



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黔南地区中二叠世碳酸盐台地边缘沉积演化及古海洋意义*
[孟琦1, 2  , 黄恒
, 黄恒1, 3 , 颜佳新1, 2 , 陈发垚1, 2 ]
, 黄恒, 陈发垚|
|

第一作者简介 孟琦,男,1993年生,中国地质大学(武汉)古生物学与地层学专业硕士研究生,主要从事碳酸盐岩沉积地质学研究。E-mail: mengqi@cug.edu.cn。
华南地区二叠纪为东特提斯地区一个大型碳酸盐台地。中二叠世茅口亚世,上扬子碳酸盐台地经历了从全盛到萎缩的重要转折。黔南地区地处该台地南缘,是通过地层结构及其变化探讨台地演化控制因素的最佳地区。文中选取黔南罗甸纳水和沫阳 2个代表性剖面,通过对茅口组的岩石类型和骨屑颗粒组合统计分析,从碳酸盐生产工厂的角度开展研究。野外和室内研究共识别出 15种岩石类型、 5种沉积相和 6种骨屑颗粒组合,并恢复了各组合对应的碳酸盐生产工厂的特征。沉积相及碳酸盐生产工厂分析表明,黔南地区自晚孤峰期开始经历了由热带浅水、暖水工厂到热带较深水、温凉水工厂的明显转变,表现为孤峰阶上部开始出现较深水沉积。这一碳酸盐生产工厂的转变现象在华南地区孤峰期—早冷坞期普遍存在,表现为台地萎缩、台地边缘被淹没。中二叠世晚孤峰期至晚二叠世吴家坪期是全球重大地史转折期,光合作用生物遭受重创、海洋生态环境急剧恶化,上述碳酸盐生产工厂的急剧转变与当时热带浅海碳酸盐生产工厂被抑制或破坏有关。
About the first author Meng Qi,born in 1993,is a master candidate of paleontology and stratigraphy in China University of Geosciences(Wuhan). He is mainly engaged in research of carbonate sedimentology. E-mail: mengqi@cug.edu.cn.
South China,part of Eastern Tethys,developed a huge carbonate platform in the Permian. During the Maokouin Subepoch of the Middle Permian,the carbonate platform experienced a transition from its prosperity to shrink. The southern Guizhou was located at the southern margin of the carbonate platform. It is a good area for discussing the controlling factors of platform evolution by analysis of stratigraphic structure and its variation. In this study,we selected the typical Nashui and Moyang sections of Luodian,southern Guizhou and studied on their carbonate factory by analyzing rock types and skeletal association statistics. As a result,15 rock types,5 sedimentary facies and 6 skeletal associations are recognized by field and indoor research. The carbonate factory features of the skeletal associations were also rebuilt. The results show that a distinct transition from tropical,shallow and warm water factory to tropical,deep and cool water factory occurred starting from the late Kuhfengian in the southern Guizhou,manifested as the beginning occurrence of