









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
山东青岛和日照滨岸潮间带现代生物遗迹组成与分布特征*
[王海邻1  , 胡斌
, 胡斌1, 2 , 宋慧波1 ]
, 胡斌, 宋慧波|
|

第一作者简介 王海邻,女,1971年生,副教授,博士研究生,主要从事沉积学和遗迹学的研究。E-mail: whl@hpu.edu.cn。
通讯作者简介 胡斌,男,1952年生,教授,博士生导师,主要从事古生物地层学、遗迹学和沉积学的教学和研究工作。E-mail: hub@hpu.edu.cn。
在山东青岛和日照滨岸潮间带可见到现代生物营造的各种遗迹。通过分析这些生物遗迹与其沉积背景的关系,取得以下成果: ( 1)研究区生物遗迹中,层面遗迹以腹足类动物的拖迹为主,层内遗迹以居住迹丰度最高,且类型多样。( 2)建立了 2个生物遗迹组合: 一是高潮区组合,主要遗迹类型包括珠带拟蟹守螺拖迹、沙蚕的拖迹、摄食迹和居住迹、四角蛤的居住迹、弹涂鱼的爬行迹和居住迹、鸭嘴海豆芽的居住迹以及螃蟹的居住迹和摄食迹;该组合产生的沉积物底层具有含泥量较高,水、氧、有机质含量均较丰富及基底较稳固等特点。二是中低潮区组合,主要由斑玉螺的拖迹和停息迹、扁玉螺的爬行迹和停息迹、寄居蟹的爬行迹、长竹蛏的居住迹、虾蛄的爬行迹和居住迹、鸭嘴海豆芽的居住迹以及螃蟹的居住迹和摄食迹组成;该组合产生的沉积物底层具有含泥量较低、粒度较粗、间隙水不易保存、有机质含量低等特点;研究区潮间带为急剧多变的海洋环境,该遗迹组合可指示温带季风气候条件下的海洋潮间带沉积环境。( 3)通过分析各种造迹生物及其遗迹的组成和空间分布特征以及其与底层沉积物和沉积环境的关系,建立了研究区潮间带沉积中的生物遗迹组合及其沉积环境分布模式。
About the first author Wang Hailin,born in 1971,is a Ph.D. student and an associate professor of Henan Polytechnic University. She is mainly engaged in researches of ichnology and sedimentology. E-mail: whl@hpu.edu.cn.
About the corresponding author Hu Bin,born in 1952,is a professor and Ph.D. supervisor of Henan Polytechnic University. He is mainly engaged in teaching and researches of paleontology and stratigraphy,ichnology and sedimentology. E-mail: hub@hpu.edu.cn.
Many traces of modern animals are found in the intertidal flat in Qingdao and Rizhao area,Shandong Province. By analyzing the relationship between the traces and the sedimentary environment,the results indicate that: (1)The major surface traces observed in the study area are the repichnia of gastropods,and the major interior traces are the domichnia. The type of the traces is multitudinous. (2)Two biogenic traces assembles are established. One is the biogenic traces assemble in the upper intertidal flat,which are mainly repichnia made by Cerithidea cingulata,repichnia,fodinichnia and domichnia made by Nereis succinea, domichnia made by Mactra veneriformis Reeve,repichnia and domichnia made by Periophthalmua cantonensis,domichnia made by Lingula anatina, domichnia and fodinichnia made by Brachyura. The characteristics of the sediment where this traces assembles produced include the high clay content,the abundance of water,oxygen and organic matter,the stable sediment. The other is the biogenic traces assemble produced in the middle and lower intertidal flat,which are mainly repichnia and cubichnia made by Natica maculosa,repichnia and cubichnia made by Glossaulax didyma,repichnia made by Solen gouldi,repichnia and domichnia made by Oratosquilla oratoria,domichnia made by Lingula anatina,domichnia and fodinichnia made by Brachyura. The characteristics of the sediment where this traces assembles produced include the lower clay content,the rough granularity,the interstitial water hold hard,the lower content of the organic matter, etc. For the intertidal flat of the study area belongs to changeable marine environment,this biogenic traces assembles can indicate the marine intertidal flat sediment environment of monsoon climate of medium latitudes. (3)By analyzing the modern animals and the traces' distribution characteristics,the relationship between the traces and the substrate,and summarizing the relationship of the traces' distribution characteristics and the sedimentary environment,the biogenic traces assemble and the distribution model of ichnoassemblages in the study area was established.
