





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
河南宜阳下三叠统刘家沟组微生物成因沉积构造演化及其对古环境变化的响应*
[邢智峰1, 2  , 周虎
, 周虎1 , 林佳1 , 王敏1, 2 , 郑伟1, 2 ]
, 周虎]
|
|

第一作者简介 邢智峰,女,1973年生,副教授,主要从事遗迹学和地球生物学研究。E-mail: xingzhifeng925@126.com。
通讯作者简介 郑伟,男,1979年生,副教授,主要从事沉积学和地球生物学研究。E-mail: zhengw99@hpu.edu.cn。
河南宜阳地区下三叠统刘家沟组红色砂岩中发育数十种微生物成因沉积构造( MISS),如皱饰构造、脱水裂痕、微生物席碎片等。按照其形态特征的演化可分为 4个不同的组合,由下往上依次为皱饰构造组合、隆脊构造组合、小型脱水裂痕组合和大型脱水裂痕组合,其分别反映了不同的古环境与古气候: ( 1)皱饰构造组合反映了弱氧化的温暖、潮湿环境;( 2)隆脊构造组合反映了富氧的温暖、潮湿环境;( 3)小型脱水裂痕组合反映了弱氧化的温暖、半干旱环境;( 4)大型脱水裂痕组合反映了富氧的炎热、半干旱—干旱环境。依据研究区的地球化学特征和岩石学特征,认为早三叠世宜阳地区的古环境由湿润、温暖、弱氧化向炎热、半干旱、富氧转变。两者比较表明,宜阳地区微生物成因沉积构造的发育与演化是对早三叠世陆相古环境细微变化的联动响应。
About the first author Xing Zhi-Feng,born in 1973,is an associate professor. She is engaged in researches of ichnography and biogeology. E-mail: xingzhifeng925@126.com.
About the corresponding author Zheng Wei,born in 1979,is an associate professor. He is engaged in researches of sedimentology and biogeology. E-mail: zhengw99@hpu.edu.cn.
There are dozens of Microbially Induced Sedimentary Structures(MISS)in the red sandstone of Lower Triassic Liujiagou Formation in Yiyang area,Henan Province,including wrinkle structures,desiccation cracks and mat chips. According to the morphology evolution of MISS,it is divided into 4 different fabrics,namely wrinkle structures fabric,petees fabric,mini desiccation cracks fabric and large desiccation cracks fabric from bottom to top. They reflect the nuance palaeoenvironment and palaeoclimate:(1)wrinkle structures fabric reflects the warm moist suboxic sedimentary environment;(2)petees fabric reflects the warm moist oxygen-enriched sedimentary environment;(3)mini desiccation cracks fabric reflects the warm semi-arid suboxic sedimentary environment;(4)large desiccation cracks fabric reflects the hot semi-arid oxygen-enriched sedimentary environment. The geochemical information also indicates that the palaeoenvironment in the Early Triassic in Yiyang area changed from warm and moist suboxic to hot and semi-arid oxygen-enriched. In conclusion,the development and evolution of MISS in Yiyang area were a response to subtle changes of continental palaeoenvironment in the Early Triassic.