deep water sediments in the upper Kuhfengian Stage. This transition was widespread from the Kuhfengian to the early Lengwuan in South China,manifested as the shrink of the carbonate platform and the drowning of the platform margin. The late Kuhfengian of the Middle Permian to the Wuchiapingian of the Late Permian was a significant turning point of global geological history where the photosynthetic organisms suffered heavy losses and the marine ecological environment deteriorated sharply. The drastic change of the carbonate factory was related to the inhibition or destruction of the shallow tropical carbonate factory.
华南地区二叠系碳酸盐岩沉积广布, 为东特提斯地区一个大型碳酸盐台地。该碳酸盐台地起源于泥盆纪中晚期, 至早二叠世栖霞亚世达到全盛, 之后自中二叠世茅口亚世逐渐萎缩, 表现为多地茅口组浅水碳酸盐岩被较深水碳酸盐岩或者硅质岩取代:在台地北侧的中、下扬子地区, 中二叠世茅口组灰岩之上广泛覆盖孤峰组硅质岩, 而且向西硅质岩层位逐渐升高, 即碳酸盐台地向西退缩(Sheng and Jin, 1994), 如鄂西恩施猫耳山剖面茅口组上部钙泥质和硅泥质沉积地层厚度仅仅十余米(Zhang et al., 2007); 台地萎缩特征在台地南部同样清楚, 如紫云四大寨剖面(较深水盆地区)(Wang et al., 2016)、罗甸纳水剖面(南缘斜坡带)(史晓颖等, 1999)和来宾铁桥剖面(孤立台地边缘)(姚尧等, 2012), 孤峰期— 早冷坞期沉积环境水深明显增加。至晚二叠世吴家坪期, 受华夏古陆和康滇古陆上升影响, 陆源碎屑岩分布范围扩大, 上扬子碳酸盐台地规模已远不及茅口亚世, 萎缩特征更加显著。晚二叠世长兴期台地规模虽有所恢复, 但是规模明显不及栖霞亚世(刘宝珺和许效松, 1994; 王立亭等, 1994; 冯增昭等, 1996; 马永生等, 2009; 郑和荣和胡宗全, 2010)。因此中二叠世孤峰期— 早冷坞期是华南晚古生代碳酸盐台地演化过程中一个由全盛到萎缩的重要转折期。
有关上扬子碳酸盐台地南部边缘演化, 一般认为与同沉积断裂活动有关, 因为该带紧邻右江盆地(或称滇黔桂台盆等)。如右江盆地中一些浅水孤立台地上晚茅口亚世的沉积序列, 可能与黔桂地区二叠纪拉张伸展和差异沉降的区域构造背景有关(杨玉卿和冯增昭, 2000; 梅冥相等, 2003; 吴浩若, 2003; 王新强和史晓颖, 2008; 杨怀宇等, 2010)。但是通过对桂中台地东南边缘沉积相结构分析, 姚尧等(2012)认为桂中茅口亚世台地边缘相的演化可能与当时同沉积断裂关系不大, 而可能与当时的古海洋条件有关。
碳酸盐台地边缘位于浅水碳酸盐台地与较深水盆地之间的过渡地带(Flü gel, 2010), 是识别台地演化历史的最好地区。黔南罗甸地区二叠系出露完整、连续, 是华南海相二叠系的标准地区之一(冯增昭等, 1994; 刘宝珺和许效松, 1994)。自20世纪90年代开始的生物地层学(如王志浩, 1991, 2000; Zhu and Zhang, 1994)和大规模露头层序地层学(如梅仕龙等, 1999a; 史晓颖等, 1999)研究, 深化和提高了黔南二叠纪地层的划分和对比, 探索了海平面变化等因素对该相带地层结构的影响(陈洪德等, 1999; 王成善等, 1999a; 梅冥相等, 2004)。前人成果及我们团队前期野外调查表明, 黔南罗甸地区中二叠统岩相分异明显(金振奎, 1996), 除发育生物礁和生物碎屑滩外, 也发育硅质岩、硅质泥岩、局限台地白云质灰岩沉积及重力流、风暴岩事件沉积(王立亭等, 1994; 薛武强等, 2012), 意味着该台地边缘在中二叠世经历过明显的、大规模进积/退积迁移。但是, 前人的研究大多基于台地演化的二维模式开展, 涉及上述台地演化的问题也往往笼统地归为构造背景成因, 因此难以对台地演化的深层原因做进一步探讨。文中以罗甸纳水剖面和沫阳剖面为重点, 在岩石类型分析和骨屑颗粒组合分析的基础上, 恢复了该地区茅口亚世碳酸盐生产工厂模式, 从碳酸盐成因的角度分析上扬子碳酸盐台地南部边缘在中二叠世茅口亚世的演化, 探讨其古海洋意义。
