




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
全新世以来浙江余姚河姆渡—田螺山遗址的古环境演变及人类活动*
[郑丽波1  , 郝秀东
, 郝秀东2 , 禚彬3 , 柳云龙3 , 王小龙1 , 朱立新1 , 余新波4 , 张甬江5 ]
, 郝秀东, 禚彬|
|

第一作者简介:郑丽波,女,1973年生,教授级高级工程师,主要从事第四纪环境与水文地质研究。E-mail: 121837499@qq.com。
通讯作者简介:郝秀东,男,1980年生,同济大学海洋与地球科学学院博士研究生,现主要从事孢粉学与环境考古研究。E-mail: xiudonghao@126.com。
通过对浙江余姚河姆渡—田螺山遗址附近的 HMD1501和 HMD1502两个钻孔的微体古生物学研究,揭示了该地区全新世以来的植被演变、气候变化、海平面变迁及人类活动。孢粉、海相沟鞭藻、淡水藻类及有孔虫内衬的记录表明: ( 1)在早全新世(对应于镇海组下部),植被群落属于常绿阔叶和针叶混交林,气候温暖潮湿,研究区属于海侵阶段。( 2)在中全新世(对应于镇海组中部),植被群落虽仍为针阔叶混交林,但气候转为温和湿润,研究区仍属于海侵阶段。( 3)由于人类活动的进一步加剧,在晚全新世(对应于镇海组上部)的植被群落中草地的面积也随之扩大,并在滨海地区开始有一些以莎草科为主的沼泽地形成,植被群落属于针阔叶混交林—草地,气候更加凉爽干燥,研究区转为海退阶段。( 4)在 ~7235— 3327 cal. yr B.P.期间,草本花粉的含量激增,特别是一些粒径不小于 35, μm 的禾本科栽培作物花粉的含量增加,说明此时人类活动开始增强,河姆渡—田螺山遗址附近出现较多的水稻种植区。
, Hao Xiudong, Zhuo BinAbout the first author:Zheng Libo,born in 1973,is a professor of Zhejiang Hydrogeologic and Engineering Geological Brigade.She is mainly engaged in researches on Quaternary environmental evolution and hydrogeology. E-mail: 121837499@qq.com.
About the corresponding author:Hao Xiudong,born in 1980,is a Ph. D. candidate of Tongji University. He is engaged in researches on palynology and environmental archaeology.E-mail:xiudonghao@126.com.
Micropaleontological record of the two boreholes(HMD1501 and HMD502)from Hemudu-Tianluoshan Sites in Yuyao of Zhejiang Province,reveals a detailed vegetation history,paleo-vegetation evolutions,palaeoclimate change,fluctuating sea level and human activity in the study area during the Holocene.Palynological results(pollen,spores,dinoflagellate,freshwater algae and foraminifer linings)suggest that: (1)Evergreen coniferous and broadleaved mixed forest developed during the Early Holocene(corresponding to the lower of Zhenhai Formation),the climate was warm and humid,and the study area was belong to a transgression stage. (2)Evergreen coniferous and broadleaved mixed forest were still developed during the Middle Holocene(corresponding to the middle of Zhenhai Formation),the climate changed into moderate and humid,and the transgression continued. (3)Due to human activity,coniferous and broadleaved mixed forest-grassland developed during the Late Holocene(corresponding to the upper of Zhenhai Formation),the climate changed into cold and dry,however,the study area began to enter a regression stage. (4)Human activities began to strengthen,and high percentage Poaceae pollen(≥35,μm)indicates rice culture activity presented at Hemudu-Tianluoshan Sites during the ca.7235-3327 cal.yr B.P..
