学的主要研究对象(Riding, 2000, 2011; 曹瑞骥和袁训来, 2006; 梅冥相等, 2006, 2007; Altermann, 2008; Dupraz et al., 2009; Gerdes, 2010; 梅冥相, 2011, 2012, 2014)。所以, 叠层石一直是地质学家采用不同方法研究和思考的主题, 而且一直被当作地球早期生命历史的代表物而得到深入调查, 因为叠层石确实为地球早期生命历史提供了间接而且复杂的证据, 而且现代叠层石确实代表着明显的生物信号而成为研究的焦点。
在现代叠层石研究中, 以下实例成为叠层石多样化构成的典型代表:(1)南极湖泊相细粒碎屑沉积物组成的大型锥状叠层石, 与小尖塔状叠层石共生, 而且主要由丝状蓝细菌特殊的生物学行为所形成(Andersen et al., 2011), 为解释古代叠层石的形成提供了一个较为特别的类比物; (2)硅质叠层石, 以美国黄石国家公园热泉水池中硅质叠层石最为典型而且研究得较为深入彻底(Bosak et al., 2009, 2010; Petroff et al., 2010; Mata et al., 2012), 新西兰North群岛的Waimangu地热田被称为煎锅湖(Frying Pan lake)的热水湖湖泊中的大型硅质叠层石也特别令人耳目一新(Jones et al., 2005); (3)碳酸盐沉积物构成的叠层石, 以下3个地点最具有代表性而且得到了长期研究, 包括巴哈马台地的粗粒叠层石(Reid et al., 2011)、澳大利亚库隆潟湖超盐度环境中的细粒叠层石(Allen et al., 2009)、巴西东南部超盐度海岸潟湖中的细粒叠层石(Delfino et al., 2012), 这为研究与钙化作用相关的而且为复杂的微生物新陈代谢活动形成的更加普遍的碳酸盐叠层石的形成机理, 提供了丰富的资料和富有智慧的思考途径。
因此, 立足于前人的研究成果, 追踪和总结现代叠层石的多样化构成, 以及多样化过程所蕴含的沉积作用和微生物新陈代谢活动特征, 对深入了解叠层石复杂的沉积学特征和生物学属性具有重要的科学意义。
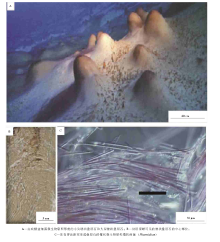
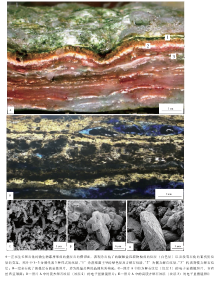
在南极东部, Untersee湖是一个面积最大(11.4, km2)和水深最深(> 160, m)的湖泊, 由于处在南纬74° 而成为一个长年被冰覆盖的湖泊, 而且是一个超级寡养的高碱性(pH=10.4)湖泊, 除了湖泊的西南角为一个小型的缺氧盆地以外, 湖泊水体富含可溶解氧气而且特别清澈透明(Andersen et al., 2011)。在湖泊底覆盖着光合作用微生物席, 这些微生物席至少可以延伸到100, m的深度, 而且为特别的丝状蓝细菌所形成, 它们构成2种特征性的叠层石( 图 1-A):(1)小尖塔状叠层石, 主要为微细鞘丝菌(Leptolyngby; 一种分叉的微细丝状蓝细菌)底栖微生物席所形成, 叠层石体的直径为10~15, mm, 高可以达到150, mm, 多呈束状发育在锥状叠层石之间, 这种形式的叠层石在南极其他地区的清水湖泊中较为普遍(Love et al., 1983; Hawes and Schwarz, 2001; Sutherland, 2009; Sutherland and Hawes, 2009); (2)锥状叠层石, 主要为蓆菌(Phormidium; 一种不分叉的丝状蓝细菌; 图1-C)底栖微生物席所形成, 这种类型的叠层石以前鲜有报道(Love et al., 1983; Hawes and Schwarz, 2001), 说明这样的叠层石可能是Untersee湖泊的独特特征, 锥状叠层石生长高出湖泊底大约50, cm, 它们的直径范围在10~60, cm之间; 形成这些锥状叠层石的纹层, 厚度为0.2~0.8, mm( 图 1-B), 与小尖塔状叠层石一样, 锥状叠层石主要为细粒黏土和有机质所构成的泥质叠层石为特征, 碳测年表明纹层可能发生在近10年的时间范围内(Andersen et al., 2011), 所以为一些描述得较好的太古宙叠层石(Altermann et al., 2006; Allwood et al., 2006, 2009; Awramik, 2006; Altermann, 2008)的生长提供了一个现代类比物。小尖塔体与锥状体的共生产出与特征性的蓝细菌构成的底栖微生物群落存在关联, 从而说明微生物行为支撑着叠层石的形态差异。
 | 图1 南极东部Untersee湖现代叠层石的总体特征Fig.1 Images showing the general features for modern stromatolites in Untersee Lake of Antarctica |
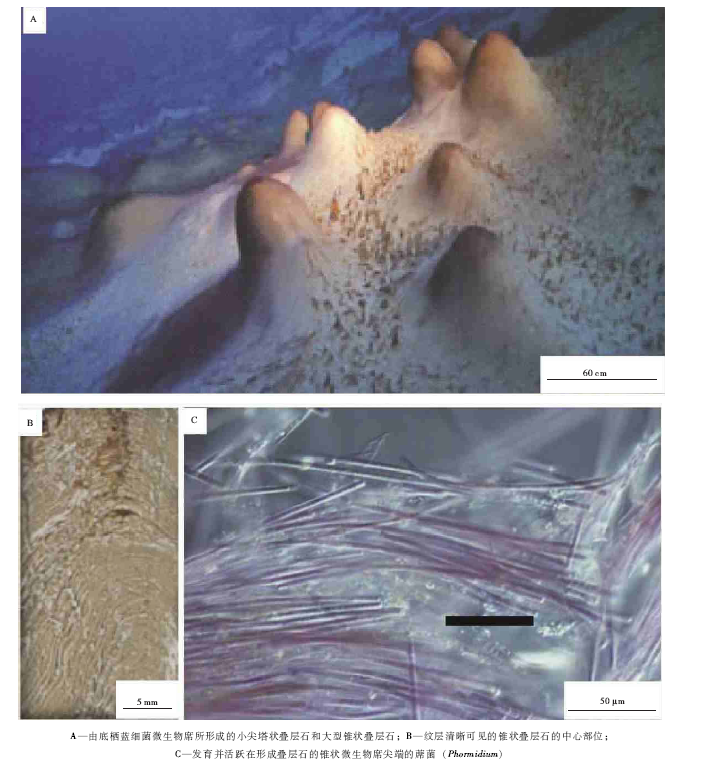
