

{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国湖相碳酸盐岩时空分布与碳氧同位素特征*
[苏玲1, 2, 3  , 朱如凯
, 朱如凯1, 2, 3 , 崔景伟1, 2, 3 , 吴松涛1, 2, 3 , 罗忠1, 2, 3 , 毛治国1, 2, 3 , 李森1 ]
, 朱如凯, 崔景伟|
|

第一作者简介 苏玲,女, 1982年生,硕士,主要从事沉积储层研究。 E-mail: suling2200@petroChina.com.cn。
通讯作者简介 朱如凯,男, 1968年生,博士,教授级高级工程师,主要从事沉积储层研究。 E-mail: zrk@petroChina.com.cn。
湖相碳酸盐岩产状多样,多以夹层形式分布在碎屑岩剖面中,也见以结核或钙质微体化石等薄层赋存于泥岩、页岩等碎屑岩中,具有层数多、单层薄、呈韵律性变化等特点。受构造背景、物源输入和古环境等因素控制,中国湖相碳酸盐沉积最早出现于二叠纪,主要发育时段为古近纪,具有沉积时间跨度大、分布面积广的特征。湖相碳酸盐岩按成因可分为原生沉积型、成岩改造型和热液喷流型 3类。中国湖相碳酸盐岩的碳氧同位素特征可揭示原生和成岩过程中水介质环境的差异性: ( 1)δ 13C和 δ 18O 密切相关,且δ 13C多正偏,指示封闭型咸水、半咸水湖泊环境;( 2)δ 13C和 δ 18O 无相关性,且δ 13C多负偏,指示开放型湖泊环境;( 3)δ 13C严重正偏,指示成岩作用时发酵带的环境受到古细菌参与的甲烷生成作用的影响。
, Zhu Rukai, Cui JingweiAbout the first author Su Ling,born in 1982,is engaged in research of reservoir sedimentology. E-mail: suling2200@petrochina.com.cn.
About the corresponding author Zhu Rukai,born in 1968,is engaged in research of reservoir sedimentology. E-mail: zrk@petrochina.com.cn.
Lacustrine carbonate rocks have various occurrences. They commonly occur as the interbeds in clastic rocks with characteristics of multi-layers,thin individual layer and rhythm. Occasionally,they occur in mudstone and shale as thin layers of concretions or calcareous microfossils. Controlled by tectonic background,provenance and palaeoenvironment,the lacustrine carbonate rocks in China show wide spatial-temporal distribution,which were firstly reported in the Permian,but mainly found in the Paleogene. Based on carbonate and oxygen isotope analysis,different water environments during primary deposition and diagenetic process are concluded: (1)Well-related δ13C and δ18O with mostly positive δ13C values,indicating a closed saltwater or semi-saltwater lake environment;(2)Completely-unrelated δ13C and δ18O values with mostly negative δ13C values,indicating an open lake environment;(3)Seriously-positive δ13C values,indicating that during diagenesis,the environment was affected by the methane generation process,which was participated by archaebacteria.
