





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
重庆巫溪红池坝长兴阶深水相沉积记录及地质意义*
[杨巍1  , 张廷山
, 张廷山2, 3 , 杨洋2, 3 , 毛凯楠1 , 张华鑫2 ]
, 张廷山|
|

第一作者简介:杨巍,男, 1985年生, 2016年毕业于西南石油大学,获工学博士学位,现为贵州理工学院副教授,主要从事沉积学与古生态学研究。E-mail: rexswpu@163.com.。
重庆巫溪红池坝剖面长兴阶上部地层出露齐全,产系列牙形刺和深水陆棚相遗迹化石。通过建立牙形刺带,对该剖面长兴阶沉积相类型及特征进行了精细研究,并讨论了区域大隆组 /长兴组界线的时代归属。长兴中—晚期开始,沉积环境由深水盆地向深水陆棚逐渐过渡,海洋生态环境的不稳定性开始显现并持续发展,基于时间序列的遗迹化石记录详细刻画了深水陆棚水体含氧量频繁加速波动过程,为系统审视二叠纪末生物与环境协同演化提供了重要依据。
About the first author:Yang Wei,born in 1985,graduated and obtained his Ph.D. degree from Southwest Petroleum University in 2016. Now he is an associate professor in Guizhou Institute of Technology,with main interests in sedimentology and palaeoecology. E-mail: rexswpu@163.com.
The deep-water sediments were well developed along the Hongchiba section in Wuxi County,Chongqing,which is represented by a series of Upper Changhsingian conodonts and deep shelf trace fossils. Based on the assemblage zone of conodont observed from the Hongchiba section,the Upper Changhsingian sedimentary feature and facies were investigated,and the controversial viewpoints on the boundary between Dalong Formation and Changxing Formation were discussed. During the Middle-Late Changhsingian,the sedimentary environment shifted gradually from a deep-water basin to a deep-water shelf, giving rise to unstable marine ecological environment. The analysis of trace fossil records with time series points to the unstable fluctuation change of oxygen levels in deep-water shelf facies through the Middle-Late Changhsingian. It also reflects the interaction between biology and environment when the critical geologic interval has been approaching.
