







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
塔里木盆地良里塔格地区奥陶系生物礁的沉积特征和成礁模式*
[孟苗苗1, 2, 3  , 樊太亮
, 樊太亮1, 2 , 卫端1, 2 , 王珊珊1, 2 ]
, 樊太亮|
|

第一作者简介 孟苗苗,女,1988年生,中国地质大学(北京)博士研究生,研究方向为碳酸盐沉积学理论与应用。E-mail: mmm1988@126.com。
塔里木盆地奥陶系生物礁储集层受到广泛关注。笔者优选塔里木盆地良里塔格地区奥陶系鹰山组顶部生物礁体( NYG-1)和一间房组底部生物礁体( NYG-2)为研究对象,通过详细的野外观察、实测和薄片鉴定,对生物礁的生物类型、岩性特征和沉积相单元进行分析。结果表明:塔里木盆地良里塔格地区鹰山组和一间房组的造礁生物主要由瓶筐石组成,与生物礁相关的岩性有障积礁灰岩、粘结礁灰岩、生屑粒泥灰岩、生屑泥粒灰岩、内碎屑泥粒灰岩、生屑颗粒灰岩、内碎屑颗粒灰岩和球粒颗粒灰岩。 NYG-1礁体发育礁基、礁核、礁顶、礁翼和礁盖 5个沉积相单元; NYG-2发育礁基、礁核与礁盖 3个沉积相单元。从生物类型来看,鹰山组的礁为托盘—障积礁,为近似圆形的台内点礁,展布无明显的方向性;一间房组的礁为托盘障积—粘结礁,属台缘礁,常呈现出礁体迁移的特征。鹰山组和一间房组成礁模式的共同点是都经历了礁基的形成、礁体的发育和礁体的衰亡 3个阶段,而差异主要表现在生物礁体的灭亡阶段: 对鹰山组生物礁而言,海平面的快速下降和高能水动力条件导致了瓶筐石的灭亡和内碎屑灰岩的产生;对一间房组生物礁而言,海平面的上升和浮游藻类生物大量发育造成造礁生物甁筐石的灭亡。以上研究对于建立符合塔里木盆地地区条件的礁体沉积模式和探索露头与井下生物礁对比研究有重要意义。
About the first author Meng Miao-Miao,born in 1988,is a Ph.D. candidate at the China University of Geosciences(Beijing). She is mainly engaged in the study of carbonate sedimentology. E-mail: mmm1988@126.com.
The Ordovician reef reservoirs in the Tarim Basin have been receiving more and more attention. The present research describes the reefs in Lianglitage area, Tarim Basin: NYG-1 reef (at the top of Yingshan Formation) and NYG-2 reef (at the bottom of Yijianfang Formation). Biotypes, lithologies, and sedimentary facies units of these two reefs were detailed documented, using detailed field observations, measurements, and thin section identifications. The results suggest that reef-building organisms are mainly composed of calathiums and that lithology associated with reefs is bafflestones, boundstones, bioclastic wackestones, bioclastic packstones, intraclastic packstones, bioclastic grainstones, intraclastic grainstones, and peloidal grainstones. Facies units in the NYG-1 reef mainly contain reef base, reef core, reef top, reef flank and reef cap. However, the NYG-2 reef consist only of reef base, reef core and reef cap. The Yingshan Formation is rich in calathium-baffle reefs, while Yijianfang Formation is dominated by calathium baffle-binding reefs. Reefs in Yingshan Formation are in the shape of circle-like and intra-platform patches, without obvious directional anisotropies. Reefs in Yijianfang Formation, in contrast, contain platform margin reefs that extensively show the vertical migration characteristics. Both formations have experienced three phases of evolutions, namely colonization, vertical aggradation, and capping. The difference lies mainly in the capping phase. More specifically, during the deposition of Yingshan Formation, the rapid decrease of sea levels and the high water energy led to the perish of calathium and the formation of grainstones, whereas the rise of sea levels and the development of considerable planktonic algae organisms caused the suffocation of calathiums. To conclude, this research provides insights into building reefs depositional models of the Tarim Basin and exploring new methods of comparative study on outcrops and underground reefs.