微生物主要繁盛于后生生物贫乏的局限环境中, 缺氧、高温、低扰动等一系列极端条件促进了微生物的发育(Riding and Awramik, 2000; 梅冥相, 2014)。在地质历史时期中, 以蓝细菌为主的微生物在碎屑沉积物的基底繁殖, 分泌的胞外聚合物(EPS)彼此相互粘结、捕获外来的细小颗粒, 聚集生长形成呈层状致密的微生物席(Noffke et al., 1997; Gerdes et al., 2000; Noffke, 2000; Bose and Chafetz, 2009; Taj et al., 2014; Sarkar et al., 2016)。不同生命活动时期的造席微生物与沉积物的相互作用, 使得微生物席以特殊形态的层面构造保存在岩层中, 被称为“ 第五种原生沉积构造” , 即微生物成因沉积构造(Microbially Induced Sedimentary Structures, 简称MISS)(Noffke et al., 2001, 2006)。Schieber等(2007)总结了微生物成因沉积构造的生长模式, 认为其形成主要有微生物席的生长(growth)、代谢(metabolism)、破坏(destruction)、腐烂(decay)与成岩(diagensis)5大过程。微生物的生命活动贯穿了整个生命演化历史, 微生物成因沉积构造不仅只发育在前寒武系(Noffke et al., 2003; 史晓颖等, 2008a), 而且在生物大灭绝时期的下三叠统(Chu et al., 2015; 于水情等, 2015)、下侏罗统(Porada and Bouougri, 2007; Petetffy, 2016)、上白垩统(Wilmeth et al., 2014)等地层中均广泛发育, 且整体呈“ 灾后泛滥” 的特点。
微生物成因沉积构造的形态与结构演化给微生物与环境系统变化的研究提供了重要的宏观信号, 是当前地球生物学的重点研究方向之一(Noffke, 2010; 陈留勤, 2013)。在晚二叠世— 早三叠世生物大灭绝后, 硅质碎屑岩地层中保存了形态各异的微生物成因沉积构造(Pruss et al., 2004), 成为了生物复苏时期环境压力变化的标志。换句话说, 保存在硅质碎屑岩中的微生物成因沉积构造见证了大灭绝后生态系统重建的全过程, 是大灭绝时期生态环境的重要组成部分。研究微生物成因沉积构造在P-T生物大灭绝后的发育演化, 对研究P-T大灭绝后生物复苏期的古生态变化有重要的解释意义。
河南宜阳地区石千峰群孙家沟组— 刘家沟组中的一套红色含砾中— 细粒砂岩沉积隶属于上二叠统与下三叠统(张利伟等, 2014), 属于陆相三角洲— 湖泊的过渡沉积, 发育了种类丰富、保存完好、数量众多的微生物成因沉积构造, 如皱饰构造、干涉波痕、微生物席碎片、气隆构造、脱水裂痕等(Tu et al., 2016; 于水情, 2014)。由于微生物成因沉积构造在不同的沉积层位类型不同, 故通过对微生物成因沉积构造种类的统计与层位对比, 可反映当时的环境气候变化, 还原宜阳地区早三叠世生物复苏期的古生态。
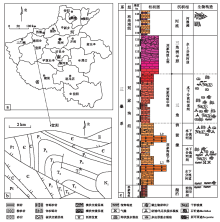
河南宜阳地区位于华北地台南缘, 属洛伊盆地宜阳凹陷区。晚二叠世扬子板块的向北俯冲与华北板块的向南俯冲造成相互挤压、抬升, 导致宜阳地区形成了陆缘近海的凹陷盆地(杨文涛, 2009), 沉积了一套以红色砂岩建造为主的三角洲— 河流沉积。研究区微生物成因沉积构造发育的剖面位于宜阳县城南部大雨淋村的河谷内(图 1), 出露良好, 自下而上依次为:上二叠统孙家沟组上部发育一套紫红色泥页岩与灰绿色薄层灰质泥岩互层沉积, 属于浅湖相。下三叠统刘家沟组可分为3段, 下部发育一套红色的中细粒砂岩沉积; 中部发育一套红色细粒砂岩与粉砂岩互层沉积, 属三角洲前缘亚相, 根据粒度度化又将其划分为水下分流河道和水下分流河道间2个微相; 上部粒度逐渐变粗, 发育一套厚层红色中砾砂岩和灰红色含砾粗粒砂岩沉积, 属于三角洲平原的水上分流河道沉积(朱筱敏, 2008)。上覆地层和尚沟组与刘家沟组呈整合接触, 为一套紫红色— 红色粉砂岩与泥岩互层沉积, 部分层位发育钙质结核, 并见Skolithos、Scoyenia遗迹化石(胡斌等, 2015), 是典型的河流沉积, 属河漫滩微相。在研究剖面上, 微生物成因沉积构造广泛发育的位置是孙家沟组的顶部、刘家沟组的下部与中部(图 1-c)。