二叠纪末地球表层系统发生的系列变化, 如缺氧事件、海洋物理化学异常、大规模海退等(吴亚生等, 2006; Algeo et al., 2007; Luo et al., 2010; 殷鸿福和宋海军, 2013), 导致全球生物组成和生态结构发生剧变(戎嘉余和黄冰, 2014), 但目前关于整个生物危机事件型式依然存在较大争议(Yin et al., 2007; Shen et al., 2011; 郑全锋等, 2013)。基于时间框架的沉积记录研究对深入理解二叠纪末生物危机事件中海洋生态系统演化具有重要作用, 尤其是不同时空背景生态系变革与生物危机事件的对应关系的深入理解, 对全面认识二叠纪末生物危机过程具有重要作用。
扬子地区是二叠系— 三叠系全球界线层型剖面所在地, 近年来随着二叠纪末生物与环境事件研究不断深入, 二叠纪末深水相沉积演化过程受到广泛关注(何卫红等, 2015), 广元上寺、广元旺苍、兴山峡口、长兴煤山等已积累了较丰富的剖面资料(Yin et al., 2007; 房强等, 2012; Shen et al., 2013; Shen et al., 2014; 何卫红等, 2015; Xu et al., 2016), 为二叠纪— 三叠纪之交生物和环境对比提供了众多依据。需要指出的是, 目前上扬子北部区深水相剖面研究程度较低, 特别是系统地层沉积研究实例较少, 极大地限制了区域生物危机事件对比。近期, 笔者在重庆巫溪红池坝发现出露连续完整的上二叠统顶部— 下三叠统底部地层剖面, 产系列牙形刺和丰富的深水相遗迹化石。文中试图聚焦红池坝剖面二叠系长兴阶上部地层, 在生物地层厘定基础之上, 研究上扬子北部区二叠纪末沉积环境演化过程, 以丰富二叠系— 三叠系之交沉积环境背景及底栖生物群多样性演化过程。
晚二叠世晚期, 峨眉地幔柱的火山喷发作用, 造成上扬子区内部产生了广泛的裂陷作用。幕式地壳沉降导致的断块差异沉降控制了区域堑垒构造格局的形成(张廷山等, 2011; 姚军辉等, 2011; 杨巍等, 2014; 王学军等, 2015), 其中深水地堑地区主要受北西向基底断裂控制, 接受大隆组沉积, 广泛分布于川北广元— 旺苍、开江— 梁平(开江— 梁平海槽)以及川东北城口— 鄂西(城口— 鄂西海)等地区(王一刚等, 2006), 与周缘浅水台地长兴组相变明显(王一刚等, 2006; 杜金虎等, 2010; 刘树根等, 2016)。
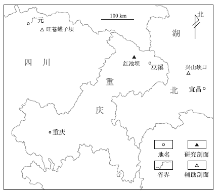
重庆巫溪地区长兴期位于川东北碳酸盐岩台地向城口— 鄂西海的转换带。区内上二叠统广泛出露, 地层分区属扬子地层区大巴山分区。大隆组厚度数米至数十米不等(牛志军等, 2001), 以红池坝剖面最为典型(图 1, 剖面起点N31° 29.319', E109° 07.150')。
 | 图 1 重庆巫溪红池坝剖面地理位置Fig.1 Location of Hongchiba section in Wuxi County, Chongqing |
红池坝剖面地层出露自上二叠统长兴组顶部至下三叠统大冶组下部, 岩性及厚度描述如下(图2):
上二叠统— 下三叠统大冶组> 10im
第1段(未见顶)
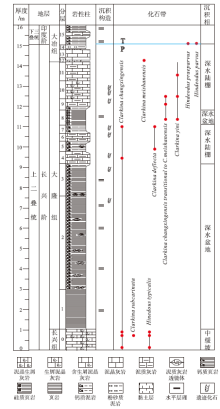
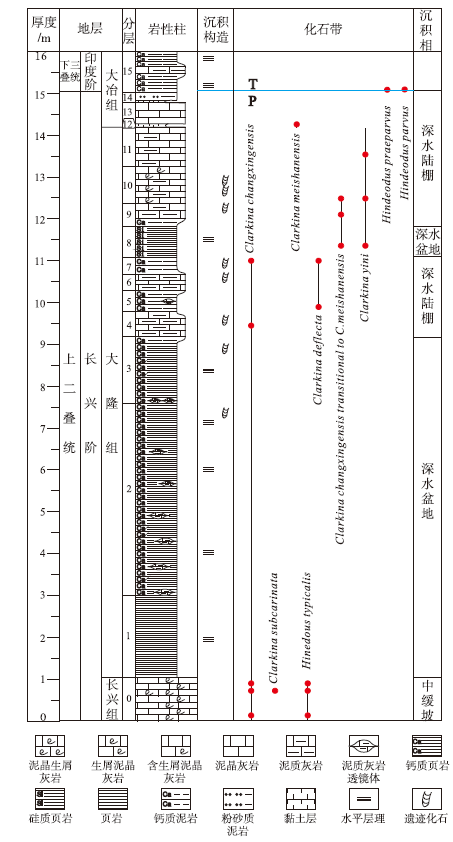 | 图 2 重庆巫溪红池坝剖面长兴阶地层综合柱状图Fig.2 Comprehensive column of the Changhsingian from Hongchiba section in Wuxi County, Chongqing |
15. 深灰色薄层钙质泥岩夹泥质灰岩, 风化后呈土黄色, 发育水平层理。底部产牙形刺Hindeodus parvus(图 3-A)及Hindeodus praeparvus(图 3-B) > 10m
14. 深灰色薄层粉砂质泥岩, 底部为厚0.06im的灰色薄层黏土岩 0.26m