生物礁的研究对于地层的精确划分和对比、古环境和旋回性事件的定量恢复等具有重要的生态学和地质学意义。此外, 生物礁内各种孔隙、孔洞非常发育, 是石油、天然气理想的储集空间, 故地质历史时期形成的生物礁蕴藏着十分丰富的石油和天然气资源, 具有重要的经济价值。长期以来, 生物礁的概念和定义是学者们一直关注的理论问题: Newell等(1953)提出了生物礁的严格定义, 即其是由生物作用形成的抗浪构造; 20世纪70年代, Dunham(1970)提出礁的双重概念(地层礁和生态礁); 范嘉松(1996)认为, 生物礁是碳酸盐岩中一种重要的岩石类型, 其由大量的、各种各样的生物堆积而成, 或者是生物作用的产物; 而最新具有代表性的生物礁定义认为其是由固着生物所建造, 本质上是原地沉积的碳酸盐岩建造(Riding, 2002)。据研究, 奥陶纪是地质历史中生物礁发育的重要时期(Wilson, 1975; Scrutton, 1997)。塔里木盆地奥陶系发育大量的生物礁体, 2005年在塔中地区Ⅰ 号坡折带发现的中国第1个奥陶系超亿吨级礁型大油气田, 被美国石油地质家学会评为“ 2005年全球重大油气勘探新发现” , 开启了塔里木盆地生物礁型油气勘探新篇章。
目前对于塔里木盆地生物礁的研究, 主要集中在生物礁的发育层位和沉积相带(高志前等, 2005; 顾家裕等, 2005)、造礁生物和组合(朱忠德等, 2006)、生物礁不同储集相带以及储集层发育控制因素(贺萍等, 2003; 张兴阳等, 2004; 邓小江等, 2007; 焦养泉等, 2011; Jiao et al., 2012)、生物礁的野外识别和井下地球物理特征(张兴阳等, 2004; 邬光辉等, 2007)等方面。巴楚良里塔格地区生物礁露头发育, 前人已做了大量研究:如顾家裕等(2001)认为塔里木盆地的生物礁主要发育在中上奥陶统一间房组和良里塔格组, 该成果结束了塔里木盆地奥陶纪“ 无礁” 的历史, 并对造礁生物和附礁生物的类型、组合和分布特征进行了研究; 罗平等(2003)利用探地雷达和野外自然伽马能谱数据采集仪对良里塔格山两侧的生物礁规模和内部结构进行了分析; 李相明和杨申谷(2006)对巴楚地区中奥陶世生物礁的造礁生物进行了研究, 认为甁筐石是主要的建礁生物; 徐康等(2012)分析了一间房组礁滩体的岩石学、沉积学、古生物学和地球化学等特征, 认为礁滩体主要产出于一间房组, 以托盘类为主要造礁生物; 郑剑锋等(2015)对一间房组礁滩复合体进行了三维地质建模, 进而建立了微相控制下的孔隙度-渗透率模型, 提高了露头地质建模的精度。
以上研究主要集中在良里塔格地区一间房组的造礁生物和沉积特征方面, 虽然丰富了对塔里木盆地奥陶系生物礁的认识, 但仍存在3个主要问题: 一是对鹰山组生物礁的研究比较匮乏, 因为鹰山组生物礁数量较少, 以前没有发现; 二是缺乏对生物礁体从宏观到微观的精细刻画; 三是对生物礁成礁模式的探讨较少。而明确这些问题, 对于建立符合塔里木盆地的礁体沉积模式、探索露头和井下生物礁对比研究的新手段有重要意义。从出露良好的地表剖面着手, 容易建立生物礁体的岩性变化以及储集层变化的预测模式(Kerans et al., 1994; Barnaby and Ward, 1995), 基于此, 文中优选巴楚露头区良里塔格山出露的奥陶系生物礁体为对象, 对鹰山组和一间房组生物礁的生物类型、岩性和沉积相单元进行对比分析, 进而对鹰山组和一间房组的生物礁成礁模式进行探讨。
塔里木盆地位于新疆维吾尔自治区南部, 南邻昆仑山和阿尔金山, 北接天山, 面积达56× 104, km2。巴楚生物礁露头区位于塔里木盆地西北缘(图 1-a)。早奥陶世, 塔里木板块周缘大洋扩张逐渐减弱; 早奥陶世末期, 塔里木板块与周缘地块边界由离散型转变为聚敛型, 板块内部构造活动较弱, 仅有局部地区存在低幅隆起; 中奥陶世, 塔里木周缘处于挤压环境, 塔里木板块内部形成隆凹相间的构造格局(朱忠德等, 2006; 赵宗举等, 2009)。古地磁资料表明塔里木板块在奥陶纪处于南纬20° ~30° (方大钧和沈忠悦, 2001), 碳氧同位素分析表明塔里木板块的古温度为24~32, ℃, 因此塔里木地区奥陶纪处于热带— 亚热带气候, 有利于生物礁的发育(齐文同和范嘉松, 2002)。
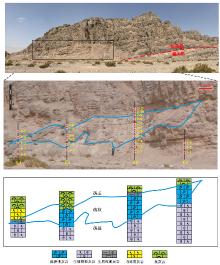