在湖盆演化过程中, 湖相碳酸盐岩一般发育于构造活动相对稳定、湖盆水体持续扩张的阶段。在淡水内陆湖盆、咸水内陆湖盆、偶具海侵的内陆湖盆和潟湖湖盆中, 均可发育各种碳酸盐岩(王英华等, 1993)。平面上, 湖相碳酸盐岩多分布于近盆缘的滨岸带和盆内浅湖中的水下隆起区, 呈小面积分布; 垂向上, 多以互层形式分布在碎屑岩剖面中, 也见以结核或钙质微体化石等薄层赋存于泥岩、页岩等碎屑岩中, 具有层数多、单层薄、呈韵律性变化等特点, 厚度比例一般不到总厚度的15%(邹才能, 2013)。湖相碳酸盐岩岩性多样, 包括生物灰岩、颗粒灰岩、礁灰(云)岩、泥晶灰岩和白云岩等(王英华等, 1993), 陆源碎屑的混杂较为普遍, 受相带控制明显, 不同相带上的碳酸盐岩类型在平面上呈连续或不连续的带状环: 滩相和礁相在滨浅湖区相对隆起的正地形顶部或斜坡地带发育, 以生物灰岩、鲕粒灰岩为主, 呈透镜状, 在高部位厚度较大, 呈不连续片状或连续带状环岸分布; 浅湖区发育白云质灰岩; 水下隆起区发育鲕粒灰岩、灰质白云岩; 半深湖区以泥灰岩为主, 多呈薄层状夹在黑色泥岩中(邹才能, 2014)。
国内外学者针对湖相碳酸盐岩岩石学特征、沉积环境、生烃特征和储集特征, 已开展了大量研究工作(Talbot and Kelts, 1990; 王英华等, 1993; Mazzullo, 2000; 刘传联等, 2001; 李红等, 2013; 柳益群等, 2013; 钱凯等, 2013; 闫伟鹏等, 2014)。由于湖相碳酸盐岩与富含有机质的湖相沉积物密切相关, 而富有机质湖相泥页岩是重要的烃源岩类型, 因此湖相碳酸盐岩的研究对于明确泥页岩形成环境、揭示有机质富集机理具有重要意义(Talbot and Kelts, 1990)。此外, 作为重要的储集岩, 湖相碳酸盐岩紧邻烃源岩发育, 源储一体, 成藏条件优越, 在油气勘探开发中已有突出表现(钱凯等, 2013; 邹才能, 2013, 2014), 如近年来, 在中国含油气盆地的湖相碳酸盐岩层中(如准噶尔盆地二叠系芦草沟组、四川盆地侏罗系大安寨段、渤海湾盆地的古近系、柴达木盆地的古近系等)发现了具一定规模的油气资源, 表明其是重要的勘探领域(张晓宝等, 2000; 廖静等, 2008; 陶士振等, 2012; 毛玲玲等, 2014; 闫伟鹏等, 2014)。
对于湖相碳酸盐岩(夹层或结核)成因, 大多数学者认为是一定构造背景下湖相沉积环境中的正常沉积作用(刘传联等, 2001)形成的, 也有学者提出其是热液作用、喷流作用的产物(郑荣才等, 2006; 柳益群等, 2013)。不同成因认识对于湖相碳酸盐岩及共生的湖相泥岩、页岩的古环境恢复、沉积相带分布预测和储集层评价都有重要影响。其中碳氧同位素是反映沉积水体环境的关键参数, 在碳酸盐岩成因、古气候和古环境演化等方面得到了广泛的应用。