20世纪70年代, 作为中国新石器时代考古史上重大发现之一, 距今约7000年的河姆渡文化的发现和发掘, 对中国、乃至世界的稻作农业的起源与发展、南方地区史前建筑状况、史前人类的生活方式演化以及全新世古气候的演化等研究均产生了广泛而深远的影响(俞为洁, 1993; 劳伯敏, 1995; 江大勇等, 1999; 黄渭金, 2002)。在诸多的研究内容中, 古人类从事农业生产活动与自然环境状况的关系研究是非常重要的一个部分。近年来的研究多集中在河姆渡文化时期(孙湘君等, 1981; 郎鸿儒, 1987; 郑云飞等, 1994; 江大勇等, 1999; 刘军, 2006; 李明霖等, 2009; 周宗尧等, 2012), 而在河姆渡文化发展之前及之后的环境状况却较少涉及。
2004年开始发掘的田螺山遗址, 距离河姆渡遗址仅7, km, 在中国新石器时代遗址中属于河姆渡文化早期的一处重要遗址, 出土了大量的干栏式建筑、骨耜、炭化稻、木桨、编织物及龟甲等具有河姆渡文化特色的先民生产、生活遗存, 此外, 该遗址与河姆渡遗址有着相近的聚落规模和相似的年代跨度(浙江省文物考古研究所等, 2007), 为我们重新审视河姆渡文化发展前后的自然环境状况提供了一个绝佳的契机。本研究拟通过对田螺山南麓HMD1501和HMD1502钻孔的孢粉学研究, 运用孢粉、海相沟鞭藻、淡水藻类及有孔虫内衬等记录来重建和恢复河姆渡— 田螺山遗址及其附近地区全新世以来的植被演变、气候变化及海平面变迁, 为进一步研究河姆渡文化时期古人类从事农业生产活动及其与自然环境的关系提供资料。
田螺山遗址位于浙江省余姚市三七市镇相岙村, 地处姚江河谷北侧低丘环绕的小盆地中部, 北面横亘四明山支脉翠屏山, 东距海岸30~40, km, 西南距河姆渡遗址约7, km。该遗址围绕一个名为田螺山的小山头分布, 周围是大片低平湿软的稻田, 面积为3万多平方米, 海拔2.3, m, 埋深2~3, m(浙江省文物考古研究所等, 2007)。本研究的2个钻孔HMD1501和HMD1502均位于田螺山南麓, 其经纬度分别为: 29° 55'48.23″N、121° 26'39.60″E; 29° 55'48.23″N、121° 21'20.75″E(图 1)。
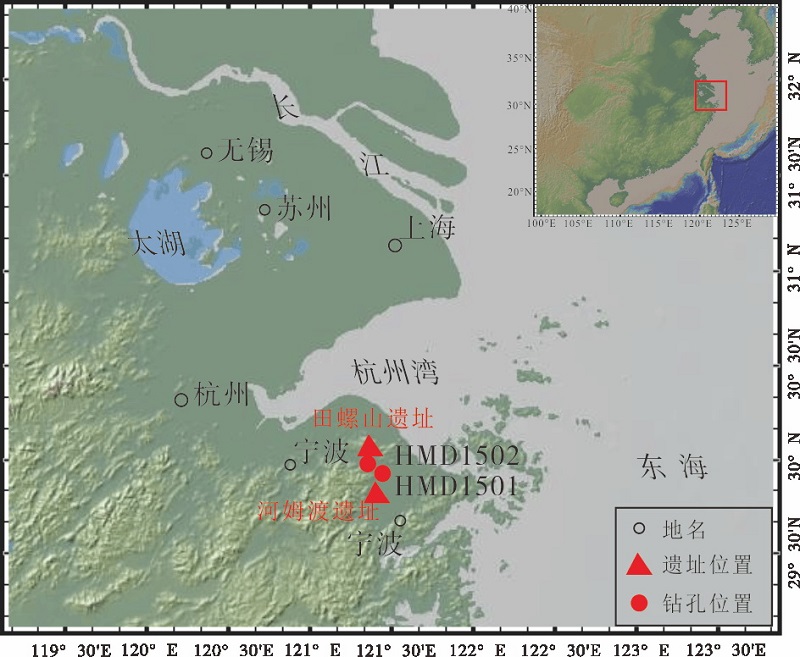 | 图1 浙江余姚田螺山HMD1501和HMD1502钻孔位置Fig.1 Location of Boreholes HMD1501 and HMD1502 at Tianluoshan of Yuyao, Zhejiang Province |
本区第四系包括中更新统、上更新统和全新统, 沉积物主要为灰色、黄灰色黏土和粉砂, 砂泥交互韵律层理发育, 另外下部有黄色砂砾石。第四系厚度介于40~100, m之间, 呈现明显向海变厚的变化规律。按照浙江省岩石地层分类标准, 共分为前港组、东浦组、宁波组和镇海组4个地层单位(浙江省地质矿产局, 1989)。本研究的2个钻孔HMD1501和HMD1502只涉及到全新统镇海组一个地层单位。
研究区属亚热带季风气候, 温和湿润, 四季分明, 雨量充沛, 冬夏季风交替明显。冬夏长(各约4个月)、春秋短(各2个月左右)。多年平均气温16.3, ℃, 7月份最热, l月份最冷, 极端最高气温39, ℃, 极端最低气温-11, ℃, 平均积温5100, ℃左右, 平均日照1900 h以上。多年平均降水量1400, mm左右, 山地丘陵一般要比平原多三成左右; 主要雨季出现在3— 6月份和8— 9月份。台风主要集中在7— 9月份, 暴雨主要出现在6月份至7月份上旬的梅雨季节和 8— 9月份的台风季节, 以9月份居多。冰雹和龙卷风出现的机率较小。龙卷风主要出现在6— 9月份, 其中7月份为多。
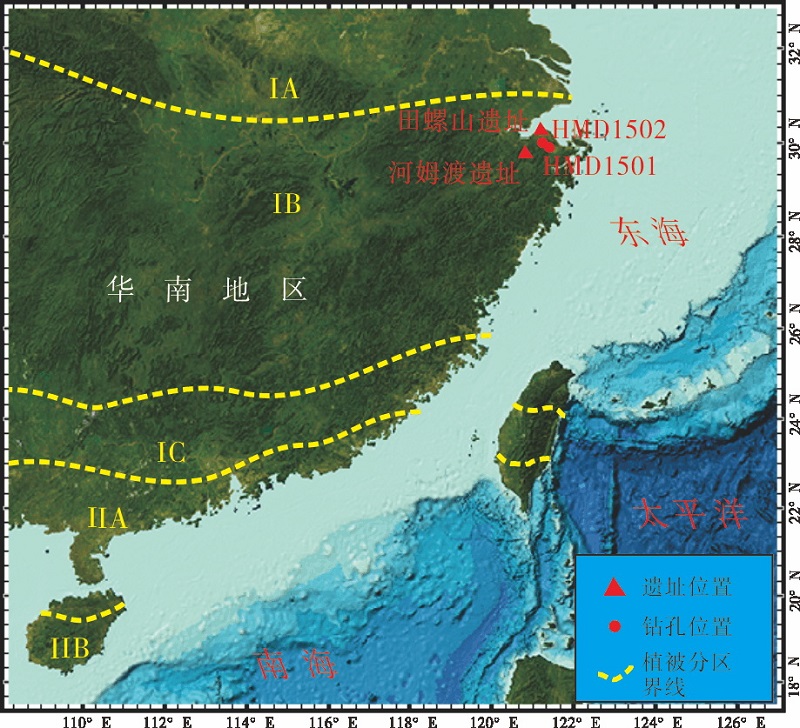 | 图2 研究区及邻区现代植被分带(修改于中国科学院中国植被图编辑委员会, 2007) IA— 北亚热带常绿— 落叶混交林; IB— 中亚热带常绿— 落叶混交林; IC— 南亚热带常绿— 落叶混交林; IIA— 北热带半常绿季雨林; IIB— 南热带季雨林和热带雨林。红色圆点为HMD1501和 HMD502钻孔位置Fig.2 Modern vegetation belts of the study area and its adjacent region(Modified after Editorial Committee of Vegetation Map of China, Chinese Academic of Sciences, 2007) |