按照微生物席的变化响应过程, 微生物成因沉积构造分为生长、代谢、破坏、腐烂与成岩5个阶段, 不同阶段发育不同特征的微生物成因沉积构造。按Schieber等(2007)的分类方案对研究区发育的微生物成因沉积构造分类后发现, 宜阳地区的微生物成因沉积构造主要有微生物席生长相关构造、微生物席破坏相关构造以及少量的微生物席腐烂(代谢)相关构造。值得注意的是, 微生物席的生长与发育是一个连续的过程, 其阶段没有明显的分界性, 某一种构造可能是多个过程的产物, 因此部分类型之间很难被分开。在垂直层面上, 还能看到微生物席以层状的形式保存在岩层中, 与沉积物形成具有明显明暗分层的微生物席层(图 2-f)(邢智峰等, 2011)。
 | 图 2 河南宜阳地区下三叠统刘家沟组皱饰构造组合a— 皱饰构造组合发育层位(7~21 m); b— 肯尼亚波痕, 是微生物席局部聚集生长而形成的似波痕构造, 无方向性, 圆拱状的脊高2~3imm; c— 微生物席碎片, 保存在砂岩层面之上, 厚度1~2imm, 主要成分是泥岩, 是微生物席被外力破坏后原地埋藏形成的, 图中笔长12icm; d, e— 象皮构造, 脊高2~3imm, 多与波痕共生; f— 微生物席纹层, 是微生物席多次迭代生长而形成的纹层, 厚度不等, 在黑色箭头处有明显凸起Fig.2 Wrinkle structures fabric of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
微生物成因沉积构造属于层面构造, 对其宏观演化特征的研究主要是对出露的层面处发育的微生物成因沉积构造种类进行统计(即分异度)。在研究剖面上, 由下而上主要发育12种微生物成因沉积构造, 分别是:象皮构造(elephant skins)、肯尼亚波痕(kinneyia ripples)、微生物席砂片(mat chips)、簇状凸起(tufts)、瘤状突起(postules)、生长脊(growth ridges)、砂火山(sand volcanoes)、卷边构造(rolls-ups)、纺锤状脱水裂痕(spindle-shaped desiccation cracks)、树枝状脱水裂痕(tree desiccation cracks)、网状脱水裂痕(net desiccation cracks)、多边形脱水裂痕(polygonal desiccation cracks)。这些构造由下至上分异成4种不同的组合。
2.1.1 皱饰构造组合(wrinkle structures fabric) 在研究剖面的7~21im处(图 2-a), 皱饰构造与小型微生物席碎片发育, 形成皱饰构造组合。
研究区发育的皱饰构造(Hagadorn and Bottjer, 1999; Porada and Bouougri, 2007)主要有2种: 一种是以脊背呈尖棱状凸起为主要特征的“ 象皮构造” (图 2-d, 2-e), 另一种的脊凸起呈圆拱状, 又被称为“ 肯尼亚波痕” (图 2-b)。前者尖棱状的脊在水平方向生长、交错汇聚形成尖峰, 席层围绕着尖峰周围又分散开来形成面积大小不一的不规则多边形, 脊高为1imm左右, 尖峰处的峰高于脊, 峰高2~3imm; 往往与波痕共生, 在波峰、波谷处均有发育, 有时会将波痕夷平。后者圆拱状的脊四散生长, 凹凸不平, 凸起的脊背与下凹处过渡圆滑, 在局部区域呈现一定的“ 聚集生长” 特性(郭荣涛等, 2012); 脊峰间距较小且沿某一方向平行排列, 但总的看来同一层的肯尼亚波痕的脊峰间距的变化依旧是毫米级, 相邻脊峰的平均间距为4~11imm。
微生物席碎片(图 2-c)呈碎屑状, 长轴长度4~6imm, 共出现2次, 均与皱饰构造共生, 证明该组合中的微生物席碎片是同期发育的部分皱饰构造被破坏后原地沉积形成的。
2.1.2 隆脊构造组合(petees fabric) 在剖面的25~43im处(图 3-a), 发育了多种微生物席成因沉积构造, 其中隆脊类构造占大多数, 并与波痕共生, 称为隆脊构造组合。