作者通过对鄂尔多斯盆地中上三叠统延长组碳酸盐岩结核碳氧同位素的分析, 同时调研已发表文献中碳氧同位素数据, 发现不同盆地、不同层位、不同岩性的碳酸盐岩碳氧同位素数据差异非常大, 特别是碳同位素的分布区间大, δ 13C值的变化范围从-10.8‰ 至17.9‰ 。文中通过对不同地区湖相碳酸盐岩碳氧同位素分析数据的对比, 以期探讨碳氧同位素反映的古环境信息, 这对于研究共生的页岩发育环境以及早期成岩阶段流体的特征与演化、寻找有利储集层、研究页岩形成环境以及早成岩阶段成岩作用均具有重要意义。
中国湖相碳酸盐岩分布范围广, 如四川盆地川中大安寨组灰岩分布面积约1.5× 104, km2; 渤海湾盆地古近系沙河街组白云岩分布面积约2.0× 104, km2; 准噶尔盆地二叠系云化岩分布面积约3000, km2; 松辽盆地白垩系青山口组灰岩分布面积约1.5× 104, km2; 柴达木盆地古近系灰岩分布面积约1.0× 104, km2。大多数湖盆的湖相碳酸盐岩以灰岩沉积为主, 部分为白云岩, 少数为碳酸盐岩与碎屑岩组成的混积岩(表 1)。不同时代湖相碳酸盐岩岩性特征与空间分布具有一定的差异性。
| 表1 中国湖相碳酸盐岩沉积学特征及碳氧同位素特征 Table1 Sedimentary characteristics and carbon and oxygen isotopic characteristics of lacustrine carbonate rocks in China |
古生界湖相碳酸盐岩主要发现在准噶尔、三塘湖等盆地, 与二叠系芦草沟组烃源岩伴生(表 1)。准噶尔盆地吉木萨尔凹陷芦草沟组中的碳酸盐岩与碎屑岩组成的混积岩中, 碳酸盐岩单层厚度普遍较薄, 一般为50~100, cm, 最厚约200, cm, 并以夹层形式赋存于泥(页)岩中。在准噶尔盆地南缘东部乌鲁木齐地区红雁池南剖面和养牛场剖面中, 见中、薄层深灰色、灰色碳酸盐岩和粉砂岩与中、厚层灰黑色泥岩和油页岩形成互层沉积(张晓宝等, 2000; 雷川等, 2012; 李红等, 2013)。三塘湖盆地中碳酸盐岩以2种沉积组合形式产出, 反映了沉积环境差异: (1)白云质泥晶灰岩、凝灰岩、白云岩及火山岩以夹层形式赋存于厚层暗色泥岩、凝灰质泥岩、白云质泥岩中, 指示了地幔热液喷积作用的背景(柳益群等, 2010, 2013); (2)白云岩、灰岩与黑色泥岩、页岩等碎屑岩呈互层沉积, 指示了湖泊沉积背景(李红等, 2012)。
中生界湖相碳酸盐岩以白垩系为主(表 1), 在松辽盆地、二连盆地、酒西盆地、四川盆地等广泛发育, 多与富有机质泥页岩伴生, 但产状差异较大。松辽盆地白垩系嫩江组暗色泥岩中发育2种碳酸盐岩: (1)层状白云岩和椭球状白云岩结核, 以层状为主; (2)泥质铁白云岩结核, 主要呈顺层状产出于灰绿色粉砂质泥岩、页岩与介形虫层组成的互层层序中(刘万洙和王璞珺, 1997; 王国栋等, 2008)。内蒙古二连盆地白音查干凹陷下白垩统碳酸盐岩以浅灰色— 深灰色泥质白云岩为主, 夹泥岩(郭强等, 2012)。