 | 图2 美国黄石国家公园现代硅质叠层石的总体特征Fig.2 Images showing the general features for modern siliceous stromatolites in Yellowstone National Park, USA |
由于缺乏矿物沉淀作用而保存潜力较低, 所以, Untersee湖的泥质叠层石还没有石化。Andersen等(2011)特别注意到, 在锥状叠层石之间, 甚至在一些锥状叠层石的侧部, 正在生长着小型小尖塔状体叠层石。小尖塔状叠层石与锥状叠层石在相同环境中的共生产出表明, 在对相同环境中不同的微生物群落正产生着不同的响应。组成这2种特征性叠层石的微生物群落存在着差异, 最为特征的是小尖塔状叠层石明显缺乏蓆菌(Phormidium)。物种构成的差异, 强烈地支持了一个解释:锥状叠层石和小尖塔状叠层石形成于不同微生物席类型中的优势物种的差异性行为, 虽然对这些叠层石的确切形成机理还不太清楚。例如, 蓆菌(Phormidium)可能更加喜欢与地面平行排列而不是垂直于地面, 它将产生一个细胞外聚合物质(EPS)层, 在这个EPS层中, 细胞在退却, 从而促使细胞沿着地面平行排列, 而且这个行为将会阻止细胞与光响应的垂向移动, 从而会促进平行移动。相反, 微细鞘丝菌(Leptolyngby)是在小尖塔状叠层石占据着优势的生物, 则可能喜欢垂直定向, 从而可以直接向上进入水柱而生长。
一个接近于连续的、穿越周围环境光照度较高的光的使用效率, 成为被冰覆盖的光合作用微生物席的一个普遍特征(Hawes and Schwarz, 2001; Schwarz et al., 2005; Vopel and Hawes, 2006); 然而生物的生产量, 在锥状体的不同部分没有产生明显变化, 但是, 在锥状叠层石顶部较高表面积的生物量, 说明了在锥状叠层石顶部单位面积的潜在性生长, 总体上要高于侧部和平坦的底部地带。
在Untersee湖的湖底, 沉积物主要是由细粒黏土矿物所组成(大小为1, μ m左右), 具有少量粒径可达50, μ m的长石块状晶体, 因此, 湖泊底的沉积物明显不同于那些较粗粒的粉砂和砂, 这些较为粗粒的沉积物可能是通过风成过程搬运并沉积在湖泊表面冰上, 说明了“ 冰川面粉(glacial flour; Chanudet and Filella, 2008)” 的脉动式注入并分布在湖泊内的基本特点。如果斜长石的风化过程实际上就是产生碱度的主要过程的话, 少量的“ 冰川面粉” 的幕式注入, 以及随后的微生物群落的恢复, 是叠层石纹层( 图 1-B)最有可能形成的基本机制。
Andersen等(2011)还特别注意到, 富含斜长石的“ 冰川面粉” 的偶尔注入, 将是保持湖泊水体高碱度的前提; 在这样的水体环境中, 硅质碎屑矿物的优势性表明, 矿物沉淀作用实质上是缺乏的, 相反, 叠层石的生长是通过黏土级别的沉积物的捕获与粘结过程, 从而形成特征性的泥质叠层石。Andersen等(2011)还进一步认为, 锥状叠层石体的生长, 要求叠层石顶部的生物量和矿物堆积作用要大于锥状体之间的平坦地带, 但是, “ 冰川面粉” 的沉落被认为是均匀的, 或者优先堆积在地势较低处, 这不会导致尖端部位的优先生长。因此, 叠层石锥状形态的生长, 要求微生物群落在其尖端部位要很快增加其生物量, 或者是通过丝状体朝向锥状体的脊部的移动而产生优先生长, 从而最终成型叠层石(Batchelor et al., 2005); 再者, 锥状体和小尖塔状叠层石的共生( 图1-A), 加上矿物沉淀作用的缺乏, 强烈地表明, 生物学行为是造成Untersee湖中的泥质叠层石形态学特征的主要起因, 这个认识进一步表明, 微生物行为与群落之间的联系是非常复杂的, 这要比光梯度变化之类的环境线索的单一响应(Batchelor et al., 2005)复杂得多。Andersen等(2011)还强调到, 细胞与细胞间的信号传输, 可能是叠层石形态学特征较为重要的生物作用因素, 例如在微生物景观中的繁殖作用和建造作用的定额传感过程(Battin et al., 2007; Decho et al., 2010), 但是, 这方面的研究还有待今后进一步调查。所以说, 对Untersee湖中的微生物群落及其环境响应的估计, 为微生物群落对叠层石形态成型过程所起的特殊作用提供了一个新约束, 从而对一个老问题产生了一个新认识, 即对形成古代叠层石的微生物活动的解释, 还存在着许多有待于深入研究的细节问题。
上文已经提及到, 硅质叠层石, 以美国黄石国家公园热泉最为典型而且研究得较为深入彻底(Bosak et al., 2009, 2010; Petroff et al., 2010; Mata et al., 2012), 而且新西兰North群岛的Waimangu地热田被称为煎锅湖(Frying Pan Lake)的热水湖湖泊中的大型硅质叠层石也特别令人耳目一新(Jones et al., 2005); 下文以Berelson等(2011)的研究成果为主介绍黄石国家公园的硅质叠层石, 以Jones等(2005)的研究成果为主介绍新西兰煎锅湖的硅质叠层石。