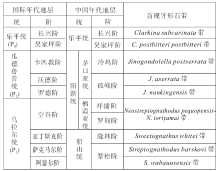
国际二叠纪年代地层系统中将二叠系分为3统9阶, 下统为乌拉尔统(Cisuralian), 中统为瓜德鲁普统(Guadalupian), 上统为乐平统(Lopingian)。文中重点研究的瓜德鲁普统自下而上分为罗德阶(Roadian)、沃德阶(Wordian)和卡匹敦阶(Capitanian), 均为牙形石带所限定(图 1)。
 | 图1 中国与国际二叠纪年代地层对比简图(据金玉玕等, 1999, 2000修改)Fig.1 Sketch map showing Chinese and international Permian chronostratigraphic correlation(modified from Jin et al., 1999, 2000) |
根据2002年全国地层委员会划分方案, 中国二叠纪年代地层自下而上分为船山统、阳新统和乐平统, 其中阳新统分为栖霞亚统(罗甸阶、祥播阶)和茅口亚统(孤峰阶、冷坞阶)(图 1)。
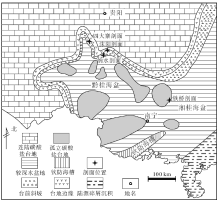
黔南地区在构造上位于扬子板块南部, 二叠纪以浅水碳酸盐沉积为主(贵州省地质矿产局, 1987), 大致沿河池— 罗甸— 紫云— 望谟— 册亨— 罗平一线发育弧形展布的生物礁沉积(沙庆安等, 1990; 刘宝珺和许效松, 1994; 焦大庆等, 2003), 向南出现台前斜坡— 较深水盆地的过渡地带(图 2), 因此台地南缘整体上具有镶边台地特征。沫阳、纳水剖面恰好位于台地到较深水盆地的过渡地带(图 2)。
 | 图 2 黔南地区中二叠世岩相古地理图(据刘宝珺和许效松, 1994修改)Fig.2 Lithofacies palaeogeographical map of the Middle Permian in southern Guizhou (modified from Liu and Xu, 1994) |
纳水剖面位于罗甸县罗苏乡纳庆村西南约5, km处, 沿罗甸至望谟312省道旁出露二叠系, 地层连续清晰。文中研究的阳新统茅口组主要为一套斜坡相沉积, 其下部以中— 厚层暗色粒泥岩— 泥粒岩为主, 中部粒度变粗, 主要为一套巨厚层砾屑灰岩, 上部为一套薄层灰泥岩— 粒泥岩, 吴家坪组底部泥岩、钙质泥岩覆于其上。
沫阳剖面位于罗甸县新沫阳镇与老沫阳村之间的柏油公路旁, 距新沫阳镇约1, km。二叠系沿沫阳河两岸出露。文中研究的阳新统茅口组下部和中部以薄— 中层灰岩夹薄层硅质岩为主, 含多层薄层砾屑灰岩, 上部主体为一套海绵礁灰岩, 顶部的薄层粒泥岩被吴家坪组底部泥岩、钙质泥岩覆盖。
在野外观察基础上, 对罗甸纳水、沫阳剖面薄片中的颗粒成分、基质类型、沉积组构、生物化石等进行了系统鉴定分析。岩石类型(以RT表示)划分及命名依据Dunham分类方案, 并将岩石特征与Flü gel(2010)分类方案中的标准微相(以SMF表示)特征进行了对比。
见海绵骨针、钙球等深水生物和少量棘皮碎屑, 生屑颗粒含量仅1%~8%, 其余均为灰泥基质(图 3-A)。与SMF1特征相似, 代表缓慢沉积的较深水环境。
 | 图 3 黔南罗甸地区纳水和沫阳剖面中二叠统岩石学特征(一) A— RT1, 基本不含生屑的灰泥岩, NS244-2; B— RT2-1, 以海绵骨针为主的粒泥岩, MY39-4; C— RT2-2, 以薄壳双壳为主的粒泥岩, MY70-4; D— RT2-3, 以钙球为主的粒泥岩, NS193-2; E— RT3-1, 含多种生屑颗粒的泥粒岩, NS201-1; F— RT3-2, 发育粒序层理的泥粒岩, 箭头指示正粒序, NS198-3; G— RT3-3, 生物掘穴泥粒岩, MY21-7; H— RT4-1, 生屑颗粒砾屑灰岩, MY30-4。Bi: 双壳, Br: 腕足, C: 钙球, F: 有孔虫, Fu:䗴, G: 腹足, T: 管壳石。单偏光, 比例尺代表1, mmFig.3 Petrological features of the Middle Permian at Nashui and Moyang sections in Luodian area, southern Guizhou(Ⅰ ) |