 | 图 1 塔里木盆地良里塔格地区中下奥陶统鹰山组和一间房组生物礁的剖面位置及综合柱状图a— 良里塔格地区在塔里木盆地的位置; b— 生物礁体观测点位置图, 主要的2个观测区域是南一沟地区(NYG)和一间房地区(YJF), 其中生物礁体的观测点有NYG-1、NYG-2、YJF-1和YJF-2。NYG-1生物礁是鹰山组沉积时期形成的, 后3者是一间房组沉积时期形成的; c— 良里塔格地区鹰山组和一间房组综合柱状图(据郭峰等, 2010, 有改动)Fig.1 Location of reef outcrops and stratigraphic column of the Middle-Lower Ordovician Yingshan and Yijianfang Formations in Lianglitage area, Tarim Basin |
奥陶系在良里塔格地区呈现为近南北向的宽缓背斜构造, 出露的地层包括鹰山组上部、一间房组、吐木休克组和良里塔格组(图1-b)。其中良里塔格地区的生物礁主要发育在鹰山组和一间房组。
鹰山组(O1-2y)沉积厚度达523, m, 可分为3段。下段为灰色— 浅灰色中— 薄层粉细晶白云岩、中细晶白云岩、内碎屑灰岩夹藻云岩、亮晶砂屑云岩和硅质岩, 水平层理和微波状层理发育, 属于半局限台地潟湖— 内碎屑滩沉积。上段为灰色— 深灰色中— 薄层亮晶内碎屑灰岩、泥粒灰岩夹粉细晶白云岩、藻灰岩和薄层或条带状硅质岩, 反映了半局限台地内碎屑滩与潟湖的交替环境(图 2-a)。顶部生屑灰岩发育, 见数个生物礁体(图1-c)。
 | 图 2 塔里木盆地良里塔格地区中下奥陶统鹰山组和一间房组生物礁野外照片a— NYG-2剖面点鹰山组上段的4个沉积旋回, 表现为黑灰色薄层泥粒灰岩、泥晶灰岩到浅灰色薄层颗粒灰岩的变化; b— YJF-2剖面点一间房组中段礁滩体特征, 礁滩体大量发育溶蚀孔Fig.2 Photographs showing reef outcrop of the Middle-Lower Ordovician Yingshan and Yijianfang Formations in Lianglitage area, Tarim Basin |
一间房组(O2yj)与下伏鹰山组呈整合接触, 厚69, m, 整体上为一套灰色、深灰色中厚层状生屑灰岩和内碎屑灰岩。下部以礁灰岩和生屑灰岩为主, 生物礁与生屑滩共生关系明显, 发育典型的进积型和退积型礁体, 为台缘礁滩相碳酸盐沉积(图 1-c; 图 2-b); 上部发育厚层含生屑泥粒灰岩。
良里塔格山北部东西两侧发育大量的生物礁体, 笔者优选出南一沟地区NYG-1和NYG-2生物礁体露头进行系统观察和取样, 并结合一间房地区 YJF-1和YJF-2生物礁(图 1-b)对礁体的叠置关系进行分析。对生物礁剖面NYG-1和NYG-2分别取5条和4条垂向测线进行详细实测, 并进行系统取样。取样时选取保留原始沉积信息的岩石样品, 尽量避开有明显后期方解石脉充填、风化、溶蚀和重结晶现象的岩石。采样间距为0.3~1, cm, 共采集49块样品(采样位置见图 3, 图 4), 其中鹰山组生物礁取样33块, 一间房组生物礁取样16块。对所有样品进行薄片磨制并鉴定, 从微观角度分析其古生物、岩石类型、沉积相单元特征。薄片在中国地质大学(北京)磨制, 采用德克萨斯大学奥斯汀分校的Nikon BX-P偏光显微镜进行观察。
 | 图 3 塔里木盆地良里塔格地区南一沟剖面鹰山组NYG-1生物礁的精细刻画Fig.3 Fine description of NYG-1 reef in the Yingshan Formation at the NYG profile in Lianglitage area, Tarim Basin |
 | 图 4 塔里木盆地良里塔格地区南一沟剖面一间房组NYG-2生物礁的精细刻画Fig.4 Fine description of the NYG-2 reef in the Yijianfang Formation of the NYG profile in Lianglitage area, Tarim Basin |