自从Weed(1889)首次在黄石国家公园的热水水池中发现并研究叠层石以来, 该地区的热水水池中的叠层石成为地质微生物学研究的代表, 近年来尤其引人注目的研究是, 从锥状叠层石的形态学属性去探索所蕴含的光合作用信号, 并有效地延伸到古代叠层石的研究之中, 产生了许多创新性认识和新概念, 如光合作用气泡及其产生的特殊的窗格组构所产生的锥状叠层石的成型过程(Bosak et al., 2009, 2010; Mata et al., 2012)以及锥状叠层石形成被解释为对营养物扩散作用的响应(Petroff et al., 2010)等等。黄石国家公园与热泉相关的热水水池, 形成了各种各样的微环境, 这些微环境的温度变化较大, 在水下2280, m处的热泉口最高温度可以达到94 ℃, 而且pH从1变化到10, 水体中富含二氧化硅, 形成了一个较为严酷的环境, 也成为各种各样的微生物群落繁盛的各种微环境, 最终发育了丰富多样的硅质叠层石。
Berelson等(2011)对黄石国家公园的现代硅质叠层石富有成果的研究, 是对有名的Obsidian Pool热泉所形成的热水水池特别的微生物生态系(Spear et al., 2005; Shock et al., 2005)中生长的叠层石( 图 2-A, 2-B)所进行的研究, 该热水水池的面积大致为600, m2, pH为5.7, 是一个弱酸性的富含二氧化硅的热水水池。两种特征性的微细纹层样式( 图 2-C)被识别出, 最普遍的纹层样式组成了叠层石的大部分(叠层石主体), 然而不太普遍的特征性纹层覆盖在主体上, 构成叠层石悬挂体。
叠层石主体由明显的亮层和暗层组成( 图 2-C, 2-D), 纹层的厚度为100~200, μ m, 为含量大于95%的隐晶质二氧化硅(蛋白石)构成; 总体上, 亮层的平均厚度为150, μ m, 而暗层的平均厚度接近100, μ m。暗层为致密填积的、定向性大致平行于纹层的丝状蓝细菌(蓆菌(Phormidium))所组成, 亮层主要为球形蓝细菌和硅藻所构成, 另外见到稀疏填集的、定向性几乎直交于纹层的丝状蓝细菌(也以蓆菌为主)所构成( 图 2-D); 更为特征的是, 亮层具有大型的洞穴( 图 2-D), 表明了微生物席内气泡的存在, 这些气泡是富含氧气的, 曾经解释和定义为光合作用气泡(Bosak et al., 2009, 2010), 而且这样的气泡在古代叠层石中形成的特殊的窗格组构还被描述为沙滤器构造(Mata et al., 2012)。很明显, 叠层石的主体是向上生长的, 而且主体的侧部和顶部则被一种致密的特征不明显的纹层所覆盖, 最终, 建造作用过程又回到主体的纹层样式, 而且这些悬挂体则代表了一个沉积作用层或叠层石内的一个相( 图 2-C)。悬挂体或覆盖层包含着少量硅化的丝状蓝细菌, 以及在主体之中没有发现的球状蓝细菌和羽状的硅藻( 图 2-E), 还见到一些分叉的飞氏菌(Fischeralla); 再者, 与主体中的亮层和暗层的对偶层形式不一样, 悬挂体存在较少的内部构造, 而且填集得较为紧密。
Berelson等(2011)还进一步阐释到, 美国黄石国家公园现代硅质叠层石纹层的2种样式, 代表了2种时间范围的驱动力:“ 叠层石主体” 的微细纹层代表了昼夜式的或接近于昼夜式的生长, 而“ 主体” 和“ 悬挂体” 组构的交互作用则代表了发生在年度范围中的热泉水池高度或体积的波动过程。这些创新性的认识, 更加证实了叠层石可能是微生物群落结构和环境驱动力的一个有价值的记录, 而且可以类比于太古宙的微细纹层叠层石(Altermann et al., 2006; Altermann et al., 2006; Awramik, 2006; Altermann, 2008; Allwood et al., 2006, 2009), 为研究古代叠层石的形成提供了许多有价值的思考途径和研究线索。
在新西兰北部, 沿着Okataina火山中心的南部边缘发育一个被称为Waimangu-Rotomahana的热水体系, 这个火山中心位于第四纪流纹岩地层和热水喷发角砾构成的崎岖不平的地域之中(Seward and Sheppard, 1986); 该地区具有一个较长的火山喷发活动历史, 包括1886年的Mt Tarawera火山喷发, 这些火山活动产生了现在的热水体系。