颗粒以远洋生屑为主, 且以单一种类的富集为特征, 生屑颗粒含量5%~70%, 基质为灰泥, 与SMF3特征一致。根据富集远洋生物种类的不同, 可将此类型分为3个亚型。
RT2-1以海绵骨针为主的灰泥岩— 粒泥岩。 颗粒以海绵骨针为主, 其他生屑见棘皮、有孔虫、钙藻、介形虫, 但含量相对较少, 破碎很严重(图 3-B)。
RT2-2以薄壳双壳为主的粒泥岩。 颗粒以双壳为主, 其壳壁很薄, 粒度相对其浅水同门类较小。有些片状双壳呈定向排列, 表现出纹理化微组构, 指示低能无扰动环境(图 3-C); 而有些壳体无序排列, 与棘皮、钙藻、腕足等的破碎颗粒混杂堆积在一起。
RT2-3以钙球为主的灰泥岩— 粒泥岩。 灰泥岩— 粒泥岩中颗粒以钙球为主, 兼有少量放射虫及泥晶化颗粒(图 3-D)。
生屑颗粒含量大于60%, 生物类型主要为棘皮、钙藻、有孔虫、䗴、腕足等, 有些还含少量海绵骨针、泥晶化颗粒。其中, 浅水源生屑颗粒较破碎, 磨圆通常较好。根据颗粒的排列特征可将此微相分为3个亚型。
RT3-1无特殊沉积结构的粒泥岩— 泥粒岩。 颗粒粒径比较均匀, 分选好, 未见特殊沉积结构(图 3-E), 多由颗粒流所形成。与SMF4特征相似。
RT3-2具有浊积特征的粒泥岩— 泥粒岩。 颗粒排列表现出特殊的沉积结构。有些亚型中颗粒定向排列; 有些颗粒自下而上变细, 发育正粒序层理(图 3-F); 还有一些颗粒的排列反映出多个不完整浊积序列的叠加。它们均有典型的浊流特征。对应SMF4中的异地浊积岩。
RT3-3生物掘穴泥粒岩。 生物掘穴产生的孔隙中填集了生屑, 使灰泥岩或粒泥岩中出现泥粒岩斑块, 颗粒成分以棘皮碎屑和似球粒为主(图 3-G)。与SMF1-生物掘穴灰岩特征一致。
砾屑可分为生屑颗粒和内碎屑2种, 基质以灰泥为主。
RT4-1生屑颗粒砾屑灰岩。 颗粒主要为䗴、钙藻、有孔虫、腕足、珊瑚、海百合茎等的碎片, 它们大多来自浅水环境。颗粒粒径超过2, mm者可达50%以上, 最大可达4.5, mm, 磨圆、分选差(图 3-H)。通常为颗粒流或风暴成因, 有些亚型中具有浊流沉积特征。与SMF5特征相同。
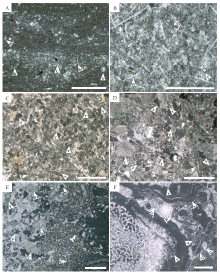
RT4-2内碎屑砾屑灰岩。 内碎屑种类多样, 砾径5~3000, mm, 其岩石特征是判断砾屑来源的重要标志。有些亚型中砾屑仅以灰泥岩为主, 无磨圆(图 4-A), 对应SMF4中单一岩屑成分的角砾岩; 有些亚型中砾屑则以粘结岩为主, 它们形状极不规则, 密集填集在一起, 基质含量很少(图 4-B), 与SMF6特征相同; 另一些亚型中砾屑成分多样, 既包含礁源粘结岩、颗粒岩, 又包含灰泥岩, 与SMF4中的复成分砾屑灰岩特征相同。
 | 图4 黔南罗甸地区纳水和沫阳剖面中二叠统岩石学特征(二) A— RT4-2, 灰泥岩砾屑灰岩, NS212-1; B— RT4-2, 粘结岩砾屑灰岩, NS219-1; C— RT5-1, 似球粒/颗粒发育泥晶套的颗粒岩, MY81-4; D— RT5-2, 含纹层的似球粒颗粒岩, 箭头指示纹层, MY82-12; E— RT6, 粘结岩, MY81-3; F— RT7-1, 硅化灰岩, 箭头指示未硅化完全的棘皮、钙藻碎屑, NS206-2; G— RT7-2, 含海绵骨针的原生硅质岩, MY27-4; H— RT7-3, 不含生屑颗粒的原生硅质岩, MY28-3。Ar: 古石孔藻, S: 海绵。 单偏光, 比例尺代表1, mmFig.4 Petrological features of the Middle Permian at Nashui and Moyang sections in Luodian area, southern Guizhou(Ⅱ ) |
RT5-1似球粒或颗粒发育泥晶套的颗粒岩。 该亚型中颗粒绝大多数发育泥晶套或完全泥晶化, 可识别出来的颗粒包括棘皮、腹足、有孔虫、钙藻, 它们以颗粒支撑的方式堆积在一起, 粒间孔隙被亮晶方解石充填(图 4-C), 指示动荡的环境, 与SMF11特征一致。
RT5-2含纹层的似球粒颗粒岩。 颗粒主要为泥似球粒, 粒度总体极小, 形状不规则, 多为次圆状, 它们以颗粒支撑方式紧密堆积, 被亮晶方解石胶结, 含有泥晶纹层(图 4-D), 可与利于陡坡稳定的SMF16-有纹层的似球粒颗粒岩的特征相比较。
以海绵为主的造礁生物被古石孔藻包覆, 另有管壳石、有孔虫、腕足等一同被粘附, 共同形成海绵藻礁(图 4-E), 与代表台地边缘礁的SMF7特征一致。