酒西盆地下白垩统下沟组发育湖相白烟囱型喷流岩, 与正常沉积的暗色页岩形成不等厚互层(郑荣才等, 2006)。辽西上白垩统义县组碳酸盐岩以夹层形式发育在火山岩— 沉积碎屑岩之中(陈登辉等, 2011)。四川盆地下侏罗统自流井组发育少量湖相碳酸盐岩, 主要是介壳灰岩与紫红色泥岩互层, 部分剖面上可见微晶白云岩夹层(王全伟等, 2006)。近年来, 作者在鄂尔多斯盆地东南缘铜川地区露头的中上三叠统延长组长7段油页岩中发现大量直径达60~100, cm、孤立分布的碳酸盐岩结核(图 1)。
 | 图 1 鄂尔多斯盆地铜川地区中上三叠统延长组湖相碳酸盐岩结核 a— 油页岩中发现的大量直径达60~100 cm、孤立分布的结核, 长7段; b— a图中结核的切断面, 长7段Fig.1 Lacustrine carbonate concretion of the Middle-Upper Triassic Yanchang Formation in Tongchuan area, Ordos Basin |
新生界湖相碳酸盐岩发育广泛(表 1), 中国东部渤海湾、苏北、广东三水、广西百色等均有发现。古近纪是湖相碳酸盐岩发育的全盛时期, 碳酸盐岩具有厚度大、面积广、层数多、储油物性好的特征(钱凯等, 2013)。渤海湾盆地歧口凹陷沙河街组湖相碳酸盐岩与泥质岩呈互层沉积, 白云岩约占碳酸盐岩体积分布的31%(廖静等, 2008); 东营凹陷沙河街组湖相碳酸盐岩主要分布在沙一段和沙四段, 夹于暗色泥岩中, 呈薄层状产出(刘传联, 1998; 刘传联等, 2001), 其中枝管藻生物礁储集层是主力产油层(王夕宾等, 2016)。辽东湾盆地古近系沙河街组产有一套仅限于深凹陷内的深湖相白云岩, 在深湖相暗色泥岩中呈薄层状产出(戴朝成等, 2008)。江汉盆地江陵凹陷古新统沙市组和新沟嘴组(王春连等, 2013)及潜江凹陷上始新统(张永生等, 2006)发育以石盐为主、固液相并存的综合性盐类矿床, 盐间地层岩性以泥岩、泥质碳酸盐岩、碳酸盐质泥岩为主; 在泌阳凹陷, 古近系发育暗色泥质白云岩、白云质泥岩, 夹薄层粉、细砂岩(黄杏珍等, 2001)。柴达木盆地古近系干柴沟组集中发育的湖相碳酸盐岩, 呈夹层状产出在碎屑岩中(毛玲玲等, 2014)。青藏高原北部地区新生代陆相地层中发育碳酸盐岩, 以古近系陆相红层和新近系湖相碳酸岩为代表(伊海生等, 2007)。此外, 在现代沉积中也见到湖相碳酸盐的产出, 如青海湖湖底沉积物中存在微米级球状白云石集合体(于炳松等, 2007); 小柴旦盐湖南岸发现的一种湖滩岩型硼质细砂岩中, 出现白云石包壳、白云石藻屑和藻团块(夏文杰和李秀华, 1986)。
| 表 2 中国湖相碳酸盐岩岩石学特征 Table2 Petrological characteristics of lacustrine carbonate rocks in China |