13. 深灰色薄层泥晶灰岩, 岩性较单一, 层面草莓状黄铁矿密集分布 0.50m
12. 黄灰色— 灰色薄层黏土岩, 产牙形刺Clarkina meishanensis(图 3-F, 3-G) 0.08m
— — — 整合— — —
上二叠统大隆组 13.18m
11. 深灰色薄层泥晶灰岩及含泥质泥晶灰岩, 草莓状黄铁矿发育。产牙形刺Clarkina yini(图 3-H) 0.95m
10. 深灰色薄层含生屑泥晶灰岩及泥质灰岩, 生屑以钙质海绵骨针为主, 含少量放射虫。产遗迹化石Palaeophycus striatus、Palaeophycus wutingensis、Taenidium satanassi及Thalassinoides suevicus, 产牙形刺Clarkina yini(图 3-I)及Clarkina changxingensis transitional to C.meishanensis(图 3-K) 0.87m
9. 下部为深灰色薄层钙质泥岩, 上部为深灰色薄层泥质灰岩, 见薄介壳与放射虫。产遗迹化石Planolites montanus、Planolites beverleyensis及Cochlichnus anguineus, 产牙形刺Clarkina changxingensis transitional to C.meishanensis(图 3-L) 0.55m
8. 灰黑色— 黑色硅质页岩。产牙形刺Clarkina changxingensis transitional to C.meishanensis(图 3-M)及Clarkina yini(图 3-J) 0.74m
7. 深灰色薄层钙质泥岩, 由放射虫定向排列构成细粒纹层, 层间见黄铁矿。产遗迹化石Treptichnus bifurcus及Planolites montanus, 产牙形刺Clarkina deflecta( 图 3-N)及Clarkina changxingensis(图 3-P) 0.40m
6. 黄灰色— 灰色薄层泥质灰岩夹灰色薄层泥晶灰岩, 见少量生屑, 磨蚀严重。产遗迹化石Rusophycus hanyangensis0.40m
5. 深灰色薄层钙质泥岩, 黄铁矿普遍。产牙形刺Clarkina deflecta(图 3-O), 发育小型泥质灰岩透镜体 0.50m
4. 深灰色薄层泥质灰岩, 见泥质水平纹层, 其中产少量海绵骨针及薄介壳, 普遍被底栖生物强烈的扰动作用改造而呈稀散分布。该层上部层面产大量遗迹化石Chondrites type-B, 产牙形刺Clarkina changxingensis(图 3-Q) 0.60m
3. 下部为灰黑色钙质页岩, 含大量放射虫及海绵骨针, 其底部深灰色泥质灰岩透镜体发育; 上部为灰黑色钙质页岩, 以放射虫为主; 顶部为深灰色— 灰黑色薄层钙质泥岩, 含海绵骨针及少量有孔虫, 层面及层间见遗迹化石Palaeophycus heberti及Taenidium irregularis1.60m
2. 灰黑色— 黑色碳质页岩, 含海绵骨针及放射虫, 具定向性。层内发育深灰色— 灰黑色泥质灰岩透镜体, 自下向上透镜体规模渐小。顶部产遗迹化石Palaeophycus tubularis, Protopaleodictyon cf. submontanum及Planolites punctatus4.58m
1. 灰黑色— 黑色碳质页岩, 含海绵骨针及放射虫 1.95m
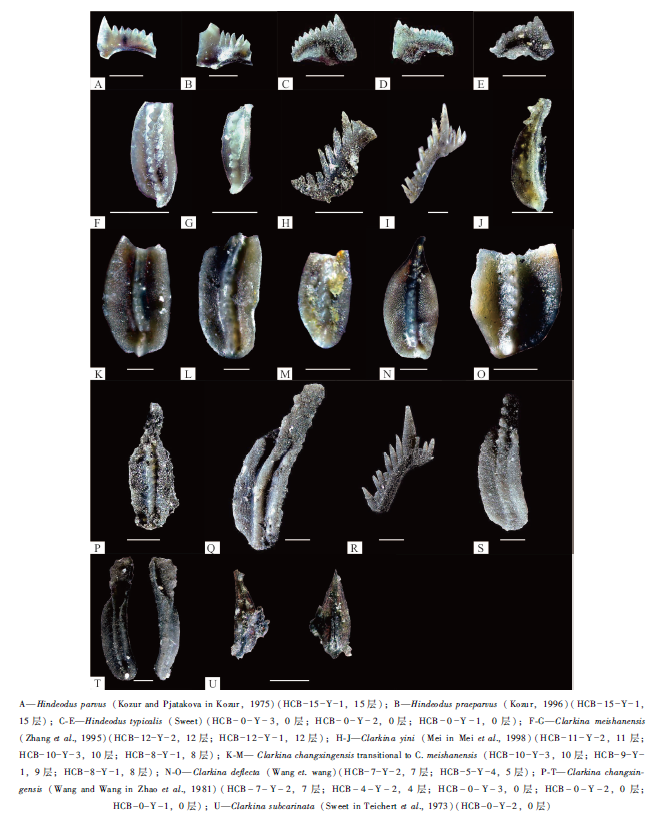 | 图 3 重庆巫溪红池坝剖面上二叠统— 下三叠统牙形刺化石(线段比例尺均代表 200iμ m)Fig.3 Conodonts from the Upper Permian-Lower Triassic in Hongchiba section of Wuxi County, Chongqing (Scale bar=200iμ m, all the individuals’ pictures were taken from upper view) |