生物礁的生物组成可以分为造礁生物和附礁生物。造礁生物构成生物礁的骨架, 研究区造礁生物由瓶筐石、海绵类、苔藓虫、棘皮动物和钙藻类等组成; 附礁生物指自由生活在礁体中的生物, 是礁灰岩中沉积物的主要来源, 在生物礁体格架的建立中起到了积极作用, 研究区附礁生物主要有腕足类、三叶虫、有孔虫、介形虫和腹足类等。
1)瓶筐石。为主要造礁生物, 常以单体形式呈零散或团簇状出现。单体呈开头端朝上、锥端朝下的杯状, 开口多为圆形或椭圆形, 开口直径为2~10, cm, 单体长度为10~30, cm。体壁为具有网格状多孔的双层结构, 网格密集, 网孔粗大, 双层体壁以壁间的轴射状针杆连接(图 5-a, 5-b, 5-c)。瓶筐石多垂直发育, 显示原地保存状态, 能抵抗风浪, 起捕获和障积生物碎屑与灰泥的作用, 常与海绵类、葛万藻和苔藓虫等共生。
2)有柄棘皮动物。为次要的造礁生物, 主要包括海百合类和海林擒类, 其化石碎屑十分常见, 往往在礁基及礁翼等部位聚集成层, 成为生屑灰岩的主要成分(图 5-d)。几乎所有礁体的基底均由数十厘米到数米的粗粒级棘皮动物碎屑为主导的生屑灰岩构成, 表明有柄棘皮动物常常是格架生物造礁前重要的先驱群落分子。
3)海绵类。一般占造礁生物的10%左右。主要化石包括: Hudsonospongia minganensis, Archaeoscyphia minganensis(Billings), A. sp., Calycocoelia pro funda(Ivanov), C. sp.等。其中Hudsonospongia在研究区较发育, 为研究区造礁海绵类的特色之一。海绵类主要产于礁基及礁翼中, 营底栖固着型滤食生活, 适宜于清静温暖和流通性很好的正常浅海生活, 因其骨骼脆弱常以海绵骨针的形式存在于生物礁体之中(图 5-e)。
4)苔藓虫。在礁灰岩中总体含量低于5%。属种有Trepostomata sp., Batostoma sp., Hallopora sp., 其中Trepostomata常见。苔藓虫常呈枝状和包覆状群体特征, 其硬体底部匍匐于海底, 向上呈直立状生长, 起造架和障积作用。成年期的苔藓虫横切面呈圆形(图 5-f, 5-g), 主要生活在温暖洁净、有一定水体能量的浅海中。由于肢体细弱, 抗浪能力较差, 苔藓虫在以瓶筐石为主要造礁生物的生物礁中的造礁作用也受到限制。大多数苔藓虫呈零星的肢体碎片保存于礁灰岩中, 少量附在瓶筐石或石海绵外壁上生长。
5)钙藻类。主要是指皮壳状藻类, 如葛万藻、蠕孔藻以及一些隐藻类等。占造礁生物总量的5%左右, 局部地区可以富集到10%。这些藻类通过披覆和粘结作用等固定灰泥和生物碎屑, 增强了生物礁体的抗浪能力。葛万藻是呈管状藻丝的蓝藻, 由胶质依鞘外形成的钙质套呈隐粒结构, 直径约10, μ m, 大小均一, 无隔壁和侧枝孔, 管体相互缠绕, 使集合体成线形、团块形(图 5-h), 在礁中大多作为粘结生物附生和缠绕于瓶筐石和海绵类的体壁之外。蠕孔藻是与珊瑚藻亲缘关系十分密切的一类红藻, 其叶常呈瘤状, 内部由放射状紧密排列的丝体组成(图 5-i), 是一间房组主要的造礁生物之一, 常与瓶筐石共生。
附礁生物的生活方式多样, 有的是固着在造礁生物骨架上的, 如钙质绿藻和某些双壳类生物; 有的是游移的, 如腹足类和双壳类; 还有的是底栖的, 如腕足类、有孔虫和三叶虫等。
1)腕足类。主要为单个壳体, 由大小不一的双瓣组成。大部分情况下保存的是壳体碎片, 显微镜下可以看到低角度交叉的纤状或平行片状结构(图 6-a)。
 | 图 6 塔里木盆地良里塔格地区中下奥陶统鹰山组和一间房组附礁生物的特征a— 含生屑粒泥灰岩, 见腕足类(Br), NYG-1-1-3, 单偏光; b— 含生屑粒泥灰岩, 见三叶虫(Tr), NYG-1-1-3, 单偏光; c— 含生屑泥粒灰岩, 单列有孔虫(Fo)可见数个孔腔相连, 有孔虫左上侧相邻的条带为海百合茎碎屑(Cr), NYG-1-1-3, 单偏光; d— 含生屑泥粒灰岩, 见介形虫(Os), NYG-1-2-1, 单偏光; e— 含生屑泥粒灰岩, 见腹足(Ga)、腕足(Br)、海百合(Cr)、海绵骨针(Sp)和三叶虫(Tr)等, NYG-1-5-1, 单偏光; f— 障积礁灰岩, 见完整的腹足化石(Ga), NYG-2-2-5, 野外照片, 参照物笔的长度为12, cm。其中, NYG样品来自于南一沟剖面Fig.6 Photos and microphotographs showing reef-inserted organisms of the Middle-Lower Ordovician Yingshan and Yijianfang Formations in Lianglitage area, Tarim Basin |