湖相碳酸盐沉积以盆内和近源沉积为主, 多以层状、薄层状、结核状、透镜状等夹于黑色泥岩、页岩和油页岩中(表 2), 其岩石学特征与形成环境密切相关(孙钰等, 2008)。仅从岩性分布就可看到, 不同地区的湖相碳酸盐岩特征具有明显差异:准噶尔盆地南缘二叠系、松辽盆地白垩系湖相碳酸盐岩以白云岩为主, 矿物组成以铁白云石为主, 含少量石英、长石和方解石, 发育微晶结构, 多以层状、透镜状或结核形式产出。三塘湖盆地、青藏高原北部、渤海湾盆地、江汉盆地的湖相碳酸盐岩中均发育白云岩, 且以泥晶白云岩为主, 但成因具有差异。三塘湖盆地、酒西盆地、二连盆地中碳酸盐岩的形成与火山活动有关, 为喷流型热水沉积白云岩(白烟囱), 以铁白云石含量高为特征, 且含热水成因的矿物如方沸石、纳沸石等, 铁、锰含量较高(郑荣才等, 2006; 郭强等, 2012; 柳益群等, 2013)。青海湖和小柴旦盐湖现代沉积物中也发现了白云石集合体、白云石包壳和白云石藻屑等。但辽西义县、柴达木盆地、四川盆地等产出的湖相碳酸盐岩主要以灰岩为主, 仅见很少量白云岩。
碳在自然界主要以自然碳、氧化碳和还原碳3种形式存在, 碳同位素的演变主要与2条循环途径相关: (1)碳同位素交换, 即CO2气体与各种水溶碳酸根原子团之间的碳同位素交换反应使碳酸盐富集13C, 总的来说, 13C趋于富集在碳的高价化合物中, 即CH4(13C最亏损)→ C→ CO→ CO2→ C
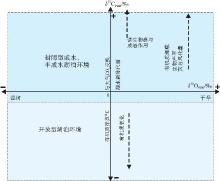
原生碳酸盐中的氧同位素可反映湖水构成和矿物沉淀温度, 其中湖水构成受地表水、地下水和雨水的注入量、流出量及蒸发量影响(Talbot and kelts, 1990)。原生碳酸盐中碳同位素记录了沉积时水体中溶解的无机碳浓度(DIC), 而溶解的无机碳中δ 13C变化与碳来源有关。此外, 其还受水体的盐度、温度、深度、氧化还原条件、生物作用以及水体与碳酸盐矿物之间的碳分馏系数影响。一般来说, 盐度升高δ 13C值增大, 温度降低δ 13C增大, 水体深度增加δ 13C降低, 缺氧还原条件下δ 13C降低, 生物降解可使δ 13C变轻(郑永飞和陈江峰, 2000)。碳来源主要包括: (1)进入水中的碳, 包括河流带入的碳酸根离子、先存碳酸盐矿物的溶解、热液流体供给的C等(李荣等, 2008); (2)与大气CO2交换(McKenzie, 1985; Lee et al., 1987); (3)湖水中的生物新陈代谢(浮游植物通过光合作用吸收较多的12C, 使表层水体中的溶解无机碳中13C含量相对增加, 进而使表层水体中形成的原生碳酸盐δ 13C值偏高)(Stiller and Hutchinson, 1980)。此外, 富含12C的有机质的埋藏量以及埋藏速度影响原生碳酸盐的碳同位素组成。同时, 碳氧同位素还受湖水滞留时间影响(McKenzie, 1985; Lee et al., 1987)(图 2), 不同类型湖盆碳氧同位素变化差异明显。