RT7-1硅化灰岩。 此微相中硅质岩主要由微晶石英构成(图 4-F), 反映了明显的硅质交代作用, 其原始岩石类型可能为多种, 硅化未完全者可恢复原始类型。
RT7-2含海绵骨针的原生硅质岩。 硅质岩中仅含海绵骨针一种生屑(图 4-G)。
RT7-3不含生屑颗粒的原生硅质岩。 硅质岩中不含生屑颗粒, 常发育水平层理(图 4-H)。
根据有关浅水碳酸盐沉积相模式, 上述岩石类型及特征清晰地反映了文中研究相带的沉积相构成。缓慢沉积的RT1, 含远洋生物的RT2, 代表浊流沉积的RT3-2及后期硅化形成的RT7-1, 组成本区盆地相(FZ1)沉积, 偶尔可见颗粒流成因的RT3-1。深水陆棚相(FZ2)包括含远洋生物的RT2, 生物掘穴形成的RT3-3以及多种硅质岩RT7, 偶尔可见RT3-1及生物含量极少的RT1。台前斜坡脚相(FZ3)在晴天背景下形成RT2和RT3-1, 风暴背景下主要形成代表风暴浊流沉积的RT3-2, RT4-1较少见。此外, 硅质岩RT7在此相带内也比较常见。砾屑灰岩RT4及泥粒岩RT3-1组成本区台前斜坡相(FZ4)沉积。台地边缘礁相(FZ5)主要包括颗粒岩RT5-1和粘结岩RT6, 含纹层的似球粒颗粒岩RT5-2仅在个别层位出现。
海平面变化和构造背景对台地边缘地层结构的控制作用已为大家所熟知。但是如何通过台地边缘沉积学研究揭示碳酸盐台地的演化, 则需要对碳酸盐沉积物的生产和堆积有较深入的了解, 也就是近年来逐渐为人们重视的碳酸盐工厂的概念和方法。浅水碳酸盐沉积物的生产, 主要受海洋生态条件的影响, 包括水体营养条件、古气候、海水化学条件等; 台地边缘的演化受碳酸盐生产工厂和生态可容空间变化的共同影响(颜佳新和严雅娟, 2014)。
大部分碳酸盐岩沉积物是由分泌碳酸盐的生物产生, 其中不少是光合作用的副产物, 因此碳酸盐的生产主要取决于光照程度。海洋50~100, m表层水体中, 特别在10, m深度内, 光合作用生物繁盛, 是碳酸盐生产的主要场所。这个具有高产率的浅水区域曾被形象地称为“ 碳酸盐生产工厂” (James, 1978)。自20世纪90年代以来, “ 碳酸盐生产工厂” 的概念逐渐得到深化, 已经识别出热带浅海工厂、灰泥丘工厂和温凉水工厂(Schlager, 2003)。骨屑颗粒组合分析是20世纪80— 90年代古气候分析的首选指标(Lees and Buller, 1972), 碳酸盐工厂分析方法的进一步完善, 使其焕发出新的活力, 成为环境营养条件分析和碳酸盐生产工厂识别的重要内容(Mutti and Hallock, 2003; Schlager, 2003; Pomar and Hallock, 2008)。
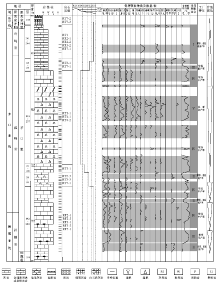
为探索茅口亚世上扬子碳酸盐台地南缘碳酸盐工厂的特征和演化, 在上述岩石类型分析的基础上, 本次研究对纳水和沫阳剖面茅口组地层进行了逐层系统的骨屑颗粒成分鉴定和统计(图 5; 图6; 统计内容为各采样位2.5, cm× 2.5, cm的单张薄片中各种骨屑颗粒的个数), 结合古生态和生境型分析, 建立了骨屑颗粒组合, 并进行了碳酸盐生产工厂分析。
 | 图 5 黔南罗甸地区纳水剖面茅口组综合地层柱状图Fig.5 Comprehensive stratigraphic column of the Maokou Formation at Nashui section in Luodian area, southern Guizhou |
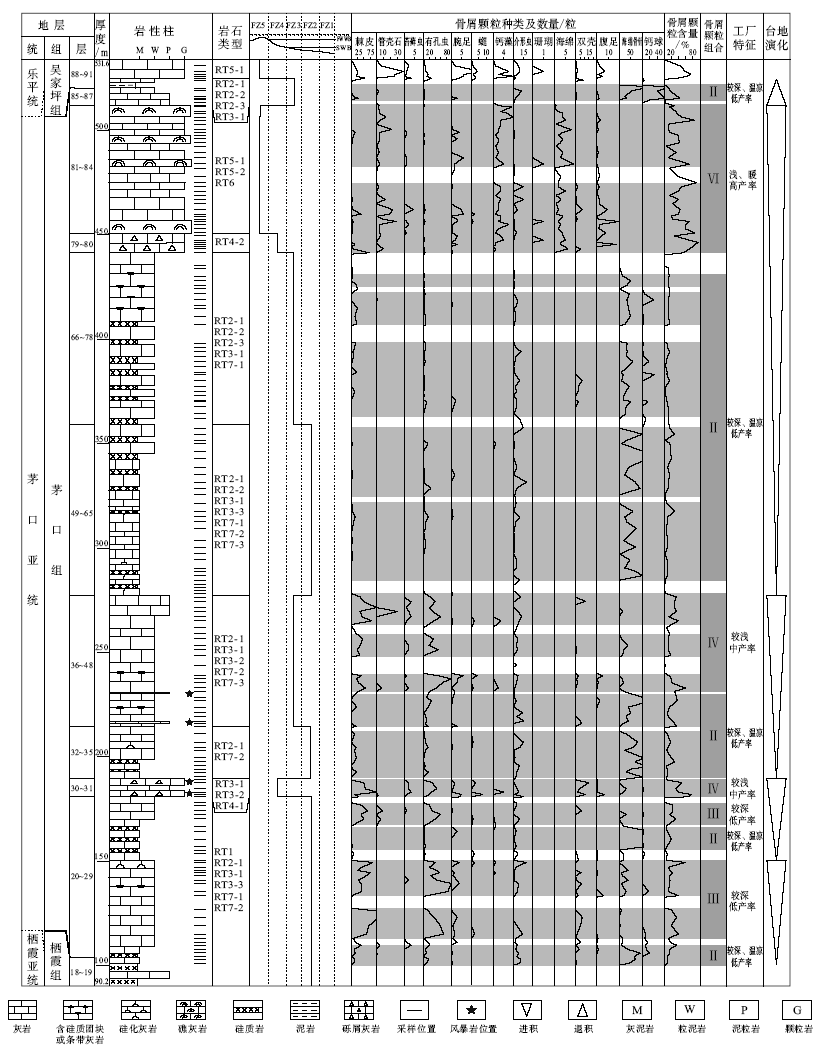 | 图6 黔南罗甸地区沫阳剖面茅口组综合地层柱状图Fig.6 Comprehensive stratigraphic column of the Maokou Formation at Moyang section in Luodian area, southern Guizhou |
以双壳为骨屑颗粒的主要成分, 其壳体小而薄, 属于典型的深水浮游类群。介形虫作为广适型生物门类, 其数量虽然较少, 但相比其他组合, 其含量明显增多且分布连续, 因此也被作为本组合的特征骨屑颗粒之一。有时还包含细砂级的棘皮碎屑以及代表深水环境的海绵骨针、钙球(图 7-A)。