遗迹学是研究现代和古代生物生活时在沉积物底层中(层面和层内)遗留下来的活动痕迹(吴贤涛, 1986; 胡斌等, 1997), 这种生物活动遗迹与沉积环境、沉积物性质、生态条件有着密切的关系。了解现代各类造迹生物的活动规律、造迹行为习性、遗迹特点和分布特征及其与沉积环境的关系, 对判识遗迹化石及其形成的古环境条件, 具有重要的实证价值和指导意义。也正是如此, 从20世纪起, 现代潮坪沉积中的生物遗迹研究就受到了国内外遗迹学者的广泛关注。国外学者已在北海(Wadden Sea)、意大利、美国的撒佩罗岛(Sapelo Island region)、乔治亚州(Georgia)和威拉帕海湾(WillapaBay)以及日本等地做过大量工作(Schafer, 1962; Frey, 1970; Frey and Basan, 1978; Frey and Howard, 1986; Seike et al., 1999, 2011; Uchman and Pervesler, 2006), 这些研究成果极大地推进了遗迹学的发展。值得指出的是, Uchman和 Pervesler(2006)较详细地论述了潮间带沉积物的沉积特性(粒度、含水率和有机质含量等)与生物遗迹形态特征的关系, Bromley(1996)讨论了潮间带造迹生物的生活习性和造迹过程。
相比而言, 中国学者对古遗迹学研究较多, 而对新遗迹学的研究起步较晚、成果相对较少(王英国, 2000; 胡斌等, 2012, 2015)。另外, 宋慧波等(2014)考察和分析了中国杭州湾庵东滨岸潮间带现代沉积物中的生物遗迹组成及其分布特征, 王珍如等(1994)探索性地研究了中国北部湾现代潮间带的造迹动物及其遗迹特征。关于山东青岛、日照滨岸潮间带中现代生物遗迹的详细研究, 以往做的工作不多, 仅王珍如等(1988)对中国青岛、北戴河现代潮间带底内生物及其遗迹类型进行了统计。本次工作主要针对研究区现代沉积物中造迹生物与其遗迹组成和分布特征以及其与沉积环境之间的关系进行较深入的分析。
青岛位于山东半岛东南部沿海、胶东半岛东部, 东、南濒临黄海, 地势东高西低, 南北两侧隆起, 中间低凹, 青岛海岸线曲折, 岬湾相间。日照位于山东半岛东南部, 总的地势背山面海, 中部高四周低, 略向东南倾斜, 山地、丘陵、平原相间分布, 日照海岸位于黄海中部, 岬湾相连, 属于比较平直的基岩砾质海岸。该区属温带季风气候, 温度适中, 四季分明, 年平均气温12.7, ℃, 受海洋性气候影响, 气候湿润, 雨量较丰沛, 年平均降水量为662.1, mm, 年平均相对湿度为73%。潮汐属正规半日潮, 平均潮差为2.8, m左右, 海水盐度为32‰ 左右。
青岛和日照滨岸潮间带平坦, 退潮后潮间带露出的面积可达数公里, 为潮间带动物提供了良好的栖息场所。潮汐运动增加了氧气的吸收和溶解, 随潮流而来的有机碎屑又为动物提供了丰富的营养物质, 故该地区潮间带动物种类多样, 遗迹丰富。本次野外观察时研究区的选定综合考虑沉积物底质类型、受人工干扰程度, 最终确定3个观察区(图1), 分别是青岛海湾大桥附近(图1中A观察区)、青岛银沙滩(图1中B观察区)和日照刘家湾附近滨岸潮间带(图1中C观察区)。
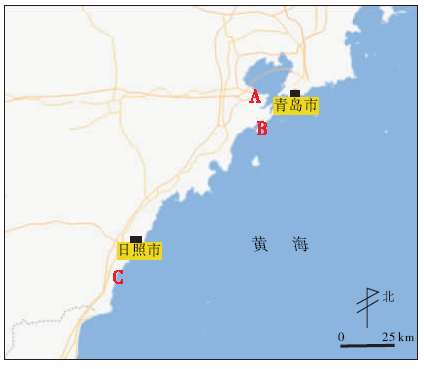 | 图1 山东青岛和日照滨岸潮间带现代生物遗迹采样点位置(底图来自Google EarthTM) A观察区— 青岛海湾大桥附近; B观察区— 青岛银沙滩; C观察区— 日照刘家湾附近滨岸潮间带Fig.1 Sampling location of modern animal traces made in intertidal flat of Qingdao and Rizhao area, Shandong Province(base map from Google EarthTM) |
青岛海岸带及邻近海域的现代沉积物类型分布特征总体表现为: 沿海岸粒度较粗, 为砂、粉砂质砂等, 向海粒度有逐渐变细的趋势; 沉积物主要为砂— 粉砂— 黏土, 局部为粉砂质黏土、黏土质粉砂和黏土质砂(肖菲和尹延鸿, 2006)。
