






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
华北中元古代鱼骨状方解石: 成因机制和古环境意义*
[汤冬杰1, 2  , 史晓颖
, 史晓颖1, 3 , 张文浩4 , 刘云3 , 吴金键3 ]
, 史晓颖|
|

鱼骨状方解石是一种特殊的碳酸盐沉积,由锯齿状亮暗交互的亚毫米级条带组成,主要见于太古宙。以往认为,鱼骨状方解石属无机化学沉淀成因,形成于水体缺氧、碳酸盐过饱和、富Fe2+、Mn2+等碳酸盐沉淀抑制剂的环境条件;在地质记录中其丰度随时间的减少反映了海洋的长期氧化趋势。文中首次在华北地台中元古界高于庄组四段微生物礁内发现了大量鱼骨状方解石。宏观观察表明,这些鱼骨状方解石主要以微生物礁孔洞充填物形式产出,明显区别于太古宙以海底沉淀形式直接产出在海底的鱼骨状方解石。显微研究发现,鱼骨状方解石晶体纤维具有沿晶体生长方向旋转消光特征,证明其内部亚晶的光学C轴从纤维底部的随机排列逐步旋转至上部垂直纤维生长方向。这符合球状晶体生长模式,需要方解石沉淀抑制剂的参与。鱼骨状方解石产出具有丰度随时间减少以及产出形式由海底沉淀向孔洞胶结物转变的特征。这些特征与海洋氧化逐渐增强以及具氧化还原敏感属性的碳酸盐沉淀抑制剂逐渐从水体中移除相吻合。笔者认为鱼骨状方解石的沉淀抑制剂为Fe2+和Mn2+,这与微生物岩无明显Ce异常和Fe2+极强的抑制能力相一致。因此,鱼骨状方解石可用于指示缺氧环境条件。此外,显微和超微研究也表明鱼骨状方解石晶体内存在有大量与其生长方向一致的菌丝体残余和与之密切伴生的有机矿物,表明微生物为鱼骨状方解石成核和初始沉淀提供了重要垫板。
About the first author Tang Dongjie,born in 1985,is an associate professor of China University of Geosciences(Beijing). He is engaged in geobiology and Precambrian geology. E-mail: dongjtang@126.com.
第一作者简介 汤冬杰,男,1985年生,中国地质大学(北京)副教授,主要从事地球生物学和前寒武纪地质研究工作。E-mail: dongjtang@126.com。
Herringbone calcite(HC),characterized by the alternation of sub-millimetric light and dark jagged bands,is a special type of carbonate mineral phase that mainly developed during the Archean. Early studies suggest that HC is a kind of inorganic chemical precipitation,formed in anoxic,calcium carbonate supersaturated,and precipitation inhibitor (Fe2+,Mn2+) ̄rich seawater. Therefore,it has been used as an indicator of anoxic seawater conditions,and its abundance decreases over geologic time is thought to have reflected the increasing oxidation of the ocean. However,recent studies suggest that the genesis of HC might have been variable. For the first time,we have found HC in the microbial reef of the Mesoproterozoic Gaoyuzhuang Formation in the North China Platform. Macroscopically,these HCs mainly occurred in the voids of microbial reef as void-filling,distinct from their Archean counterparts which largely occurred as seafloor precipitation. Microscopically,the HC fiber shows a characteristic of rotated distinction along the crystal growth direction,indicating that the internal crystals of each HC fiber was once rotated from the bottom to the top. This is in accordance with the spherulitic growth pattern,therefore,requiring the participation of the calcite precipitation inhibitors. The secular decrease in HC abundance and the changes in their occurrence pattern from seafloor precipitation to void-filling,suggest that HC precipitation inhibitors should be redox-sensitive elements,which have been continuously removed from seawater with the oxidation of the ocean. Thus,we think that the major inhibitors for HC precipitation are Fe2+ and Mn2+,and HC can be used as a mineral indicator for anoxic environment,which is consistent with the Ce anomaly results(without obvious anomalies)in the microbial reef and the presence of strong inhibitor effect of Fe2+. In addition,both microscopic and ultramicroscopic observations revealed that there are a large number of bacterial filamentous relics and closely associated organominerals concentrated along the axis of HC fibers,indicating that the microbes have provided favorable sites for the initial nucleation and subsequent growth of HC crystals.