 | 图7 黔南罗甸地区纳水和沫阳剖面茅口组骨屑颗粒组合特征 A— 双壳— 介形组合; B— 海绵骨针组合; C— 棘皮— 深水有孔虫组合; D— 棘皮— 苔藓虫— 浅水有孔虫组合; E— 棘皮— 绿藻— 组合; F— 海绵— 古石孔藻组合。Ar: 古石孔藻, Ba: 腕足, Bi: 双壳, By: 苔藓虫, C: 棘皮, Da: 粗枝藻, F: 有孔虫, Fu: 䗴, G: 腹足, O: 介形虫, P: 似球粒, S: 海绵, SS: 海绵骨针, T: 管壳石。单偏光, 比例尺代表1, mmFig.7 Features of skeletal grain association of the Maokou Formation at Nashui and Moyang sections in Luodian area, southern Guizhou |
该组合中骨屑颗粒几乎全部为深水异养型, 仅分布于纳水剖面顶部的较深水沉积环境中。盆地位于弱光— 无光带, 营养物质丰富但水体溶氧量较低, 因此该组合中骨屑颗粒总含量很低, 且以浮游型生物为主。它们零星分布于灰泥基质中, 碳酸盐产率极低。该组合反映出一种较深水、温凉水极低生物产率的碳酸盐生产工厂, 是台地淹没的关键识别标志。
以含有丰富的海绵骨针或钙球为特征, 常伴生细砂— 砂级的有孔虫、薄壳双壳、管壳石、海百合茎, 但数量相对较少。其中, 有孔虫包含广适型的Eotuberitina属及反映深水环境的节房虫(图 7-B)。
该组合分布于风暴浪基面以下的环境中, 海绵骨针、钙球都是典型的较深水环境生物, 前者往往还能反映较低的水温, 如加拿大北极地区二叠纪末的硅质海绵骨针组合和中下扬子地区茅口组的硅质海绵骨针组合(Beauchamp and Grasby, 2012)。贵州南部早二叠世仍然可能受到温凉水上升流的影响(Shi and Grunt, 2000), 但是否可以持续到中二叠世仍不得而知。部分骨屑颗粒, 如管壳石来源于浅水地区, 而浮游类型来自于表层水体。这样一种以异养型为主、多来源的骨屑颗粒组合, 是台前斜坡脚及以下地区较深水、温凉水低生物产率碳酸盐生产工厂的体现。
以棘皮和深水有孔虫的大量富集为特征, 此外还包含管壳石、绿藻、海绵骨针、钙球、腕足、苔藓虫、双壳(图 7-C)。
该组合中的代表性分子有孔虫主要包括营底内生活的Nodosaria属、Nodosinelloides属等, 它们发育细小的直列式壳, 壳壁薄, 结构简单, 是典型的深水相有孔虫。浅水来源的光能自养型生物— 绿藻出现较少。该组合中虽然同样含有海绵骨针及钙球, 但它们的含量和代表性远不及前述组合。这一组合中的骨屑颗粒呈现出多来源、异养型为主、兼有少量自养型的特征, 是较深水低生物产率碳酸盐工厂的体现。
含有丰富的棘皮和有孔虫, 苔藓虫相对较少。有孔虫以古串珠虫及小栗虫为主, 前者壳体呈柱状, 大而壁厚, 营底栖表生附着, 是浅水动荡、阳光充足条件下的典型分子, 后者则为局限环境的代表。此外, 该组合中还包含浅水来源的光能自养型生物粗枝藻、管壳石和珊瑚, 异养型生物腕足、三叶虫、壳体大而厚的软体动物以及深水来源的海绵骨针(图 7-D)。
该组合同样以异养型生物为主, 其特征分子海百合茎和苔藓虫, 常常出现于浪基面之下的沉积环境, 如广西来宾铁桥剖面栖霞组下部的BBC组合(沙庆安等, 1990), 也可以出现在水温相对较低、陆源碎屑物质影响较为明显的地方, 如保山地区二叠系永德组(颜佳新等, 2003), 是一种广适型生物分子, 广泛见于古代和现代非暖水碳酸盐沉积中。该生产工厂发育于营养物质丰富的弱光带, 相比组合Ⅲ , 本组合中浅水组分, 特别是浅水自养型组分的含量明显增加, 代表一种较浅水中等生物产率碳酸盐生产工厂。
仍含有丰富的棘皮类, 更为突出的是, 组合中浅水源骨屑颗粒的含量进一步增加, 尤以䗴和粗枝藻为代表。似球粒作为光能自养型组合的代表分子(James, 1997)在此组合中也大量出现。有孔虫以古串珠虫和壳体侧壁加厚、可抵抗较强水动力的厚壁虫为主。珊瑚以及古石孔藻包覆的海绵在该组合中同样较为常见。此外, 浅水环境中营底栖生活的腕足、软体等生物的含量较其他组合也有所增加(图 7-E)。
该组合中的特征分子, 绿藻和䗴, 广泛发育于古生代热带暖水碳酸盐台地沉积中(Beauchamp, 1994)。本次研究该组合见于纳水剖面的砾屑灰岩中, 砾屑虽然是异地沉积, 但它们很好地保留了该地区浅水生物碎屑滩生产工厂的记录。该组合中浅水光合自养型骨屑颗粒含量达到了纳水剖面茅口亚世的峰值, 结合其伴生有底栖异养型分子的特征, 认为该组合是一种浅水、暖水环境中台地边缘滩相高生物产率碳酸盐工厂的体现。