日照近岸沉积区主要沉积物类型为砂、粉砂、泥质粉砂和砂— 砂粉— 泥。两城河以北, 岸线曲折、沉积物类型多变; 两城河以南, 岸线平直, 多为滩。在平面上沉积物类型呈现为与海岸平行的规则条带状分布, 垂直岸线方向沉积物有明显的分带性, 由岸向海依次由中砂到细砂、极细砂、粉砂质砂或砂质粉砂, 再过渡到泥质粉砂、泥质砂、砂— 粉砂— 泥, 粒度变化表现为由粗到细再到粗(周莉等, 1983)。
研究区滨岸潮间带沉积中, 非生物成因沉积构造十分发育。青岛海湾大桥附近和青岛银沙滩这2处潮间带地形平缓, 潮水运动方向主要为垂直岸线方向, 沉积物颗粒均匀, 层理主要是水平层理, 有时可见冲洗交错层理。日照刘家湾附近潮间带宽度约8~10, km, 地形起伏不平, 因此潮水流动方向多变, 潮流方向多为垂直岸线和平行或斜交岸线方向, 这就造成了该区沉积物表面波痕丰富、复杂多样, 波脊线呈垂直、平行和斜交岸线的都有, 常见的有对称波痕、不对称波痕、尖顶的或削顶的波痕、干涉波痕、舌形波痕等(图 2), 其中平行直脊波痕在高潮区发育普遍, 舌形波痕在中、低潮滩发育较普遍。
 | 图2 山东青岛和日照滨岸潮间带常见沉积构造 A— 潮间带沉积物表面的舌形波痕; B— 潮间带沉积物表面的平行直脊削顶波痕; C— 潮间带沉积物表面的平行直脊尖顶波痕; D— 潮间带沉积物表面的泥裂; E— 潮间带沉积物中的冲洗交错层理。A至C照片来自于C观察区, D照片来自于A观察区, E照片来自于B观察区Fig.2 Common sedimentary structures in intertidal flat of Qingdao and Rizhao area, Shandong Province |
根据野外观察和描述, 研究区潮间带沉积物中常见的造迹生物有12种, 包括节肢动物门的螃蟹(Brachyura)、虾蛄(Oratosquilla oratoria)和寄居蟹(Paguridae), 腕足动物门的鸭嘴海豆芽(Lingula anatina), 脊索动物门的弹涂鱼(Periophthalmua cantonensis), 软体动物门的四角蛤(Mactra veneriformis Reeve)、扁玉螺(Glossaulax didyma)、斑玉螺(Natica maculosa)、珠带拟蟹守螺(Cerithidea cingulata)和长竹蛏(Solen gouldi), 环节动物门的沙蚕(Nereis succinea)以及鸟类等。它们所造的遗迹类型主要是运动迹(拖迹、爬行迹、足辙迹)、停息迹、生殖迹、排泄迹、居住迹、觅食迹(摄食迹)以及鸟类足迹(表 1)。
| 表1 山东青岛和日照滨岸潮间带造迹生物种类及其遗迹类型 Table1 Trace-makers and their trace types in intertidal flat of Qingdao and Rizhao area, Shandong Province |
螃蟹(Brachyura)属节肢动物门软甲纲, 栖息于潮间带沉积物内, 退潮后外出活动, 以沉积物表面的有机碎屑为食, 在潮间带的高、中低潮区均有分布, 数量较多, 具群聚性, 但由于受人类活动的影响, 本研究区观察到的螃蟹形体较小。
研究区观察到的螃蟹所营造的遗迹主要是居住迹、摄食迹和足辙迹。居住迹就是居住潜穴(图 3-A), 穴口圆形, 与底表齐平, 大部分穴口周围有散落的掘穴砂球(图 3-B, 3-C, 3-D), 砂球有时为砂黄色, 与表层沉积物颜色相同, 有时呈黑灰色, 说明穴道底部已进入沉积物的还原层。穴道圆形, 与底表近似垂直, 表面光滑, 等径, 直径0.5~1.5, cm, 深度15~25, cm。穴道大小与动物个体大小有关, 深度与季节有关, 一般冬季因动物越冬穴道较深。有时穴口周围有一低矮的圆形泥墙, 亦叫潜穴烟囱(图 3-F), 内径与穴道口径相等, 对穴道起到保护作用, 一般高潮区含泥量较高的沉积环境中潜穴烟囱较高, 且整齐坚固, 砂质沉积环境中的潜穴烟囱低矮, 不易保存。