鱼骨状方解石(herringbone calcite, 简写为HC)由锯齿状亮暗交互的亚毫米级条带组成, 其独特的矿物学特征被认为是由内部纤维状晶体所含亚晶的光学轴从底到顶逐渐旋转所导致(Sumner and Grotzinger, 1996a)。鱼骨状方解石多见于太古宙, 少量见于古元古代, 主要以海底沉淀形式广泛产出; 但在此后, 鱼骨状方解石丰度显著减少, 仅在新元古代和显生宙个别时段偶见以孔洞充填形式出现, 而在中元古代早期迄今没有报道(Sumner and Grotzinger, 1996a, 1996b; Grotzinger and James, 2000)。鱼骨状方解石这种长期演变, 以及要求缺氧条件下Fe2+、Mn2+等抑制剂参与的特征(Sumner and Grotzinger, 1996a), 已被用作反映海水氧化还原条件演变的重要证据(Sumner and Grotzinger, 1996b)。碳酸盐沉淀抑制剂可吸附在晶体生长面, 从而降低碳酸盐沉淀速率并改变晶体形态, 也可通过提高颗粒表面自由能来抑制成核(Sumner and Grotzinger, 1996b)。鱼骨状方解石被认为是抑制剂改变晶体形成的典型代表(Sumner and Grotzinger, 1996b)。笔者发现北京延庆高于庄组四段微生物礁的孔洞充填物主要为鱼骨状方解石, 这为研究其成因机制和古海洋环境条件提供了重要材料。
有关鱼骨状方解石的成因争议很大。晶体光学观察研究表明, 鱼骨状方解石内的亚晶由下至上逐渐旋转并最终大体平行于晶体长轴方向, 因此认为其生长模式反映了沉淀抑制剂导致的球状晶体沉淀(spherulitic precipitation)模式(Keith and Padden, 1963; Sumner and Grotzinger, 1996a)。由于鱼骨状方解石主要形成于太古宙至古元古代的缺氧海洋条件下, 因此推测Fe2+和/或Mn2+是导致鱼骨状方解石沉淀的主要抑制剂(Sumner and Grotzinger, 1996a, 1996b), 而其产出丰度和形式的长期演变被认为是海水氧化导致这些抑制剂从海水中移除的结果(Sumner and Grotzinger, 1996a, 1996b)。但通过对显生宙个别鱼骨状方解石的研究, 有学者认为除Fe2+、Mn2+这2种抑制剂外, Mg2+和/或Sr2+等碳酸盐沉淀抑制剂也能导致鱼骨状方解石的形成(Tourre and Sumner, 1999)。甚至有人认为, 即使没有抑制剂参与, 仅缺氧条件也可导致鱼骨状方解石形成(de Wet et al., 2004)。这就从机理上对Summer和Grotzinger(1996a)提出的鱼骨状方解石的球状晶体生长模式(要求沉淀抑制剂参与)提出了质疑。
由此可见, 虽然鱼骨状方解石是一种潜在的古氧相指示矿物, 但将其应用于古氧相分析的前提是确定其沉积机制及其所需抑制剂类型。为确定鱼骨状方解石的成因模式并分析其古环境意义, 文中以延庆高于庄组四段微生物礁内的鱼骨状方解石为研究对象, 进行晶体光学显微观察、碳同位素和主微量元素测试等综合研究。
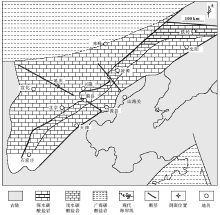
研究区位于北京延庆北部(图1), 属华北克拉通北缘燕辽裂陷槽中段。自古元古代末期(约1800, Ma)吕梁运动导致华北最终克拉通化之后, 其构造体制发生了根本性变化, 从此前的碰撞造山转化为大规模的陆内裂谷阶段(Wan et al., 2003)。在古元古代末, 响应于Columbia超大陆裂解, 华北克拉通在北东缘形成了燕辽裂谷。其内自下而上沉积了厚达万米的长城系(1800— 1600, Ma, Pt1)、蓟县系(1600— 1400, Ma, Pt2)和青白口系(1000— 800, Ma, Pt3), 在蓟县系和青白口系之间存在约400, Ma的沉积间断(高林志等, 2009)。