以钙质海绵及钙藻的大量富集为特征。该组合中常见古石孔藻包覆海绵构成的礁骨架, 具有华南地区茅口亚世生物礁的典型特征(范嘉松和吴亚生, 2005), 也是二叠纪特提斯热带地区常见的骨屑颗粒组合(Wahlman, 1985)。礁骨架间被大量细砂级似球粒和亮晶胶结物充填。除骨架生物外, 该组合还包含绿藻、海百合茎、苔藓虫、腹足及珊瑚(图 7-F)。此组合属于典型的光能自养型骨屑颗粒组合, 发育于阳光充足、营养物质较贫乏的台地边缘, 是浅水、暖水环境中台地边缘生物礁相高生物产率碳酸盐工厂的体现。
台地边缘相带是异地沉积非常发育的地区, 这在上述岩石类型识别中已经有较好体现。从本次探索实践看, 结合沉积特征和生态分析, 完全可以有效地区分原地/异地组分, 再造碳酸盐生产工厂。从本次对骨屑颗粒的统计结果来看, 骨屑颗粒虽然来源较广, 但是底栖/浮游分子、浅水/深水分子、自养/异养型分子、暖水/温凉水分子的组合在剖面中各段分异明显, 特别是异地特征型组分(反映来源特征)区别显著, 因此利用骨屑颗粒组合恢复台地边缘相带碳酸盐生产工厂同样是可行的。本次在研究区内共识别出6种骨屑颗粒组合, 论其多样性是其北侧的碳酸盐台地区和南侧的黔桂盆地不可比拟的, 这无疑为建立黔南地区茅口亚世碳酸盐生产工厂模式奠定了很好的物质基础(图 8)。
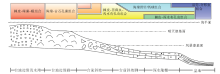
 | 图 8 上扬子碳酸盐台地南缘茅口亚世碳酸盐生产工厂恢复模式Fig.8 Model of carbonate factory reconstruction of southern margin of Upper Yangtze carbonate platform during the Maokouin Subepoch |
近20年来, 围绕茅口亚世海平面变化的研究, 无论是在黔桂地区(梅仕龙等, 1999b; 梅冥相等, 2002; 焦大庆等, 2003; Wignall et al., 2009), 还是扬子地区(李祥辉等, 1999; 王成善, 1999b), 甚至在全球范围(Haq and Schutter, 2008), 都认为孤峰期至早冷坞期是一个海平面下降的时期。近年来, 对茅口亚世峨眉山玄武岩的成因研究取得了重要进展。有研究认为由峨眉山玄武岩构成的峨眉山大火成岩省的成因与地幔柱隆升有关, 并且导致扬子碳酸盐台地西部茅口组上部遭受剥蚀(He et al., 2003, 2010)。若此, 茅口组地层序列也应该是向上变浅的。但是上述台地南缘的沉积微相及碳酸盐生产工厂分析结果表明, 上扬子碳酸盐台地南缘茅口亚世的演化过程并不完全如此。黔南地区自孤峰期开始沉积相和碳酸盐生产工厂变化明显, 在沉积序列上表现为由进积到退积的显著转变(图 5; 图6)。
纳水剖面沉积从浊流多发和含有深水生物的盆地相, 逐渐演化为滑移— 崩落和碎屑流极为发育的台前斜坡相。在此期间, 每一沉积旋回相对前一旋回, 其中的暖水分子和光能自养型分子比重逐渐增加。从以海绵骨针/钙球组合为代表的较深水、温凉水工厂, 逐步转变为以棘皮— 绿藻— 䗴组合为代表的浅水、暖水工厂。从沉积序列角度看是沉积环境水体逐渐变浅, 从碳酸盐台地演化角度看是台地向外(南)逐渐增生进积, 而且增生速度较高以致台前斜坡逐渐变陡, 直至发育一套巨厚层砾屑灰岩。同样的趋势在沫阳剖面也很清晰, 剖面下部的2组风暴岩分别记录了其台前斜坡相和台前斜坡脚相沉积背景。之后, 从富含海绵骨针的深水陆棚相, 经过碎屑流发育的台前斜坡相, 演化为台地边缘礁相。碳酸盐工厂则从最初的较深水、温凉水工厂逐步转为以海绵— 古石孔藻组合为代表的浅水、暖水工厂, 暖水和光能自养型分子含量的逐渐增多, 同样反映台地进积。因此当时台地顶部及边缘的热带浅水工厂状态良好, 为上扬子碳酸盐台地的增生进积提供了充足的物源。
至晚孤峰期, 纳水剖面中部巨厚层砾屑灰岩被一套薄— 中层灰泥岩— 粒泥岩覆盖(地质年代据史晓颖等, 1999), 台前斜坡相沉积逐渐转为盆地相沉积。此后, 沫阳地区生物礁被一套薄层含大量深水生物的粒泥岩— 泥粒岩覆盖, 转为台前斜坡脚相沉积。从碳酸盐生产工厂的角度来看, 2个剖面从热带浅水、暖水工厂分别转为以双壳— 介形虫组合、海绵骨针/钙球组合为代表的较深水、温凉水工厂。综合2个地区骨屑颗粒组合特征可以推测, 自晚孤峰期开始, 上扬子碳酸盐台地南缘由南(纳水)向北(沫阳)依次形成了一套表层浮游异养型(薄壳双壳)、底栖异养型(海绵骨针、介形虫、深水有孔虫)和表层自养型(管壳石)的生物组合, 地层记录中以温凉水异养型占绝对优势。碳酸盐生产工厂的这一转变, 可能预示了当时生态环境的急速恶化。直至晚二叠世长兴期, 随着热带浅水、暖水工厂的逐渐恢复, 华南地区生物礁得以繁盛, 上扬子碳酸盐台地的规模重新扩大。