 | 图3 山东青岛潮间带现代沉积物中的螃蟹遗迹 A— 居住迹, 垂直管状潜穴; B— 沉积物表面上的螃蟹潜穴开口及周围散布的掘穴砂球; C— 螃蟹潜穴开口及摄食迹; D— 螃蟹摄食爪痕和摄食砂球; E— 退潮后沉积物表面密布的螃蟹摄食迹; F— 含泥质较高的沉积物表面的螃蟹潜穴泥墙。A至E照片来自于B观察区, F照片来自于A观察区Fig.3 Traces made by Brachyura in modern intertidal deposits of Qingdao area, Shandong Province |
摄食迹由动物摄食过程产生的爪痕和食渣砂球组成。爪痕是动物摄食时留下的线型凹痕, 连续或间断, 位于穴口周围或附近, 呈辐射状排列、沿穴口对称排列或位于穴口一侧(图3-C, 3-D)。摄食爪痕受沉积物性质影响, 当表层沉积物中有机质含量低时, 动物需远离穴口觅食, 因此摄食爪痕和食渣砂球的分布较散乱, 无规律, 摄食迹的清晰度与沉积物粒度有关, 粒度细, 爪痕清晰, 粒度粗则模糊不清。食渣砂球呈圆形, 直径3~4, mm, 数量极多, 一般在退潮后4 h左右即可在沉积物表面观察到密布的螃蟹潜穴开口及其周围散布的食渣砂球, 一个穴口周围可形成600~1100粒(图 3-E)。摄食迹的特点是分布于居住潜穴附近的沉积物表面, 在退潮期间快速形成, 涨潮时受潮水冲刷而破坏。
研究区砂质沉积环境中螃蟹居住迹、摄食迹丰度较高, 表面常见大量掘穴砂球、食渣砂球和摄食爪痕, 但由于沉积物粒度较粗, 螃蟹从沉积物表面爬过后一般难以观察到其爬行迹。泥质沉积环境中螃蟹潜穴丰度高, 当沉积物含水率较低时, 潜穴口泥墙较高且较坚固(图3-F), 同时可观察到螃蟹在沉积物表面爬行时留下的清晰的足辙迹。
鸭嘴海豆芽(Lingula anatina)属腕足动物门无铰纲无穴目, 外形为双叶状, 背腹两壳呈扁平鸭嘴形, 绿色。背壳较小, 后部较圆; 腹壳较大, 后部较尖。壳表光滑, 生长线明显, 壳缘外套生有刚毛, 伸出壳外。肉茎粗而长, 圆柱形, 由壳后端伸出, 生活于潮间带细沙质或泥质底内, 借肌肉收缩挖掘泥沙, 营穴居生活。海豆芽从早寒武世出现, 至今已经历了漫长的地质时代, 是古海洋生态学研究中的主要指相动物, 其典型的生物遗迹特征也可作为海洋潮间带沉积环境判定的重要参考。研究区潮间带沉积物中鸭嘴海豆芽数量分布最多, 在日照刘家湾附近的潮间带砂质沉积物中每平方米的数量可达38个, 观察到的鸭嘴海豆芽壳体长2.5~3.5, cm, 宽1.2~1.8, cm, 壳厚0.5~0.8, cm, 肉茎长8~18, cm。
研究区观察到的鸭嘴海豆芽造的遗迹主要是居住迹, 为竖直的潜穴, 分为上部的壳体穴道和下部的肉茎穴道两部分(图 4-A, 4-B)。壳体穴道由动物壳体上下运动时挤压周围沉积物形成, 横截面为扁双凸镜形(图 4-C), 深度为10~20, cm, 是壳体长度的4~6倍, 穴径略大于壳体的宽度和厚度, 一般在壳体穴道上部1/3处有一纵脊, 是动物上升运动时壳体以略倾斜的角度左右摆动对穴壁压磨的结果。潜穴穴口与底表齐平, 通常有以下2种形态: 一种是由3个并列的小孔组成的三孔式(图 4-D, 右上侧), 孔径1~2, mm, 中间的小孔略大于两侧的小孔, 有时在3个小孔的周围有透镜形的砂脊(图4-D, 左下侧), 三孔式主要是由动物外套膜前缘的刚毛组成的两侧的进水孔和其间的出水孔在静水条件下所形成。另一种为单孔式(图 4-D, 左上侧), 穴孔1个, 双透镜形, 是动物壳体局部伸出底表时形成的。潜穴下部为肉茎穴道, 其长度约为总穴道的1/2左右, 是肉茎在沉积物内呈最大限度收缩时挤压周围沉积物而形成的。
 | 图4 山东日照潮间带现代沉积物中的鸭嘴海豆芽遗迹 A— 鸭嘴海豆芽潜穴; B— 鸭嘴海豆芽潜穴侧面; C— 鸭嘴海豆芽壳体穴道横截面形状; D— 鸭嘴海豆芽潜穴在沉积物表面的穴口, 左上角为单孔式, 右上侧为三孔式、周围无砂脊, 左下侧为三孔式、周围有砂脊Fig.4 Traces made by Lingula anatina in modern intertidal deposits of Rizhao area, Shandong Province |