2)三叶虫。未见完整的三叶虫化石, 仅在显微镜下见到三叶虫的各种切面, 如头甲、胸甲和尾甲等。其呈奇特的彩带状、鞍状以及勾状等(图 6-b), 属活动的底栖海洋性生物, 高盐度环境中不能生存。
3)有孔虫。主要由低镁方解石组成, 外形呈球状、锥状、管状、纺锤状等, 按照房室排列有单列、双列和三列之分(图 6-c)。
4)介形虫。由2个形状相似的壳瓣组成, 常呈蛋卵形、纺锤形、半椭圆形和梨形等。介形虫大小通常不超过1, mm, 已脱离的完整壳体中充填亮晶方解石, 壳瓣显示出细微的柱状结构(图 6-d)。
5)腹足类。常呈球形、锥形、塔形和帽型等, 螺壳大小一般为5~40, mm, 最小仅1, mm, 最大达200, mm, 在薄片上表现为一大一小相连的2个旋环(图 6-e)。野外也见有直径约10, cm的腹足类化石(图6-f)。
碳酸盐岩类型及特征是分析沉积环境的重要依据(Flü gel, 2004)。通过对NYJ-1和 NYJ-2生物礁详细的野外观察、系统取样和薄片鉴定, 结合碳酸盐岩颗粒类型、基质类型、沉积组构和生物化石等特征, 并依据Embry 和 Klovan(1971)、 Folk(1962)的碳酸盐岩定名方法, 共识别出8种岩石类型: 障积礁灰岩、粘结礁灰岩、生屑粒泥灰岩、生屑泥粒灰岩、内碎屑泥粒灰岩、生屑颗粒灰岩、内碎屑颗粒灰岩和球粒颗粒灰岩。
1)障积礁灰岩。含有原地和准原地骨架, 由原地块状瓶筐石等造礁生物组成坚固骨架, 由藻类等组成准原地骨架, 原地和准原地骨架的组合对灰泥基质和附礁生物等起到障积和捕获作用。
2)粘结礁灰岩。主要是通过原地生物的胶结作用形成的, 原地生物以钙藻类和钙质海绵等为主, 常构成礁顶的包壳粘结岩。藻粘结礁灰岩主要发育在一间房组生物礁顶部(图 7-a)。
3)生屑粒泥灰岩。以泥晶胶结为主, 先期较强的水动力使生物破碎, 后期滞留在安静环境中。生物碎屑包括腕足类、三叶虫、腹足类、双壳类和有孔虫等(图 6-a, 6-b, 6-c)。
4)生屑泥粒灰岩。颗粒特征较生屑粒泥灰岩明显, 颗粒之间主要被灰泥充填, 也可见少量细粒的亮晶方解石散布于灰泥基质中, 可能为重结晶的灰泥或细小生物碎屑。岩石总体呈颗粒支撑结构。颗粒依然以生物碎屑为主, 生屑包括棘屑、腕足类和海绵骨针等, 大部分生物碎屑破碎且杂乱堆积, 不具有成层性(图 7-c)。除生物化石外, 岩石中还可见少量球粒和藻团块等颗粒。
5)内碎屑泥粒灰岩。主要由极细内碎屑和泥晶组成, 其中内碎屑呈次圆到圆状, 分选较好, 粒径平均约0.1, mm, 颗粒边界模糊。颗粒之间泥晶充填, 局部亮晶胶结。生屑较少, 可见蠕孔藻、球松藻、介形虫和棘屑等(图 7-f)。
6)生屑颗粒灰岩。主要由棘皮动物碎屑组成, 亮晶方解石胶结。棘皮动物中主要见海百合茎, 其呈不规则颗粒或球粒, 分选差; 偶尔含少量双壳类、介形虫等生物碎屑(图 7-d, 7-e)。
7)内碎屑颗粒灰岩。是一种以内碎屑为主要组分的异化颗粒灰岩, 亮晶胶结。内碎屑颗粒磨圆度好, 分选中等, 偶含少量介形虫等生物碎屑。内碎屑颗粒灰岩野外呈薄层状产出(图 7-g)。NYG-1礁核顶部的颗粒分2个粒级, 分别为 0.2~0.5, mm和1~2, mm, 反映出其遭受到2期水动力的影响(图 7-h); 长条形颗粒有一定的方向性, 占颗粒总量的30%。而在礁侧缘, 颗粒分选磨圆较好, 粒径0.1~0.2, mm, 含有少量磨圆较好的生物碎屑, 相较于礁核顶部具有较弱的水动力条件(图 7-i)。
8)球粒颗粒灰岩。是通过胶结、凝聚或蓝藻黏液粘结的碳酸盐岩, 亮晶方解石胶结。颗粒或球粒不规则且无内部结构, 颗粒边界模糊。偶尔含双壳类和介形虫等生物碎屑(图 7-b)。
NYG-1礁体长6, m、高2, m, 形态上略呈低幅度的穹隆状。根据岩石类型、颗粒类型、含量及粒度、沉积结构和构造特征, NYG-1礁体可划分为5个沉积相单元, 自下而上分别是礁基、礁核、礁顶、礁翼和礁盖(图 3)。
礁基为生物礁的生长基座, 横向上连续稳定。岩性主要是以棘皮动物碎屑为主的生屑颗粒灰岩, 其次是含生屑泥粒灰岩。棘屑以海百合茎为主(图 7-d, 7-e), 粒径介于0.5~2, mm之间, 占生屑总量的70%~80%。其他生屑(双壳类和介形虫等)含量为5%~10%, 并含有10%~20%的亮晶胶结物。