 | 图1 华北地台中元古代高于庄期古地理背景(据Tang et al., 2013a, 有修改)Fig.1 Simplified palaeogeographic map of the Mesoproterozoic Gaoyuzhuang stage in North China Platform(modified from Tang et al., 2013a) |
延庆高于庄组上部(第三段)凝灰岩的高精度锆石SHRIMP U-Pb年龄(1559± 12, Ma)和锆石LA-MC-ICP-MS年龄(1560± 5, Ma)很好地限定了该组的沉积时限(李怀坤等, 2010)。据下伏大红峪组中上部火山岩夹层的锆石U-Pb年龄1622— 1625, Ma(陆松年和李惠民, 1991; Lu et al., 2008), 可将高于庄组底界年龄限定为约1600, Ma, 因此整个高于庄组的沉积时限大致为1600— 1530, Ma(李怀坤等, 2010)。
华北地台中部的高于庄组自下而上可划分4个亚组(或段)( 图 2): 官地亚组(高一段)、桑树鞍亚组(高二段)、张家峪亚组(高三段)和环秀寺亚组(高四段)。在研究区内, 高一段主要为浅— 深灰色中厚层含燧石条带白云岩夹薄层微生物席白云岩, 底部有含砂质云岩; 其中叠层石较为发育, 局部形成叠层石礁。高二段主要由中厚层白云岩及薄层钙质、泥质白云岩组成, 下部发育含锰页岩, 上部白云岩中由微生物席诱发的层面构造特别发育。高三段由深灰色中厚层白云岩和薄板状白云质灰岩组成, 并发育有大量微生物席纹层、凝块石和结核(Mei, 2008), 上部夹有较多的暗色页岩薄层。高四段由浅灰色中厚层白云岩及块状微生物礁组成, 主要形成于潮下带环境(Mei, 2007; Tang et al., 2016)。
 | 图2 华北中元古界蓟县系和延庆高于庄组地层序列Fig.2 Lithological columns of the Mesoproterozoic Jixian System in North China and Gaoyuzhuang Formation in Yanqing County |
文中研究的样品采自北京延庆干沟、八道河以及下马鹿沟村; 研究的标本80余块, 磨制光面10个、探针片100余张。宏观特征主要基于野外露头和抛光光面观察; 低倍显微特征观察使用Zeiss Discovery V20型立体显微镜; 高倍显微以及晶体光学特征观察使用Zeiss Scope A1型偏光显微镜; 超微组构观察使用场发射扫描电镜(FESEM)。为避免污染, 扫描分析之前对样品进行了去离子水超声波清洗; 为增强样品导电性和图像质量, 样品表面进行了喷Pt处理, 涂层厚约10, nm。FESEM型号为Zeiss Supra 55, 工作距离5~15, mm, 加速电压10~20 kV, 电子图像拍摄探头为Inlens。
微量元素测试使用Bruker Aurora M90型电感耦合等离子体质谱仪(ICP-MS)。为避免陆源碎屑对碳酸盐岩矿物稀土浓度的影响, 样品预处理使用了5%的乙酸。类似方法已经成功应用于多例碳酸盐岩稀土元素分析(e.g., Nothdurft et al., 2004; Ling et al., 2013; Tang et al., 2016)。样品前处理:先将50, mg样品粉末加入到50, mL的离心管内, 然后加入大约40, mL的蒸馏水和500 ng的内标物Rh, 再加入2.5, mL的乙酸(纯度为98%, 使用亚沸蒸馏法进行提纯)用于溶解碳酸盐矿物。待反应完成后, 将溶液配备至50, mL。将离心管封闭后震荡约1 h, 然后以3000 r/min的转速离心3, min。离心管内上部的溶液用于微量元素的测试。为评估陆源碎屑对稀土配分模式的影响程度, 部分样品采用了传统的HNO3/HF溶样方法将样品完全溶解后再做测试, 具体操作流程同Guo等(2013)。绝大部分元素的测试精度优于± (5~10)%。测试在中国科学院地球化学研究所完成。主量元素测试使用Spectro公司的EDXRF手持元素分析仪, 测量误差小于2%。