虾蛄(Oratosquilla oratoria)属节肢动物门软甲纲口足目, 主要生活于潮间带中潮区至低潮区的泥质沉积物中, 穴居生活, 涨潮时游出穴外在水中游泳或爬行, 落潮后潜入有积水的穴道内, 以小虾等小型动物为食。
虾蛄造的遗迹主要是层面上的爬行拖迹和层内的居住潜穴。爬行拖迹是虾蛄爬过沉积物表面时其舌形尾扇划擦沉积物表面留下的扁平条带状拖痕, 痕迹直或微弯曲, 较浅, 宽度一般0.5~1.0, cm, 与其尾扇宽度一致(图 5-A)。居住潜穴是由动物附肢挖掘而成, 穴道口处一般有泥丘或不整齐的瘤状堆积物, 是动物挖掘洞穴时清理出的沉积物经水流冲击后形成的。穴道为Y型或U型, 垂直于底表或略倾斜, 穴径2~3, cm, 一般上部较细, 向下逐渐变粗, 深度可达18~50, cm。由于沉积物较软, 其潜穴较难观察和保存。
 | 图5 山东日照潮间带现代沉积物表面的虾蛄拖迹(A)和弹涂鱼爬行迹(B)Fig.5 Traces made by Oratosquilla oratoria(A)and Periophthalmua cantonensis(B) in modern intertidal deposits of Rizhao area, Shandong Province |
弹涂鱼(Periophthalmua cantonensis)也叫跳跳鱼, 属脊索动物门辐鳍鱼纲, 具有两栖性, 可在潮间带沉积物内或表层生活。
弹涂鱼造的遗迹包括居住潜穴和表层的爬行迹。其居住潜穴多为“ Y” 形, 上部穴道直径一般为2~3, cm, 深度一般5~8, cm, 由于沉积物较软, 其潜穴一般难以观察和保存。
爬行迹是生物在泥质沉积物表面爬行及跳跃过程留下的痕迹(图 5-B)。研究区观察到的爬行迹长度15~80, cm, 与其爬行距离有关, 而宽度与动物个体大小有关。中间有时有拖痕, 拖痕一般宽2~5, mm, 是其身体后部拖过沉积物表面所留下的痕迹。两侧对称排列的弧形是其胸鳍和尾鳍的压痕: 胸鳍压痕较大, 位于外侧, 长度0.5~1.5, cm, 宽度0.3~0.5, cm; 尾鳍压痕较小, 位于内侧, 长度0.2~0.5, cm, 宽度0.1~0.3, cm。
观察区的弹涂鱼主要分布在高潮区含泥量较多的沉积地带, 当表层沉积物粒度较小、含水率适中时, 其痕迹清晰, 胸鳍压痕中可见明显的纹理; 当含水率过高的泥质汤底或含水率过低沉积物表面较固结时, 其爬行迹不够清晰。
四角蛤(Mactra veneriformis)属于软体动物门双壳纲帘蛤目, 主要栖息于潮间带高潮区或中潮区, 底内穴居。
观察区四角蛤造的遗迹主要是居住迹, 是动物向沉积物内挖掘潜穴时, 由于壳体的挤压作用和2个水管经常伸缩活动, 在底内沉积物中形成明显且具固定形状的居住潜穴。潜穴整体上与底表垂直, 分为上部的水管穴道和下部的壳体穴室2部分(图 6-A): 由于其2个水管已全部愈合, 因此上部的水管穴道近似圆形, 长2~3, cm, 穴径0.3, cm左右, 在沉积物表面有开孔; 下部的壳体穴室与动物壳体形状一致, 长3.1~3.3, cm, 宽2.5~3.0, cm, 表面光滑。研究区四角蛤居住迹主要分布在高潮区, 丰度较高, 在1 m× 1 m范围内可观察到80~125个, 对底表向下5~6, cm深度的沉积物扰动剧烈(图 6-B), 形成密集的潜穴。
 | 图6 山东青岛潮间带现代沉积物中四角蛤造的遗迹 A— 四角蛤潜穴; B— 四角蛤在6, cm深度内沉积物中的扰动特征。所有照片均来自于A观察区Fig.6 Traces made by Mactra veneriformis in modern intertidal deposits of Qingdao area, Shandong Province |
沙蚕属(Nereis succinea)环节动物门多毛纲须虫目, 栖息于潮间带的泥质沉积物中。研究区观察到的沙蚕主要分布在高潮区含泥量相对较高的砂质沉积环境中, 一般体宽约5, mm, 体长8~15, cm。沙蚕造的遗迹主要有运动拖迹、居住迹和排泄迹。运动拖迹是沙蚕在底层表面爬行时留下的, 浅U形, 一般3~10, cm长, 1~2, mm宽, 直或微弯曲, 不分枝, 两端常与居住潜穴洞口相连(图 7-A)。居住迹是沙蚕潜入沉积物内部时形成的管状居住潜穴, 圆形, 等径, 直径大小与沙蚕大小有关, 该考察区潮间带的沙蚕潜穴直径1~2, mm, 深度一般5~8, cm, 有垂直管状、” Y” 形、” U” 形, 表面光滑, 颜色与表层沉积物相同, 沙蚕造的潜穴穴口在表层呈圆形, 穴口或穴道内有时有粪粒产生(图 7-B, 7-C), 粪粒直径约0.2, mm, 在穴口呈圆锥状粪丘分布或散布于穴口周围。