研究区植被属中亚热带常绿阔叶— 落叶混交林北部亚地带, 浙闽山丘甜储、木荷林区(中国科学院中国植被图编辑委员会, 2007; 图2)。历史上森林植被茂密, 但由于解放前近百年累遭摧残, 后又复受破坏, 所以目前原始植被常绿阔叶林已几乎绝迹, 代之以针叶林、灌丛等次生植被及人工植被类型(吴征镒, 1980)。主要种类包括: 青冈属(Cyclobalanopsis), 苦槠(Castanopsis sclerophylla), 栲属(Castanopsis), 石栎(Lithocarpusglaber), 枫香(Liquidambar formosana), 马尾松(Pinus massoniana)等(《浙江森林》编辑委员会, 1993)。
本研究的 HMD1501 和 HMD1502 两个钻孔分别为 2015年5月份钻取的, 本次钻探为科学钻探, 采用加套管取心的方法, 共钻取的深度分别为 46.51 m 和 44.59 m, 在保证岩心质量的前提下, 每孔综合取心率达到 98% 以上。岩心的实验室处理在南京师范大学地理科学学院实验室, 按照剖样照相— 室内编录— 分样— 采样的流程处理。岩心经切割机剖样后, 一半密封放入冷库, 覆盖保鲜膜冷藏保存; 另一半照相存档, 再进行室内编录和分样, 在分样过程中, 将泥炭层、植物残体、贝壳等取出单独标记作为测年材料。 本研究分别选取其上部 24.01~0.61 m 和 29.61~1.11 m 进行孢粉学分析, 其具体岩性及采样方法如下:
2.1.1 HMD1501孔岩性 24.01~11.21, m, 以褐灰色黏土为主, 时有粉砂团块或薄层出现, 具层理, 含丰富的贝壳碎片; 11.21~2.57, m: 中间和底部为褐灰色黏土, 顶部为灰黑色黏土, 含植物残体斑块, 并分别在2.57~2.77, m和4.20~4.35, m处出现2层黑色泥炭; 2.57~1.01, m: 上层黏土具非常微弱的层理, 下层粉质黏土, 具微弱或明显水平层理, 底部与下层沉积为侵蚀突变; 1.01~0, m: 深灰色耕植土为主, 出现大量暗褐色斑点(可能与现代植物根系有关), 较致密(图3)。
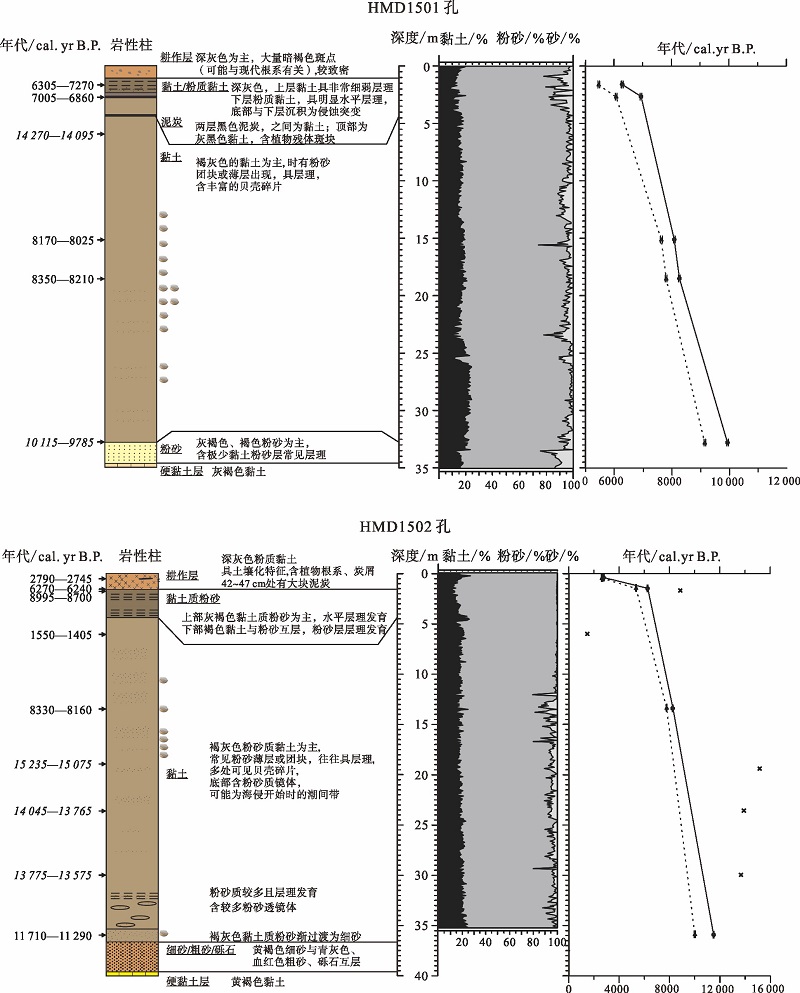 | 图3 浙江余姚田螺山HMD1501和HMD1502钻孔年龄— 深度曲线 黑色三角形为常规年龄, 黑色圆点为矫正后的日历年龄, “ 斜体字” 和“ × ” 表示异常的年龄值Fig.3 Age-depth curves of Boreholes HMD1501 and HMD1502 at Tianluoshan of Yuyao, Zhejiang Province |
2.1.2 HMD1502孔岩性 29.61~18.71, m, 以褐灰色粉砂质黏土为主, 出现粉砂薄层或团块。18.71~4.41, m: 以褐灰色粉砂质黏土为主, 常见粉砂薄层或团块, 往往具层理, 多处可见贝壳碎片; 4.41~1.51, m: 上部以褐灰色黏土质粉砂为主, 水平层理发育; 下部为褐灰色黏土与粉砂互层, 粉砂层层理发育; 1.51~0, m: 深灰色粉质黏土, 具土壤化特征, 含植物根系、炭屑, 并在0.42~0.47, m处出现大块泥炭(图3)。
2.1.3 样品采集 样品按照0.8, m等间距进行采集, 分别采集了HMD1501孔上部0.61~24.01, m岩心30份样品, 以及HMD1502孔上部1.11~29.61, m岩心36份样品, 共采集了66份样品。
所有孢粉样品的实验室处理均在上海师范大学旅游学院进行。实验室处理采用传统的过筛法(Brown, 2008), 即先加盐酸去除钙质, 再加氢氟酸去除硅质, 最后在超声波中用7, μ m网筛过滤。每个样品的孢粉统计数在250粒左右。在统计孢粉数目的同时记录样品中出现的海相沟鞭藻、淡水藻类、苔藓孢子及有孔虫内衬等, 孢粉图谱百分含量按孢子与花粉总数计算, 海相沟鞭藻、淡水藻类及苔藓百分含量以孢子、花粉、海相沟鞭藻、淡水藻类及苔藓的总数计算。此外, 舒军武等(2007)研究显示, 现代水稻花粉粒径大小在34~38.5, μ m之间, 据此, 本研究依据禾本科花粉粒径大小将其划分为≥ 35, μ m和< 35, μ m两组, ≥ 35, μ m的禾本科花粉中很可能含有较多的具有人工栽培作物的禾本科植物(如水稻等)的花粉。
孢粉(藻类)浓度计算采用“ 外加标志法” (Maher, 1981), 即将已知浓度的现代石松孢子(Lycopodium)(27, 637粒/片)作为标志添加到每一份样品中, 再依据鉴定和统计结果进行浓度计算。具体孢粉浓度计算公式如下:
孢粉浓度值(粒/mL)=
孢粉及沟鞭藻类等的鉴定和统计是在Leica DM4000 B光学投射显微镜10× 40倍下进行, 对个体极小的孢粉放大至 10× 60倍。孢粉及沟鞭藻类等的拍照使用Leica DFC450数码相机。使用Tilia(1.7.16)软件建立孢粉、藻类百分比及浓度的孢粉谱。
| 表1 浙江余姚田螺山HMD1501和HMD1502钻孔AMS14C年龄数据 Table1 Results of AMS14C dating in Boreholes HMD1501 and HMD1502 at Tianluoshan of Yuyao, Zhejiang Province |