无机碳氧同位素分析在内华达大学地球科学系同位素实验室完成。将50~100, μ g的碳酸盐粉末置于自动反应器(Kiel-Device)中与100%的磷酸反应约10, min, 反应温度为70, ℃, 反应所产生的气体自动输入到质谱仪(Finnigan Delta Plus)中进行同位素分析, 分析结果采用Vienna-PeeDee belemnite(VPDB)标准。内部标样的C和O同位素误差小于0.05‰ 。
鱼骨状方解石是延庆高四段微生物礁内的主要胶结物, 其体积百分含量最高可达15%(图 3-A至3-F)。近距离观察显示, 鱼骨状方解石呈锯齿状亚毫米级别亮暗交互条带(图 3-G)。其产出形式主要有2种: (1)沿锥叠层石的轴或轴两侧的微生物席产出, 明显受控于叠层石形态(图 3-A, 3-B); 局部放大可见沿叠层石纹层分布的鱼骨状方解石, 厚毫米至厘米级不等, 横向断续延伸, 并可截断叠层石纹层(图 3-B)。(2)呈斑块状或不规则角砾状产出, 具破碎沉积特征(图 3-C, 3-D, 3-E); 以这种形式产出的鱼骨状方解石排列很不规则( 图 3-D, 3-E, 3-F), 部分呈棱角状(图 3-E), 推测为微生物礁原生组构的破碎产物(图 3-D, 3-F), 或破碎后再胶结的产物(图 3-E)。
 | 图3 北京延庆高于庄组四段鱼骨状方解石宏观特征Fig.3 Macroscopic characteristics of herringbone calcite(HC) of the Member 4 of Gaoyuzhuang Formation in Yanqing County, Beijing |
延庆高四段鱼骨状方解石的这2种产出方式与加拿大中元古界Dismal Lakes群锥叠层石礁体内鱼骨状方解石的产出特征一致(Bartley et al., 2015)。一般认为, 这种叠层石的轴部微生物席向上生长速率较快, 从而疏松多孔; 轴周边的微生物席虽然生长相对较慢, 但由于受到水的浮力和轴部快速生长微生物席的拖拽也多原始孔洞, 从而为第1种形式产出的鱼骨状方解石提供了大量空间(Bartley et al., 2015)。由于这种微生物岩多孔的特征也导致其容易破碎, 从而出现第2种形式产出的鱼骨状方解石(Bartley et al., 2015)。
微生物礁内的孔洞主要被鱼骨状方解石充填胶结, 如叠层状凝块石柱间(图 4-A)以及其他不规则的空间内(图 4-B)均被鱼骨状方解石充填。有些较小的孔洞可完全被鱼骨状方解石充填, 孔洞边缘的鱼骨状方解石呈纤维状垂直于孔洞边缘整齐排列, 但孔洞内部的鱼骨状方解石杂乱排列, 且纤维变短加粗(图 4-C, 4-D)。有些较大的孔洞不能完全被鱼骨状方解石充填, 孔洞的核心主要为自形的白云石(图 4-A)。
 | 图4 北京延庆高于庄组四段鱼骨状方解石显微特征Fig.4 Micrographic characteristics of herringbone calcite(HC)of the Member 4 of Gaoyuzhuang Formation in Yanqing County, Beijing |