 | 图7 山东青岛潮间带现代沉积物中沙蚕造的遗迹 A— 沉积物表面沙蚕造的拖迹; B— 沙蚕潜穴在沉积物表面的穴口, 周围分布有粪粒; C— 沙蚕“ U” 形潜穴, 部分被粪粒填充。所有照片均来自A观察区Fig.7 Traces made by Nereis succinea in modern intertidal deposits of Qingdao area, Shandong Province |
与宋慧波等(2014)在杭州湾庵东滨岸潮间带现代沉积物表面发现的沙蚕遗迹(宋慧波等, 2014)相比, 青岛和日照潮间带现代沉积物中沙蚕遗迹的形态与其形状相同, 但遗迹丰度较低, 扰动程度也较低。
扁玉螺(Glossaulax didyma)属软体动物门腹足纲中腹足目。扁玉螺贝壳呈半球形, 背腹扁而宽。壳顶低小, 螺旋部较短, 体螺层宽度突然加大。壳面光滑无肋, 生长纹明显。壳面呈淡黄褐色, 壳顶为紫褐色, 基部为白色。其斧足发达, 可包被贝壳, 用来挖掘泥沙, 使身体埋于沉积物中, 或半潜入沉积物内巡捕食物。多分布在潮间带中低潮区低洼有水的地方。
观察区内扁玉螺造的遗迹主要是运动迹和停息迹。运动迹是动物在沙上爬行时由于前足锄沙的作用而留下的痕迹, 研究区观察到的扁玉螺壳体大小3~4, cm, 其运动迹为不规则的U形带状爬行迹, 表面光滑, 一般与波痕共生, 宽度约5, cm, 两侧有明显的砂脊, 是动物运动时其前足将前方的泥沙铲向两侧面形成的, 长度可达3~9, m(图 8-A)。停息迹是动物全部潜入沉积物内时形成的囊状停息迹, 在沉积物表面凸起呈丘状(图 8-B)。
 | 图8 山东日照潮间带现代沉积物中的扁玉螺遗迹 A— 拖迹, 与波痕共生; B— 囊状停息迹Fig.8 Traces made by Glossaulax didyma in modern intertidal deposits of Rizhao area, Shandong Province |
斑玉螺(Natica maculosa)属软体动物门腹足纲中腹足目, 壳体厚, 体螺层底宽, 略平, 壳体尺寸6~8, mm。运动时身体背着壳体在沉积物表面匍匐运动, 以附着在沉积物中的藻类为食。一般分布在潮间带中低潮区, 其分布受潮流和波浪的影响较大, 局部高度密集, 在30 cm× 30 cm面积内可有90~100个(图 9-D)。
 | 图9 山东日照潮间带现代沉积物中的斑玉螺和扁玉螺遗迹 A— 斑玉螺“ C” 形拖迹; B— 斑玉螺直形、微弯曲形拖迹; C— 半嵌入沉积物停息的扁玉螺; D— 密集分布的扁玉螺拖迹和停息迹; E— 扁玉螺停息迹示意图Fig.9 Traces made by Natica maculosa and Glossaulax didyma in modern intertidal deposits of Rizhao area, Shandong Province |
观察区内斑玉螺造的遗迹主要是运动迹和停息迹。其运动迹为浅U形带状拖迹, 宽度与壳体底部宽度一致, 两侧有明显略高于底表的砂脊, 从形状上来看, 斑玉螺运动拖迹形状不规则, 大体上有直形、微弯曲形、“ C” 形等(图 9-A, 9-B)。停息迹是动物全部潜入沉积物内或半潜入沉积物内时形成的, 当动物半潜入沉积物内时形成凹陷囊状表生半浮痕停息迹(图 9-C)。当动物全部潜入沉积物内时, 沉积物表面呈丘状, 有小孔与外界保持联系(图9-D, 9-E)。
寄居蟹(Paguridae)属于节肢动物门软甲纲十足目, 常寄居在空螺壳内, 体躯左右不对称, 腹部较柔软, 可卷曲于螺壳中。活动时头胸部伸出壳口, 靠其3对步足拖动身体及螺壳一起运动。