煎锅湖(Frying Pan Lake), 占据着Echo火山口的一部分, 这个火山口形成在1886年的Tarawera火山喷发过程之中(Houghton and Scott, 2002)。煎锅湖地表面积大约为38000, m2(Scott, 1994), 湖泊底较为平坦, 水体深度平均为6, m, 在热泉和气体从湖泊底的沉积物中的喷气孔排泄的地方, 局部的水体深度大于 20, m(Keam, 1981; Houghton and Scott, 2002); 湖泊的水体为硫酸盐型, pH为5.4~5.8(Glover et al., 1994)。在2003年2月份以及2004年3月份, 在出水水道南侧, 生长着叠层石的表面水体具有的温度为47~51, ℃, pH值为5.5~5.8; 在煎锅湖的滨线周围的几个地点以及从泄水渠出口向下游的200, m内的地带, 发现了正在生长的百合大浮叶状叠层石( 图 3-A; Jones et al., 2005)。
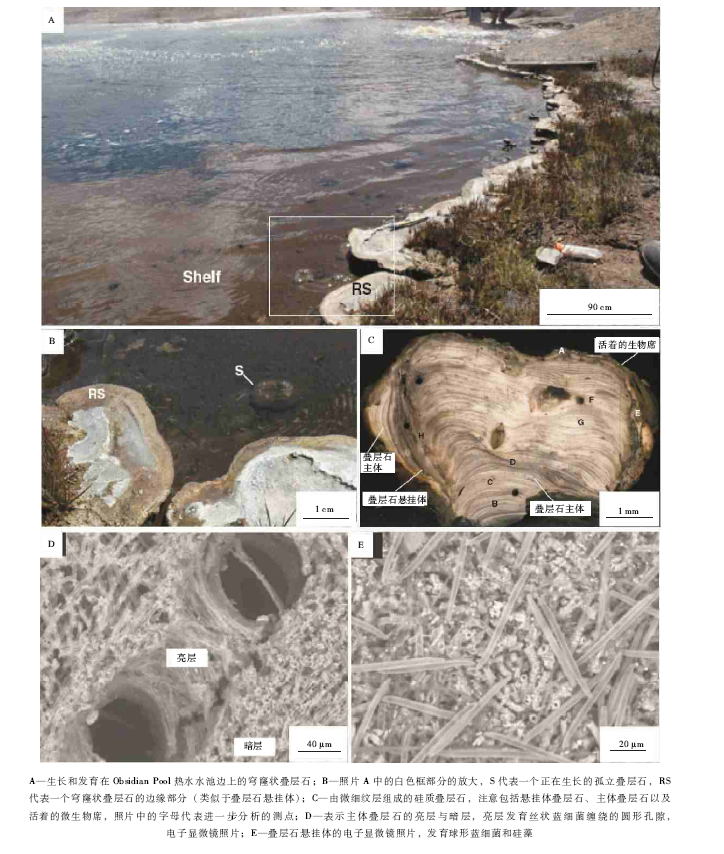
 | 图3 煎锅湖硅质叠层石的总体特征(修改自Jones et al., 2005)Fig.3 Images showing general features for modern siliceous stromatolites in Frying Pan Lake(modified from Jones et al., 2005) |
靠近出水道底面大致1, m深的地方, 被暗绿色的微生物席所覆盖( 图 3-A), 各种形态和大小的柱状体以不同的高度突出在出水道的底面(图3-B), 沿着出水道的边缘和轴向部分; 一旦叠层石柱状体的顶面向上生长到水面, 微生物席就会在水面向外延伸生长而产生一个圆形或卵圆形的百合大浮叶状( 图 3-C)。百合大浮叶状叠层石的上部表面, 具有一个突出的边缘(3~5, mm高), 支撑着与周围水体接触的部分, 也是暗绿色的, 在潮湿的时期, 覆盖着百合大浮叶状叠层石表面内部的平坦部分的就具有一个被浅绿色带所包围的桔红色内部带( 图 3-C); 在进一步干化之后, 内部分带现象就随之消失。随着水面下降, 气泡将穿过微生物席而上升, 并且在暴露的微生物席上产生小型的成串小气泡( 图 3-C), 这些气泡应该是蓝细菌光合作用的结果(Bosak et al., 2009, 2010; Mata et al., 2012)。
进一步观察表明, 煎锅湖硅质叠层石( 图 3, 图4), 主要为不分叉的丝状蓆菌(Phormidium)所形成, 其次是分叉的丝状蓝细菌飞氏菌(Fischerella), 少量生物构成包括过渡性而且有隔膜的丝状微生物、直径较小的丝状微生物、有孔的或没有孔的孢子以及硅藻(Jones et al., 2000, 2001, 2002, 2005), 这些生物形成纹层明显而且包含着生长孔隙的硅质叠层石( 图 3-D); 叠层石内部的生长孔隙, 实际上就是 图3-C所示的大量气泡的石化了的残余物, 从而代表了蓝细菌光合作用形成的气泡(Bosak et al., 2009, 2010; Mata et al., 2012)。
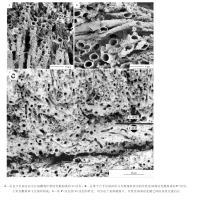
煎锅湖硅质叠层石的纹层主要为2种样式( 图 3-E):(1)“ P-纹层” , 以致密交织的、平躺着的丝状蓝细菌(主要为蓆菌)硅化鞘为特征( 图 4-B); (2)“ U-纹层” 要比P-纹层具有一个更加多样的生物群, 以直立的丝状蓝细菌为特征, 主要为蓆菌所构成, 包括一些飞氏菌以及其他难以识别的微生物( 图 4-A)。第3种纹层样式被定义为“ M-纹层” (Jones et al., 2000, 2001, 2002, 2005), 以细胞外黏液、黄铁矿草莓状体和硅藻为特征, 而且以不太明显的纹层或透镜层的形式局部覆盖在U-纹层之上。单个蓆菌丝状体定向性从横向到纵向的转变, 标定了从P-纹层到U-纹层的转变( 图 3-E, 图4-C)。
很清楚, 煎锅湖的百合大浮叶状叠层石( 图 3, 图4)是微生物成因的, 具有几乎全部为光合作用蓝细菌丝状体组成的构造格架, 这些丝状体形成了二氧化硅沉淀作用模板。叠层石的生长受到局部性的水体和气候条件之间综合作用的复杂变化所控制。实际上, 叠层石内部的纹层化作用, 常常归因于依赖于时间的这些环境条件的旋回性变化。来自于煎锅湖出水道中“ 百合大浮叶状” 叠层石的信息表明, 这些内部纹层的结构与环境条件存在关联, 而且P-纹层、U-纹层、M-纹层构成的旋回, 必然与一些旋回性变化的环境因素的变化相关联, 但是, 很难将这些纹层与特定的环境参数联系起来(Jones et al., 2000, 2001, 2002, 2005)。形成柱状体和百合大浮叶状叠层石的微生物席, 正在活跃地硅化, 这些叠层石形成在一个酸性(pH为5.6~5.8)而且较浅的热水水体的滨岸平台之中, 沿着出水道的边缘发育。从较厚的绿色底栖微生物席开始的柱状叠层石体的向上生长, 一直生长到空气与水的界面上, 而且通过超过垂向生长的横向快速生长而形成一种平坦状的百合大浮叶状的衬板状; 变大了的衬板状叠层石体将遮蔽着下伏的柱状体, 从而限制了光合作用而降低了柱状体的横向扩张( 图 3)。形成表面衬板体的微生物群落, 要比形成柱状体的微生物群落更加多样化, 而且具有俯卧式和直立纹层的交互过程( 图 3-E, 图4-C), 还可能包含有富集硅藻分泌的细胞外黏液的纹层。特定的叠层石宏观样式, 以及特定的微生物构成, 为解释古代叠层石提供了许多有价值的线索。