鱼骨状方解石纤维体呈锯齿状平行排列, 厚0.5~1, mm(图 4-E, 4-F)。其消光特征独特, 正交偏光下条带下部不消光, 由下至上逐渐转变为全消光(图 4-F)。将偏振光系统逆时针旋转45° , 消光位置也由晶体上部转移至下部(图 4-G, 4-H)。这种旋转消光特征, 表明鱼骨状方解石下部亚晶的光学C轴具有随机排列特征, 但沿晶体生长方向, 亚晶的C轴会逐渐旋转直至大体垂直于晶体的生长方向。
通常认为鱼骨状方解石具有无机化学沉淀成因, 其内部不太可能保存微生物组构(Sumner and Grotzinger, 1996a)。但在高于庄组鱼骨状方解石内保存有微生物丝状体残余。这些丝状体不仅数量多, 而且主要集中在晶体轴部, 呈规律性分布(图 5)。在早期硅化的鱼骨状方解石内可见大量保存较好的微生物丝状体(图 5-A, 5-B)。通过白卡技术(Folk, 1987)可以清晰显示丝状体轮廓(凸显有机质部分), 其直径主要集中在20, μ m, 长数百微米(图 5-C)。未被硅化的鱼骨状方解石纤维内的丝状体沿晶体生长方向展布, 但保存较差, 直径约20, μ m的丝状体轮廓仅隐约可见。高倍显微观察显示这种级别的丝状体内可见直径约0.6, μ m、长数微米至几十微米不等的次级微小丝状体, 这些微小丝状体也沿晶体生长方向分布( 图 5-D至5-J), 推测可能属细菌丝状体的鞘和胞外聚合物(EPS)进一步降解的物(Tang et al., 2015)。
 | 图5 北京延庆高于庄组四段鱼骨状方解石的显微生物特征Fig.5 Microscopic characteristics of microbial fabrics in herringbone calcite(HC)of the Member 4 of Gaoyuzhuang Formation in Yanqing County, Beijing |
运用场发射扫描电镜对部分硅化的鱼骨状方解石进行超微组构观察, 发现其内部保存有大量直径达10, μ m、长达100, μ m的丝状体(图 6-A), 与显微镜下观察到的微生物丝状体对应(图 5-C)。在丝状体的边缘保存有大量黏液状EPS构造以及粒径为50~200, nm的有机矿物纳米颗粒(图 6-B, 6-C, 6-D), 与同期微生物岩内发现的有机质残余以及伴生的有机矿物(Tang et al., 2013a, 2013b, 2013c, 2014, 2015)十分类似。
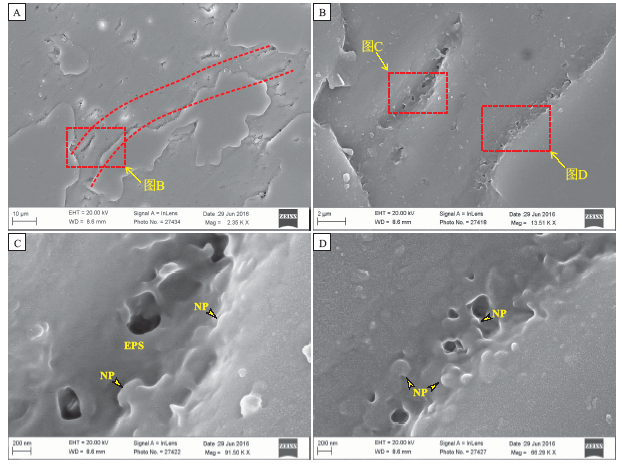 | 图6 北京延庆高于庄组四段鱼骨状方解石的超微生物特征Fig.6 Ultrastructure of microbial fabrics in herringbone calcite(HC)of the Member 4 of Gaoyuzhuang Formation in Yanqing County, Beijing |
鱼骨状方解石以及微生物礁其他组成部分的稀土配分模式如 图7所示。由图7可见, 用乙酸与用HNO3/HF溶样所测结果的稀土配分模式和稀土总量类似, 表明微生物礁受陆源碎屑影响微小, 故以下将不再区分这2种方法。鱼骨状方解石(n=7)的REE含量为4.104± 1.258 μ g/g、Th含量为0.082± 0.185 μ g/g、Y/Ho值为53.266± 3.948、PrSN/YbSN值为0.565± 0.241、Ce/Ce* 值为0.903± 0.093; 微生物席(n=8)的REE含量为5.964± 1.341 μ g/g、Th含量为0.315± 0.220 μ g/g、Y/Ho值为50.637± 4.013、PrSN/YbSN值为0.342± 0.102、Ce/Ce* 值为0.944± 0.059; 晶洞(n=5)的REE含量为6.944± 2.571 μ g/g、Th含量为0.019± 0.029 μ g/g、Y/Ho值为50.425± 6.632、PrSN/YbSN值为0.300± 0.144、Ce/Ce* 值为0.926± 0.097; 全岩(n=4)的REE含量为5.079± 1.534 μ g/g、Th含量为0.215± 0.166 μ g/g、Y/Ho值为54.611± 6.999、PrSN/YbSN值为0.362± 0.031、Ce/Ce* 值为0.949± 0.044。