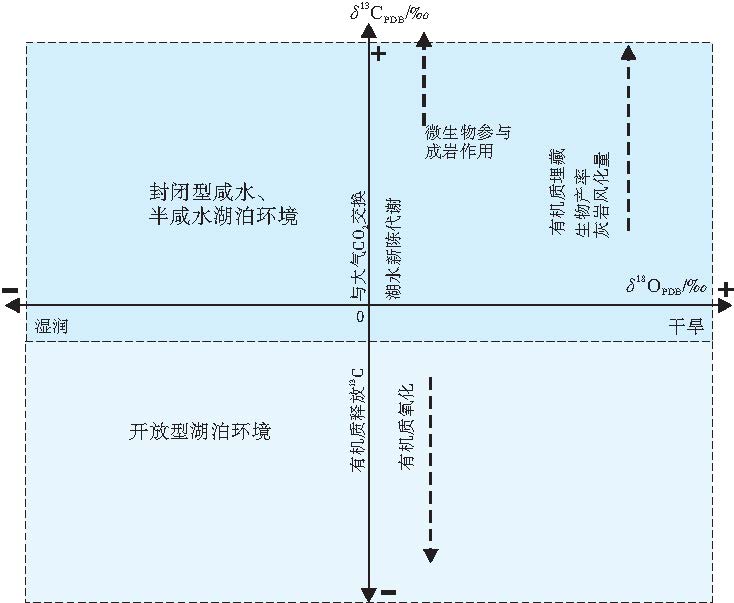 | 图 2 湖水和碳酸盐沉积物中 δ 18O 和δ 13C变化的主要控制因素(据伊海生等, 2007, 有修改)Fig.2 Major controlling factors on δ 18O and δ 13C variations of lake water and carbonate sediments (modified from Yi et al., 2007) |
开放型湖盆中, δ 13C受地表水中固定的无机碳浓度、光合作用和CO2呼吸循环 δ 18O 的影响(McKenzie, 1985); 而蒸发作用对δ 18O 的影响很小。由于注入水停留时间短, 碳氧同位素的影响因素不同, 因此在开放型湖盆中二者的变化趋势呈不相关(Talbot and Kelts, 1990)。
封闭型湖盆中, 碳氧同位素具有很强的相关性和共变趋势。氧同位素的变化大部分是水体平衡变化的结果, 同时, 表层富12C的CO2优先脱气, 水体中富集13C(Talbot and Kelts, 1990)。水体贮存时间长, 蒸发作用主导了同位素的组成(Fontes and Gonfiantini, 1967; Gat, 1981; Gonfiantini, 1986)。当然, 用初级生产力变化来解释湖相碳酸盐岩同位素的变化(Stiller and Hutchinson, 1980; McKenzie, 1985), 尚不能有力地解释封闭型湖盆中溶解的无机碳的组成及碳酸盐的沉积特征, 因此需要考虑微生物对其的影响, 这在下文详细叙述。但是, 封闭型湖盆中独有的碳氧同位素共变趋势, 是湖盆形态、气候、地质背景和水文作用共同影响的结果(Talbot and Kelts, 1990)。
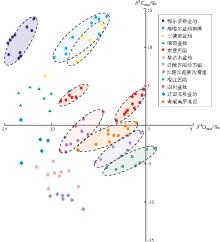
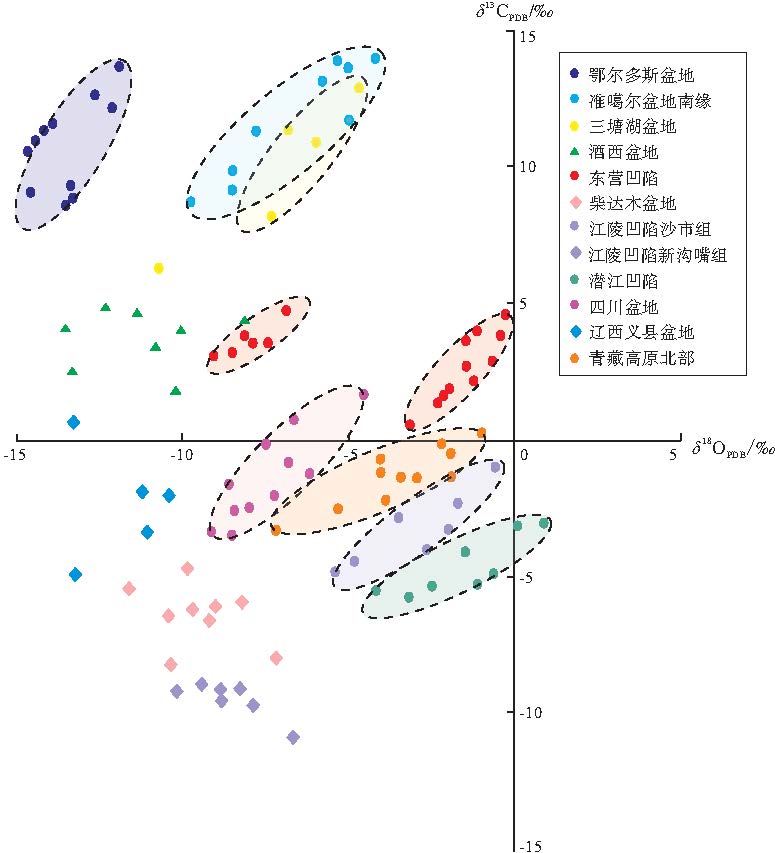
氧同位素大于-10‰ 可认为属于原生型碳酸盐岩, 基本不受成岩作用影响(曲长胜等, 2017), 但鄂尔多斯盆地三叠系延长组湖相碳酸盐岩结核、酒西盆地和辽河义县盆地碳酸盐岩明显遭受了成岩作用影响, 导致其不能被用来判断原生水体的环境。文中建立以δ 13C值为纵坐标、δ 18O值为横坐标、0为原点的坐标体系(Talbot and Kelts, 1990; 刘传联等, 2001), 根据样品的分布投点位置, 判断沉积时的古环境(图3)。其中, 若样品投点主体位于第1、2象限、或原点附近, 且δ 13C和δ 18O呈明显相关性, 指示了封闭型咸水— 半咸水湖盆; 若样品投点落在第3象限, 且δ 13C和δ 18O之间不相关, 则指示了开放型淡水湖盆环境。笔者将中国典型湖相碳酸盐岩碳氧同位素数据进行投点分析(图 3), 数据来源于发表的文献及本单位实验数据(表1)。根据投点位置判断碳酸盐岩沉积时的水体环境, 总的来说可分为3类:
1)δ 13C和 δ 18O 相关性密切, 且δ 13C以正偏为主。典型代表包括准噶尔盆地二叠系芦草沟组层状碳酸盐岩、三塘湖盆地二叠系芦草沟组泥晶白云岩、东营凹陷沙一段和沙四段碳酸盐岩等, 碳酸盐岩主要以结核、互层或夹层形式赋存于富有机质页岩中, 整体沉积粒度较细, 有机质丰度高。碳酸盐 δ 13C和 δ 18O 相关性明显, 且δ 13C以正偏为主、δ 18O以负偏为主, 表明碳酸盐形成于湖水滞留时间相对较长、封闭型的咸水— 半咸水湖泊体系中, 且δ 13C正偏可能与微生物有关。
2)δ 13C和 δ 18O 相关性密切, 且δ 13C多为负偏。典型代表包括四川盆地侏罗系大安寨段、青藏高原北部、江陵凹陷沙市组和潜江凹陷的湖相碳酸盐岩等, 碳酸盐岩与细粒沉积物主要以互层形式发育, 整体有机质丰度稍低于第1种类型, δ 13C和 δ 18O 偏负, 投点在原点附近, 且具相关性, 表明它们发育在蒸发作用明显、湖水滞留时间相对较长的封闭型咸水— 半咸水湖泊体系中。
3)δ 13C和 δ 18O 相关性弱。典型代表包括江陵凹陷新沟嘴组和柴达木盆地(王春连等, 2013; 毛玲玲等, 2014), δ 13C和 δ 18O 相关性弱, 表明其发育环境属于水体滞留时间短的开放性湖泊系统中。
作者认为湖相碳酸盐的形成除受原生碳酸盐形成时的水体环境影响外, 后期成岩改造也对其形成演化产生重要影响, 因此仅依靠原生碳酸盐不能完全反映湖相碳酸盐岩的形成与演化。成岩碳酸盐岩的氧同位素往往是在继承先存矿物氧同位素组成的基础上, 反映其形成时的孔隙水温度和组成; 且其氧同位素组成也与碳酸盐矿物的种类有关(同一流体、相同温度情况下形成的方解石、白云石, 其氧同位素组成因分馏系数的差异而不同)(胡作维等, 2012), 同时, 细菌等微生物活动是影响碳酸盐岩碳氧同位素变化的关键因素。
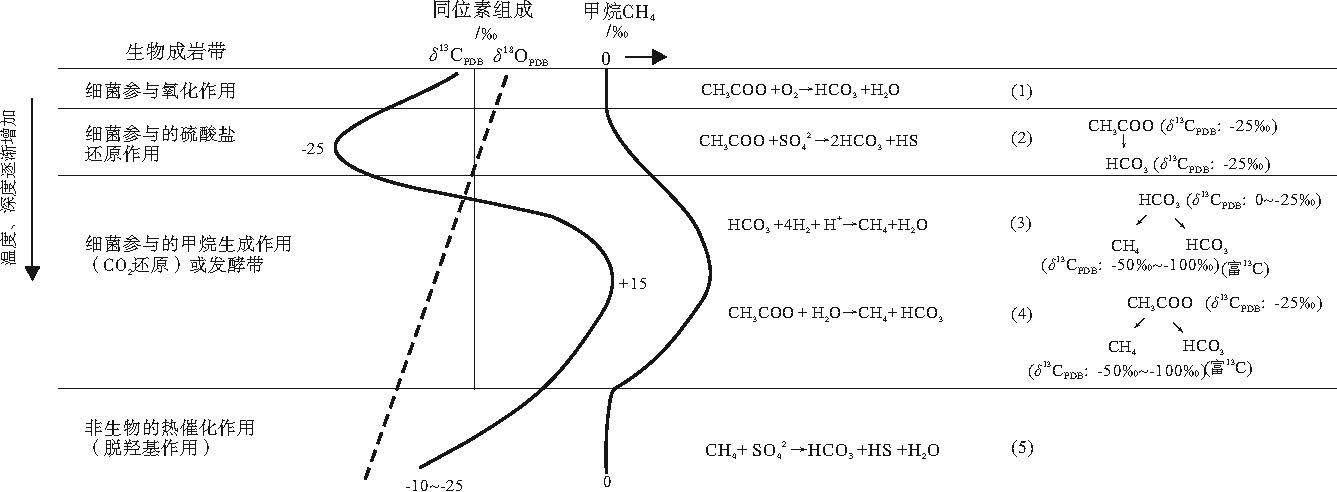 | 图4 不同成岩作用带的碳氧同位素特征(修改自Talbot and Kelts, 1990; Mazzullo, 2000)Fig.4 Characteristics of carbon and oxygen isotops in different diagenetic belt (modified from Talbot and Kelts, 1990; Mazzullo, 2000) |
鄂尔多斯盆地三叠系延长组湖相碳酸盐岩结核、准噶尔盆地二叠系芦草沟组碳酸盐岩受成岩作用影响明显。湖相碳酸盐岩多与富有机质沉积物伴生, 富有机质沉积物包含大量的活性有机物, 微生物活跃, 其主要通过2种方式影响碳酸盐的形成: 第1种是通过改变孔隙水的pH和碱性, 引起碳酸盐分解和沉淀; 第2种是对沉积有机质的降解, 这是主要的方式。微生物对沉积有机质的降解作用是一系列的氧化还原反应。生理学决定微生物具有分带性, 氧化剂有效性也随深度增加而变化, 所以不同水深的微生物氧化还原反应决定了碳酸盐成岩、碳酸盐岩碳同位素组成特征, 具体可分为3种(图 4)。