礁核为生物礁的主体, 造礁生物以瓶筐石和粘结藻类为主(图 5-a), 附礁生物有腕足类、介形虫、有孔虫、腹足类和三叶虫等营底栖生物。在礁格架间发育有泥— 微晶球粒、生屑和内碎屑等滞留沉积。瓶筐石主要发育在礁核底部, 长× 宽介于1cm× 3cm到3cm× 6cm之间。瓶筐石含量从礁核底部向上部和翼部递减。
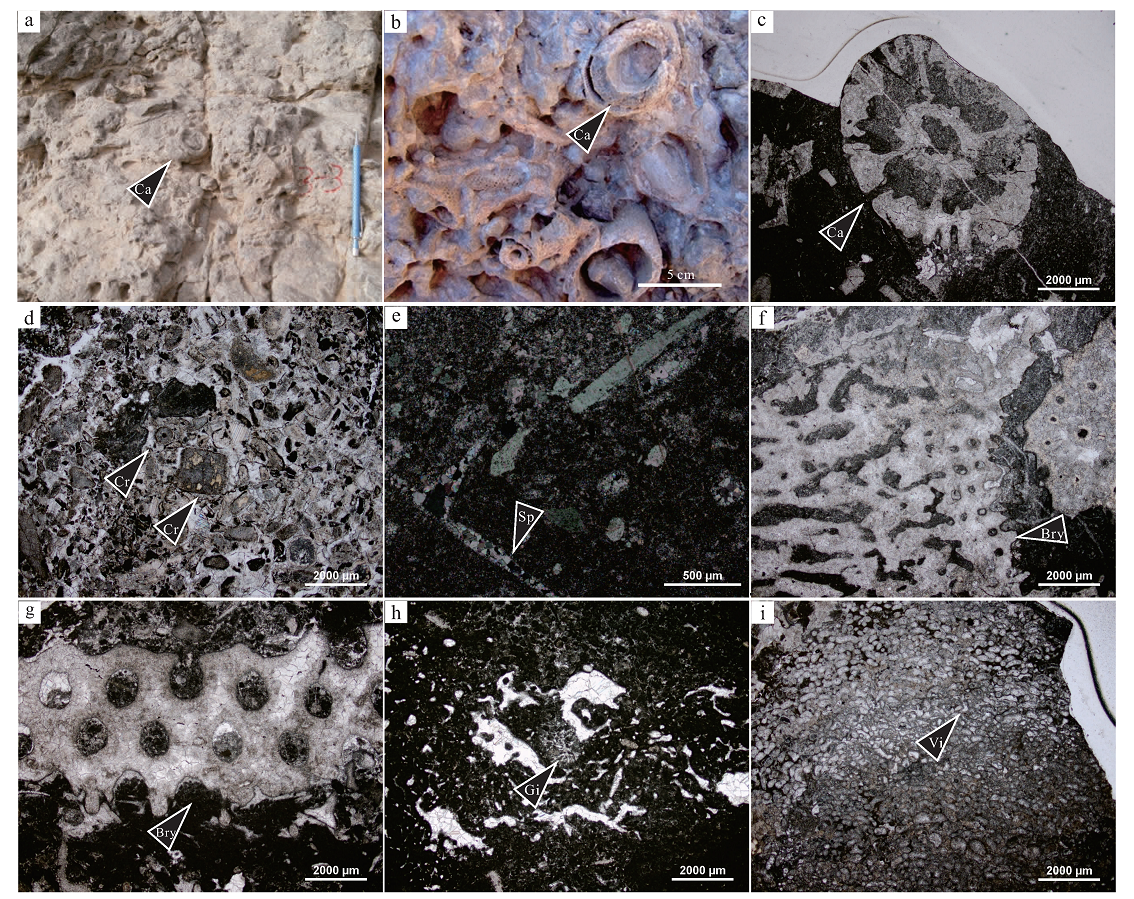 | 图 5 塔里木盆地良里塔格地区中下奥陶统鹰山组和一间房组造礁生物的特征a— 障积礁灰岩, 见瓶筐石(Ca), NYG-1-3-3, 野外照片; b— 障积礁灰岩, 见簇状发育的瓶筐石(Ca), YJF-1, 野外照片; c— 障积礁灰岩, 瓶筐石横截面(Ca)上见双层体壁以壁间的轴射状针杆连接, NYG-2-2-10, 单偏光; d— 棘屑灰岩, 海百合茎(Cr)多呈圆形、半圆形、长方形和多边形等, NYG-2-2-1, 单偏光; e— 含生屑泥粒灰岩, 见海绵骨针(Sp), NYG-1-1-3, 正交光; f— 障积礁灰岩, 见苔藓虫(Bry), 切面上可见卵圆形的虫管、花瓣状的间隙孔以及小而圆的刺孔, NYG-2-2-7, 单偏光; g— 障积礁灰岩, 见苔藓虫(Bry), NYG-1-2-1, 单偏光; h— 藻粘结灰岩, 见钙藻类葛万藻(Gi), NYG-1-3-3, 单偏光; i— 粘结礁灰岩, 见钙藻类蠕孔藻(Vi), NYG-2-2-11, 单偏光。其中, NYG样品来自于南一沟剖面, YJF样品来自于一间房剖面Fig.5 Outcrop photos and photomicrographs showing reef-building organisms of the Middle-Lower Ordovician Yingshan and Yijianfang Formations in Lianglitage area, Tarim Basin |
礁顶位于礁核上部, 为灰色— 深灰色薄层状, 主要由泥晶灰岩和生屑泥粒灰岩组成。生屑含量为5%~10%。
礁翼发育于礁核的四周, 下部与礁基接触, 上部与呈层状藻灰岩组成的礁盖有明显的突变接触关系(图 8)。主要由生屑颗粒灰岩和含生屑泥晶灰岩组成。颗粒分选较差且没有成层性。从礁翼不同位置样品的薄片鉴定分析可知, 从礁顶至礁缘方向生屑含量变少、泥晶成分增多。
 | 图 8 塔里木盆地良里塔格地区南一沟剖面鹰山组NYG-1生物礁L2测线的岩石学特征a— 测线L2的宏观照片以及取样位置, 测线位置见图 3; b— 生屑灰岩, 生屑见海绵骨针(Sp)、介形虫(Os)、腕足类(Br)等, 泥晶胶结, 样品2-1, 单偏光; c— 含生屑泥晶灰岩, 见腕足类, 泥晶胶结, 样品2-2, 单偏光; d— 含内碎屑生屑灰岩, 生物以棘皮类海百合茎(Cr)为主, 亮晶胶结, 样品2-3, 单偏光; e— 内碎屑灰岩, 亮晶胶结, 样品2-4, 单偏光; f— 内碎屑灰岩, 亮晶胶结, 样品2-5, 单偏光; g— 内碎屑灰岩, 亮晶胶结, 样品2-6, 单偏光Fig.8 Outcrop photo and photomicrographs showing lithological characteristics of the Yingshan Formationof Line L2 at NYG-1 reef of NYG profile in Lianglitage area, Tarim Basin |
礁盖与下伏礁核呈过渡关系, 主要由薄层状亮晶颗粒灰岩组成, 见少量生物碎屑。原因可能是原来未固结的灰泥丘和生物碎屑等受波浪或潮汐再次搅动作用以及间歇性暴露作用, 形成大量磨圆度较好的内碎屑颗粒(图 8-e, 8-f, 8-g)。
NYG-2礁体长25, m、高5, m, 呈SE-NW向展布, 且礁体底部凹凸不平、顶部平缓。纵向上可分为礁基、礁核和礁盖3个相单元(图 4)。礁基以亮晶棘屑灰岩为主, 棘屑主要为海百合茎, 颗粒粗大, 风化后颗粒松散、表面凹凸不平(图 7-d)。薄片上棘屑大小为0.5~2, mm, 含量70%~80%, 分选磨圆差, 亮晶胶结(图 7-e)。礁核为生物礁的主体, 造礁生物以瓶筐石和钙质海绵为主(图 5-b), 附礁生物有腕足类和腹足类等营底栖生物, 在礁格架间有生屑泥晶灰岩和藻泥晶灰岩的滞留沉积。礁盖与下伏礁核界限明显, 主要由薄互层展布的灰色泥、亮晶藻砂屑灰岩、泥晶生屑灰岩及藻粘结灰岩组成。
