{kind=link}
地球历史中的巨型氧化作用事件:了解古地理背景演变的重要线索*
[梅冥相 ]
]
]
|
|

作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作。 E-mail: meimingxiang@263.net。
在塑造我们的星球环境的过程之中,分子氧起着关键的作用。大气圈和海洋中氧气的出现及其浓度随着时间的变化,与地球上的主要变化存在强烈关联,诸如构造重组、气候波动和生物进化。针对地球大气圈氧气含量的上升,多年研究的结果肯定了 2个基本事实:( 1)地球最早期的大气圈是缺乏氧气的;( 2)今天的大气圈则为 21%的氧气所组成。由于地质历史时期大气圈氧气水平的大多数地质标志,只是意味着存在与缺乏,这就为确定大气圈氧气含量上升的时间进程带来很多困难。即使如此,一系列地质证据已经表明,一个从缺氧的到含氧的大气圈的转变,大致发生在 2.5—— 2.0, Ga,这个转变被定义为巨型氧化作用事件( GOE)。近年来的深入研究发现,几个主要证据表明,在前寒武纪——寒武纪过渡时期的大约 850—— 540, Ma,发生了"第二次巨型氧化作用事件( GOE-Ⅱ)",还被进一步定义为新元古代巨型氧化作用事件( NOE)。再者,大气圈氧气水平在显生宙还存在着一个特别的上升,这次变化在石炭纪晚期接近一个峰值为 150% PAL(现代大气圈氧气含量水平),所以,也可以定义为一次巨型氧化作用事件,即显生宙的巨型氧化作用事件( POE)。因为蓝细菌光合作用造成的氧气生产,曾经导致了大气圈与海洋的氧化作用,反过来为需氧呼吸作用和大型而且复杂的、最终富有智慧的生物进化,提供了基本条件;因此,大气圈氧气上升,是与地球动力学过程紧密相关的地球生物学过程的作用结果,从而成为了解漫长的地质历史时期古地理背景演变的重要线索。从古元古代的 GOE,经过新元古代的 NOE,到显生宙的 POE,这些巨型氧化作用事件的内在特征、作用结果与基本属性,尽管存在着较大的差异,但是,从这些概念的出现到对它们的形成机理的探索性研究,涌现出了许多新概念和新认识;追索这些新概念和新认识,将为了解地球大气圈氧气上升的复杂历史所代表的一个特别的地球上生物学过程,提供一些有益的重要线索和思考途径。
Molecular oxygen has played a crucial role in shaping our planetary environment. The emergence of oxygen in the atmosphere and the oceans and its changing concentration through time is strongly linked to major changes on Earth such as tectonic reorganization,climatic perturbations and biological evolution. Two facts are known with certainty that are resulted from the long-term study on the rise of atmosphere oxygen,that are:(1)Earth’s earliest atmosphere was essentially devoid of oxygen,and(2)today’s atmosphere is composed of 21% oxygen. It should be emphasized that most geological indicators of atmospheric oxygen levels in geological time imply only presence or absence,which led to many difficulties on study of the rise of atmosphere oxygen. Even though,a battery of geological indicators suggest that a shift from an anoxic to an oxic atmosphere some time between 2.5 and 2.0 billion years ago,which is known and defined as the Great Oxidation Event(GOE). Further research in resent years demonstrates that several lines of evidence point to a second‘Great Oxidation Event’(GOE~Ⅱ)during the Precambrian-Cambrian transition between about 850 and 540,Ma,which is herewith referred to as the Neoproterozoic Oxygenation Event(NOE). In practice,the considerable variation in atmospheric oxygen levels through Phanerozoic time peaked near 150% PAL in the late Carboniferous,which is herewith defined as the Phanerozoic Oxygenation Event(POE). Because photosynthetic oxygen production by cyanobacteria led to oxygenation of the atmosphere and oceans,in turn provided an elementary condition for the aerobic respiration and the evolution of large,complex and ultimately intelligent organisms,the rise of atmosphere oxygen can be defined as the action product of geobiological process that is genetically related to the geodynamics,which become an important clue for the further understanding of the evolution of palaeogeographical background of the Earth. Furthermore,there are many intrinsically differences with respect to their consequences and merits among these Oxygenation Events, i.e. the GOE,the NOE and the POE,but the concept of themselves and the study on their forming mechanism demonstrate many new ideas and cognitions. Tracing these new ideas and cognitions will be helpful for the further understanding of this particular geobiological process represented by the rise of atmosphere oxygen,which can provide many useful thinking approaches and researching clues for the future research.
一个全新的化学探索惊人地表明, 地球大气圈中最为丰富的2种气体, 是氮气和氧气。大气圈中氮气的含量, 自从地球形成以来一直保持恒定, 在大气圈中的代谢转换时间, 被估计为大约1000, Ma左右; 再者, 作为一种惰性或非反应性气体, 氮气在大气圈中的丰度变化, 不受生物过程驱使(Berner et al., 2003; Berner, 2006)。