 | 图 3 河南宜阳地区下三叠统刘家沟组隆脊构造组合a— 隆脊构造组合发育层位(25~43 m); b— 瘤状突起, 呈圆拱状, 未连结成脊, 高1~2 mm; c— 簇状突起, 尖脊高2~3 mm; d— 网状生长脊, 脊高3~4 mm; e— 放射状隆脊和瘤状突起, 被黑色虚线框住的部分为隆脊的聚集部位, 条形虚线为向其聚集的脊, 瘤状突起分布在右下区域; f— 砂火山和逃逸构造, 黑色框中为砂火山构造的放大图, 黑色虚线是典型的气体逃逸构造Fig.3 Petees fabric of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
当微生物席某一点过量生长或是微生物群落生命活动释放出的气体汇聚在席底某一点使席层表面隆起时, 其以瘤状(图 3-b, 3-e)、簇状(图 3-c)的形式表现在层面上。随着微生物群落的繁盛和生命活动越来越频繁, 隆起相互连通, 就形成放射状(图 3-e的黄线区域内)、星状的隆起, 如星状脊、帐篷脊等(Noffke et al., 2003)。研究区发育的隆脊构造多为簇状凸起, 隆起高2~3imm, 直径5~8imm; 部分簇状凸起聚集后形成的不规则生长型隆脊轴长为20~30imm, 但产出较少。层面发育的波痕受微生物席生长活动的干涉而被夷平, 波谷内发育有簇状凸起或生长脊(图 3-d), 在微生物成因沉积构造活动较弱的区域波痕又重新显现。
此外, 在27im处还发现微生物席腐烂相关构造(图 3-f)。微生物腐烂分解后释放出的气体在沉积物表面作用形成的构造被称为微生物席腐烂相关构造。在研究区, 较为稀疏的席层破裂形成类似雨痕的气体逃逸构造(图 3-f的白线区域内), 而较厚的席层破裂后在重力作用下向外侧翻卷, 经成岩作用后石化成类似火山口状的“ 砂火山” 构造(图 3-f的黑框区域内)。
2.1.3 小型脱水裂痕组合(mini desiccation cracks fabric) 在剖面的60~80im处(图 4-a), 主要发育卷边构造与纺锤状的脱水裂痕, 形成小型脱水裂痕组合。
 | 图 4 河南宜阳地区下三叠统刘家沟组小型脱水裂痕组合a— 小型脱水裂痕组合发育层位(60~80 m); b— 卷边构造, 微生物席开裂后较薄的边缘脱水后上卷形成卷曲的边缘, 研究区发现的卷边构造边缘厚度为2~3 mm; c— 纺锤状脱水裂痕, 席层轻微开裂后形成的纺锤状裂口被外来的砂质填充而形成的脱水裂痕, 开裂方向多与波痕的走向平行; d— 多边形脱水裂痕, 大片的席层开裂成不规则块状, 席片间隙被填充而形成具网状的多边形脱水裂痕; e— 树枝状脱水裂痕, 微生物席从某一点呈扩散状开裂, 裂隙被填充后形成类似树枝状的脱水裂痕Fig.4 Mini desiccation cracks fabric of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
微生物席暴露一段时间后会脱水开裂, 裂开的缝隙被外来的沉积物填充, 因为被填充的沉积物多为砂质颗粒并多发育在砂岩中, 又被称为“ 砂裂” (Schieber et al., 2007)。纺锤状脱水裂痕的裂隙长10~30imm, 宽1~4imm, 脊高1~4imm, 多与波痕共生(图 4-c)。微生物席脱水开裂后, 由于开裂的边缘部分脱水后上翘并翻卷, 沿着被破坏的席表面边缘围成一圈, 形成独特的卷边构造(flip-overs)(图 4-b)。该层位发育的卷边构造卷起的边缘厚度小于3imm, 规模较小。裂隙被外来沉积物填充, 裂隙之间相互连通形成了多边形的(图 4-d)、树枝状的(图 4-e)脱水裂痕。
2.1.4 大型脱水裂痕组合(large desiccation cracks fabric) 在剖面的84~97im处(图 5-a), 脱水裂痕的形态与规模逐渐增大, 不再以小型的“ 纺锤状” 裂痕为主, 取而代之的是大型的、厚度较大的网状、曲线型的脱水裂痕, 多与波痕共生, 构成大型脱水裂痕组合。
 | 图 5 河南宜阳地区下三叠统刘家沟组大型脱水裂痕组合a— 大型脱水裂痕组合发育层位(84~97 m), 规模更大, 席层更厚, 表明此时微生物群落最为繁盛; b, c— 大型网状脱水裂痕, 脊宽10~20 mm, 脊高5~15 mm; d— 正弦状脱水裂痕, 在波谷中沿波痕走向发育, 脊高2 mm, 脊宽4 mm; e— 脱水裂痕底部, 并无“ V” 字型底; f— 曲型脱水裂痕, 较厚的席层开裂后由于脱水作用使得边缘萎缩而聚集形成圆弧状, 随后裂隙被填充从而形成曲型— 圆弧状的脱水裂痕, 在其下层还发育有皱饰构造Fig.5 Large desiccation cracks fabric of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