相比较而言, 鱼骨状方解石和晶洞的Th含量低于微生物席和全岩, 表明前者形成于受陆源碎屑影响更小的相对局限环境(孔洞)或具有更快的沉淀速率导致混入的陆源碎屑少; 鱼骨状方解石的其他各项指标与微生物礁其他组成部分相似, 表明沉积鱼骨状方解石的孔洞与海水流通良好。
 | 图7 北京延庆高于庄组四段鱼骨状方解石及其围岩的REE+Y配分图比较(A— 鱼骨状方解石; B— 微生物席; C— 晶洞; D— 全岩)Fig.7 REE+Y distribution patterns of herringbone calcite(HC)and its surrounding rocks of the Member 4 of Gaoyuzhuang Formation in Yanqing County, Beijing |
受陆源碎屑影响较小的非骨架碳酸盐岩被认为是记录海水稀土配分模式的重要载体(e.g., Webb and Kamber, 2000; Kamber and Webb, 2001; Nothdurft et al., 2004; Ling et al., 2013, Tang et al., 2016)。高于庄组微生物礁各种组成部分的REE和Th含量显著低于后太古宙澳大利亚页岩(PAAS)的REE(183 μ g/g)和Th(14.6 μ g/g)含量(McLennan, 2001); Y/Ho值显著高于PAAS的相应值27, 而与现代海水的该比值(44~74)接近; PrSN/YbSN值明显小于1, 稀土配分图左倾(图 7), 相似于现代海水的配分模式, 但区别于PAAS平坦的配分模式(PrSN/YbSN值接近1)。Ce异常按照Ce/Ce* =CeSN/(P
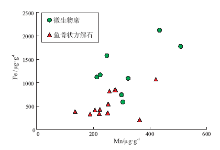
对鱼骨状方解石和微生物席内的碳酸盐沉淀抑制剂Fe和Mn的浓度测定结果见图8。由图8可见, 在鱼骨状方解石和微生物席内的Fe、Mn含量无显著差异; 前者的Fe、Mn浓度分别为549± 273 μ g/g和256± 76 μ g/g(n=12), 后者的Fe、Mn浓度分别为1273± 520 μ g/g和319± 103 μ g/g(n=8)。鱼骨状方解石内的Fe、Mn含量与加拿大中元古界Dismal Lake群微生物礁以及美国寒武系Ledger组微生物礁内鱼骨状方解石的Fe、Mn含量(de Wet et al., 2004; Bartley et al., 2015)类似。
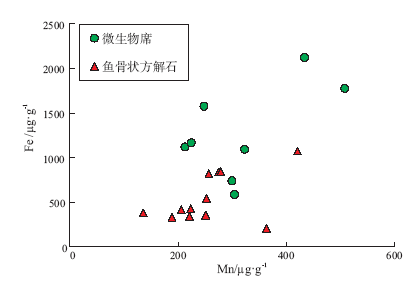 | 图8 北京延庆高于庄组四段鱼骨状方解石和微生物席内沉淀抑制剂含量对比Fig.8 Concentration of precipitation inhibitors in herringbone calcite(HC)and microbial mats of the Member 4 of aoyuzhuang Formation in Yanqing County, Beijing |