与氮气相反, 氧气, 作为地球大气圈中第2丰富的气体, 则是一种高度反应的气体, 表现出以下基本的作用特征:(1)如果不存在一个连续性来源, 大气圈中的氧气会变得快速亏损(Keeling et al., 1993); (2)由于常常与地幔中的还原剂进行化学反应, 氧气存在于极为不平衡的热力学状态; (3)在太阳系内, 地球上二价气态的氧气, 所具有的较高浓度是非常独特的, 而且在太阳系内的大约20秒差距(parsec)内的其他行星上, 还没有发现过(Kasting, 1993); (4)在宇宙中, 氧是第3丰富的元素(排在氢和氦之后; Anders and Grevesse, 1989), 按照重量计算, 氧元素还是地球的地壳中最丰富的元素(Barrow and Tipler, 1986)。宇宙中的氧元素(O), 通过所谓的"主线"核反应序列所产生, 与其他元素结合在一起, 被输送到早期的地球上; 在早期地球的增生作用与分异作用过程中, 通过加热和变冷的序列式循环过程, 氧与硅、碳反应并结合, 形成2种主要的阴离子, 这样的阴离子与金属阳离子作用在一起, 构成了地壳和地幔的基本物质, 而且氧(O)与氢(H)反应形成水(Holland, 1984); 另外的水, 可能是通过陨星和彗星输送到行星地球表面。不管其来源如何, 同位素数据表明, 在地球增生作用之后约200, Ma, 地球表面就含有液态水(Falkowski and Godfrey, 2008; Falkowski and Isozaki, 2008), 代表了水圈的早期形成, 为生命的起源提供了基本的地质背景。
虽然水可以通过紫外光的作用产生, 但是, 这个反应只能够产生极少量的氧气, 这是因为强烈的负反馈效应所致(Falkowski and Isozaki, 2008; Falkowski and Godfrey, 2008)。地球上压倒优势的氧气来源, 还是水的光合生物氧化作用(photobiological oxidation), 也就是著名的生氧光合作用; 在这个过程中, 光养生物, 以光作为能量来源, 运用二氧化碳和水合成碳氢化合物并释放氧气, 而且将氧气作为副产品, 但是, 究竟什么过程产生了生氧光合作用, 这种能量转换无疑是自然界中最为复杂的过程(Olson and Blankenship, 2004; Nisbet et al., 2007)。在年到千年的时间尺度内, 生氧光合作用与呼吸作用紧密耦合, 结果, 纯粹的氧气产生实质上近于零; 也就是说, 如果没有岩石中的有机质的埋藏, 在大气圈中就只有少量的自由分子氧气。所以说, 氧气的光合作用生产是一个复杂的地球生物学过程(殷鸿福等, 2004; 徐桂荣等, 2005; 谢树成等, 2011; 梅冥相, 2012, 2014), 而且地球大气圈氧气含量水平的上升, 则是一个与地球动力学过程存在紧密关联的地球生物学过程(Falkowski and Isozaki, 2008; Falkowski and Godfrey, 2008; Condie et al., 2009; 梅冥相, 2010; Eriksson et al., 2010; Grotzinger et al., 2011; 梅冥相和高金汉, 2015; 梅冥相和孟庆芬, 2015, 2016)。
就像Kump(2008)指出的那样, 大多数古代大气圈氧气水平的地质标志, 只是意味着存在与缺乏, 因为一直到今天, 科学界还面对着2个基本的问题:(1)在生氧光合作用期间造成水分解的过程和机制, 还不清楚; (2)不知道究竟是什么机制控制了地球大气圈气体的浓度(Falkowski and Isozaki, 2008)。围绕着古代大气圈氧气含量水平的估计, 虽然还存在很大的不确定性, 但是, 关于大气圈氧气上升的时间进程和粗略样式, 已经取得了一些重要的概念和认识:(1)最早期的地球大气圈肯定是缺氧的, 其氧气含量水平肯定要小于现代大气圈的水平(PAL)的0.1%, 而且可能更低(小于10-5 PAL; Pavlov and Kasting, 2002), 而且支持这个结论的证据, 没有排除在微生物席中或地球表面水体中的太古宙小型氧气绿洲(梅冥相等, 2007; Hoashi et al., 2009; 梅冥相, 2011a, 2011b, 2012, 2014; Crowe et al., 2013; Olson et al., 2013; Riding et al., 2014)存在的可能性; (2)在2.5——2.0, Ga, 大气圈氧气含量水平快速上升, 实质上可能达到了百分之几的PAL, 由于明显的上升速度和奇特的属性, 这个时间段的大气圈氧气含量的上升被定义为巨型氧化作用事件(GOE; Kirschvink et al., 2000; Canfield, 2005); (3)大致在800——600, Ma期间, 大气圈氧气水平发生了第2次增加(Canfield and Teske, 1996), 伴随着这次事件的是深部大洋的氧化作用和多细胞动物的出现, 这就是新元古代巨型氧化作用事件(NOE; Canfield, 2005; Campbell and Allen, 2008; Holland, 2009; Campbell and Squire, 2010; Kump et al., 2011; Grotzinger et al., 2011; Shields-Zhou and Och, 2011; Tzipermana et al., 2011; Och and Shields ̄Zhou, 2012; Sperlinga et al., 2013); (4)大致在500, Ma左右, 氧气可能接近现代水平(21%), 而且自从500, Ma以来波动在15%~35%之间(Berner et al., 2003; Berner, 2006); (5)大气圈氧气水平在整个显生宙的变化, 所具有的最为特征的变化是在石炭纪晚期接近一个峰值为150% PAL(大气圈氧气含量达到了31%), 这就是文中所称的显生宙大气圈氧化作用事件(POE)。这些初步的结论, 是科学家们从零零星星的线索进行综合分析的结果, 就像冯增昭教授(2004)所倡导的"单因素分析多因素综合"的方法体系来进行区域古地理重建一样, 充满着科学家们的无穷智慧, 从而成为了解地质历史中古地理背景演变的重要线索。
在生命历史中, 针对所有的生物化学创造来讲, 利用阳光氧化水的机制所表征的生氧光合作用, 无疑是最伟大的创造, 因为早期的蓝细菌, 不但使用水作为燃料而且还具有一个几乎没有限制的能量(太阳光)供应优势; 再者, 在生氧光合作用期间, 作为废料的氧气生产作用, 确实明显改变了地球海洋、大陆和大气圈构成(Sessions et al., 2009)。大气圈氧气含量的上升, 又反过来促进生物圈的进化与革新; 例如, 大致在GOE时期, 发生了从原核生物到真核生物的进化; 在NOE期间, 发生了从单细胞到复杂结构的多细胞生物的进化(Och and Shields ̄Zhou, 2012)。
从古元古代的GOE, 经过新元古代的NOE, 到显生宙的POE, 这些概念的出现到对它们的形成机理的探索性研究, 展示出了许多新概念和新认识。这些巨型氧化作用事件的内在特征、作用结果与基本属性存在着较大的差异, 这正是追逐这些新概念和新认识的强大驱动力, 从而成为文中的主要内容。
运用各种同位素体系对陨石进行测年, 包括对那些短周期的以及现在已经灭绝的放射性核素的测年, 融合成大型的气体行星的物质来源于早期被太阳风吹到外部星云的那些物质, 包括现在已经亏损的挥发性元素组成的物质, 因此, 地球的形成时间, 就被推断为T0=4567, Ma(Amelin et al., 2009; Nyquist et al., 2009; Bouvier and Wadhwa, 2010)。
冥古宙(4567——4030, Ma), 代表了初生地球时期(Van Kranendonk et al., 2012), 这个时期的地球动力学过程, 存在着以下重要变化:(1)在4460——4450, Ma, 地球已经有了现在的大小, 地核已经分异出, 而且地球已经获得了大气圈(Allè gre et al., 1995); (2)发生在4500, Ma形成月亮的巨星撞击作用事件之后, 大致在4400——4300, Ma, 地球的岩浆海洋已经完全分异(Blichert ̄Toft and Arndt, 1999); (3)岩浆海洋的冷却过程, 可能会导致一个不稳定的地幔, 这样的地幔会发生翻转而最终产生一个巨大的铁镁质地壳(Kramers, 2007); (4)在4000, Ma左右, 地幔已经获得了现行的氧化——还原状态, 这是由于He2+损耗到太空而且与氧化剂相混合的结果, 这些氧化剂包括三价铁(Fe3+)、水和碳酸盐(Catling et al., 2001; Delano, 2001)。相应地, 发生在冥古宙重要的地球生物学过程包括:(1)生命起源, 大致发生在4100, Ma, 而且生命起源最有可能发生的环境, 是那些发育在超镁铁质海洋地壳上的碱性而且低温的热水体系(Holm et al., 2006; Russell and Hall, 2006; Martin and Russell, 2007; Holm and Neubeck, 2009), 最为重要的是, 超镁铁质海洋地壳的蛇纹石化作用, 可以作为能量来源来促进生命的起源和发育(Russell et al., 2010); (2)产甲烷生物, 发生在4100——3800, Ma之间(Holm et al., 2006; Russell and Hall, 2006)。