在该组合中, 微生物席发生大规模的开裂, 裂隙之间相互连接形成不规则网状(图 5-b, 5-c)。该层位的脱水裂痕脊宽10~20imm, 脊高5~15imm, 形成的网格大小不一, 形态与“ 泥裂” 类似, 但纵切面底部一般呈弧形或矩形(图 5-e), 并不发育向下收敛的“ V” 字形底(邢智峰, 2010)。微生物席在波谷中的选择性生长还使其发育了呈正弦状的脱水裂痕(图 5-d), 平均脊宽12imm, 平均脊高15imm; 在波痕层面上还发育有曲线型脱水裂痕(图 5-f), 这类脱水裂痕弧度大, 几近于圆弧状, 但砂脊高度较低, 为2~5imm, 宽度8imm, 并与其他类型的脱水裂痕共生。
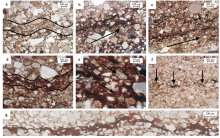
在发育微生物成因沉积构造的岩层中有明显的明暗纹层变化, 微生物席所粘结的沉积物颗粒较小, 一般小于围岩的粒径, 形成了1层或数层的细小颗粒层(图 6-a)(Noffke et al., 1997; Schieber, 2004; Schieber et al., 2007)。偏光镜下, 不透光的黏土矿物丝状体包绕着以粉砂为主的细小颗粒, 在水平面上平行排列, 具定向性(图 6-b, 6-c)。以黏土矿物为主的丝状体呈波曲状起伏且厚度不一(图 6-d), 较厚的地方应是微生物群落富集的区域, 且其厚度变化也有可能受太阳黑子活动周期的影响(Tang et al., 2014)。丝状体并非连续地生长, 有时以条状和点状的形态保存在岩层中(图 6-f)。当黏土矿物沉积较多时, 能够形成丝状体(filament), 当沉积物颗粒相对较大时, 微生物席能够捕获并固定沉积物使其保持定向排列。微生物席层所捕获的矿物主要以石英、长石为主, 能够观察到少量的方解石, 此外在丝状体边缘能看到被包绕的红色呈间断式分布或线状分布的绢云母(图 6-e)。根据微生物席层的生长习性来看, 应是微生物通过对有机质的降解或捕捉从水流中获得的(邢智峰等, 2011; 郑伟和刑智峰, 2015)。丝状体的厚度并不均匀, 微生物席群落富集的区域丝状体呈圆拱式的凸起(图 6-g), 可由凸起方向判断微生物席层的顶底。
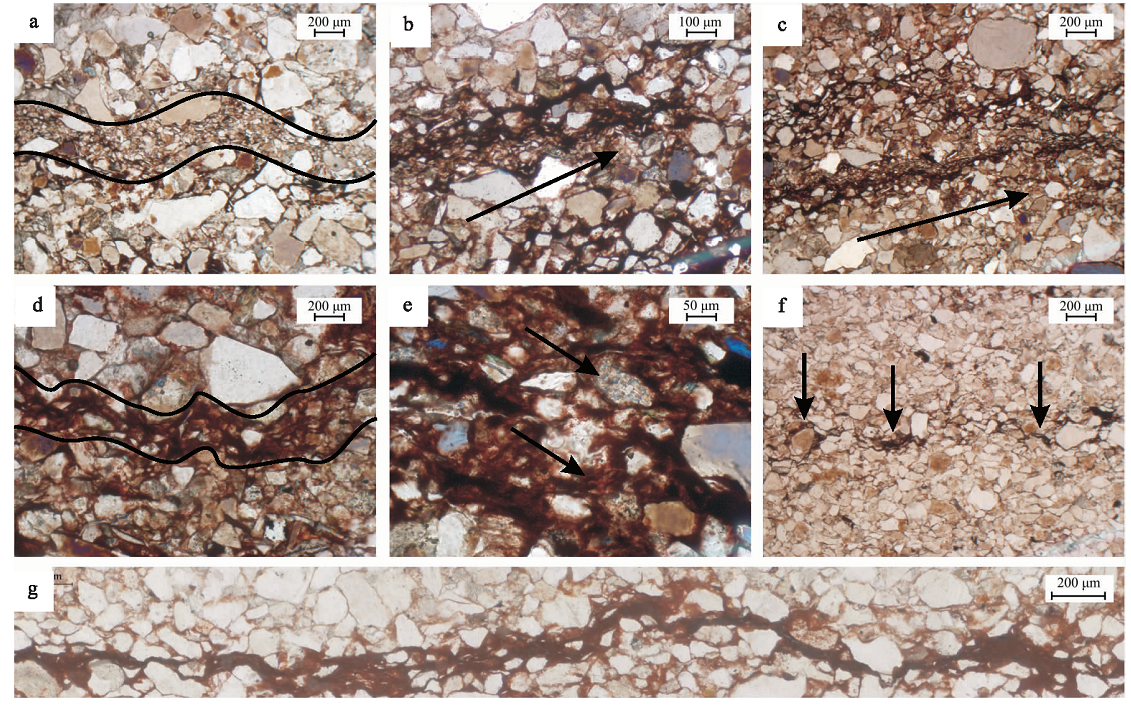 | 图 6 河南宜阳地区下三叠统刘家沟组微生物成因沉积构造显微特征a— 细小颗粒层, 黑线区域内可明显观察到粒度变化; b— 定向颗粒与暗层, 黑色箭头为颗粒排列方向, 暗层周围的颗粒呈小型化; c— 定向颗粒层, 较小的颗粒也在微生物席的粘结作用下具有定向性, 黑色箭头为颗粒排列方向; d— 波曲层, 微生物席代谢活动形成的凸起或凹陷呈不规则的波动型曲线; e— 暗层丝状体中的绢云母与红色的疑似赤铁矿(黑色箭头处); f— 不连续丝状体(黑色箭头处); g— 连续的具圆拱形凸起的丝状体Fig.6 Microscopic features of Microbially Induced Sedimentary Structures of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
沉积岩中的常量元素与微量元素含量一方面取决于岩石的性质, 另一方面也受到沉积时的气候条件、水动力条件的控制(郑德顺等, 2017)。因此, 通过测定沉积岩的常量与微量元素的含量和比值, 可还原岩石沉积时期的古气候信息, 如古盐度、古温度等(Dasch, 1969; Getaneh, 2002; 田晓雪等, 2005; Kalugin et al., 2007; 曾艳等, 2011; 胡俊杰等, 2017)。
本次从研究区采集了12块刘家沟组样本、1块和尚沟组底部样本, 岩性以细砂岩、粉砂岩为主。剔除样品的风化、蚀变部分后, 将样品敲碎、清洗烘干后磨成200目以下的粉末待测试。常量元素的测试使用X荧光射线光谱仪(XRF), 微量元素的测试使用等离子耦合质谱仪(ICP-MS), 元素分析误差均小于5%。测试样品与数据均保存于河南省生物遗迹与成矿过程重点实验室。
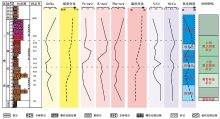
在对古气候的研究中, 盐度大小表明蒸发作用的强弱, 蒸发强度越大, 盐度越大(冯兴雷等, 2014)。Sr/Ba值通常用来表示古盐度的变化, Sr的迁移能力强于Ba, 随着远离湖(海)岸Sr/Ba值逐渐增大, 在淡水环境中小于0.61, 在半咸水环境中为0.61~1, 在海洋环境中大于1(熊小辉和肖加飞, 2011)。所测刘家沟组样品的Sr/Ba值在0.1~0.3之间, 平均值0.14(图7), 指示其是淡水环境。从纵向序列上看, 刘家沟组的B3、B8、B14的水体盐度相对较高, 反映了气候相对干燥、蒸发强度较高的气候特征。整体看来, 气候正在由湿润向干旱过渡。