观察区内寄居蟹造的遗迹主要是爬行迹, 由3对步足的爪痕和螺壳的拖痕共同组成。寄居蟹前进方向不同, 可形成3种不同形状的爬行迹: 当寄居蟹前进时, 由于第1对大螯的爪痕被螺壳的拖痕抹掉, 因此其爬行迹由两侧对称排列的第2、3对步足的爪痕和中间的螺壳拖痕组成, 有时两侧的第2、3对步足爪痕不明显, 仅可见其螺壳拖痕, 常呈“ C” 形、弧形弯曲或直线形, 宽度约5, mm, 此时的运动迹(图 10-A)与斑玉螺、珠带拟蟹守螺的运动拖迹相似。当寄居蟹后退时, 其爬行迹由3对步足的爪痕组成, 其中位于中间大且深者为其第1对大螯爪痕。当寄居蟹侧向运动时, 其爬行迹由侧向排列的3对步足爪痕和螺壳的拖痕组成(图 10-B)。寄居蟹爬行迹(图 10-C)分异度高, 但丰度较低。
 | 图10 山东日照潮间带现代沉积物中的寄居蟹造的遗迹 A、B— 寄居蟹爬行拖迹; C— 寄居蟹爬行拖迹素描图Fig.10 Traces made by Paguridae in modern intertidal deposits of Rizhao area, Shandong Province |
研究区观察到的寄居蟹体长2~3, cm, 体型较小, 其运动迹多形成于刚刚退潮后含水率较高且粒度较细的沉积物表面, 沉积物粒度较粗时其运动迹模糊不清难以辨认。
珠带拟蟹守螺(Cerithidea cingulata)属软体动物门腹足纲, 贝壳塔形, 呈尖锥形, 高35, mm左右, 宽度约为高度的1/3。生活于沉积物表层, 研究区所观察到的遗迹为其运动拖迹, 主要分布在潮间带高潮区。该运动拖迹是动物在沉积物表面匍伏运动时拖拉着贝壳在表层沉积物上划擦而形成(图 11-A), 拖迹宽度4~8, mm, 呈直线形、弧形弯曲或任意弯曲形, 犁沟状, 无分枝, 偶有交切, 属表生半浮痕遗迹。
 | 图11 山东青岛和日照潮间带现代沉积物中珠带拟蟹守螺造的拖迹和长竹蛏的居住迹 A— 沉积物表面珠带拟蟹守螺造的拖迹, A观察区; B— 长竹蛏造的居住潜穴, C观察区Fig.11 Traces made by Cerithidea cingulata and solen gouldi in modern intertidal deposits of Qingdao and Rizhao area, Shandong Province |
与宋慧波等(2014)在杭州湾庵东滨岸潮间带现代沉积物表面发现的珠带拟蟹守螺遗迹相比, 青岛和日照潮间带沉积物中的拖迹形态与杭州湾地区形态类似, 但其遗迹的丰度和分异度均较低, 仅发现其运动拖迹, 未观察到停息迹和生殖迹。
长竹蛏(Solen gouldi)属软体动物门双壳纲, 研究区观察到的长竹蛏长6~8, cm, 体宽约0.8, cm, 两壳合抱呈竹筒状, 前后两端开口, 壳质薄脆, 壳表光滑, 呈淡绿色或黄褐色。长竹蛏生活于潮间带中下潮区至浅海的沙泥沉积物中, 将身体大部分埋入砂泥中, 以强而有力的锚形斧足直立于沉积物中生活。
观察区内长竹蛏造的遗迹主要是居住迹, 竖直穴道, 与底表垂直; 穴道上下等粗, 横切面与壳体的高度、宽度一致; 穴壁光滑, 穴道深18~20, cm(图 11-B)。
研究区观察到的生物遗迹包括层面遗迹和层内遗迹。层面遗迹以腹足类动物的爬行拖迹为主, 主要有运动拖迹、觅食迹(摄食迹)、停息迹, 有时可见沙蚕粪粒, 造迹生物主要有珠带拟蟹守螺、沙蚕、弹涂鱼、虾蛄、斑玉螺、扁玉螺、寄居蟹、螃蟹。层内遗迹以居住迹丰度最高, 且类型多样, 主要有垂直管状、“ Y” 形、 “ U” 形等, 造迹生物主要是螃蟹、沙蚕、鸭嘴海豆芽、长竹蛏、虾蛄和弹涂鱼等。该区生物种类丰富, 遗迹类型多样。
基于生物遗迹的空间分布特征及其沉积物底层的特点, 可以划分出2个不同沉积背景条件下的生物遗迹组合。组合1为高潮区生物遗迹组合(图 12), 遗迹类型包括珠带拟蟹守螺拖迹、沙蚕拖迹、摄食迹和居住迹(居住潜穴内有时可见粪粒)、四角蛤居住迹、弹涂鱼爬行迹和居住迹, 鸭嘴海豆芽居住迹以及螃蟹的居住迹和摄食迹。在含泥量相对较高的沉积环境中, 四角蛤居住迹丰度较高, 对0~6, cm深度的沉积物扰动程度强烈。该组合产生的沉积物底层具有含泥量较高, 水、氧、有机质含量均较丰富、较稳固等特点, 因此适于生物生存, 造迹生物及其遗迹种类较丰富, 生物遗迹较易保存和观察。