文中选取12个鱼骨状方解石、8个晶洞和20个微生物席共40个样品做无机碳氧同位素分析。由图9可见, 所有样品的 δ 18O 同位素值均大于-10‰ , 而且碳、氧同位素之间不存在相关关系, 表明样品的同位素组分未受明显的蚀变影响。其中鱼骨状方解石、晶洞、微生物席的碳同位素值分别为-0.94‰ ~-0.61‰ (平均-0.81‰ )、-1.27‰ ~-0.90‰ (平均-1.06‰ )、-1.10‰ ~-0.50‰ (平均-0.81‰ ); 氧同位素值分别为-4.42‰ ~-2.13‰ (平均-3.90‰ )、-6.50‰ ~-10.00‰ (平均-8.44‰ )、-5.54‰ ~-3.80‰ (平均-4.76‰ )。鱼骨状方解石与微生物席的碳氧同位素组成接近; 晶洞的碳同位素值较鱼骨状方解石和微生物席的值略低, 氧同位素则更低(图 9)。
 | 图9 北京延庆高于庄组四段鱼骨状方解石及其围岩的无机碳氧同位素组成对比Fig.9 Inorganic carbon and oxygen isotopic compositions of herringbone calcite(HC)and its surrounding rocks of the Member 4 of Gaoyuzhuang Formation n Yanqing County, Beijing |
在氧化水体中, 由于可溶的Ce3+会被氧化成为低溶解度的Ce4+, 而优先被水体中的Mn-Fe氢氧化物、有机质以及黏土矿物吸附而从水柱中清除(Byrne and Sholkovitz, 1996)。因此, 现代氧化海洋中沉淀的微生物碳酸盐岩表现出明显的Ce负异常(e.g., Webb and Kamber, 2000; Kamber and Webb, 2001)。相反, 沉淀于次氧化至缺氧水体中的碳酸盐岩不会表现出Ce负异常。这是由于从氧化水体中沉降的Mn-Fe氢氧化物颗粒会被溶解, Ce会从这些颗粒中释放并被还原为溶解度高的Ce3+(German and Elderfield, 1990; German et al., 1991; Byrne and Sholkovitz, 1996)。
高于庄组微生物礁的低REE和Th含量、高Y/Ho值、缺乏显著重结晶等特征综合表明, 它们记录了当时的古海水稀土特征(e.g., Bau and Dulski, 1996; Kamber and Webb, 2001; Nothdurft et al., 2004; Slack et al., 2007)。其缺乏或具弱Ce负异常(Ce/Ce* 范围为0.76~1.06)表明, 微生物礁形成于次氧化— 缺氧环境中, 很可能为氧化还原界面附近。
鱼骨状方解石沿晶体生长方向的旋转消光特征(图 4-G, 4-H)表明, 其内部亚晶随晶体的生长逐步发生了旋转。鱼骨状方解石晶体的这种生长特征符合球状晶体生长模式(Sumner and Grotzinger, 1996a), 而这种模式需要有沉淀抑制剂的参与才能实现。但是, 碳酸盐沉淀抑制剂的种类很多, 如Fe2+、Mn2+、Zn2+、Pb2+、Fe3+、Co2+、Be2+、Ni2+、Cd2+、Ba2+、Sr2+、Mg2+、S
在显生宙的鱼骨状方解石内有较多白云石析出, 据此有学者认为Mg2+是导致鱼骨状方解石沉淀的主要抑制剂(Tourre and Sumner, 1999)。但是, Mg2+的抑制能力较Fe2+的抑制能力低4个数量级以上(Meyer, 1984), 而且在高Mg/Ca值条件下形成的碳酸盐矿物应该以文石为主而非方解石。因此, 笔者认为Mg2+并不是导致高于庄组鱼骨状方解石沉淀的主要抑制剂类型。