 | 图4 煎锅湖硅质叠层石纹层的微生物构成(修改自Jones et al., 2005)Fig.4 Images showing microbial composition for laminae of modern siliceous stromatolites in Frying Pan Lake(modified from Jones et al., 2005) |
碳酸盐沉积物构成的现代叠层石, 以下列实例为典型代表:(1)巴哈马台地的粗粒叠层石(Reid et al., 2011); (2)澳大利亚鲨鱼湾的细粒叠层石(Allen et al., 2009); (3)巴西东南部超盐度海岸潟湖中的细粒叠层石(Delfino et al., 2012)。这些海相碳酸盐叠层石复杂的微生物席群落及其微生物新陈代谢活动、以及特别且复杂的钙化作用机制, 代表着能够揭示古代叠层石形成的沉积作用和微生物新陈代谢过程的许多有用的信息, 所以成为识别古代叠层石的关键和窗口, 而且为深入理解前寒武纪微生物碳酸盐岩工厂和微生物席沉积学提供了很多重要的思考途径和研究线索(梅冥相, 2011, 2012, 2014)。
形成在正常海条件下的叠层石, 首先发现在巴哈马的Schooner沙州和Lee Stocking 群岛(Dravis, 1983; Dill et al., 1986), 这些叠层石生长在搬运着砂的潮汐水道中, 研究发现叠层石的原始增生作用是通过砂颗粒的粘结与捕获作用完成。相似的叠层石, 后来在巴哈马的其他地点的进一步发现, 引起了Reid等学者的持续性研究(Reid et al., 1999, 2000, 2011)并取得了丰富的成果, 而且涉及到有机质的分解和生产作用、微生物席的石化作用、叠层石的沉积序列、微生物群落的季节性分布以及这些作用过程之间复杂的相互作用的综合研究(Stolz et al., 2009)。
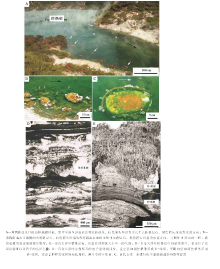
巴哈马Little Darby群岛的叠层石, 生长在潮下带水深为1~2, m的鲕粒沙州中( 图 5-A), 形成高出海底30~50, cm的穹窿状叠层石, 这些叠层石常常融合成生物礁体( 图 5-B), 在一些叠层石穹隆体的侧部生长着伞藻(Acetabularia; 图5-C)。叠层石的增生作用发生在柔软的微生物席中, 微生物席沿着叠层石构造向上生长的面分布, 在叠层石的增生作用过程中, 2个最基本的构成单元是:(1)裂须菌(Schizothrix)微生物席的生长和发育; (2)由水流和波浪提供的碳酸盐砂颗粒的注入。风和波浪的动荡程度旋回性, 是向叠层石供应的砂颗粒的脉动性行为的特征, 记录在毫米级别的细粒纹层中( 图 5-D, 图6-A)。可以滑动的裂须菌香毛簇(图 6), 是主要的生产者, 正在活跃生长的裂须菌微生物席总是保留在叠层石的顶面( 图 5-D, 图6-A), 而且随着叠层石的增生作用向上移动, 从而形成穹窿状叠层石。
 | 图5 巴哈马Little Darby沙州碳酸盐叠层石的总体特征(修改自Reid et al., 2011)Fig.5 Images showing general features of modern carbonate stromatolites in Little Darby Cay of Bahamas(modified from Reid et al., 2011) |
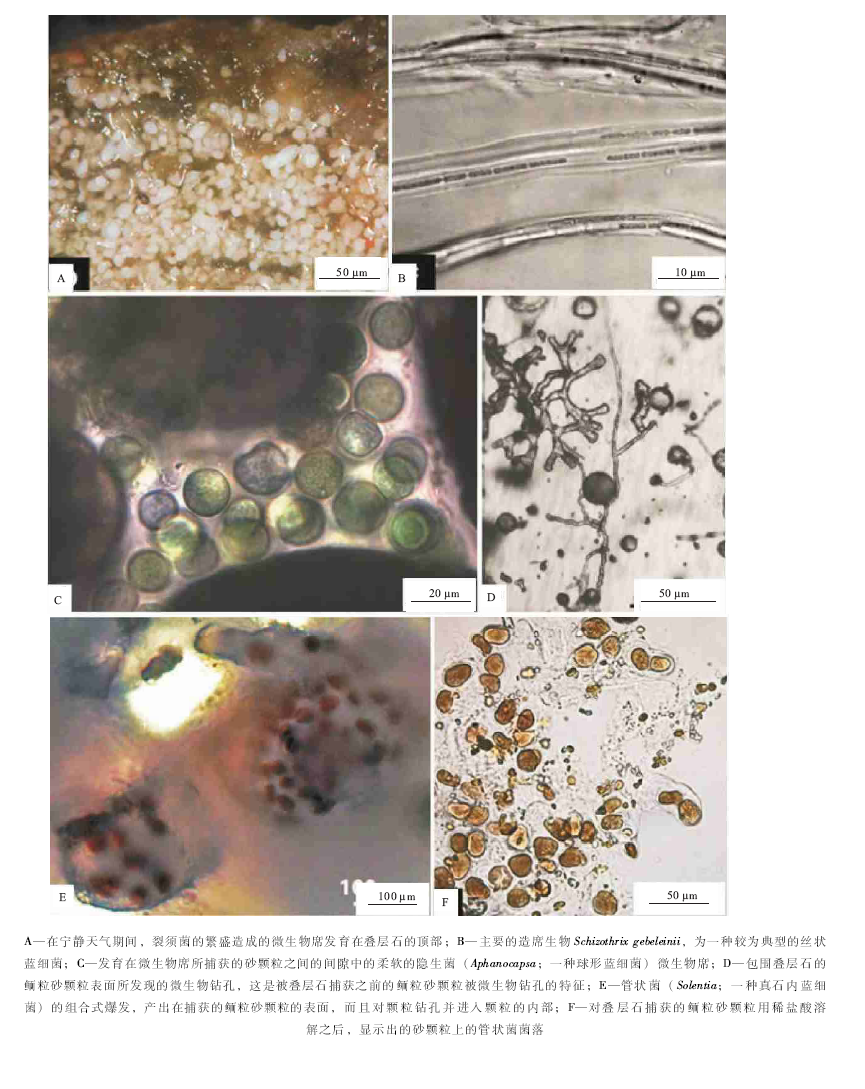 | 图6 巴哈马Little Darby沙州碳酸盐叠层石的微观特征(修改自Reid et al., 2011)Fig.6 Images showing microscopic features of modern carbonate stromatolites in Little Darby Cay of Bahamas(modified from Reid et al., 2011) |
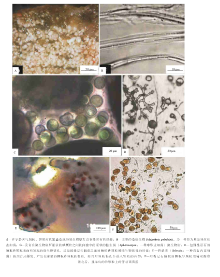
巴哈马Little Darby群岛的叠层石, 主要的造席生物是Schizothrix gebeleinii( 图 6-B), 这种蓝细菌具有高度移动性而滑动的香毛簇, 呈束状形式被包裹在细胞外聚合物的鞘中(Golubic and Browne, 1996)。次要的组成单元是那些快速滑动的生物, 属于蓝细菌属的裂须菌(Schizothrix)、颤菌(Oscillatoria)和螺旋菌(Spirulina), 以及少量的厌氧细菌如硫酸盐还原细菌等等, 这个组合的生物信号, 已经在Highborne沙洲的叠层石中被注意到(Foster and Green, 2010)。在波浪动荡作用期间, 一些颗粒粘附在微生物席上, 逐渐埋藏了蓝细菌。蓝细菌通过正向的趋光性对埋藏作用响应, 滑动移出其鞘, 而且继续在构造的顶面保持着最大限度的生物量。这些年轻的丝状体包含着单个的香毛簇, 随后通过破碎作用复制而且彼此相邻滑动, 最终形成鞘中的束状物( 图 6-B)。