AMS14C测年材料选取沉积物中的植物残片、贝壳及有机沉积物全样, 共15份测年样品, 经过简单预处理后送至 Beta Analytic公司进行测年, 结果见表1。采用最新版树轮矫正程序IntCal13(http: //www.radiocarbon.org/IntCal13.htm.)来进行日历年龄的校正, 校正后的日历年龄往往存在一个或多个年龄分布区间, 通常选取在2σ 置信区间下相对概率最大的矫正年龄区间(Talma and Vogel, 1993; Reimer et al., 2013)。
在地层中AMS14C年龄出现倒转的原因很多, 如侵蚀再沉积、生物扰动、采样时新老炭的交叉污染等, 而在河口、滨海地区, 因于水动力较强, 侵蚀再沉积造成的年龄倒转则更为常见(Pederson et al., 2005; 刘进峰, 2009)。如采集于HMD1501孔5.9, m处的有机沉积物测年结果明显偏老, 因此, 在建立该孔全新世地层年龄框架时也剔除了该倒转的年龄值。同理, HMD1502孔 1.7, m处, 其14C年龄比同一钻孔的 13.41, m的贝壳测年结果要偏老, 故剔除了该倒转的年龄值; 同时, 在该钻孔19.41, m、23.58, m 及29.95, m共3处的有机沉积物测年结果均比35.91, m的植物残片测年结果要偏老, 且比全新世的下限还要偏老, 结合本钻孔的岩性描述表明, HMD1502孔的全新统与晚更新统的界线在40, m附近, 故也剔除了这3份倒转的年龄值; 另外, 在6.01, m处的植物残片测年结果明显偏年轻, 因此, 在建立HMD1502孔全新世地层年龄框架时也剔除了该倒转的年龄值。
剔除了如上所述的倒转年龄值, 结合HMD1501和HMD1502两个钻孔的粒度分析结果, 即沉积物的粒度变化较小, 据此我们假设其沉积速率变化也较小, 通过线性内插建立了这2个钻孔全新世地层的年龄框架, 其具体地层深度与相对年龄详见图3。
HMD1501和HMD1502钻孔共鉴定出花粉和孢子85科属(图 4, 图5), 其中针叶类12属, 热带亚热带阔叶乔木和灌木类29科属, 温带阔叶乔木和灌木类13科属, 陆生草本类20科属, 蕨类孢子9科属, 以及苔藓孢子2属。具体孢粉类型及生态学划分(吴征镒, 1980)详见表2。
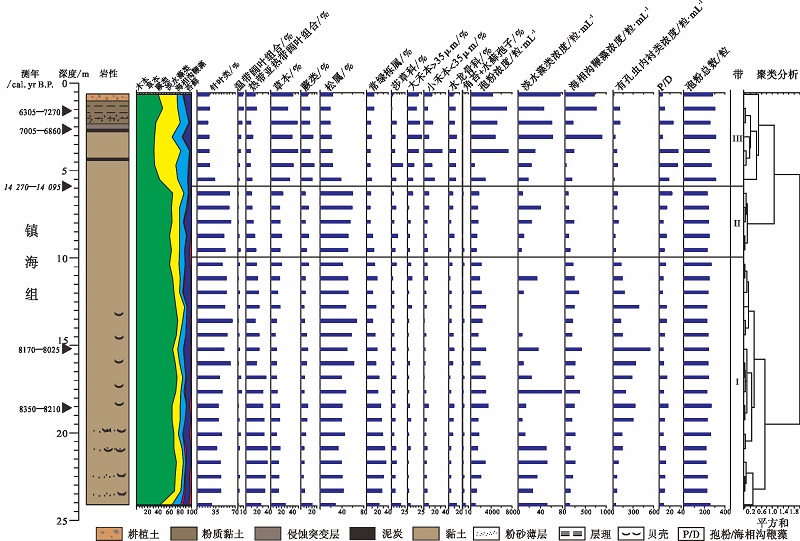 | 图4 浙江余姚田螺山HMD1501孔主要孢粉型的百分比及浓度图 斜体字表示异常年龄值Fig.4 Percentage and concentration diagram of major palynomorphs in Borehole HMD1501 at Tianluoshan of Yuyao, Zhejiang Province |
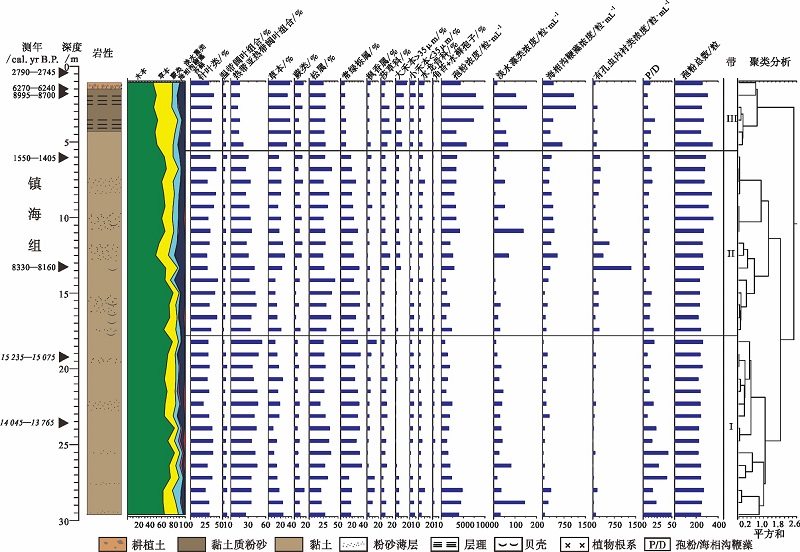 | 图5 浙江余姚田螺山HMD1502孔主要孢粉型的百分比及浓度图 斜体字表示异常年龄值Fig.5 Percentage and concentration diagram of major palynomorphs in Borehole HMD1502 at Tianluoshan of Yuyao, Zhejiang Province |
| 表2 浙江余姚田螺山HMD1501和HMD1502钻孔孢粉类型及生态学划分(划分据吴征镒, 1980) Table2 Pollen taxa and ecological grouping in Boreholes HMD1501 and HMD1502 at Tianluoshan of Yuyao, Zhejiang Province(ecological grouping is based on Wu, 1980) |
3.2.1 HMD1501孔孢粉组合特征 HMD1501孔共鉴定出花粉和孢子80科属(图 4)。其中针叶类11属, 热带亚热带阔叶乔木和灌木类25科属, 温带阔叶乔木和灌木类14科属, 陆生草本类19科属, 蕨类孢子9科属, 以及苔藓孢子2属。孢粉组合整体上以木本为主, 主要属种为松属(Pinus)和常绿栎属(Quercus-E)。草本次之, 主要为禾本科(Poaceae)和莎草科(Cyperaceae)。
此外, 蕨类主要以水龙骨科(Polypodiaceae)为主(图 4)。本钻孔还保存着大量的海相沟鞭藻和有孔虫内衬(其成分与孢粉壁类似, 可以在地层中很好地保存下来), 以及少量的淡水藻类。
依据HMD1501孔孢粉和藻类的百分比及浓度变化特征将其分为3个孢粉组合带— — 带 Ⅰ 、带Ⅱ 、 带Ⅲ (图 4), 由下至上描述如下:
带Ⅰ :~8970— 7631 cal. yr B.P.(24.01~9.81, m)
本带以木本为主, 含量为 77%(平均值, 下同), 其中针叶含量为47%, 主要为松属(45%), 热带亚热带阔叶含量较高(28%), 主要为常绿栎属(22%), 温带阔叶含量较低(2.4%)。草本含量为 14%, 主要为禾本科(7.1%)和莎草科(6.8%)。蕨类含量最低, 为 8.9%, 主要为水龙骨科(5.7%)和芒萁属Dicranopteris(1.7%)。
本带的孢粉浓度为2091粒/mL。
带Ⅱ :~7631— 7235 cal. yr B.P.(5.81, m~9.81)
本带仍以木本为主, 其含量略有下降, 为71%, 其中针叶含量略有增加, 为56%, 主要为松属(55%), 热带亚热带阔叶含量下降明显(13%), 主要为常绿栎属(8.9%), 温带阔叶含量变化不大(1.7%)。草本含量有所增加, 为18.5%, 主要为禾本科(10.9%)和莎草科(7.1%)。蕨类含量略有增加, 为10.5%, 主要为水龙骨科(6.2%)和芒萁属(2.7%)。