 | 图 7 塔里木盆地良里塔格地区中下奥陶统鹰山组和一间房组岩性特征a— 藻粘结礁灰岩, 钙藻(Al), 泥晶胶结, NYG-2-3-3, 单偏光; b— 球粒颗粒灰岩, 藻粒(Al), 亮晶胶结, NYG-1-1-2, 单偏光; c— 生屑泥粒灰岩, 含腕足类(Br)和海绵骨针(Sp)等生物碎屑, 泥晶胶结, NYG-1-2-1, 单偏光; d— 生屑颗粒灰岩, 以海百合茎碎屑(Cr)为主, NYG-2-2-1, 野外照片, 参照物镜头盖直径 5, cm; e— 生屑颗粒灰岩, 以海百合茎碎屑(Cr)为主, 亮晶胶结, NYG-2-1-2; f— 泥粒灰岩, 泥晶胶结, NYG-1-1-7, 单偏光; g— 内碎屑颗粒灰岩, NYG-1-2-6, 野外照片, 参照物铅笔长10, cm; h— 内碎屑灰岩, 亮晶胶结, NYG-1-3-5, 单偏光; i— 内碎屑颗粒灰岩, 偶见介形虫(Os)和双壳类(Bi), 亮晶胶结, NYG-1-3-5, 单偏光。其中, NYG样品来自于南一沟剖面Fig.7 Outcrop photographs and photomicrographs showing lithological characteristics of the Middle-Lower Ordovician Yingshan and Yijianfang Formations in Lianglitage area, Tarim Basin |
基于野外对生物礁的精细刻画和分析, 文中从造礁生物特征、礁体结构、礁体数量和大小3个方面对鹰山组和一间房组界面上下的生物礁体的差异进行分析。
首先, 造礁生物特征存在差异。鹰山组和一间房组的造礁生物均为瓶筐石, 但鹰山组瓶筐石个体小, 托盘生物口径约2cm× 3cm, 长4~5, cm, 以单体形式生长, 部分瓶筐石呈倒伏状, 含量占礁核10%~30%; 一间房组生物礁中瓶筐石个体较大, 托盘生物口径5cm× 10cm, 长达20, cm, 呈簇状直立生长, 含量可达整个礁核的60%~80%。就个体大小而言, 一间房组生物礁是鹰山组生物礁的2倍以上。就整个礁核规模而言, 一间房组生物礁是鹰山组生物礁的3~5倍。
其次, 沉积相单元也不相同。鹰山组生物礁由礁基、礁核、礁顶、礁翼和礁盖5个部分构成, 一间房组生物礁礁翼和礁顶不发育。鹰山组生物礁礁盖由平行层理发育的颗粒灰岩组成, 说明礁体的灭亡受水动力影响较大, 但一间房组生物礁礁盖由藻粘结礁灰岩组成, 说明生物礁的灭亡一定程度上与大量发育的浮游藻类有关。
最后, 生物礁的特征各异。鹰山组生物礁呈点礁, 单个生物礁体长6~10, m、高1~3, m, 且大部分生物礁体呈缓丘状, 仅在鹰山组顶部发育, 规模小, 数量少。一间房组已发现的生物礁有70个, 点礁一般长20~40, m、高1~15, m, 其中宽度小于25, m的占70%, 高度小于5, m的占75%, 礁体宽度和长度之比为2~12, 平均为4.6。一间房组生物礁比鹰山组大, 具有迁移的特征(图 9; 焦养泉等, 2011)。
 | 图 9 塔里木盆地良里塔格地区一间房组生物礁的垂向加积和侧向迁移特征(剖面位置见图 1)黄色虚线代表生物礁体的边界, 黄色箭头代表生物礁体的迁移方向, YJF-1剖面点Fig.9 Vertical accretion and lateral migration of reefs of the Yijianfang Formaiton in Lianglitage area, Tarim Basin(profile lacation Shows in Fig.1) |
总的来说, 从生物类型来看, 鹰山组的礁为托盘障积礁, 一间房组的礁为托盘障积— 粘结礁。从礁体特征来看, 鹰山组的礁为近似圆形的台内点礁, 展布无明显的方向性; 一间房组的礁为台缘礁, 常呈现出礁体的迁移特征。