上述自孤峰期开始的这种碳酸盐生产工厂的转变现象在华南地区有着广泛的代表性。在上扬子碳酸盐台地南缘(贵州紫云, 早冷坞期)(Wang et al., 2016)、北缘(四川上寺, 早冷坞期; 湖北猫耳山, 早冷坞期)(Zhang et al., 2007; Sun et al., 2008)及桂中孤立台地边缘(广西来宾, 早孤峰期)(姚尧等, 2012), 均是一套较深水沉积覆盖在浅水沉积之上, 沉积序列与文中研究的剖面极为相似(图 9)。四川上寺剖面早冷坞期出现了保存完好的浮游型薄壳双壳与菊石共生的生物组合(图 9)(Yan et al., 2008)。因此, 从孤峰期至早冷坞期, 整个台地范围是萎缩的, 台地边缘明显被淹没。
 | 图9 华南地区茅口亚世台地萎缩代表性剖面Fig.9 Representative sections showing shrink of platform during the Maokouin Subepoch in South China |
华南地区二叠纪位于赤道低纬度地区, 从碳酸盐生产工厂的角度看, 本区孤峰期至早冷坞期应该是有利于浅水台地顶部暖水碳酸盐生产工厂发育的时期。在此背景下, 健康的碳酸盐台地顶部生产工厂可以产生大量碳酸盐沉积物, 并通过向外运移使得台地得以向外扩展(进积), 出现向上变浅的沉积序列。上述碳酸盐台地萎缩、台地边缘被淹没、碳酸盐工厂的急剧转变, 应该与当时热带浅海碳酸盐工厂受到抑制, 甚至遭受破坏有关。
中二叠世晚孤峰期至晚二叠世吴家坪期是显生宙一个重大的地质历史转折期, 期间发生的一系列地质事件, 足以影响当时的生态系统和古海洋条件, 进而影响碳酸盐沉积的产生和堆积。特别是前乐平统生物灭绝事件(Stanley and Yang, 1994; 金玉玕等, 1995), 它与二叠纪末期生物大灭绝事件共同构成古生代末期两阶段式生物大灭绝事件(Yin et al., 2012), 殃及了当时65%~71%的海生物种(或58%属)(Erwin et al., 2002; Racki and Wignall, 2005), 尤其是对光合作用生物及与光合微生物共生的生物, 如钙藻、䗴、造礁珊瑚以及巨型双壳Alatoconchidae产生了重要影响(Isozaki and Aljinović , 2009; Groves and Wang, 2013)。很明显, 生物灭绝直接打击了当时浅水碳酸盐工厂的生产者。除此之外, 同时期也是地球环境, 如环境缺氧、古气候异常等(Bond et al., 2010a, 2010b; Isozaki et al., 2007; Yan et al., 2013)的急剧变化时期, 这些变化直接影响到当时碳酸盐沉积物生产者的生态条件, 或者说当时海洋生态条件是急剧恶化的, 极不利于碳酸盐沉积物的生产。因此, 这一时期生物灭绝和生态条件的恶化, 通过分别影响碳酸盐沉积物生产者和生产者的生态条件, 极有可能是华南地区碳酸盐台地萎缩的直接原因。期待后续研究能从成因角度有更进一步深化。
1)详细的野外和室内研究, 在贵州罗甸纳水、沫阳剖面茅口组地层中共识别出15种岩石类型及5种沉积相(盆地相、深水陆棚相、台前斜坡脚相、台前斜坡相和台地边缘礁相)。
2)通过系统地骨屑颗粒鉴定和统计, 在上述2个剖面茅口组中识别出了6种骨屑颗粒组合(双壳— 介形虫组合、海绵骨针/钙球组合、棘皮— 深水有孔虫组合、棘皮— 苔藓虫— 浅水有孔虫组合、棘皮— 绿藻— 䗴组合、海绵— 古石孔藻组合)。结合沉积特征和古生态分析, 恢复了其碳酸盐生产工厂的特征。
3)沉积相及碳酸盐工厂分析表明, 自晚孤峰期开始, 黔南地区碳酸盐生产工厂经历了由热带浅水、暖水工厂到热带较深水、温凉水工厂的明显转变, 表现为孤峰阶上部开始出现较深水沉积。
4)黔南地区碳酸盐台地边缘相带中二叠世孤峰期开始出现的碳酸盐生产工厂的转变现象, 在华南地区具有普遍性, 表现为台地萎缩、台地边缘被淹没。
5)中二叠世晚孤峰期至晚二叠世吴家坪期是显生宙一个全球重大地史转折期, 光合作用生物遭受重创、海洋生态环境急剧恶化。上扬子碳酸盐台地南部边缘孤峰期开始出现的碳酸盐生产工厂的急剧变化, 与当时热带浅海碳酸盐工厂被抑制或破坏有关。
致谢 野外和室内工作得到了李飞副教授、王夏博士、薛武强博士、严雅娟博士、蔡嘉骅硕士、武思琴硕士、甘元露硕士的帮助,审稿人提出了宝贵的建议,在此一并表示衷心感谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|