本带的孢粉浓度下降明显, 为1351粒/mL。
带Ⅲ :~7235— 5633 cal. yr B.P.(5.81~0.61, m)
本带木本含量急剧下降, 其含量甚至与草本含量相当, 分别为41.3%和40.6%, 其中针叶含量为23.8%, 主要为松属(23.2%), 热带亚热带阔叶含量持续下降(11.8%), 主要为常绿栎属(9.1%), 温带阔叶含量有所增加(5.8%)。草本含量为 40.6%, 主要为禾本科(33.1%)和莎草科(7.9%)。蕨类含量有所增加, 为18.2%, 主要为水龙骨科(11.7%)和芒萁属(2.8%)。
本带的孢粉浓度显著增加, 为4567粒/mL。
3.2.2 HMD1502孔孢粉组合特征 HMD1502孔共鉴定出花粉和孢子84科属(图 5)。其中针叶类12属, 热带亚热带阔叶乔木和灌木类28科属, 温带阔叶乔木和灌木类13科属, 陆生草本类20科属, 蕨类孢子9科属, 以及苔藓孢子2属。孢粉组合整体上以木本为主, 主要属种为松属、常绿栎属和枫香属(Liquidambar)。草本次之, 主要为禾本科和莎草科。蕨类主要以水龙骨科为主(图 5)。
此外, 本钻孔也保存了大量的海相沟鞭藻和有孔虫内衬, 并伴有少量的淡水藻类。
依据HMD1502孔孢粉和藻类组合特征将其分为3个孢粉组合带— — 带Ⅰ 、带Ⅱ 、带Ⅲ (图 5), 由下至上描述如下:
带Ⅰ :~10, 589— 7644 cal.yr B.P.(29.61~18.01, m)
本带以木本为主, 含量为 77%, 其中针叶含量为31.4%, 主要为松属(27.7%), 热带亚热带阔叶含量较高(40%), 主要为常绿栎属(26.6%), 温带阔叶含量较低(5.2%)。草本含量为 16.4%, 主要为禾本科(7.3%)和莎草科(6.9%)。蕨类含量最低, 为6.9%, 主要为水龙骨科(4.2%)和芒萁属(1.3%)。
本带的孢粉浓度为2137粒/mL。
带Ⅱ :~7644— 6773 cal.yr B.P.(18.01~5.51, m)
本带仍以木本为主, 其含量略有下降, 为72.8%, 其中针叶含量略有增加, 为35.9%, 主要为松属(31.1%), 热带亚热带阔叶含量有所下降(32.7%), 主要为常绿栎属(24%), 温带阔叶含量略有下降(4.2%)。草本和蕨类含量略有增加, 分别为18.8%和8.5%, 主要为禾本科(8.2%), 莎草科(7.6%), 水龙骨科(5.1%), 凤尾蕨属Pteris(1.3%)和芒萁属(1.04%)。
本带的孢粉浓度略有增加, 为2562粒/mL。
带Ⅲ :~6773— 3327 cal. yr B.P.(5.51~0.61, m)
本带木本含量急剧下降, 为54%, 其中针叶含量略有下降, 为34.8%, 主要为松属(27.1%), 热带亚热带阔叶含量持续下降(14.6%), 主要为常绿栎属(7.5%)和枫香属(4.3%), 温带阔叶含量略有增加(4.6%)。草本含量急剧增加, 达35.3%, 主要为禾本科(19.6%)和莎草科(11.2%)。蕨类含量有所增加, 为10.7%, 主要为水龙骨科(6.7%)和芒萁属(1.6%)。
本带的孢粉浓度显著增加, 为6372粒/mL。
3.3.1 HMD1501孔地层划分 依据岩性和孢粉、海相沟鞭藻类浓度, 并结合AMS14C测年数据, 将HMD1501孔上部0.61~24.01, m岩心分为4个层位(图 6), 从下到上依次为:
1)24.01~11.21, m: 浅海相沉积
褐灰色黏土为主, 时有粉砂团块或薄层出现, 具层理, 含丰富的贝壳碎片。平均粒径为5.26~7.37φ , 黏土、粉砂和砂含量分别为16.19%(平均值, 下同), 78.17%和5.66%。孢粉和沟鞭藻浓度偏低, 分别为: 2123粒/mL 和240粒/mL。两个采样点(15.11, m和18.51, m)的AMS14C年龄分别为8170~8025 yr B.P. 和8350~8210 yr B.P.。
2)11.21~1.01, m: 滨岸湖沼相沉积
分别在2.57~2.77, m和4.20~4.35, m处出现两层黑色泥炭, 中间和底部为褐灰色黏土, 顶部为深灰色黏土/粉质黏土, 具微弱或明显水平层理。黏土含量略有降低, 为13.63%, 粉砂和砂含量略有增加, 分别为79.22%和7.14%, 平均粒径为5.76~7.56φ 。孢粉和沟鞭藻浓度均迅速增加, 分别为: 4203粒/mL和346粒/mL。3个采样点(5.9, m, 2.67, m和1.61, m)的AMS14C年龄为14, 270~14, 095 yr B.P., 7005~6880 yr B.P. 和6305— 6270 yr B.P.。
3)1.01~0.61, m: 耕作层
深灰色耕植土为主, 出现大量暗褐色斑点(可能与现代植物根系有关), 较致密。黏土和粉砂含量略有降低, 分别为13.22%和75.82%, 砂含量略有增加, 且波动较大, 为11.11%, 平均粒径为5.83~6.90 Ф 。孢粉和沟鞭藻浓度均有所增加, 分别为4258粒/mL和712粒/mL。