太古宙, 是地壳与生物圈得到确立的地质历史时期(4030——2420, Ma; Van Kranendonk et al., 2012), 占据了幼年期地球(Juvenile Earth; 4030——2780, Ma)以及青年期地球(Adolescent Earth; 2780——1780, Ma)的大部分时期, 经历了一个特别的地球动力学过程和地球生物学过程。太古宙的地球动力学过程, 曾经被Van Kranendonk 等(2012)总结为以下几个重要阶段:(1)地球早期的原始地壳的形成(4030——3530, Ma); (2)稳定克拉通代表的岩石圈形成(3490——2820, Ma); (3)太古宙晚期地壳的快速生长(2780——2630, Ma); (4)太古宙末期地壳生长的衰减(2630——2420, Ma)。伴随者上述地球动力学过程, 太古宙时期发生了以下阶段性的地球生物学过程:(1)太古宙的地球大气圈是贫乏氧气的, 在生物圈的氧化作用和生物地球化学循环中, 蓝细菌起着一个基本而且重要的作用(Blank and Så Nchez-Baracaldo, 2010), 氧气在大气圈中的聚集作用, 大致开始于3000——2500, Ma 的生氧光合作用起源与进化之后(Payne et al., 2011), 而且发生在2900, Ma地球上最古老的冰川作用(Young et al., 1998; Nisbet et al., 2007; Williams, 2008)所代表的大气圈变冷过程, 正是地球大气圈起源与进化之后早期生命作用过程的重要表现; (2)在太古宙晚期地壳快速生长的时期(2780——2630, Ma), 所伴随的是生氧光合作用的发展, 相对于不生氧光合作用的原始海洋生产率来说, 生物生产率增加了1至2个数量级(Canfield et al., 2005; Rosing et al., 2006), 异化的铁还原作用(DIR)是光合作用多样化的结果, 发现在细菌与古菌中, 包括超嗜热菌、硫酸盐还原细菌、硝酸盐还原细菌以及产甲烷细菌, 涉及到各种各样的电子授体(Lovley, 2004), 而且被认为是地球上最早的微生物新陈代谢活动(Lovley, 2004), 这些地球生物学过程, 成为在太古宙晚期微生物大爆发的重要标志; (3)伴随者太古宙末期(2630——2420, Ma)地壳生长的衰减, 则是微生物生命在这个作用时期的繁盛和进一步扩张, 微生物明显开始泵吸出的氧气, 而且会充满海洋中的还原剂储库, 尤其是可溶解的还原性铁, 从而在很多大陆上沉淀了大量的条带状铁建造(BIF)(Catling et al., 2001), 实际上, 氧气的第1个化学痕迹, 就在这个时期被识别出, 表明了一个朝向氧化大气圈的转变作用的开始(Siebert et al., 2005; Anbar et al., 2007; Wille et al., 2007)。
今天, 科学家们为我们提供了一个早期生命起源和进化的图像(Van Kranendonk et al., 2012):(1)生命起源发生在4100, Ma之前; (2)产甲烷生物作用发生在4100——3800, Ma之间; (3)厌氧的产甲烷生物, 发生在3100, Ma之后; (4)光养生物发生在3200, Ma之前; (5)需氧的产甲烷生物发生在2800——2500, Ma之间。但是, 这个生命起源与进化的图像, 还存在着许多细节问题需要进一步调查和研究, 例如3460, Ma的氧化海洋中形成的原生赤铁矿(Hoashi et al., 2009)、3430, Ma的锥状叠层石生物礁(Allwood et al., 2006; Awramik, 2006), 以及由这些记录代表的"太古宙氧气绿洲"(Riding et al., 2014; 梅冥相和高金汉, 2015; 梅冥相和孟庆芬, 2015, 2016), 说明生氧光合作用这样复杂的新陈代谢作用已经发生在3500, Ma之前或者更早(Altermann, 2008)。生氧光合作用究竟是什么时候进化的?这个问题意义重大, 这是因为蓝细菌造成的光合作用氧气生产曾经导致了大气圈与海洋的氧化作用, 反过来为需氧呼吸作用和大型而且复杂的、最终富有智慧的生命进化提供了基本条件(Catling et al., 2005); 但是, 光合作用起源的证据, 即这个基本的生物学过程是如何开始而且如何发展的问题, 可能消失在时间的长河之中(Bekker et al., 2004; Olson and Blankenship, 2004; Nisbet et al., 2007; Buick, 2008; Payne et al., 2011; Schopf, 2011; Olson et al., 2013; 梅冥相和高金汉, 2015; 梅冥相和孟庆芬, 2015, 2016)。
针对太古宙早期几乎接近缺氧的大气圈条件, 尽管存在很多标志, 但是, 最为可能而且普遍存在的代表物, 是形成在与大气圈直接接触的岩石中的地球化学代表物(Buick, 2008)。古土壤, 成为大气圈雨沉降物造成的基岩滴定作用的记录, 潜在性地成为最好的代表物, 但是又很少保存, 因为古土壤常常形成在海平面之上的陆生序列中, 因而容易受到侵蚀作用破坏。只有一个较为肯定的太古宙古土壤实例的报道, 形成在澳大利亚西北部2.76, Ga的 Mt Roe玄武岩上(Rye and Holland, 1998)。在这个古土壤剖面中, 以下特征表明了太古宙大气圈的实质性缺氧状况:(1)氧化——还原敏感性较高的金属, 尤其是还原性的铁构成的菱铁矿和黄铁矿, 发育在被较为强烈的风化过程所淋滤过的剖面上部, 进一步说明了缺氧的条件; (2)铈, 保存为更加还原的状态, 也进一步表明了一个贫乏氧气的条件。早期的研究也表明, 在与大气圈气体混合得较好的浑浊河水所沉积的碎屑矿物沉积物中, 氧化——还原敏感的碎屑矿物, 包括黄铁矿、沥青铀矿和菱铁矿(Rasmussen and Buick, 1999), 将太古宙大气圈的含氧量水平约束在0.1 PAL, 即0.01~0.001 PAL的水平上。最后, 独立于质量的硫同位素S33和S36的分馏作用, 一种记录在太古宙地表海水中沉淀的硫矿物的普遍特征, 强烈地说明了火山成因的含硫气体进入一个缺乏氧气的而且为紫外线再次作用的大气圈中(Farquhar et al., 2000), 这些独立于质量的硫同位素信号, 难以发育在存在氧气分压条件, 表明大气圈的氧气含量水平大致为10-5 PAL(Pavlov and Kasting, 2002)。
还需要强调的是, 在GOE之前, 即使在缺氧的大气圈之下, 分子氧的实质性的量, 可能局部聚集在浅水环境中, 因为这样的环境中有利于蓝细菌的生产作用, 并且将有效地孤立起来以至于氧气没有完全被马上消除掉, 从而形成"太古宙氧气绿洲(Archean oxygen oases)"(Riding et al., 2014)。GOE之前数亿年的大气圈中可能发生过的瞬时性的氧气"阵发性氧气"(Anbar et al., 2007; Kaufman et al., 2008)、2.8, Ga的太古宙海相灰岩(Riding et al., 2014)、3.46, Ga的氧化海洋中形成的原生赤铁矿(Hoashi et al., 2009)、3.43, Ga的叠层石生物礁(Allwood et al., 2006; Awramik, 2006), 说明了产生氧气的生氧光合作用在地球上的出现, 明显要早于GOE出现的时代(梅冥相和高金汉, 2015; 梅冥相和孟庆芬, 2015, 2016)。最为有意义的是, 在南非大致2.9, Ga的Pongola超群中, 还包含着地球上最为古老的冰川作用证据, 表明了在那个时候可能存在着局部寒冷的大气圈条件(Von Brunn and Gold, 1993; Young et al., 1998), 而且甚至是一个弱氧化的大气圈(Ono et al., 2006), 尽管对这个时候的较小的δ 33S异常还存在另外的解释(Farquhar et al., 2007), 因为大陆冰川作用可能要求一个大致在20, ℃或者更低的平均地表温度(Kasting, 1987)。