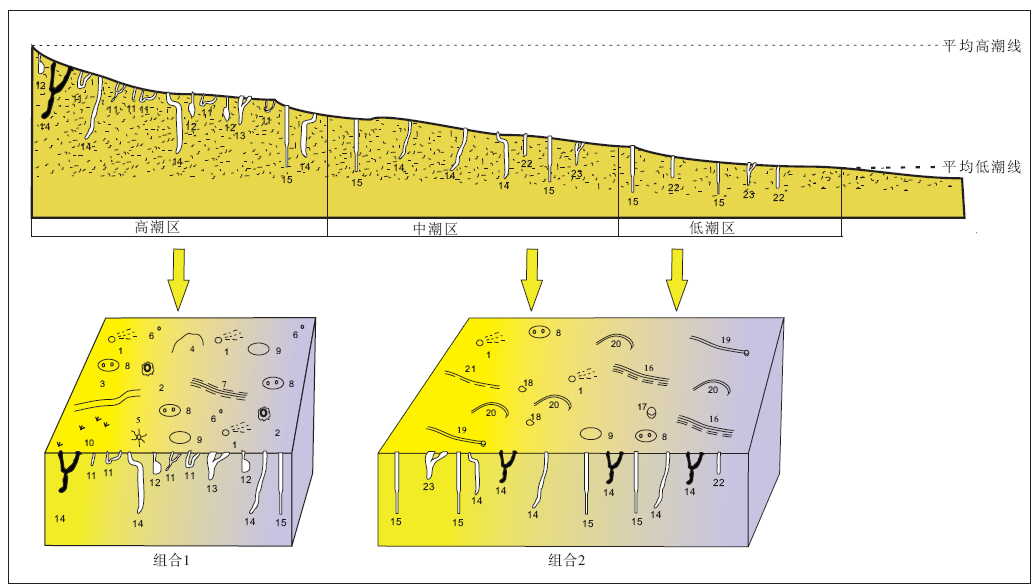 | 图12 山东青岛和日照潮间带现代沉积物中的生物遗迹组合及其沉积环境分布模式 1— 螃蟹居住潜穴开口; 2— 螃蟹居住潜穴开口, 周围有泥墙; 3— 沙蚕运动拖迹; 4— 珠带拟蟹守螺拖迹; 5— 沙蚕摄食迹; 6— 沙蚕居住潜穴开口; 7— 弹涂鱼爬迹; 8— 鸭嘴海豆芽潜穴开口, 双孔形; 9— 鸭嘴海豆芽潜穴开口, 单孔形; 10— 鸟足迹; 11— 沙蚕“ U” 形、“ Y” 形、垂直管状居住潜穴; 12— 四角蛤潜穴; 13— 弹涂鱼“ Y” 形潜穴; 14— 螃蟹“ J” 形、“ Y” 形、垂直管状居住潜穴; 15— 鸭嘴海豆芽潜穴; 16— 寄居蟹拖迹; 17— 扁玉螺停息迹; 18— 斑玉螺停息迹; 19— 斑玉螺拖迹; 20— 扁玉螺拖迹; 21— 虾蛄拖迹; 22— 长竹蛏居住潜穴; 23— 虾蛄“ Y” 形居住潜穴Fig.12 Distribution model of ichnoassemblages and their sedimentary environments in modern intertidal deposits of Qingdao and Rizhao area, Shandong Province |
组合2为中低潮区生物遗迹组合(图 12), 遗迹类型主要由斑玉螺运动拖迹和停息迹、扁玉螺爬行迹和停息迹、寄居蟹爬行迹、长竹蛏居住迹、虾蛄的爬行迹和居住迹, 鸭嘴海豆芽居住迹以及螃蟹的居住迹和摄食迹组成, 其中斑玉螺拖迹和停息迹局部密集, 扰动剧烈, 主要分布在退潮后低洼积水处附近。该组合产生的沉积物底层具有含泥量较低、粒度较粗、间隙水不易保存、有机质含量低等特点, 因此底内动物类型单调, 但适应者居住数量较多, 居住迹丰度较高。
研究区潮间带为急剧多变的海洋环境, 该环境下的生物类型与半咸水环境的杭州湾庵东滨岸潮间带中生物类型和遗迹组合特征(宋慧波等, 2014)有明显的不同。由于鸭嘴海豆芽是古海洋生态学研究中的主要指相动物, 其典型的生物遗迹特征也可作为海洋潮间带沉积环境判定的重要参考。研究区的遗迹组合可指示温带季风气候下的海洋潮间带沉积环境。
1)在青岛和日照滨岸潮间带观察到的生物遗迹包括层面遗迹和层内遗迹, 层面遗迹以腹足类动物的爬行拖迹为主, 主要有运动拖迹、觅食迹(摄食迹)、停息迹, 有时可见沙蚕粪粒, 造迹生物主要有珠带拟蟹守螺、沙蚕、弹涂鱼、虾蛄、斑玉螺、扁玉螺、寄居蟹、螃蟹。层内遗迹以居住迹丰度最高, 且类型多样, 主要有垂直管状、“ Y” 形、 “ U” 形等, 造迹生物主要是螃蟹、沙蚕、鸭嘴海豆芽、长竹蛏、虾蛄和弹涂鱼等。
2)基于生物遗迹的空间分布特征及其沉积物底层的特点, 建立了2个不同沉积背景条件下的生物遗迹组合: 一是高潮区生物遗迹组合, 主要遗迹类型包括珠带拟蟹守螺拖迹、沙蚕拖迹、摄食迹和居住迹(居住潜穴内有时可见粪粒)、四角蛤居住迹、弹涂鱼爬行迹和居住迹, 鸭嘴海豆芽居住迹以及螃蟹的居住迹和摄食迹。该组合产生的沉积物底层具有含泥量较高, 水、氧、有机质含量均较丰富、基底较稳固等特点, 因此适于生物生存, 造迹生物及其遗迹种类较丰富, 生物遗迹较易保存和观察。二是中低潮区生物遗迹组合, 主要遗迹类型由斑玉螺运动拖迹和停息迹、扁玉螺爬行迹和停息迹、寄居蟹爬行迹、长竹蛏居住迹、虾蛄的爬行迹和居住迹, 鸭嘴海豆芽居住迹以及螃蟹的居住迹和摄食迹组成。该组合产生的沉积物底层具有含泥量较低、粒度较粗、间隙水不易保存、有机质含量低等特点, 因此底内动物类型单调, 但适应者居住数量较多, 其居住迹丰度较高。青岛和日照潮间带为急剧多变的海洋环境, 该遗迹组合可指示温带季风气候下的海洋潮间带沉积环境。
3)通过分析各种造迹生物及其营造的遗迹组成特点与底层沉积物的关系, 总结了这些生物遗迹的空间分布特征与沉积环境的关系, 从而建立了青岛和日照潮间带沉积物中的生物遗迹组合及其沉积环境分布模式。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|