3.3.2 HMD1502孔地层划分 依据岩性和孢粉、沟鞭藻浓度, 并结合AMS14C测年数据, 将HMD1502孔上部1.11~29.61, m岩心分为4个层位(图 7), 从下到上依次为:
1)29.61~18.71, m: 潮下带沉积
褐灰色粉砂质黏土为主, 出现粉砂薄层或团块。平均粒径为6.03~7.67 Ф , 黏土、粉砂和砂含量分别为18.80%, 76.83%和4.41%。孢粉和沟鞭藻浓度偏低, 分别为: 2087粒/mL 和130粒/mL。两个采样点(23.58, m和19.41, m)的AMS14C年龄分别为, 14, 045~13, 765 yr B.P.(舍去)和15, 235~15, 075 yr B.P.(舍去)。
2)18.71~4.41, m: 浅海相沉积
以褐灰色粉砂质黏土为主, 常见粉砂薄层或团块, 往往具层理, 多处可见贝壳碎片。黏土含量变化不大, 为18.04%, 粉砂含量略有增加, 为79.12%, 砂含量略有降低, 且波动较大, 为2.92%, 平均粒径为5.69~7.99 Ф 。孢粉和沟鞭藻浓度均有增加, 分别为: 2644粒/mL 和275粒/mL。 两个采样点(13.41, m和6.01, m)的AMS14C年龄为8330~8160 yr B.P. 和1550~1405 yr B.P.(舍去)。
3)4.41~1.51, m: 滨岸湖沼相沉积
黏土质粉砂, 上部以褐灰色黏土质粉砂为主, 水平层理发育; 下部为褐灰色黏土与粉砂互层, 粉砂层层理发育。黏土含量略有下降, 为15.38%, 粉砂含量略有增加, 为84.56%, 砂含量急剧下降, 为0.06%, 平均粒径为6.34~7.78 Ф 。
孢粉和沟鞭藻浓度持续增加, 分别为: 7047粒/mL 和718粒/mL。一个采样点(1.7, m)的AMS14C年龄为8995~8700 yr B.P.(舍去)。
4)1.51~0.41, m: 耕作层
深灰色粉质黏土, 具土壤化特征, 含植物根系、炭屑, 并在0.42~0.47, m处出现大块泥炭。黏土含量变化不大, 但波动较大, 为15.33%, 粉砂和砂含量变化不大, 分别为84.63%和0.06%, 平均粒径为6.41~7.58 Ф 。孢粉和沟鞭藻浓度均有降低, 分别为: 4306粒/mL 和374粒/mL。两个采样点(1.5, m和0.44, m)的AMS14C年龄分别为6270~6240 yr B.P. 和2790~2745 yr B.P.。
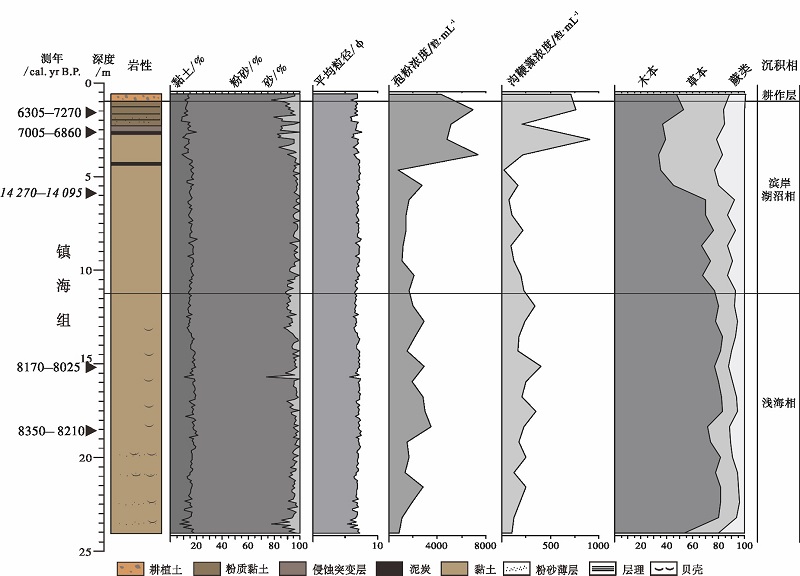 | 图6 基于岩性、粒度、孢粉、沟鞭藻浓度的浙江余姚田螺山HMD1501孔沉积相划分 斜体字表示异常年龄值Fig.6 Sedimentary facies division based on lithology, grain size, pollen and dinoflagellates concentration of Borehole HMD1501 at Tianluoshan of Yuyao, Zhejiang Province |
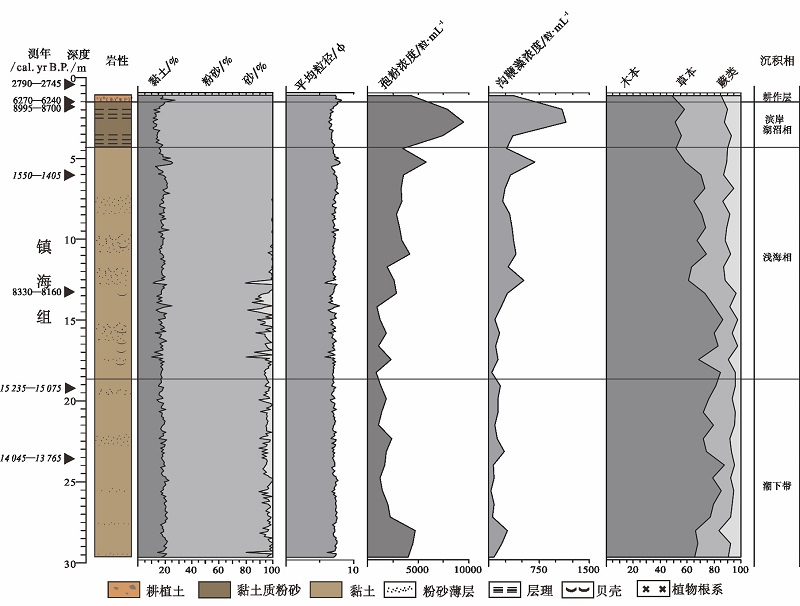 | 图7 基于岩性、粒度、孢粉、沟鞭藻浓度的浙江余姚田螺山HMD1502孔沉积相划分 斜体字表示异常年龄值Fig.7 Sedimentary facies division based on lithology, grain size, pollen and dinoflagellates concentration of Borehole HMD1502 at Tianluoshan of Yuyao, Zhejiang Province |
HMD1501和HMD1502两个钻孔的孢粉结果表明, 研究区全新世以来气候环境与植被形态存在着较大的波动, 即早期相对稳定而中后期存在着剧烈的波动。
4.1.1 早全新世 对应于HMD1501和HMD1502两个钻孔的带 Ⅰ , 根据测年结果(表 1; 图3), 推测年代分别为:~8970— 7631 cal.yr B.P.和~10, 589— 7644 cal. yr B.P., 此时研究区刚刚经历晚更新世“ 新仙女木” 事件之后进入全新世, 气候开始迅速转暖变湿, 处于气候波动上升的早全新世前北方期和北方期。
孢粉记录显示, 针叶类(主要为松属; 47%和31.4%)和热带亚热带阔叶类(28%和40%)的含量最高, 草本花粉(14%和16.4%)和蕨类孢子(8.9%和6.9%)的含量次之, 温带阔叶类(2.4%和5.2%)含量最低(图 4, 图5)。显示当时研究区及附件地区的植被群落属于常绿阔叶和针叶混交林, 松和常绿栎为优势种, 林下天然生长的禾本科(< 35, μ m)植物为草本群落的优势物种, 而在地势低洼处, 莎草、香蒲等湿生草本植物和喜阴湿的水龙骨、芒萁、凤尾蕨等蕨类植物也有分布, 说明气候开始转为温暖湿润。
4.1.2 中全新世 对应于HMD1501和HMD1502两个钻孔的带Ⅱ , 根据测年结果(表 1; 图3), 推测年代分别为:~7631— 7235 cal. yr B.P. 和 ~7644— 6773 cal. yr B.P., 处于中全新世大西洋期。