V、Ni、Cr、Co等微量元素的丰度体现了沉积环境的氧化还原性质(Yang et al., 2004; Tribovillard et al., 2006; Wang et al., 2015; 胡俊杰等, 2017)。对氧化还原环境的判断可以用V/Cr值、Ni/Co值等参数比较出来(Jones and Manning, 1994; Rimmer, 2004; 苗建宇等, 2007; Meng et al., 2017)。 当V/Cr值小于2时指示氧化环境, 2~4.25指示贫氧环境。研究区的V/Cr值在0.40~1.0之间, 平均值为0.64, 表明当时刘家沟组氧浓度含量虽然有一定变化, 但整体是富氧的。Ni/Co值也可以表示沉积环境的古氧相, 一般来说小于5是氧化环境, 5~25表示缺氧环境。研究区刘家沟组的Ni/Co值在3.09~5.56之间, 平均值为4.39, 表明刘家沟组整体为富氧环境, 个别时期氧浓度含量出现下降。结合2条不同的氧浓度指标变化曲线来看, 早三叠世宜阳地区的氧浓度呈波动性上升, 整体处于氧化环境。值得注意的是, V、Ni、Co这些微量元素的含量还受到物源区的影响。在整个早三叠世, 华北板块南缘隆起, 发生大规模的海退现象, 整体形成了向北东方向倾斜的斜坡, 宜阳地区形成了一套以砂泥建造为主的陆源近海的湖盆沉积。水体以淡水为主, 物源来自西南方向的伏牛古陆和西北方向的中条古陆(王惠勇, 2006), 沉积物是典型的长距离搬运碎屑颗粒, 故V/Cr、Ni/Co的比较方法适用于此。
此外, Fe、Mg、K等常量元素的含量也对气候变化有一定的指示意义, 潮湿的环境下Fe元素易胶结成Fe(OH)3并快速沉淀在水底, 使得沉积物中Fe元素的含量相对较高, Mg、K等元素不易沉淀, 含量较低; 当温度上升环境趋于干旱时, 水分的蒸发导致含有Ca、Mg、K、Na、Ba等元素的盐类析出形成沉淀赋存在沉积物中, 使其含量增加(陈敬安等, 1999)。研究区Fe的含量在3~4img/g之间, 在刘家沟组中部有一小段呈上升趋势, 其余均为下降; K、Mg元素含量的波动并不明显, 整体呈增加的趋势, 说明其整体环境属于半干旱型气候, 中间有一段时间为暂时的湿润气候。
地球化学分析(图 7)表明, 研究区处于一个炎热、半干旱的氧化— 弱氧化环境中, 温度不断上升, 刘家沟组下部沉积时期气候相对温暖、湿润、富氧; 中部沉积时期氧含量略有降低, 温度升高, 湿度降低, 期间有短暂的波动; 上部沉积时期气温进一步上升, 从氧化环境过渡到了弱氧化环境, 最终变成了炎热、半干旱的气候。将地球化学数据变化与微生物成因沉积构造演化相互结合来看, 皱饰构造发育于湿润温暖、相对缺氧的环境, 隆脊类构造发育于半干旱、温暖、相对富氧的环境中, 小型脱水裂痕类构造发育在半干旱、炎热的氧化环境中, 大型脱水裂痕更适合发育在干旱、炎热的氧化环境中。
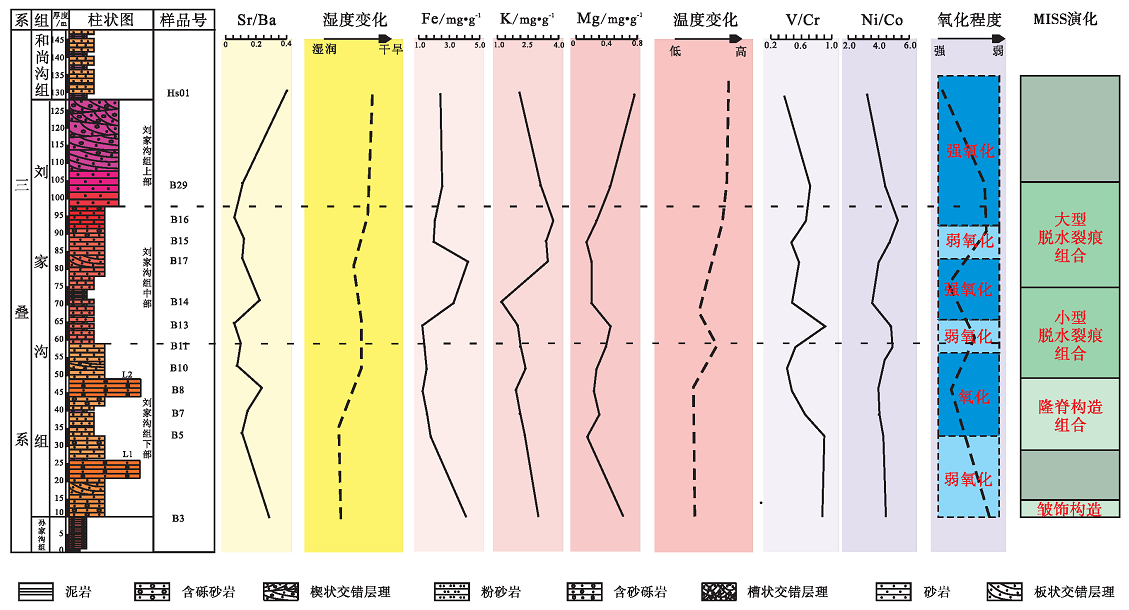 | 图 7 河南宜阳地区下三叠统刘家沟组地球化学数据变化与微生物成因沉积构造演化的关系Fig.7 Relationship between geochemical data and evolution of Microbially Induced Sedimentary Structures of the Lower Triassic Liujiagou Formation in Yiyang area, Henan Province |
Noffke等(2001)总结了微生物成因沉积构造在潟湖与无障壁海岸的分布情况, 指出潮坪环境是最适宜微生物成因沉积构造形成的环境。在高能的开阔潮坪带微生物席破坏构造广泛发育, 皱饰构造不发育或较少发育; 在相对低能的潟湖, 微生物成因沉积构造的类型比较丰富, 皱饰构造、气隆、生长脊等构造大量发育(史晓颖等, 2008b; 邢智峰, 2010; 李涛, 2011; 黄秀等, 2010)。
3.2.1 沉积相变化与微生物成因沉积构造组合的演化关系 宜阳地区的上二叠统— 下三叠统隶属于石千峰群, 作为典型的陆相地层, 其是一套以含砾砂岩为主的三角洲— 湖泊过渡沉积(图 1), 研究区剖面对应的沉积环境是湖相三角洲前缘带发育的网状河道。后生生物的贫乏、三角洲相复杂多变的水动力条件、较浅的水体以及气候的变化成就了微生物成因沉积构造繁盛的客观条件。