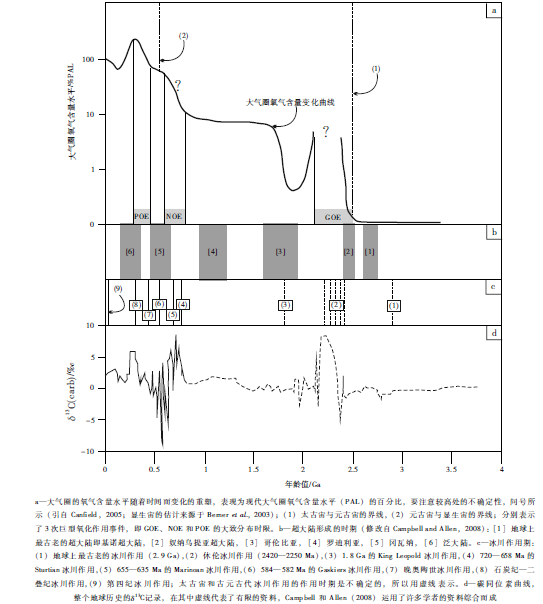
大气圈氧气的第1次明显的聚集和出现, 即巨型氧化作用事件(GOE; 图1; Holland, 2002), 大致发生在2.4——2.0, Ga(10×108年), 这是一个大地构造、气候与生物聚变的时期(Kirschvink et al., 2000; Barley et al., 2005); 一系列地质证据已经表明, GOE代表了一个从缺氧到含氧大气圈的巨大转变(Kirschvink et al., 2000; Canfield, 2005)。
 | 图1 地质历史时期的主要的地质过程Fig.1 Diagram showing the main geological processes |
今天的主流观点是, 早期的地球大气圈本质上缺氧的, 而且由紫外线对水的光解作用所产生的氧气, 大致只相当于10-14 PAL(现代大气圈水平)(Catling and Zahnle, 2002); 在2.32, Ga左右, 而且在一个地质范畴内属于短时间阶段内, 转变成一个氧化的大气圈(Bekker et al., 2004; Buick, 2008)。
地质学家们数十年的研究积累, 总结出了5个证据的线索, 支持了大气圈氧气含量水平在这个时期明显的增加(Campbell and Allen, 2008; Holland, 2009; Campbell and Squire, 2010):(1)氧化-还原敏感的矿物沥青铀矿、黄铁矿和菱铁矿, 在太古宙沉积物中较为普遍, 但是在年轻于2.3, Ga的河流沉积物中较为罕见(Holland, 2002); (2)在老于2.2——2.3, Ga的古土壤中, 表现出铁的移动性证据, 说明Fe(Ⅲ )不是太古宙土壤明显的构成组分(Rye and Holland, 1998); (3)大致出现在2.3, Ga的红层表明, 在这个时候的土壤中, Fe(Ⅲ )已经变成重要的组分(Campbell and Allen, 2008; Holland, 2009); (4)生物诱发的独立于质量的34S的分馏作用在2.4——2.3, Ga的突然增加(Cameron, 1982), 对这个变化最有可能的解释是海洋中的硫酸盐浓度的增加, 归因于在2.4, Ga之后一个更加氧化的大气圈中黄铁矿风化作用强度的增加(Canfield et al., 2000); (5)在2.45, Ga左右, 33S独立于质量的分馏作用的突然减弱, 因为33S的分馏作用多为大气圈二氧化硫的紫外辐射光解作用所产生, 要求紫外射线深深穿过一个不含臭氧的而且接近于缺氧的大气圈(Farquar and Wing, 2003)。
关于发生在这个地质历史时期的大气圈氧气上升的幅度和范围, 已经取得了以下重要的信息和认识:(1)大致在2.5, Ga, 大气圈的氧气含量逐渐穿越了一个可以氧化黄铁矿的较低的水平阀值(10-8 至 10-5 PAL; Kump, 2008); (2)在2.4——2.3, Ga期间, 达到了现代大气圈含量水平(PAL)的10~5以上(Kirschvink et al., 2000; Young, 2002; Canfield, 2005), 甚至达到了百分之几的PAL(Sessions et al., 2009); (3)在2.1——2.0, Ga期间, 还可能达到1% PAL(Kump, 2008; Kump et al., 2011)。总的来讲, 在大致2.4, Ga, 处在大约10-5, PAL的水平, 则经历了独立于质量的硫同位素分馏作用(S-MIF)的损耗, 然后是一个穿越古元古代早期日益增长的大气圈氧气上升过程; 在2.0, Ga, 最终达到一个1% PAL的水平(Kump, 2008; Kump et al., 2011)。如 图1所示, 尽管取得了上述重要的认识, 但是, 在GOE期间, 大气圈氧气含量水平上升的最高值, 还是难以准确标定而需要进一步的研究(Och and ShieldsZhou, 2012)。
在GOE之前的太古宙晚期(2.78——2.63, Ga), 地球动力学过程与地球生物学过程的综合作用, 是广泛的地幔熔融过程和地壳形成作用所开启的, 这些作用还产生了一个或多个的超大陆(Superia与Sclavia; 或Kenorland; Aspler and Chiarenzelli, 1998; Eriksson et al., 2010; 梅冥相, 2010)。这些变化与下列变化相呼应, 即在大约2.7, Ga大气圈开始聚集氧气, 而且在2.5——2.0, Ga累积成GOE(Kirschvink et al., 2000; Holland, 2002; Farquhar et al., 2000; Bekker et al., 2004; Canfield, 2005), 并且在大约1.78, Ga之后进入一个环境稳定的时期(Van Kranendonk et al., 2012)。
在GOE之前的太古宙末期(2.42——2.25, Ga), 可能是在4.0, Ga之后的地质记录中, 具有代表性的时间段, 被解释为全球性岩浆活动关闭了的时期(Condie et al., 2009), 虽然在这个时期地幔活动的完全关闭是不可能的(Berman et al., 2010; French and Heaman, 2010), 但是, 全球性地质活动在太古宙晚期(2.78——2.63, Ga)的地幔——地壳喷裂之后, 明显减弱了。岩浆活动减速的一个可能的原因(O’ Neill et al., 2007; Ernst, 2009), 是地幔的明显变冷过程(Davies, 1995)。伴随着这个构造活动明显受到限制的时期, 是全球大气圈的变冷过程以及同时发生的大气圈氧气含量水平的上升, 从而产生了广泛的冰川沉积(Kirschvink et al., 2000; Canfield, 2005)。
大气圈的变冷过程本身, 可能对地幔产生了一个反馈效应, 进一步减慢了地幔对流并限制了板块构造活动和这个时期的地壳生长(Lenardic et al., 2008; Landuyt and Bercovici, 2009)。地幔温度的减小, 会导致大陆岩石圈的增厚和变硬过程, 当与海洋岩石圈变冷和减薄过程耦合的时候, 就会导致大陆的暴露过程, 并产生在这个时代之前的岩浆活动与热水体系变化之间平衡状态的变化, 以及在此之后, 岩石暴露的增加与岩石风化过程之间的水文循环平衡状态的变化(Flament et al., 2008)。因此, 最有可能的是, 随着太古宙晚期(2.78——2.63, Ga)的地壳大量形成, 以及太古宙末期(2.42——2.25, Ga)的地幔变冷过程所耦合的大气圈变冷过程, 为微生物的繁盛和扩张提供了一个基本的地质背景, 尤其是生产氧气的蓝细菌的繁盛和扩张, 从而激发了古元古代的GOE( 图 1)的产生。
GOE的起因, 还在强烈争论着, 所提出的假说包括:(1)地幔气体氧化——还原状态的变化(Kump and Barley, 2007), 陆上火山活动及其更加具有氧化性的气体(就是二氧化硫)的增加会导致大气圈的氧气上升, 这反映了大陆的成熟作用, 而且这个认识被Holland(2009)所推崇, 他认为大气圈氧化作用状态的逐渐增强产生于火山气体中的CO2/H2O 以及 SO2/H2O比率值的增加; (2)海水营养物供应(Campbell and Allen, 2008), 认为超大陆的发育将向海洋提供营养物(铁和磷), 为蓝细菌的繁盛提供了一个媒介, 从而造成光合作用氧气的明显上升(Lindsay and Brasier, 2002; Squire et al., 2006), 因此, 就像Melezhik(2006)所认识到的那样, 古元古代大气圈的氧气上升, 与同时代冰川作用即休伦冰期存在着重叠式发育的特点; (3)氧气的还原性储库的减小(Bjerrum and Canfield, 2002), 大气圈氧气含量水平在大约2.4, Ga的上升, 是由于还原剂(还原性的硫和铁)储库在海洋中被填满的结果, 这与随着一个变冷的地球体系所造成的较弱的火山活动所导致的进入大气圈的还原性火山气体的体积减小作用在一起(Barley et al., 2005); (4)2个反馈式的稳定状态的转变(Goldblatt et al., 2006), 即在早期的低氧的稳定状态与一个较为年轻的高氧气的稳定状态, 随着对流层的紫外线屏蔽作用产生的变化而发生变化; (5)大气圈甲烷水平的减小(Zahnle et al., 2006; Konhauser et al., 2009), 也就是说, 大气圈的氧气含量水平的上升, 是甲烷温室效应大气圈崩溃的结果, Konhauser 等(2009)进一步认为, 海洋中镍的亏损是变弱的科马提岩火山活动和地幔变冷的结果, 从而造成产甲烷微生物的窒息, 以及在氧气上升之前或者在同时代大气圈甲烷的含量的降低; (6)生氧光合作用本身的进化(Kopp et al., 2005), 具体讲, 在2.4, Ga之前, 蓝细菌局限在淡水环境, 之后, 进入海水环境并多样化(Blank and Så nchez-Baracaldo, 2010), 最终导致大气圈氧气含量的上升, 因为在一个相对较为稳定的背景中有利于蓝细菌的繁盛和扩张, 因此, 就像Melezhik(2006)所认识到的那样, 针对古元古代氧气上升与冰川作用存在着重叠式的起因。