那些“ 成熟的叠层石纹层” , 即形成在平静阶段的蓝细菌多重香毛簇的丝状体, 也就是说, 形成在没有砂的融合作用的蓝细菌生物量堆积和聚集作用时期( 图 6-A)。主要的原始生产物的贡献者是裂须菌(Schizothri)的细胞外聚合物(EPS)构成的鞘, 同时经历了有机营养细菌的降解作用, 就像Highborne的叠层石那样(Decho et al., 2005), 还涉及到硫酸盐还原反应所代表的厌氧呼吸作用(Baumgartner et al., 2006)。另外一个主要的生物量生产作用, 发生在被捕获颗粒间而且处在遮蔽状态的间隙式空间中, 这样的间隙式空间有利于产生那些嵌入在柔软的EPS中的球状蓝细菌菌落, 如隐生菌(Aphanocapsa; 一种球形蓝细菌)的生长和沉落(图 6-C)。
在巴哈马Little Darby群岛的叠层石中, 第2个组成单元, 为那些具有自己的生物活动历史的鲕粒砂颗粒所组成, 这些被形成叠层石的微生物席所捕获的鲕粒然后又变得稳定。在融合进入微生物席之前, 鲕粒砂颗粒就预先被钻孔改造过, 只有少数正在被钻孔的颗粒由活着的真石内菌居住( 图 6-D)。真石内菌的生态条件, 经历了一个主要变化, 这个变化发生在颗粒被捕获和被稳定在叠层石构造之中的一个复杂过程之后, 从而导致了管状菌(Solentia)的组合式爆发(Wild et al., 2006; Tribollet et al., 2010)( 图 6-E), 管状菌产出在被微生物席捕获的砂颗粒的表面, 而且钻孔进入颗粒的内部( 图 6-F); 最为重要的是, 管状菌的石内繁盛与生物席的石化作用过程同步(Reid et al., 1999, 2000, 2011)。Little Darby的另外一个基本特征是, 一些叠层石对沉积物供应的局部遮蔽, 将会产生藻类过度生长的较长的停滞作用期( 图5-C), 然而叠层石的埋藏可能主要发生在主要的风暴作用期。
今天, 形成在超盐度海水环境中的叠层石的最好实例是澳大利亚西部的鲨鱼湾中的Hamelin Pool, 这些叠层石最早为Logan在1954年首先进行系统研究(Logan, 1961); 根据表面形态特征, 曾经将形成叠层石的微生物席大致划分为9种不同的形态, 即胶粒状、凝胶状、光滑形态、针垫状、毛簇状、脓疱状、薄膜状、网状和小气泡状微生物席(Playford, 1990)。不同形态学特征的微生物席, 为不同的微生物群落所形成; 但是, 鲨鱼湾Hamelin Pool的叠层石, 主要是微生物对沉积物的捕获与粘结过程的结果(Reid et al., 2003):(1)微晶碳酸盐(泥晶)的微生物沉淀作用, 就像在这些叠层石中的骨架和胶结物一样, 是叠层石形成作用的一个基本过程; (2)微生物捕获与粘结过程, 是潮间带叠层石增生作用的主要机制, 形成细粒颗粒状的和砂屑的结构; (3)微生物沉淀作用则是潮下带的主要机制, 形成了细粒的泥晶叠层石; (4)在鲨鱼湾叠层石中, 微生物沉淀的碳酸盐泥晶较细, 就像很多古代的泥晶质叠层石那样。因此, 在一个共同环境中, 丰富的叠层石和微生物席的排列和发育, 为研究不同类型的微生物席构成对叠层石成型和石化作用的生物与物理参数, 提供了一个独特的机会。
就像早期的研究所表明的那样, 从光滑的微生物席到脓疱状的微生物席, 可能发生在柱状叠层石的生长过程期间(Allen et al., 2009), 而且这2种类型的微生物席主导着Hamelin Pool潮间带的叠层石(图 7-A), 有时候发现它们还覆盖在潮间叠层石的表面( 图 7-B)。脓疱状微生物席( 图 7-C)具有一个暗色色素的表面层, 在表面层之下没有见到清楚的纹层, 虽然可以见到绿色的(蓝细菌的)、粉色和黑色的(细菌的)着色斑状物, 从而表明微生物的空间分布确实发生在这种生物席的中极不规则的窗格组构内。相反, 光滑的微生物席, 表面颜色较浅, 具有较低的地势起伏( 图 7-D)。
 | 图7 鲨鱼湾Hamelin Pool碳酸盐叠层石的总体特征(修改自Allen et al., 2009)Fig.7 Images showing general features of modern carbonate stromatolites in intertidal region of Hamelin Pool, Shark Bay(modified from Allen et al., 2009) |