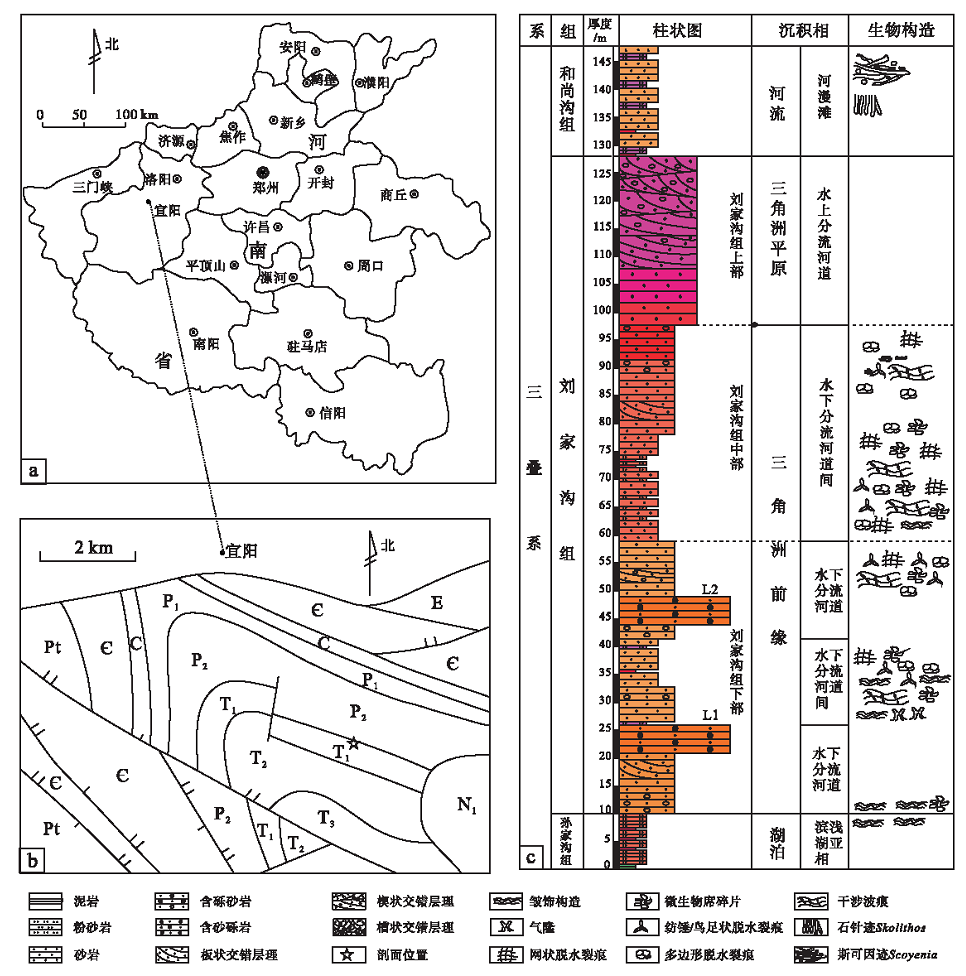 | 图 1 河南宜阳地区下三叠系剖面位置及综合地层柱状图Fig.1 Sketch showing section location and comprehensive stratigraphic column of the Triassic in Yiyang area, Henan Province |
从研究区微生物成因沉积构造所在的地层层序位置来看, 大量的皱饰构造、隆脊构造和气隆, 即微生物席生长相关构造, 发育于孙家沟组的上部和刘家沟组的下部, 主要产出在以含砾中细粒砂岩为主的岩层中。该层位平行层理发育, 并发育粉砂岩夹层, 属于湖相三角洲前缘的水下分流河道间微相, 其中水下河道由于冲积物侧向堆积而形成的水下浅滩, 充分的细粒沉积物供给, 给微生物生长提供了有利的物质条件, 形成了形态各异、种类繁多的微生物席生长相关构造, 构成了第1类“ 皱饰构造组合” 和第2类“ 隆脊构造组合” 。该层位与海相微生物席成因沉积构造相对应的沉积相应是潟湖和有障壁海岸。
在研究区的刘家沟组下部, 微生物席成长繁盛一段时间后, 水动力条件加强, 洪水事件发生, 形成暂时性水道, 大量具有同心纹层的砾石堆积。待水动力降低后, 细粒沉积物重新沉积, 微生物席的生长得以恢复。
在刘家沟组中部, 岩性以中— 细粒砂岩为主, 粒度由细变粗, 含有泥砾和粉砂岩夹层, 微生物成因沉积构造开始了从以生长相关构造为主到破坏相关构造的过渡, 平行层理逐渐被交错层理取代, 部分层位还发育有双向交错层理。沉积相由具有局限性的水下分流河道间微相向开阔的三角洲平原亚相转变, 水体更浅。微生物席破坏相关构造广泛分布于该组层序中, 纺锤状、鸟足状、网状、波曲状的脱水裂痕和厘米级的微生物席碎片与层面的波痕共生, 形成了独特的层面构造。对应的微生物成因沉积构造组合为“ 脱水裂痕组合” , 根据脱水裂痕的种类变化与厚度大小, 将其分为小型脱水裂痕组合和大型脱水裂痕组合2类。总的来说, 脱水裂痕的规模在逐渐扩张, 厚度不断增加, 表明微生物席的暴露时间更长、吸附的沉积物更多、对环境的改造程度越来越大, 与无障壁海岸中保存的微生物成因沉积构造种类相似。
从宜阳地区微生物成因沉积构的发育过程与沉积环境的变化关系来看, 微生物成因构造的演化呈现一定的阶段性。孙家沟组上部和刘家沟组下部发育了数量可观的以微生物席生长相关构造为主的皱饰构造、隆脊构造和气隆。随着水平面的下降, 部分微生物席短暂暴露, 所接触的外界温度升高, 席层中的水分蒸发, 部分席体开始收缩、坍塌、破裂、卷曲, 水流带来的新的沉积物将裂隙填充, 形成了形态各异的脱水裂痕与微生物席碎片。脱水裂痕取代了皱饰构造成为了微生物成因沉积构造的主要类型, 并且规模进一步扩大, 种类也在不断增多。随着外界温度的不断升高和水体进一步变浅, 越来越多的微生物席被改造, 出现了大规模的多边形脱水裂痕和卷边构造。在刘家沟组上部, 湖盆的收缩使其出露湖面, 成为了三角洲平原的水上分流河道, 水动力条件突然增强, 取而代之的是一套具槽状交错层理的灰色厚层含砾粗粒砂岩, 不再适宜微生物席的发育。
从古地理环境上来说, 宜阳地区的微生物成因沉积构造主要发育在细砂岩及粉砂岩的层面上, 并且多与波痕共生。彼时的宜阳地区处于三角洲— 湖相的过渡环境中, 水动力适中且水流方向多变, 上游所携带的大量沉积物也给微生物席的发育提供了物质基础。相对封闭的水下浅滩环境中, 微生物席接受水体的保护, 微生物生长相关构造发育较多。在水体更浅处, 尽管微生物席层更厚, 但其在枯水期易暴露出水面, 使得微生物席脱水、开裂、卷曲, 被沉积物填充而形成大量的微生物席破坏相关构造(Shi et al., 2008; Mata and Bottjer, 2009; Slowakiewicz et al, 2016)(图 8)。
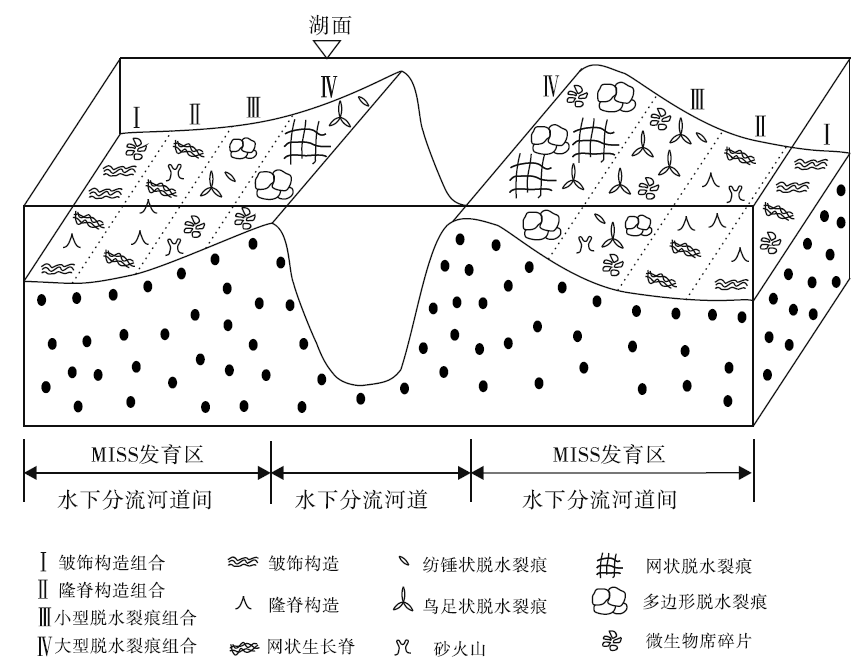 | 图 8 陆相微生物成因沉积构造分布与沉积相的耦合关系Fig.8 Coupling relationship between sedimentary facies and distribution of continental Microbially Induced Sedimentary Structures |
从古气候变化上来说, 宜阳地区早三叠世的整体气温是不断上升的, 微生物席的出现指示了其半干旱的气候条件。微生物成因沉积构造的阶段性的幕式演化细致地描绘了宜阳地区早三叠世气候的变化过程, 即温暖半湿润— 温暖半干旱— 炎热半干旱。