孢粉记录显示, 针叶类(56%和35.9%)含量略有增加, 热带亚热带阔叶类(13%和32.7)和温带阔叶类(1.7%和4.2%)的含量均有下降, 草本花粉(18.5%和18.8%)和蕨类孢子(10.5%和8.5%)的含量均有增加(图 4, 图5)。表明当时研究区及附件地区的植被群落虽仍为针阔叶混交林, 但是阔叶木本植物开始下降, 反之, 草本和蕨类的含量均开始有增加。说明其在林下或低洼处已经开始小规模的分布一些草地。特别是之前的建群优势种常绿栎属的含量急剧下降, 而草本含量, 尤其是具有栽培性的禾本科植物(花粉粒径大于等于35, μ m)含量的增加, 反映出人类活动已经开始影响植被的群落结构, 气候开始转为温和湿润。
4.1.3 晚全新世 对应于HMD1501和HMD1502两个钻孔的带 Ⅲ , 根据测年结果(表 1; 图3), 推测年代分别为:~7235— 5633 cal. yr B.P.和~6773— 3327 cal. yr B.P., 处于晚全新世亚北方期。
孢粉记录显示, 针叶类(23.8%和34.8%)和热带亚热带阔叶类(11.8%和14.6%)含量均有下降, 温带阔叶类(5.8%和4.6%)和蕨类孢子(18.2%和10.7%)的含量略有增加, 草本花粉(40.6%和35.3%)的含量急剧增加(图 4, 图5)。喜干的草本花粉, 尤其是禾本科的含量急剧增加, 而喜暖湿的热带亚热带阔叶成分持续下降, 温带阔叶成分略有增加, 反映出在人类活动的进一步影响下, 当时植被群落中的草地也随之扩大, 并在滨海地区开始由一些以莎草科为主的沼泽地形成, 植被群属于针阔叶混交林— 草地, 气候转为凉爽干燥。
海平面升降所引起的风暴潮、洪涝威胁和盐水入侵等方面的变化, 直接影响着全新世时期的沿海地区古人类的生存环境。中国东部陆架的海平面变化不仅受到全球冰量的控制, 也会受到区域构造运动的影响, 但是, 纵观过去中国东部的海平面研究, 由于严重缺乏全球背景, 加上测年的不足, 使得各种曲线之间很难进行对比(李从先和闵秋宝, 1981; Chen and Stanley, 1998; Hori et al., 2001; 朱诚等, 2003; Zong, 2004; 史威等, 2008; Chen et al., 2008; 李从先和范代读, 2009; Song et al., 2013)。田螺山HMD1501和HMD1502钻孔地处沿海地区, 较好地保存了沟鞭藻类及有孔虫内衬等海相化石, 可以进行海平面变迁研究, 为中国东部全新世以来的海平面变迁研究提供一些化石证据。
沟鞭藻类绝大多数生长在海洋里, 因此, 在进行HMD1501和HMD1502两个钻孔研究时, 依据其浓度以及P/D(孢粉总数/海相沟鞭藻总数)的比值, 可以反映离岸距离, 来恢复和重建故海岸线、海侵和海退事件(Morzadec-Kerfourn, 2005; van Soelen et al., 2010)。一般来说, P/D值高时, 则反映离岸距离较近, 属于海退时期; 反之, P/D值低时, 则反映离岸距离较远, 属于海侵时期。同海相沟鞭藻类一样, 有孔虫也是生活在海洋里, 是进行古海洋研究的一个重要的指标。本研究中, 由于提取孢粉及藻类的实验过程中用到盐酸和氢氟酸, 钙质及硅质的有孔虫壳体会被反应掉, 但是有孔虫内部的有机质部分— — 有孔虫内衬, 却能很好地保存下来, 因此, 可以综合海相沟鞭藻化石及有孔虫内衬的浓度变化来判断海侵与海退事件。
纵观田螺山HMD1501和HMD1502两个钻孔, 海相沟鞭藻类和有孔虫内衬几乎出现在每一份样品中, 这可能与研究区靠近海边的缘故, 由于一次异常的天气, 如台风、风暴潮等, 都可能将海水带上海岸, 从而, 在地层中留下了部分海相化石。此外, 也与剖面的分辨率有关, 本研究是0.8, m间隔采样, 其较低的分辨率也可能漏掉一些没有海相化石的层位。但是, 仍可以通过海相沟鞭藻类及有孔虫内衬的多寡来判断离岸的远近, 进而推断海平面的升降。
4.2.1 早— 中全新世海侵期 对应于HMD1501和HMD1502两个钻孔的带Ⅰ 、带Ⅱ , 根据测年结果(表 1; 图3), 推测年代分别为:~8970— 7235 cal. yr B.P.和~10, 589— 6773 cal. yr B.P., 此时研究区刚刚经历晚更新世“ 新仙女木” 事件之后进入全新世, 气候开始迅速转暖变湿, 处于气候波动上升的早全新世前北方期、北方期和大西洋期。
HMD1501孔的P/D的平均值较低, 为11.4; 有孔虫内衬的浓度较高, 为126粒/mL; 淡水藻类的浓度较低, 为20.55粒/mL。HMD1502孔也有类似现象, 有孔虫内衬的浓度为175.5粒/mL, 淡水藻类的浓度为36.2粒/mL, P/D值稍高, 为15.55(图 4, 图5)。综合2个钻孔的记录来看, 研究区在早— 中全新世时期属于海侵期。此外, 淡水藻类的浓度较低, 也说明河流及湖泊的影响较小, 研究区主要受海水侵蚀的影响。
4.2.2 晚全新世海退期 对应于HMD1501和HMD1502两个钻孔的带 Ⅲ , 根据测年结果(表 1; 图3), 推测年代分别为:~7235— 5633 cal. yr B.P.和~6773— 3327 cal. yr B.P., 处于晚全新世亚北方期。
HMD1501孔的P/D的值有所增加, 达18.6; 有孔虫内衬明显降低, 为56粒/mL; 淡水藻类的浓度有所增加, 为45粒/mL。HMD1502孔也有类似现象, 有孔虫内衬的浓度下降明显, 为91.4粒/mL; 淡水藻类的浓度有所增加, 为70.6粒/m; P/D值略有降低, 为11.4(图 4, 图5)。综合2个钻孔的记录来看, 研究区在晚全新世时期属于海退期。此外, 淡水藻类的浓度的增加, 也说明河流及湖泊的影响开始增强。田螺山地区在晚全新世处于海退时期, 为古人类开展农业活动提供了广阔的场所, 进而孕育了灿烂的河姆渡— 田螺山文化。