研究发现, 色球菌(Chroococcus; 图7-E)是脓疱状微生物席中独特的蓝细菌, 而微鞘菌(Microcoleus; 图7-F)和鞘丝菌(Lyngbya; 图7-G)则是光滑微生物席中独特的蓝细菌; 2种微生物席共同具有的蓝细菌大致为16种, 如拟色球菌(Chroococcidiopsis; 图7-H)和螺旋菌(Spirulina; 图7-I)等等(Allen et al., 2009)。更为重要的是, 一个多样化的新陈代谢活动可能出现在脓疱状和光滑的微生物席之中, 较为普遍的新陈代谢活动被认为出现在Hamelin Pool的微生物席之中:(1)蓝细菌的生氧光合作用( 图 7-C和7-D中的绿色层所代表); (2)不生氧光合作用(绿屈挠菌(Chloroflexi)、一些蓝细菌、阿尔发原细菌(Alphaproteobacteria)和其他微生物所造成; (3)硫化物氧化作用(伽马原细菌Thialkalivibrio和其他生物所造成, 图7-C和7-D中的粉色层所代表); (4)硫酸盐还原反应( 图 7-C和7-D中的黑色层所代表); (5)氮素代谢活动(一些肌动蛋白细菌如Frankia sp., 蓝细菌如微鞘菌和其他生物所造成)。
细菌种群、微生物以及矿物形成过程以及它们的产物, 在巴西东南部Lagoa Vermelha的超盐度海岸潟湖中得到了系统全面的研究, 而且微生物席及其之下的沉积物, 还被作为一个定义高镁方解石和白云石形成的边界条件的独特体系而加以研究(Vasconcelos et al., 2006)。现在出现于Lagoa Vermelha的化学、矿物学和生物条件, 可能在前寒武纪较普遍, 而且在Lagoa Vermelha的高盐度微生物席中, 完成了与碳酸盐矿物形成相关联的微生物新陈代谢过程, 可能已经出现在前寒武纪; 因此, 作为一种生物矿物的微生物碳酸盐岩, 可能成为地质历史时期新陈代谢的记录。
Lagoa Vermelha的微生物席, 平均厚度为3, cm, 而且由交互的碳酸盐纹层和未石化的有机质纹层所组成; 自生碳酸盐沉淀作用区域, 被嵌入在细胞外聚合物质(EPS)以及微生物之中。在微生物席内的Ca-Mg碳酸盐岩的连续纹层, 类似于叠层石状的构造。没有识别到明显的由微生物席造成的颗粒捕获和粘结过程, 而且没有石化的有机质纹层反而显示出一个微生物群落的层理现象, 这些现象已经通过微生物带染色所表现出的成层过程而观察到( 图 8-A)。这些叠层石的显微组构多为球粒和泥晶团块(lumps), 具有镁方解石的优势性以及白云石的证据, 也包含有其他类型的矿物, 如石膏和石盐(Delfino et al., 2012; 图8-B)。
微生物席显示出2层生氧光合作用进化蓝细菌, 表面层(0~2, mm)由那些带有嗜盐的蓝细菌如黏球菌(Gloeocapsa)、螺旋菌(Spirulina)、微鞘菌(Microcoleus)所居住, 直接下伏于其下的纹层(2~4, mm)主要含有丝状蓝细菌如螺旋菌和微鞘菌。在蓝细菌层之下5~12, mm, 已经检测到大量的光合硫化物氧化细菌(Vasconcelos et al., 2006)。进一步的研究已经揭示出, 球状或球形蓝细菌, 例如隐生菌(Aphanothece)和色球菌(Chroococcus), 在超盐度环境的微生物席中是主要的构成生物, 这些生物对高盐度已经高度适应; 裂须菌(Schizothrix)和微鞘菌(Microcoleus)是相对较为大型的丝状蓝细菌, 在数量上相对较少, 但是却占据着较大的生物体积(Delfino et al., 2012)。
Lagoa Vermelha微生物席包含了分散的方解石和镁— 方解石层, 生氧光合作用和厌氧呼吸作用产生了微生物席顶层的方解石沉淀作用, 而且硫化物氧化作用与硫酸盐还原作用则产生一个特殊的成岩作用过程, 这个过程将促进高镁方解石和钙白云石的沉淀作用( 图 8-A中纹层1, 图8-C)。尤其特别的是, 图8-A中所示的纹层2和纹层3的镁方解石沉淀物, 表现为特别的圆形和卵圆形的晶体形式, 确实不发育斜方六面体的晶体形式( 图 8-D, 8-E)。矿物颗粒的电子能谱分析(EDS)表明(Vasconcelos et al., 2006), 镁的摩尔浓度可以达到12%, 显示出高镁方解石的存在。
 | 图8 巴西Lagoa Vermelha现代碳酸盐叠层石的总体特征Fig.8 Images showing general features of modern carbonate stromatolites in Lagoa Vermelha, Brazil |