3.2.2 微生物成因沉积构造演化对全球环境变化的响应 晚二叠世末期开始的印支旋回运动, 将各大板块汇集在一起形成泛大陆(Scotese, 1994)。古欧亚板块沿着东昆仑— 玛沁— 略阳深断裂带与昆仑— 秦岭— 大别山2条构造带将其分成了陆地与海洋(任纪舜, 1984)。华北板块、塔里木板块与西伯利亚板块的南部部分共同形成一个狭长的古陆, 发育了一系列大小不等的内陆河湖盆地, 与中国西南、华南地层区辽阔的特提斯洋构成了典型的“ 南海北陆” 格局。宜阳地区作为华北板块的最南缘, 是连接古特提斯洋与陆地的重要通道(图 9)。作为水系下游的入湖口, 物源供给充分, 即有北部火山喷发带来的火山灰, 又有西部河流上游带来的碎屑沉积物供给, 给陆相微生物成因沉积构造的形成提供了地理条件。在整个早三叠世, 出现了全球性的火山事件与贫氧事件。以西伯利亚火山活动为代表的全球性火山事件导致的气候温室效应越来越明显。岩浆的喷发和涌出导致了陆地上的大火事件, 大火的燃烧、喷发作用产出的火山灰、释放出的有害气体直接或间接地导致了陆地生物的死亡和灭绝。燃烧过程消耗大量的氧气, 生成CO、CO2并且放出大量的热, 进一步加速了大气圈的贫氧化和温室化。经牙形石实验测定表明二叠世末期的古海洋表面平均温度高达40i℃以上, 陆表平均温度甚至会在50i℃以上(程明, 2014)。
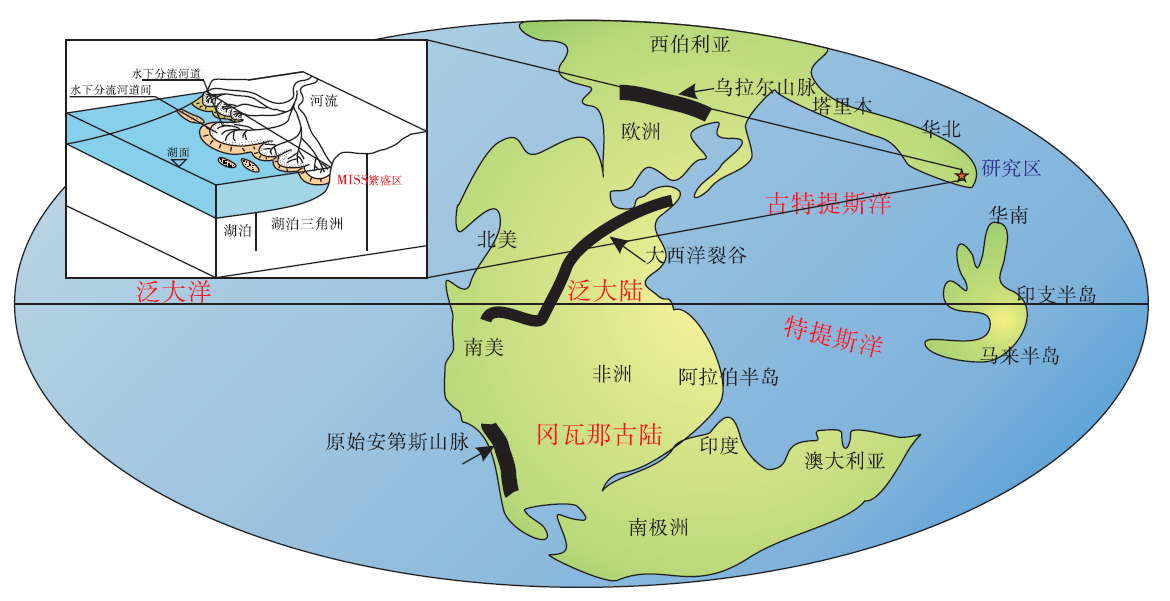 | 图 9 早三叠世河南宜阳地区的全球地理位置(据Scotese, 1994)Fig.9 Location of Yiyang area of Henan Province in globe geographic map during the Early Triassic(after Scotese, 1994) |
以蓝细菌为代表的微生物位于食物链的底层, 在后生生物繁盛的晚古生代一直处于弱势(殷鸿福和童金南, 1997)。在P-T大灭绝开始后, 由于大部分捕食者的灭绝, 西伯利亚火山喷发带来的大量矿物质和全球大火并缺氧的极端环境(沈文杰, 2007), 使得它们开始在浅滩处快速地繁衍、生长成席, 形成了一个相对封闭的“ 避难所” , 短时间内占领了生物圈, 成为复苏期单调生态系统的代表(罗根明等, 2014)。随着火山的休眠和构造运动的减弱, 在中三叠世— 晚三叠世, 整体极端环境得以改变, 大气圈氧浓度含量升高, 后生生物的生存环境改善, 微生物再一次在开阔的沉积环境中消失, 回归到了特殊的极端环境中, 生态系统又重新丰富了起来。
河南宜阳地区上二叠统— 下三叠统的陆相碎屑岩地层中发育了大量的微生物成因沉积构造, 发育的微生物席生长相关构造有皱饰构造、簇状凸起和干涉波痕; 发育的微生物席破坏相关构造有脱水裂痕、卷边构造和微生物席碎片, 脱水裂痕尤为发育。从下至上又可划分为4个不同的组合, 表明微生物成因沉积构造演化的不同阶段。三角洲— 湖泊的过渡相沉积环境给微生物群落的发育提供了适中的水动力条件和充足的物质基础, P-T之交的大灭绝杜绝了后生生物对微生物群落的捕食, 微生物成因沉积构造才得以广泛发育于此。相应地, 微生物席对外来沉积物的粘结、捕获和对基底的夷平作用, 弱化了水流, 改变了沉积环境。微生物成因沉积构造形态与规模的演化能够体现出沉积环境细微的变化, 及时对外界环境的变化产生反馈, 温度、水流、气候条件的精细变化都会不同程度地影响微生物席的生长, 有助于对研究区沉积环境的细分。
微生物成因沉积构造在整个剖面的生长变化情况和地球化学数据同时表明: 在P-T之交的大灭绝后, 宜阳地区的气温正在逐渐升高, 气候也由湿润向干旱过渡。陆相沉积环境中的微生物群落在整个时期都很活跃, 其沉积产物也呈阶段性的变化, 具有一定的波动性。这一细小的波动性变化可以反映出沉积环境的细微改变, 微生物成因沉积构造的演化与沉积环境的变化具有相互耦合的动态平衡。
陆相P-T界线也是近年来研究的焦点之一, 然而由于陆相环境的局限性, 生物化石并不能很好地保存, 锆石测年的程序繁琐精度也还有待提高(刘兵等, 2014; 赵英利等, 2016)。海相的P-T界线位于微生物岩层的底界, 若将微生物生态系统的短暂性繁盛作为P-T大灭绝时期的多幕灭绝(童金南和殷鸿福, 2009)中的生物复苏标志(沈树忠等, 2010), 那么碳酸盐岩中的微生物岩与陆源碎屑岩环境中的微生物成因沉积构造应是等时的, 可以将微生物成因沉积构造的首现作为陆相P-T界线的辨别标志。多年以来, 陆相P-T界线的位置一直存在争议, 微生物成因沉积构造作为一种易于识别、在后生生物大灭绝时期分布广泛、存在时间短的原生沉积构造, 对陆相P-T界线的厘定具有极为重要的参考价值。
(责任编辑 张西娟; 英文审校 白国平)
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|