另外, δ 56Fe值在元古宙早期的变化, 也支持了GOE代表的大气圈氧气含量的上升, 表现在:(1)在2.5——2.3, Ga期间, 异化的铁还原作用(DIR)受到压制(Johnson et al., 2008a, 2008b), 因为细菌硫酸盐还原反应(BSR)活动的加强, 将会造成海水硫化物含量的增加, 硫化物含量的增加会滴定铁元素, 造成支撑DIR的铁的可用性降低; (2)在大致2.4, Ga之后, δ 34S值的增加, 记录了海水硫酸盐含量的增加; (3)在大致2.3, Ga, 即大气圈氧气含量水平的初始上升之后, 海水的δ 56Fe值演变成接近于零的状况, 这正好是处在一个含氧气的大气圈之下的特征性标志(Rouxel et al., 2005; Anbar et al., 2007)。这些假说本身, 实际上就充分表明了地球大气圈氧气上升是一个与地球动力学过程密切相关的地球生物学过程。
2250——2060, Ma, 是以再次复兴的全球性岩浆活动为特征的时期, 但是与其太古宙晚期的全球性岩浆活动期还不一样, 伴随着这个事件所发生的是碳酸盐岩δ 13C构成的极端高的正异常以及明显的碳同位素异常波动, 这就是著名的Lomagundi-Jatuli碳同位素正异常事件(LJE)(图 1-d; Melezhik et al., 2005, 2007)。其基本事实是这些代表着全球性碳酸盐岩中碳循环的一个较大的碳循环波动(Karhu and Holland, 1996), 而且是在地球历史进程中任何已知的沉积碳酸盐岩中最大幅度的δ 13C正异常(图1; Melezhik et al., 2007), LJE似乎是古元古代其他的突出事件序列中的一个整体, 它们之间的关系是非常复杂的而且只是得到部分揭示(Melezhik et al., 2005)。更为有趣的是, 这个时期大气圈氧气含量水平的逐渐增加, 也正是第1次出现真核生物化石的时间(2100, Ma; Han et al., 1992; Albani et al., 2010), 还可能第1次代表了在大气圈氧气含量水平上升所直接激发的生命复杂性得到加强的进化事件。
伴随着大气圈氧气含量水平的上升, 元古宙早期(或太古宙晚期)真核生物起源, 确实是一次重要的地球生物学事件。一个引人注目的认识是, 真核生物的主要进化和发展, 发生在2.1——1.8, Ga, 这可能是在大气圈的氧气水平增加到1%~2% PAL的时候、真核生物获得了叶绿体的结果(Knoll, 1992)。尽管确切的时间点还难以确定, 但是, 在中元古代的1.5——1.4, Ga, 很明显, 来自于宏观化石和微化石的证据均表明了真核生物开始多样化, 而且在1.2, Ga左右, 化石记录表现出明显的真核生物多样性; 微化石证据还表明, 推测的菌类亲缘关系和陆生微生物群落, 发育在更加氧化的环境之中(Knoll, 1992, 2013; Javaux et al., 2004; Parnell et al., 2010)。
在GOE之后的古元古代晚期, 地球明显经历了一个长时间的环境稳定期, 以较为平稳的碳同位素变化为特征(图 1-d), 被形象化地描述为"令人烦恼的10×108年(boring billion 1000, Ma)", 大致从1.85, Ga持续到 0.85, Ga(Holland, 2006, 2009), 具有少量或没有大陆冰席, 伴随着一个生物革新速率减小的记录。针对这个特殊的时期, Buick(2007)认为, 在有效铜缺乏的情况下, N2O可能逃逸到大气圈, 这种强有力的温室效应气体的持续供应, 可能有助于这个时期免于大陆冰席的覆盖(Knoll, 2013); 但是, 这个认识, 受到1.8, Ga 的King Leopold冰川作用的挑战(图 1-c中的"(3)"所示), 这一次冰川作用的存在(Williams, 2005, 2008), 颠覆了以前关于2200——800, Ma期间没有发生冰川作用的普遍认识, 因为这一次冰川作用还被解释为与哥伦比亚超大陆的汇聚(图 1-b中的"[3]"所示)存在关联; 但是, 这一次冰川作用代表的大气圈变冷过程以及对应的氧气上升, 还有待于进一步研究。更要注意的是, 在这个较长的时期, 大气圈的氧气含量水平约束得较差, 推测处于5%~18% PAL的水平(Canfield and Teske, 1996), 较多的讨论还涉及到这个时期的深部大洋的化学构成和氧化——还原条件。在1.80, Ga, 条带状铁建造(BIF)的消失得出的结论是, 大气圈的氧气含量水平增加到足以向深海通气, 以至于可溶的铁不能持久保留下来(Holland, 1984, 2006); 但是, 这个认识, 就像Slack和Cannon(2009)将BIF在1.85, Ga的消失, 解释为巨型Sudbury地外流星的撞击事件的结果那样, 即撞击事件造成全球性海洋深部缺氧水体与浅水含氧水体混合作用而产生不利于BIF形成的环境, 还存在不同的认识和解释; Canfield(2005)认为, 不是氧化作用, 而是深部大洋在大约1.8, Ga变成硫化海洋, 并作为海水硫酸盐储库建造的自然结果造成了BIF的消亡, 即硫酸盐随后在深部大洋中变成最为丰富的氧化剂, 而且海洋转变成缺氧和硫化的静海, 从而阻碍了BIF的形成, 这样的作用过程是通过硫酸盐还原反应的增强所产生的氧气亏损和硫化物建隆的结果, 也是伴随着大气圈氧气含量的上升、光合作用蓝细菌激发的硫酸盐还原细菌繁盛的结果(Meyer and Kump, 2008)。更为关键的是, BIF的消亡, 实际上要年轻于以前的认识, 新的数据得出的结果是发生在1.78, Ga, 要晚于巨型Sudbury地外流星的撞击事件(Van Kranendonk et al., 2012)。
Canfield(1998, 2004)曾经建议到, 中元古代的海洋是一个分层的海洋, 具有一个氧化的表面海水层, 以及一个静海的、而且以一个全球性的硫化氢储库为特征的深水水体为特征, 这样的海洋属于典型的"静海"模型(Meyer and Kump, 2008), 从而被誉称为"Canfield"海洋(Van Kranendonk et al., 2012)。这个模型得到了海水中较低的硫酸盐浓度(大致为现代值的5%~15%)、以及一个缺氧而且硫化的深部水体特征的支持(Shen et al., 2002, 2003; Arnold et al., 2004), 也得到了块状硫化物矿床大量发育的支持。
大气圈氧气含量水平, 在这个时代被估计为8%~15%PAL(图 1-a)。Canfield(2005)以及 Buick(2007)均认为, N2O在这个时代可能已经变成较为重要的温室效应气体。从一个富铁的而且贫乏氧气的古元古代海洋, 转变成一个中元古代的硫化海洋, 被认为是与大气圈氧气含量水平上升相关联的下列作用过程的结果:(1)海洋的铁与氧气储库的充满; (2)暴露的大陆上的岩石中的硫化物的得到增强的风化过程, 向海洋输入了更多的硫酸盐, 从而导致更加广泛的微生物异化硫酸盐还原反应而产生更多的硫化物。这样一个"Canfield"海洋(Van Kranendonk et al., 2012), 经历了哥伦比亚超大陆的汇聚和裂解过程(图 1-b中的"[3]"所示; Rogers and Santosh, 2002; Zhao et al., 2002; Reddy and Evans, 2009), 哥伦比亚超大陆, 形成于1.92——1.76, Ga的全球性地体增生和造山活动(包括超高温的变质作用; Santosh et al., 2007), 在这个时期, 氧气水平已经增加到这样的程度, 以至于在一些黑色页岩沉积中发育天然裂变反应器(即富集铀), 大气圈氧气含量水平的连续增加, 还导致了中元古代早期真核生物的连续发育(Van Kranendonk et al., 2012)。在这个时期大气圈的氧气含量水平继续上升(Canfield, 2005), 还得到了年轻于大约1.8, Ga的岩石的块状菱铁矿层缺乏的进一步印证(Ohmoto et al., 2004); 但是, 在元古宙大气圈中的甲烷浓度, 可能还保持在较高的水平, 通过大气圈中的一层较薄的有机薄雾而使大陆保持温暖(Kasting, 2005)。中元古代晚期, 还经历了一个主要的超大陆, 即罗迪利亚超大陆(图 1-b中的"[4]"所示), 在1300——900, Ma的形成(Li et al., 2008, 2013); 再者, 罗迪利亚超大陆的穿时性裂解, 通过广泛的裂谷过程大致发生在825, Ma, 这大致是广泛的新元古代(成冰纪)冰川作用首次开始的时间, 而且持续到600, Ma, 即新的冈瓦纳超大陆(图 1-b中的"[5]"所示)还处在汇聚阶段的时候(Li et al., 2008, 2013), 从此, 地球进入了又一个新的发展时期。