在Lagoa Vermelha, 生物地球化学条件表明, 碳酸盐形成作用2种不同的机制发生在沉积物不同的深度。硫酸盐还原细菌活动以及碱度在叠层石表面纹层之下的缺氧部分, 正在控制着白云石沉淀作用以及从无序到化学计量白云石的“ 老化过程” (Vasconcelos and McKenzie, 1997; van Lith et al., 2002), 然而, 硫化物氧化作用以及由此造成的pH值降低, 则通过发生在含氧— 缺氧界面的镁方解石的沉淀作用而促进白云石形成(Moreira et al., 2004)。总之, 微生物新陈代谢过程及其所伴随的碱度变化, 主要造成了Lagoa Vermelha微生物席内的碳酸盐沉淀作用, 并形成特别的还可能包含着原生白云石的细粒叠层石。进一步研究表明, 如 图8-A所示的叠层石纹层, 表明了4种类型的生物在活动(Warthmann et al., 2011):(1)产生氧气的蓝细菌; (2)消耗氧气的异养细菌; (3)厌氧硫化物— 氧化作用的紫色硫细菌; (4)在较深沉积物中的产生硫化物的硫酸盐和硫的还原细菌。更为重要的是, 紫色硫细菌的原始的新陈代谢活动是通过硫化氢的氧化作用来吸收二氧化碳, 甚至在含氧光合作用进化之前, 厌氧的硫细菌可能曾经对生物调节的碳酸盐岩的形成具有重要贡献, 这种生物调节的碳酸盐岩将产生化石微生物席, 最终形成普遍的碳酸盐纹层化作用和有名的叠层石。
已经被普遍接受的观点是, 在现代微生物席中蓝细菌是主要的建造者(Awramik and Sprinkle, 1999; Gerdes et al., 2000; Jing et al., 2005; Roeselers et al., 2006; 梅冥相等, 2006, 2007; Lau et al., 2009; Gerdes, 2010; 梅冥相, 2011, 2014), 这是一个正统的方法学所推崇的思考途径范例, 也是蓝细菌形态类型所造成的一个范例(Gerdes et al., 2000; Westall, 2005; Gerdes, 2010; Chacó nb, 2011); 要强调的是, 绿色和紫色硫细菌的微生物群落, 包括化能有机异养微生物、发酵微生物、化能有机自养微生物铁还原和硫酸盐还原细菌, 均是现代微生物席重要的功能性和结构性构成(Baumgartner et al., 2006; Chacó nb, 2011)。这些特殊的蓝细菌、硫酸盐还原细菌、硫化物氧化细菌等表征的微生物席的功能性和结构性构成, 在形成现代碳酸盐叠层石的微生物席群落结构中得到了最为明显的体现(图 5至图8)。
南极冰湖中的蓝细菌微生物席及其所形成的泥质叠层石(图 1; Andersen et al., 2011), 黄石国家公园(Bosak et al., 2009, 2010; Petroff et al., 2010; Berelson et al., 2011; Mata et al., 2012)以及新西兰热水湖泊(Jones et al., 2005)中的蓝细菌形成的硅质叠层石, 不但表现出蓝细菌强烈的生态忍耐性, 以及在微生物席的发育之中起的关键性作用, 而且也展示出现代叠层石的多样化构成, 尽管对一些细节问题还有待于进一步研究, 如南极冰湖中的泥质叠层石的生长机理、以及热水酸性湖泊中蓝细菌鞘及基质的硅化作用机制等等。虽然在元古宙和太古宙的碳酸盐叠层石中识别出了蓝细菌化石(Altermann et al., 2006; Altermann, 2008), 但是, 还是难以准确回答蓝细菌生物席就是古代叠层石的前体(Chacó nb, 2011); 这方面的研究, 运用徐桂荣等(2005)的“ 协同进化” 理念, 去理解蓝细菌的形态和生态革新对形成叠层石的影响(Blank and Sá nchez-Baracaldo, 2010), 就显得更加重要。
冯增昭(2013)主编的《中国沉积学》(第二版)囊括了中国沉积学家的智慧和新成果, 为今天的沉积学研究, 提供了重要的理论基础和学科生长点。徐桂荣等(2005)《生物与环境的协同演化》、殷鸿福等(2004)《生物地质学》和谢树成等(2011)《地球生物学》一起, 为沉积学家在地球生物学框架下开展沉积学研究, 提供了一些重要的思考途径。曹瑞骥和袁训来(2006)的《叠层石》, 成为古生物学家对叠层石类型高度总结的典型代表。这些汇聚了许多学者的智慧, 而且具有鲜明的时代特色的宏篇巨著, 为沉积学家研究叠层石这一个地球早期的“ 宏观化石” , 提供了许多重要的思考途径和线索。
还应该强调的是, 随着“ 蓝绿藻” 最终被修订为“ 蓝细菌” , 而且“ 藻席” 被进一步修订为“ 微生物席” (Krumbein, 1979; Rippka et al., 1979; Brock et al., 1994), 多年来将“ 藻” 简单地代指“ 微生物” 而产生的一些不规范的沉积学术语, 如“ 藻灰结核” 、“ 藻屑” 、“ 藻鲕” 、“ 藻叠层石” 等等, 应该予以抛弃, 因为“ 地质学专业术语是地质学定义明确的专业用语, 它反映了地质学基本特征及其释义” (杨建超等, 2007); 现代叠层石的多样化构成, 也进一步说明了形成叠层石的微生物席, 具有复杂的微生物群落和复杂的新陈代谢活动, 并非只是藻类生物的生命活动结果。
责任编辑 庞凌云 李新坡)
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|