人类活动对植被的变化有着非常深远的影响, 主要体现在乔木花粉的减少, 先锋植物花粉的增加, 栽培作物花粉大量出现等(Scaife, 1988; McAndrews and Boyko-Diakonow, 1989; Whittington et al., 1991)。此外, 郑卓等(2004)指出, 全新世蕨类孢子含量增高可能反映人类活动的增强。因此, 在本研究中, 就可以通过乔木花粉(如常绿栎属等)的减少, 以及草本花粉(尤其是花粉粒径大于等于35, μ m的禾本科栽培作物的花粉)和蕨类孢子的大量出现, 来指示全新世时期的人类活动。
随着河姆渡遗址中大量稻谷遗存及农具的发现, 引发了亚洲稻作起源及其传播的大讨论。此后, 中国报道有水稻遗存的新石器时代遗址大约有170处, 主要分布在长江中游和下游地区, 少数位于华南地区和黄河流域(裴安平和熊建华, 2004)。其中, 最引人注意的遗址多发现在长江三角洲地区, 伴随着一系列新石器早期文化遗址的发现, 提供了越来越多的水稻种植的直接或间接的证据, 例如, 浙江的河姆渡、罗家角、上山遗址; 太湖周围的草鞋山遗址; 上海的崧泽及昆山的绰墩等遗址均发现了水稻遗存(黄宣佩和张明华, 1980; 郑云飞等, 1994, 2001; 黄翡等, 1998; 萧家仪等, 2004; Zheng and Jiang, 2007), 从而将水稻种植的历史可能最早提前到 1万年以前(Jiang and Liu, 2006; 郑云飞等, 2007; Zheng and Jiang, 2007)。
在本研究中, 田螺山遗址的HMD1501和HMD1502两个钻孔的孢粉记录显示, 在全新统镇海组沉积晚期, 对应于晚全新世亚北方期, 年代约为:~7235— 3327 cal. yr B.P., 孢粉组合发生了较大的变化。其中, 最突出的变化就是草本花粉的含量激增, 特别是花粉粒径大于等于35, μ m的禾本科栽培作物花粉含量显著的增加。与此同时, 蕨类孢子的含量也有明显的增加。然而, 乔木花粉(如常绿栎属等)含量存在较明显的下降(图 4, 图5)。Ren和Beug(2002)提出, 木本植物花粉含量明显减少, 蕨类植物孢子含量增加, 可能与森林的衰退及农业文明有关, 先民们种植水稻时的开荒伐薪有关, 从而导致了森林面积进一步减少。
由于禾本科花粉的传播范围较小, 因此, 粒径大于等于35, μ m 的禾本科栽培作物花粉的大量增加可能指示水稻田的存在(Li et al., 2006)。在2004年田螺山遗址的考古现场, 发现了大量河姆渡文化时期的水稻遗存, 如稻谷壳及炭化米粒等遗物(浙江省文物考古研究所等, 2007), 将其与 HMD1501和HMD1502两个钻孔的孢粉结果相互印证, 证实在~7235— 3327 cal. yr B.P.期间古人类农业活动较频繁, 在河姆渡— 田螺山遗址附近存在较多的水稻种植区。
综合河姆渡— 田螺山遗址HMD1501和HMD1502两个钻孔的孢粉、藻类及有孔虫内衬等的记录, 恢复和重建了全新世以来田螺山地区的植被环境及海平面变迁, 具体如下:
1)在早全新世气候回暖期, 研究区的植被群落属于常绿阔叶和针叶混交林, 松和常绿栎为优势种, 林下天然生长的禾本科植物为草本群落的优势物种, 而在地势低洼处, 莎草、香蒲等湿生草本植物和喜阴湿的水龙骨、芒萁、凤尾蕨等蕨类植物也有分布, 说明气候开始转暖。
2)在中全新世大西洋期阶段, 研究区的植被群落虽仍为针阔叶混交林, 但是阔叶木本植物开始下降, 反之, 草本和蕨类的含量均开始有增加。说明其在林下或低洼处已经开始小规模的分布一些草地。特别是之前的建群优势种常绿栎属的含量急剧下降, 而草本含量, 尤其是具有栽培性的禾本科植物含量的增加, 反映出人类活动已经开始影响植被的群落结构, 气候仍保持温和湿润。
3)在晚全新世亚北方期阶段, 喜干的草本花粉, 尤其是禾本科的含量急剧增加, 而喜暖湿的热带亚热带阔叶成分持续下降, 温带阔叶成分略有增加, 反映在人类活动的进一步影响下, 当时植被群落中的草地也随之扩大, 并在滨海地区开始由一些以莎草科为主的沼泽地形成, 植被群属于针阔叶混交林— 草地, 气候转为凉爽干燥。
4)综合海相沟鞭藻、有孔虫内衬及 P/D记录来看, 研究区在早— 中全新世时期属于海侵时期, 晚全新世时期属于海退时期。田螺山地区在晚全新世时期处于海退时期, 为古人类开展农业活动提供了广阔的场所, 进而孕育了灿烂的河姆渡— 田螺山文化。
5)HMD1501和HMD1502两个钻孔的孢粉资料均显示, 在全新世晚期, 草本花粉的含量激增, 特别是一些大于等于35 μ m的禾本科栽培作物花粉的含量增加, 与此同时, 蕨类孢子的含量也有所增加, 然而, 木本植物花粉含量明显减少, 结合田螺山遗址的考古遗存资料, 证实在~7235— 3327 cal. yr B.P.期间古人类农业活动较频繁, 在河姆渡— 田螺山遗址附近存在较多的水稻种植区。
(责任编辑 李新坡)
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|