古元古代GOE之后, 深部海洋还保持一个缺氧状态, 就出现了一个相对较为新颖的元古代海洋模式, 这就是一个得到广泛接受的氧化-还原分层模式, 在大约1.84, Ga之后表现为广泛的缺氧状况, 甚至可能影响到透光带。但是, 缺氧-含铁条件, 可能是前寒武纪海洋的主要特征(Poulton and Canfield, 2011), 凭借此, 元古宙海洋的氧化——还原结构的空间变化至少盛行在GOE之后、并一直持续到元古宙——显生宙界线, 而且硫化条件可能限制了近滨环境, 与今天海洋中的氧气最小带相似(Poulton et al., 2010)。
成冰纪与埃迪卡拉纪组成的新修订的新元古代(Van Kranendonk et al., 2012), 是一个极端动荡的时代, 表现在:(1)罗迪利亚超大陆的裂解; (2)成冰纪的全球性冰川作用; (3)冰川作用之后朝向更加稳定气候的转变; (4)壮观而且快速的复杂多细胞生命的进化。在这个环境波动的框架内, 新元古代巨型氧化作用事件(NOE; 图1-a), 处在一个生物与非生物领域之间的关键界面上(Och and Shields ̄Zhou, 2012)。过去数十年的持续研究, 使得前寒武纪——寒武纪过渡时期大气圈和海洋氧气浓度的上升的现象, 已经越来越得到确信。与中元古代"令人烦恼的1000, Ma"形成对照, 元古宙的最后300, Ma(850——541, Ma), 被形象化地称作"咆哮的300, Ma(Roaring 300, Ma; Holland, 2009)", 反映在碳同位素异常事件的巨大变化之中(图 1-d)。
所推断和总结出的NOE(Campbell and Allen, 2008), 主要基于以下证据:(1)高分辨率的碳和硫同位素数据, 来自于阿曼Huqf超群的新元古代晚期的海相沉积物之中, 表明了这个时期的氧气水平的3次增加(Fike et al., 2006); (2)δ 34S值的明显增加, 沉积物中的硫酸盐和硫铁矿的硫同位素差异已经在澳大利亚和纳米比亚被发现(Hurtgen et al., 2005; Halverson and Hurtgen, 2007); (3)来自于加拿大纽芬兰的新元古代晚期的沉积物中的化学与生物活跃铁的比率值的变化(Canfield et al., 2007); (4)在663——551, Ma期间的黑色页岩中氧化——还原敏感的钼含量的特征性增加(Scott et al., 2008); (5)这个时期对应着 87Sr/86Sr 值的明显增加(Campbell and Allen, 2008)。在NOE期间, 向大气圈增添了9×1017 kg氧气, 至少是GOE所产生氧气的10倍(Campbell and Squire, 2010)。
最为重要的是, 碳同位素记录的极端变化, 在这个时期期存在一些独特之处(图 1-a; Kump, 2008; Campbell and Squire, 2010; Kump et al., 2011; Och and Shields ̄Zhou, 2012):(1)在800, Ma之后, 一个明显长时间的δ 13Ccarb正异常事件, 曾经被解释为有机碳埋藏的加强, 因而对应的是地球表面环境氧化作用动力的释放; (2)在埃迪卡拉纪中期的极端δ 13Ccarb负异常事件, 则表明了一个很大规模的有机碳储库的氧化作用, 可能是大洋循环过程得到增强、以及所产生的深部大洋通气作用的结果。关于精确的生物地球化学机制, 也存在一些质疑, 这样的碳循环的波动, 在没有全球性氧储库的巨大变化的情况下是难以解释的, 尤其是碳同位素异常明显的全球性特征。对埃迪卡拉纪中期的极端负异常事件(图 1-c中"(6)"所示的Gaskiers冰川作用之后; Narbonne et al., 2012), 在阿曼Shuram组的原始发现的而且被命名为Shuram异常事件, 这样的异常事件紧紧地超前于进化和演变事件, 包括大型后生动物的起源和动物的生物矿化作用的起源之类的进化与演变事件(Grotzinger et al., 2011)。现行普遍的解释, 代表了一个潜在性的尝试性假说, 即大气圈含氧量明显在大约550, Ma之后的上升, 同时与之对应的是在埃迪卡拉纪末期宏体动物的多样化事件。另外, 整个新元古代一直进入寒武纪的海水沉积物中的 87Sr/86Sr 值的上升, 可能说明了化学风化过程作用强度以及向海洋提供的营养物的上升, 从而造成了包括光合作用蓝细菌的原始生产率的上升、以及有机碳埋藏速率的上升。元古宙晚期的Shuram异常事件, 以极端碳同位素负异常为特征, 与上文所述的Lomagundi-Jatuli碳同位素正异常事件(LJE)(Melezhik et al., 2005, 2007)所代表的元古宙早期的大规模碳循环形成对照, 所产生的图1-d所示的碳循环波动的特征性曲线, 以及所暗含的成因机理和科学意义, 正在成为科学家们探索的重要主题。
被称作"咆哮的300, Ma"(大致为850——550, Ma), 大致从成冰纪晚期到埃迪卡拉纪早期, 氧气含量水平肯定明显上升, 大气圈的氧气含量水平被认为从5%~10% PAL增加到60%~100% PAL(Canfield, 2005), 形成新元古代巨型氧化作用事件(NOE; 图1-a; Kump, 2008; Campbell and Squire, 2010; Kump et al., 2011; Och et al., 2012); 大致在500, Ma左右氧气可能接近现代水平(21%), 而且5×108年来至今天波动在15%~35%之间。
在大约750——580, Ma, 越来越得到一致认识的是, 3次大陆间的、甚至是全球性的冰川作用事件的发生; 冰川作用的证据, 来源于那些沉积在被动裂谷盆地中的混杂砾岩, 这些裂谷盆地形成于罗迪利亚超大陆裂解期间(Shields-Zhou and Och, 2011, 2012; Van Kranendonk et al., 2012)。根据界线地层的U-Pb锆石年代学特征, 3次冰川作用包括(图 1-c中的"(4)、(5)、(6)"所示):(1)720——658, Ma的Sturtian冰川作用; (2)655——635, Ma的Marinoan冰川作用; (3)584——582, Ma的Gaskiers冰川作用。如果将这些冰川作用理解为大气圈变冷过程的话, 它们与几乎同时代的碳同位素异常事件代表的大规模碳循环波动、以及这个时期的大气圈氧气含量水平上升之间的复杂耦合关系, 还是一个有待于进一步探索的重大科学命题。
NOE表征的新元古代巨大转变, 代表了全球性碳循环的所有方面的一级变化(风化过程、生物扰动作用、碳的埋藏、有机生产率、氧化——还原状态), 但是, 它们的结果和复杂的相互作用机制, 还有待于探索与研究(Och et al., 2012)。在600——800, Ma, NOE所表征的大气圈氧气水平的上升, 可能是那些具有较高新陈代谢速率的生物, 尤其是后生动物起源与进化的主要控制因素, 另外一个重大变化是深部大洋的氧化作用(Sessions et al., 2009; Holland, 2009; Grotzinger et al., 2011; Och et al., 2012; Van Kranendonk et al., 2012; Sperlinga et al., 2013)。
对于NOE的成因, Campbell 和 Squire(2010)将其解释为下列地球动力学过程的结果。东冈瓦纳与西冈瓦纳的汇聚, 不但产生一个冈瓦纳超大陆(图 1-b的"[5]"所示), 而且了一个大约8000, km长的环冈瓦纳超级山脉带, 这些山脉的侵蚀基底, 包括了最古老的含硬柱石蓝片岩和榴辉岩, 以及高压变质岩石, 这些低热梯度和高压变质岩石的突然出现, 意味着冈瓦纳造山带相对较冷而且更加坚硬, 与早期的超大陆汇聚相联系, 支撑着高山山脉的发育。因此, 在地势起伏与侵蚀作用速率之间, 存在着一个对数线性关系, 而且在沉积作用速率与有机碳埋藏速率之间存在着一个线性关系。环冈瓦纳超级山脉, 要高于较早期的超大陆汇聚期间所形成的山脉, 而且这些山脉的快速侵蚀作用向海洋和河流释放了大量的基本营养物, 包括铁和磷, 从而激发了藻类与蓝细菌的爆发; 这个过程, 反过来造成了光合作用氧气生产作用的明显加强, 而且在这个时期的快速沉积作用, 又促进了在光合作用期间产生的有机质和生物成因的黄铁矿的较高的埋藏速率, 以至于它们不能与氧气反应, 最终导致了大气圈氧气的一个支撑性的上升。这次事件向大气圈增添了9×1017 kg氧气, 至少是GOE所产生氧气的10倍。这些富有理性的认识, 成为现今对NOE成因相对合理的解释。