通常, 生物礁的发育可分为3或4个独立而连续的演化阶段(James and Debrenne, 1980; Marshall and Davies, 1982; Riding, 2002; Montaggioni, 2005; Grö tsch, 2009)。结合生物礁的宏观展布、岩性和生物组合特征, 对良里塔格地区鹰山组和一间房组生物礁的成礁模式进行了探讨, 共同点是生物礁都经历了礁基的形成、礁体的发育和礁体的衰亡3个阶段。对于前2个阶段而言其差异较小, 在礁体的衰亡阶段两者差别较大。
适宜的环境使得海百合类生物在砂屑滩上定居, 之后海百合类大量繁盛, 形成海百合丛林, 因为仍受水动力的影响且没有障壁格架, 所以海百合等棘屑往往被海浪、潮汐等水流作用破碎。由于水动力作用较下部砂屑滩沉积时期弱, 因此这些棘屑并未发生长距离搬运, 而是堆积形成了正向地貌单元。棘屑滩的出现改变了水深、微地貌, 增加了光照、氧气, 为造礁生物的发育奠定了物质基础。
在前期发育的棘屑滩基础上, 瓶筐石等造礁生物开始迅速繁殖, 与藻类等形成缠绕格架, 同时粘结和障积灰泥和砂屑、生物屑, 最后逐渐发育成障积丘体。礁体的障壁作用使得水动力减弱, 并在格架礁体间滞留灰泥, 这些灰泥进一步促进了藻类的发育和造礁生物的生长。生物作用、粘结作用和障积作用共同促进了礁体迅速生长, 进而形成较高的凸起地貌单元。
鹰山组生物礁礁体的衰亡过程是: 随着礁体的快速生长, 礁体顶部开始分叉、缩小, 抗浪作用锐减。当礁体顶部到达海平面位置时, 强大的水动力对礁体造成破坏, 这种破坏包括对已成礁体的破坏和对礁体发育的阻碍。此时发育较多的是生屑灰岩, 因礁核顶部水动力最强, 所以附礁生物和造礁生物碎屑往往滞留在礁体的翼部, 但由于前期礁体格架仍具有抗浪作用, 导致灰泥往往滞留在礁体的翼部, 且越远离礁核沉积的泥晶灰岩越多、生屑含量越少。随着水动力的破坏作用持续, 造礁生物和附礁生物完全失去了适宜的生存条件, 而这些正向的地貌单元对颗粒灰岩的发育极为有利, 因此就形成了以内碎屑为主的颗粒滩(图 10-a)。
一间房组生物礁礁体的衰亡过程是: 海平面的小幅度快速上升以及浮游藻类生物的繁盛, 使得浮游藻类的光合作用和呼吸作用加强, 藻类死亡后被喜氧微生物分解利用, 消耗大量的氧气, 在海平面一定深度形成相对厌氧环境, 进而导致生物礁体因为缺氧窒息而死。大量藻类覆盖在生物礁体之上, 形成藻纹层灰岩(图 10-b)。一间房组生物礁的衰亡是由海平面小幅度上升引起的原因有2个: 一是海平面的上升或者下降都对生物礁的发育有影响(沈安江等, 2001), 而并非只有常规认识的海平面下降才能导致生物礁的衰亡; 二是一间房组下段是一个海平面上升的过程(徐康等, 2012), 那么就单个生物礁序列来说也应该是一个海平面上升的结果。郑剑锋等(2015)也认为, 一间房组生物礁的衰亡是海平面上升作用的结果。因此, 笔者认为, 一间房组下段NYG-2生物礁体的衰亡是海平面上升和浮游藻类的大量发育造成的。
1)塔里木盆地良里塔格地区奥陶系鹰山组和一间房组的造礁生物主要为瓶筐石; 附礁生物主要有腕足类、三叶虫、有孔虫、介形虫和腹足类等。与生物礁相关的岩性有障积礁灰岩、粘结礁灰岩、生屑粒泥灰岩、生屑泥粒灰岩、内碎屑泥粒灰岩、生屑颗粒灰岩、内碎屑颗粒灰岩和球粒颗粒灰岩。NYG-1礁体发育礁基、礁核、礁顶、礁翼和礁盖5个沉积相单元, NYG-2礁体纵向上分为礁基、礁核和礁盖3个沉积相单元。
2)鹰山组瓶筐石的大小和数量远不及一间房组, 鹰山组瓶筐石是一间房组瓶筐石的先驱。从生物类型来看, 鹰山组的礁为托盘障积礁, 一间房组的礁为托盘障积— 粘结礁; 从礁体特征看, 鹰山组生物礁为近似圆形的台内点礁、展布无明显的方向性, 一间房组生物礁为台缘礁、常呈现出礁体的迁移特征。
3)鹰山组和一间房组生物礁演化的共同点是都经历了礁基的形成、礁体的发育和礁体的衰亡3个阶段。差异主要表现在生物礁体的衰亡阶段: 对鹰山组生物礁而言, 海平面快速下降造成的高能水动力条件导致了造礁生物灭亡; 对一间房组生物礁而言, 海平面的上升促使藻类生物大量发育, 使得造礁生物窒息而死。
致谢 感谢中国石化勘探开发研究院陈跃老师和中国地质大学(北京)高志前老师的野外指导, 感谢美国德克萨斯大学Ian Duncan 教授、李兆奇博士后和姜磊博士后对薄片鉴定和论文思路方面的指导。特别感谢国家留学基金委的资助与支持!
(责任编辑 张西娟; 英文审校 龚承林)
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|