1)细菌参与的氧化作用。分子氧主导的有氧环境中, 嗜盐喜氧细菌参与代谢作用。
2)细菌参与的硫酸盐还原作用。缺氧海相环境中, 不稳定的有机质被硫酸盐还原菌氧化, 相比其他成岩阶段, 其要求快速的硫酸盐还原反应。在湖泊中, 溶解的硫酸盐浓度相对较低(Livingstone, 1963; Davison, 1988), 所以硫酸盐还原菌的作用不太明显。
3)细菌参与的甲烷生成作用。湖相沉积物中, 低浓度的硫酸盐和富有机质的硫酸盐池快速耗尽, 使细菌产甲烷作用成为早成岩阶段的主导因素(Talbot and Kelts, 1990)。生物甲烷生成作用有2个途径: (1)海相环境下, 硫酸盐还原过程中, 有机质去碳酸基释放CO2或HC
通常, 与嗜盐喜氧细菌、硫酸盐还原细菌及甲烷厌氧氧化作用有关的碳酸盐岩δ 13C总体呈现偏负的特征, 这是由于碳酸盐岩部分碳源来自于微生物降解有机质形成的贫13C的重碳酸根; 在细菌参与的甲烷生成作用中, 由于碳同位素分馏作用, 形成贫13C的CH4和富13C的CO2, 从而形成相对富集13C的碳酸盐矿物。
基于上述认识, 笔者认为鄂尔多斯盆地碳酸盐结核、准噶尔盆地白云岩δ 13C严重正偏, 可能是发酵带内细菌参与的甲烷生成作用导致的, 甲烷菌活动引起有机质碳同位素分馏。青藏高原地区叠层石灰岩类、东营凹陷沙一段和沙四段碳酸盐岩同位素显示13C富集, 可能也与微生物活动有关。
1)湖相碳酸盐岩垂向上多以夹层形式分布在碎屑岩剖面中, 也见以结核或钙质微体化石等(夹)薄层赋存于泥岩、页岩等碎屑岩中, 具有层数多、单层薄、呈韵律性变化等特点。中国湖相碳酸盐岩沉积最早出现于二叠纪, 主要发育时段为古近纪, 表现出沉积时间跨度大、分布面积广的特征。
2)碳酸盐岩碳氧同位素受到沉积初期原始水体环境及成岩改造双重作用控制, 具有阶段性演化特征: (1)碳酸盐沉积初期, 碳氧同位素的相关性可反映湖盆的水文环境; (2)碳酸盐岩成岩阶段, 碳氧同位素尤其是13C的分布, 能够反映不同成岩时期发生的微生物作用过程。
3)基于湖相碳酸盐岩碳氧同位素值, 发现δ 13C和 δ 18O 相关性可揭示沉积水介质环境的差异性: (1)δ 13C和 δ 18O 密切相关, 且δ 13C多为正偏, 指示了封闭型咸水、半咸水湖泊环境; (2)δ 13C和 δ 18O 无相关性, 且δ 13C多为负偏, 指示了开放型淡水湖泊环境; (3)δ 13C严重正偏, 指示了成岩作用时发酵带的环境受到古细菌参与的甲烷生成作用的影响。微生物在碳酸盐岩形成过程中的作用是不容忽视的, 通过碳氧同位素分析来研究微生物碳酸盐岩, 有待继续深入。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|