要特别指出的是, NOE表征的新元古代巨大转变, 代表了全球性碳循环的所有方面的一级变化(风化过程、生物扰动作用、碳的埋藏、有机生产率、氧化——还原状态), 但是, 它们的结果和复杂的相互作用机制, 还有待于探索与研究(Och et al., 2012)。虽然元古宙早期的GOE的概念已经被广泛接受, 但是, 由于针对新元古代NOE的时间进程和变化程度的约束还是相对较少, 从而产生了一些争议; NOE合适的证据, 包括大规模气候波动的背景、与罗迪利亚超大陆裂解和冈瓦纳大陆再次汇聚相关的构造抬升, 还有埃迪卡拉纪生物群和寒武纪"大爆发"所反映的主要的生物革新(Och et al., 2012; Sperlinga et al., 2013)。总体较高的δ 13C值, 可能是有机碳埋藏的结果, 而且有机碳埋藏的加强, 又是地球表面环境在大约800, Ma之后, 氧化能力释放的结果; 在大约580, Ma之后, 极端的δ 13C负值的全球性记录(Shuram异常事件; Fike et al., 2006; Grotzinger et al., 2011), 强烈表明了可溶有机碳储库(DOC)的氧化作用, 其中在555, Ma的峰值, 与埃迪卡拉纪宏体生物群的突然多样化事件相重合。朝向新元古代与寒武纪界线, 87Sr/86Sr 值的增加, 表明了大陆风化过程的增强, 为新元古代晚期较高的有机生产作用和埋藏过程提供了有利条件。所以说, 与元古宙早期的GOE一样, 元古宙晚期的NOE, 是一个与复杂的地球动力学过程(超大陆裂解、冰川作用代表的大气圈变冷、碳同位素异常代表的海洋碳循环波动等)紧密相关的地球生物学过程, 这个特别的地球生物学过程反过来激发和促进了相关的生物革新。
伴随着NOE代表的大气圈氧气含量水平的上升, 可能是生物多样性和复杂性最为明显飞跃的重要原因, 最为重要的生物事件是复杂的多细胞动物的起源, 如:(1)阿曼的海绵生命标志物(Knoll and Carroll, 1999; Knoll, 2003; Gaidos et al., 2007; Saul, 2009; Gaidos, 2010); (2)850, Ma岩石中的后生动物组织级别的结构性证据(Neuweiler et al., 2009); (3)中国的滞育卵与动物胚胎(Yin et al., 2007; Wang et al., 2009); (4)Marinoan冰川作用前的岩石中发现的钙化的海绵后生动物化石(Maloof et al., 2010); (5)在新元古代冰川作用结束后不久, 早期动物进化的化石, 较为独特地保存在各种海相沉积岩中, 包括成型为球形、碟状、多叶体、蠕虫状以及其他的放射状和双叶状的实体结构(Narbonne, 2005; Canfield et al., 2007)。更为有趣的是, 较深水的底栖软体宏体化石(埃迪卡拉生物群; Narbonne, 2005; Yuan et al., 2011; Narbonne et al., 2012), 较为宽泛地与Marinoan冰川作用事件(655——635, Ma; 图1-c中的"(5)"所示)结束后、大气圈氧气增加到大约0.1 APL水平的上升相重合, 这个时期还以产生于永冻层融化的明显甲烷释放、以及海洋酸化作用为特征(Fike et al., 2006; Canfield et al., 2007; McFadden et al., 2008; Shields-Zhou and Och, 2011; Shield ̄Zhou et al., 2012)。但是, 一直到冰期之前, 后生动物没有得到真正的发展, 因为地表环境可能通过Marinoan冰川作用后的氧气上升而发生改变(Bao et al., 2008; Love et al., 2009; Yuan et al., 2011; Shields-Zhou and Och, 2011; Shields ̄Zhou et al., 2012)。
大气圈与海洋中的氧气历史, 一直引起科学家的兴趣, 尤其是针对早期动物进化的环境约束的关注(Knoll, 2013), 因为没有任何一个长周期变化轨迹具有如此的吸引力。越来越多的地球化学数据, 支持了以下假说:一个更加持续氧化的海洋, 只是到了埃迪卡拉纪晚期才真正出现, 更有可能是在580——560, Ma之后(Li et al., 2010; Johnston et al., 2012), 从而代表了一个受到延迟的生物圈的氧化作用(Kah and Bartley, 2011)。就其本身而言, 这与分子钟所估计到的800, Ma开始了动物多样性并不矛盾(Erwin et al., 2011), 但是, 这个状况确实约束了适用于最早期动物的进食样式和生命体制计划(Knoll, 2013)。这个推断和认识, 主要基于以下事实, 即成冰纪到埃迪卡拉纪中期的任何双侧动物世系, 没有见到任何肉食性的证据; 假定生态与物理环境是紧密联系的, 肉食动物的起源, 可能是一种新陈代谢昂贵的觅食策略, 而且明显受到大气圈氧气含量水平上升的驱动(Erwin et al., 2011)。
新元古代巨型氧化作用事件(NOE)之后, 在600, Ma左右, 即在新元古代晚期, 大气圈的氧气分压才接近现代的水平(Berner et al. 2003; Canfield et al. 2007); 大气圈氧气水平在整个显生宙所具有的变化是在石炭纪晚期接近一个峰值为150% PAL(大气圈氧气含量达到了31%), 在侏罗纪早期处于一个低值为60% PAL(大气圈氧气的含量为12%), 然后上升到现代的水平(21%)。来源于详细的模拟, 运用了硫和碳的同位素质量平衡(Berner, 2006), 表明了硫酸盐中的 δ 34S 与碳酸盐中的δ 13C同位素的变化(图 1-d)发生在360——260, Ma期间, 这归因于大气圈氧气含量水平从350, Ma的80% PAL增加到260, Ma的140% PAL(相当于31%; 图1-a), 这是地球历史进程中, 大气圈氧气含量水平唯一一次超过现代值的一次显著的大气圈氧气上升, 所以被誉称为显生宙的巨型氧化作用事件(POE; 图1-a)。最为有趣的是, 在Berner模型中(Berner, 2006), 350——275, Ma、以及650——525, Ma, 增添到大气圈中的氧气量的计算结果均是9×1017 kg(Campbell and Squire, 2010)。还需要强调的是, 关于420, Ma至现在的大气圈氧气含量水平, 较紧密的限定和约束, 是通过木炭化石堆积的连续记录设置的, 即:(1)在60% PAL的氧气含量水平之下, 火焰不可能持续; (2)由于高于大约160% PAL的氧气水平, 将造成野火频率和活力的大大增强, 从而破坏森林生态体系, 所以在石炭纪晚期接近的一个峰值, 被设置为140%或150% PAL(Kump, 2008)。
Berner(2006)长期研究的结果表明, 由于陆生植物的扩张所造成的风化作用速度的增加, 会加强埋藏有机碳的保存而抗拒微生物对有机碳的降解和破坏, 大量褐煤的埋藏, 在石炭纪和二叠纪煤系地层中较为普遍, 也可能是造成350——275, Ma氧气增加的主要原因(Berner, 2006), 从而导致了从泥盆纪到石炭纪的大气圈二氧化碳的主要下降事件, 并产生了全球性的变冷过程(石炭纪至二叠纪冰川作用; 图1-c中的"(8)"所示)和相应的大气圈氧气上升(POE), 还与泛大陆(图 1-b中的"[6]"所示)汇聚作用存在着关联(Campbell and Allen, 2008; Och et al., 2012; 梅冥相和刘少峰, 2013)。POE的形成, 那个时期节肢动物和爬行状动物的巨型畸形动物的发育, 支持了这个认识, 因为这个生物学现象也应该归因为大气圈氧气浓度的上升(Payne et al., 2011)。Berner(2006)还进一步指出:(1)POE代表的大气圈氧气上升的主要原因, 被确信为大型陆生维管植物的进化, 因为大型陆生维管植物造成了有机质埋藏的增加(以及由此而造成的氧气生产的增加), 而且这是一种新的抗生物降解的物质的介入, 这种新物质就是木质素; (2)就像这个时期较为丰富的化石木炭所证实的那样, 木质素和其他植物碎屑在古生代晚期(~360——260, Ma)的沼泽中的过剩埋藏, 导致了巨量煤的沉积而且增强了野火的发生。