鱼骨状方解石在中元古代之前以海底沉淀形式广泛产出, 其后则主要以微生物礁孔洞充填物的形式零星产出(Sumner and Grotzinger, 1996a, 1996b; Grotzinger and James, 2000)。其产出丰度和产出形式的长期演变很可能表明: 海水中的沉淀抑制剂在中元古代之前浓度较高, 之后明显降低, 至显生宙则基本没有。符合这种特征的抑制剂最可能为氧化还原敏感元素— — Fe2+和Mn2+。它们在前寒武纪缺氧水体中浓度高, 在显生宙氧化水体中浓度低(Sumner and Grotzinger, 1996a, 1996b); 而且Fe2+和Mn2+具有极高的碳酸盐沉淀抑制能力, 实验证明在Fe2+参与下可形成似鱼骨状方解石晶体(Meyer, 1984; Sumner and Grotzinger, 1996a, 1996b; Sumner and Tourre, 2000)。高于庄组微生物礁的稀土特征表明, 其沉积的底层海水处于Mn-Fe氧化还原界面附近。由于在研究区鱼骨状方解石主要形成于相对局限的孔洞环境, 并且伴随有有机质的降解, 故其沉积环境的氧浓度可能更低, 为缺氧环境。这就表明高于庄组的鱼骨状方解石的确形成于低氧环境, Fe2+、Mn2+可能是主要的沉淀抑制剂。因此, 鱼骨状方解石可以作为古氧相指示矿物, 反映缺氧水体环境, 结合其产出特征可进一步确定是底层海水缺氧或是孔洞水体缺氧。
华北中元古界高于庄组的鱼骨状方解石仅以孔洞充填形式产出, 而不像太古宙至早元古代那样, 可以海底沉淀形式直接沉淀。这可能主要受控于以下3个因素:
1)太古宙至中元古代, 海水的碳酸盐饱和度有所下降, 鱼骨状方解石沉淀需要通过有机质降解提升微环境碳酸盐饱和度, 从而诱发沉淀。
2)中元古代, 海水氧化程度有所升高, 需要相对局限的环境(孔洞)通过有机质降解消耗氧气, 以便增加鱼骨状方解石沉淀抑制剂的浓度。
3)碳酸盐饱和度降低后需要更低的碎屑输入速率, 否则鱼骨状方解石的生长很容易因陆源碎屑的输入而中断。
微生物活动对鱼骨状方解石沉淀的作用长期以来多被忽视。尽管微生物的降解一方面可降低微环境的氧浓度来增强鱼骨状方解石沉淀抑制剂的浓度, 另一方面可形成重碳酸盐从而增加微环境碱度和碳酸盐饱和度促进鱼骨状方解石沉淀, 但长期以来, 无论是以海底沉淀形式还是以微生物礁孔洞胶结物形式产出的鱼骨状方解石, 均被认为属无机成因(Grotzinger and James, 2000; Bartley et al., 2015)。导致这种认识的关键原因是从未找到微生物促进鱼骨状方解石沉淀的直接证据。本研究不仅在鱼骨状方解石内发现了大量微生物丝状体残余, 而且发现这些丝状体集中在鱼骨状方解石晶体内部, 排列方式与晶体生长方向一致(图 5)。超微组构观察进一步表明与微生物丝状体和EPS密切共生的有机矿物纳米颗粒(图 6)。这些特征充分表明, 微生物丝状体不仅是被包围在鱼骨状方解石晶体内部残余, 而且很可能通过微生物代谢活动(如铁还原过程、细菌硫酸盐还原过程)诱发了鱼骨状方解石的沉淀, 并为鱼骨状方解石成核提供了重要垫板。鱼骨状方解石与微生物席的碳同位素组成相近, 这可能表明前者与后者类似, 具有生物成因; 它们的碳同位素未明显负偏, 可能与当时海水巨大的溶解无机碳库(DIC)有关(Bartley and Kah, 2004)。
文中首次报道了华北中元古代的鱼骨状方解石, 通过研究得出以下结论:
1)华北中元古界高于庄组四段发育大量以孔洞胶结物形式产出的鱼骨状方解石。
2)高于庄组的鱼骨状方解石与其他地区和时代产出的鱼骨状方解石类似, 均符合球状晶体生长模式, 其沉淀需要有碳酸盐沉淀抑制剂的参与。
3)Ce异常特征指示高于庄组鱼骨状方解石形成于缺氧环境。结合分析鱼骨状方解石产出丰度和产出形式的长期演变规律, 以及各类碳酸盐沉淀抑制剂的能力, 提出氧化还原敏感元素Fe2+、Mn2+是控制鱼骨状方解石沉淀的主要抑制剂; 因此沉积岩中鱼骨状方解石的发育可以指示缺氧环境条件。
4)鱼骨状方解石晶体内部保存有与其生长方向一致的生物丝状体残余和有机矿物, 表明微生物代谢和有机质降解促进了鱼骨状方解石沉淀, 并为鱼骨状方解石的初始沉淀提供了成核垫板。
作者声明没有竞争性利益冲突.
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|