发生在350——275, Ma之间的大气圈氧气的上升, 还被解释为形成于泛大陆汇聚期间的超基山脉的快速侵蚀作用的结果; 这个山链, 大约为8000, km长, 宽度可达1000, km(Matte, 2001), 大小类似于冈瓦纳超级山脉; 最为有趣的是, 在Berner模型中(Berner, 2006), 350——275, Ma、以及650——525, Ma, 增添到大气圈中的氧气量的计算结果均是9×1017 kg(Campbell and Squire, 2010)。与Berner(2006)的解释结合在一起, 说明了POE也是一个与特别的地球动力学过程(超大陆汇聚产生的超级山链、冰川作用等)紧密相关的地球生物学过程。
相似地, 在白垩纪之后被子植物的扩张, 至少部分影响了新近纪晚期的全球变冷过程和相应的大气圈氧气上升, 从40, Ma以来到现在的氧气上升, 似乎还与喜马拉雅山脉的隆升存在着一些联系(Berner, 2006; Campbell and Allen, 2008; Campbell and Squire, 2010)。相似的情况是, 哺乳动物的个体大小, 在古近纪的增大, 也归因于大气圈氧气含量水平的上升(Falkowski et al., 2005; Payne et al., 2011)。氧气可用性的增加可能会促进哺乳动物的辐射(Butterfield, 2009; Payne et al., 2011), 就像NOE与动物后生起源和进化之间的复杂关系那样, 鸟类和哺乳动物的中生代进化、以及随后的大型有胎盘哺乳动物的始新世多样性, 可能得益于侏罗纪到现在的大气圈氧气含量水平上升(Falkowski et al., 2005)。
发生在元古宙早期(2.5——2.0, Ga)的GOE, 大气圈氧气含量水平快速上升, 尽管可能只是达到了百分之几的PAL, 但是, 由于明显的上升速度和奇特的属性, 这个时间段的大气圈氧气含量的上升被定义为巨型氧化作用事件(GOE; Kirschvink et al., 2000; Canfield, 2005), 标定了地球大气圈第1次从一个缺氧的大气圈转变成一个含氧的大气圈。大致在800——600, Ma期间, 大气圈氧气水平发生了第2次增加(Canfield and Teske, 1996), 这就是新元古代巨型氧化作用事件(NOE), 伴随着这次事件的是深部大洋的氧化作用和多细胞后生动物的出现(Canfield, 2005; Campbell and Allen, 2008; Holland, 2009; Campbell and Squire, 2010; Grotzinger et al., 2011; Kump et al., 2011; Shields-Zhou and Och, 2011; Tzipermana et al., 2011; Och et al., 2012; Sperlinga et al., 2013), 在NOE之后, 大致在500, Ma左右, 地球大气圈含量增加到现代水平, 标定了显生宙一个相对稳定的大气圈氧气含量水平的开始。在显生宙, 最为特征的变化是在石炭纪晚期, 一个接近于150% PAL的大气圈氧气含量峰值(大气圈氧气含量达到了31%), 这就是文中所称的显生宙大气圈氧化作用事件(POE)。如果将大气圈氧气含量水平的上升, 理解为一个主要的地球生物学过程的话, 就像图1-a中的GOE、NOE和POE所代表的那样, 因为产生氧气的光合作用过程本身就表明了氧气的生物成因, 那么, 这一个复杂的地球生物学过程与地球动力学过程(超大陆旋回、全球性碳同位素异常事件代表的碳循环的大规模波动、冰川作用代表的大气圈变冷过程等)之间的复杂关系和响应机制, 将是一个永恒的主题。
一直到今天, 在生氧光合作用期间产生水分解的过程和机制还不清楚, 也不知道究竟是什么因素控制了地球大气圈气体构成的浓度(Falkowski and Godfrey, 2008; Falkowski and Isozaki, 2008; Kump, 2008)。前一个问题, 应该在数十年内, 借助于高分辨率的光合体系结构、以及复杂的生物物理学途径来测度电子转化反应, 而得到解决(Barber, 2008); 后一个问题, 将很难得到约束, 随着一个更好的模型将更加综合的生物地球化学测量综合起来, 可能会得到一个较好的了解(Hayes and Waldbauer, 2006)。
在地球的生命体系中, 已经成为主要的转变能量途径的非平衡氧化——还原反应网的进化, 很多问题确实不知道。不可避免的是, 氧气成为最终的电子储库。但是, 氧气的生物化学特征还是一个谜, 我们对氧气的光合作用反应的生产, 还没有完全了解, 或者说, 光合作用产生的氧气在新陈代谢和生物地球化学循环中的反馈, 也是一个没有得到完全了解的问题; 尽管在《物种起源》出版之后已经经历了150年, 生命的起源始终是一个黑箱(Falkowski and Isozaki, 2008; Falkowski and Godfrey, 2008)。
地球历史中的3次重要的巨型氧化作用事件, 是复杂的地球生物学过程与复杂的地球动力学过程相耦合的结果(图 1), 尽管还存在很多细节问题需要进一步调查和研究, 但是, 科学家们多年的持续性研究还是为我们提供了一个地球大气圈氧气上升的综合图像, 在这个艰辛的研究过程之中, 从多学科研究到高度综合, 与冯增昭教授(2004)倡导的"单因素分析多因素综合"作图法来进行区域古地理重建的富有智慧的思维方法, 具有异曲同工之妙; 还需要强调的是, 冯增昭(2013)主编的《中国沉积学》(第2版), 不但是"单因素分析多因素综合"富有建树的思维方法的拓展, 而且为今天的沉积学研究, 提供了重要的理论基础和学科生长点。因此, 立足于前人的研究成果, 总结地质历史时期的巨型氧化作用事件的特征和属性, 推崇、倡导和学习"单因素分析多因素综合"的创造性思维和科学研究方法, 将是对中国古地理学先哲冯增昭先生90华诞的最好祝愿!
元古宙早期的GOE, 基本上得到了确定; 然而, 元古宙晚期的NOE, 是近年来得到强调的一个新概念。再者, 将石炭纪晚期接近一个峰值为150% PAL(大气圈氧气含量达到了31%)的大气圈氧气上升, 理解为POE, 可能会有助于理解地球历史进程中大气圈氧气含量水平上升的复杂过程和反馈机制。形成这些巨型氧化作用事件的内在成因、启动机制的差异性和共同点, 还没有得到完全的了解, 例如:(1)GOE的形成, 可以解释为一个稳定构造背景和岩石圈变冷过程产生的微生物的繁盛与扩张的结果, 相应的是一个几乎同时代的大气圈变冷过程和冰川作用期, 但是, 这个似乎合理的多重控制机制又不太适合新元古代的NOE; (2)巨型氧化作用事件均对应于明显的碳同位素异常事件, 但是, GOE与POE主要对应的是正异常代表的碳循环波动, 而NOE则对应着明显的负异常, 这些差异代表的地球动力学和地球生物学过程, 还有待于进一步研究; (3)巨型氧化作用事件与超大陆旋回之间, 也是一个非常复杂的关系, 因为GOE和NOE似乎主要对应着超大陆的裂解期, 但是, POE则与泛大陆的汇聚作用存在关联; (4)气候与地表氧化作用增强之间的联系的准确属性, 还是一个需要进一步思考的问题, GOE、NOE和POE总是与广泛冰川作用期存在着一个接近同时性的时间关系, 成为确实让人惊奇的现象; 再者, 发生在2.9, Ga的地球上最古老的冰川作用(图 1-c中的"(1)"所示)所代表的局部寒冷的大气圈和相应的弱氧化的大气圈, 还没有得到足够的重视, 1.8, Ga的King Leopold冰川作用所蕴含的全球变化, 也需要进一步研究才能得到阐释。这些问题的提出, 将成为今后采用"单因素分析多因素综合"的富有智慧的思维方法进行深入研究的强大驱动力。地质历史时期的巨型氧化作用事件, 以及所涉及到的复杂的地球动力学过程和地球生物学过程, 为拓宽沉积学的研究范畴提供一些有益的思考途径和研究线索; 追逐这些重要概念及其所产生的若干新认识和新问题, 希望能够激发同行的关注而最终起到一个抛砖引玉的作用, 并以此祝贺古地理学先哲冯增昭先生90华诞!
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|
| [153] |
|
| [154] |
|
| [155] |
|
| [156] |
|
| [157] |
|
| [158] |
|
| [159] |
|
| [160] |
|
| [161] |
|
| [162] |
|
| [163] |
|
| [164] |
|
| [165] |
|
| [166] |
|
| [167] |
|
| [168] |
|
| [169] |
|
| [170] |
|
| [171] |
|
| [172] |
|