— — — 整合— — —
上二叠统长兴组厚度> 5m
0. 灰色中— 厚层生屑泥晶灰岩, 局部生屑富集为泥晶生屑灰岩。生屑以棘皮、有孔虫、绿藻、腕足类、双壳类为主, 磨蚀普遍。层间见斑状白云化。产牙形刺Clarkina changxingensis(图 3-R, 3-S, 3-T), Hindeodus typicalis(图 3-C, 3-D, 3-E)及Clarkina subcarinata( 图 3-U)
多数岩石地层单元划分均具有穿时性, 而生物地层划分为建立等时地层格架对比提供了重要依据。虽然二叠纪末生物灭绝事件导致全球超过90%的海洋生物种相继灭绝(Erwin, 2006), 但仍有部分晚二叠世牙形刺属种延续至早三叠世, 因此牙形刺在上二叠统— 下三叠统生物地层划分对比中扮演重要角色。室内处理及分析表明, 红池坝剖面牙形刺保存较为完整, 大多数属于Clarkina属和Hindeodus属的Pa型分子。根据Clarkina属和Hindeodus属内种的地层分布及演化关系, 将剖面划分为4个牙形刺带, 自下而上分别为:
1)Clarkina changxingensis带
该带以Clarkina changxingensis(Wang and Wang in Zhao et al., 1981)分子为主体, 以Clarkina yini首现为顶界, 因剖面起点(长兴组顶部)可能处于此牙形类化石带内, 所以该带底界不清。该带包括剖面0~7层(图 2), 共生分子包括Hindeodus typicalis, Clarkina subcarinata, Clarkina deflecta。
2)Clarkina yini带
该带以Clarkina yini Mei, 1998分子为主体, 底界为Clarkina yini首现, 顶界至Clarkina meishanensis首现。包括剖面8~11层(图 2), 相当于煤山D剖面24层, 其中有共生分子趋向于由Clarkina changxingensis向Clarkina meishanensis过渡。
3)Clarkina meishanensis带
该带分布于剖面12层(图 2), 以Clarkina meishanensis(Zhang et al., 1995)分子为主体, 以较高丰度为特征, 未见其他带分子。另外, 该黏土层中检出火山成因石英, 可与煤山剖面生物主灭绝界线对比。
4)Hindeodus parvus带
该带底界为Hindeodus parvus首现, 分布于剖面15层底部; 未见顶。共生分子包括Hindeodus praeparvus。由于Hindeodus parvus为三叠系底部带化石, 因此剖面15层底部相当于煤山剖面27层C底界( 图 2), 是重要的二叠系— 三叠系分界标志(殷鸿福等, 2001)。
1)长兴组顶部生屑泥晶灰岩
位于Clarkina changxingensis带内, 为长兴组顶部中— 厚层沉积。薄片中常见丰富的棘皮、有孔虫、绿藻、腕足类、双壳类生物碎屑, 其中棘皮类碎片磨蚀严重, 而腕足类、双壳类生物碎片则相对平直, 这些经搬运改造的生屑密集填集于灰泥基质中, 生物扰动明显(图 4-A), 反映其形成于中缓坡环境, 后者空间上分布较广。
2)大隆组底部碳质/钙质泥页岩
该时期由底栖生物控制的碳酸盐生产过程基本停止, 以暗色富有机质页岩和海绵骨针与放射虫组合为标志和特征(1~3层)(图 4-B), 覆于以生物成因组分为主的碳酸盐沉积之上, 前者指示深水盆地环境, 与后者在垂向上表现出明显相变。碳质/钙质泥岩中发育规模不等的泥质灰岩透镜体(图 4-C), 垂向上表现为微弱正粒序(鲍马序列D-E段), 表明沉积时可能位于盆地边缘, 间歇受到远源浊流作用影响。
3)大隆组下部泥质灰岩/钙质泥岩
主要岩性为薄层泥质灰岩及钙质泥岩, 间夹薄层泥晶灰岩, 草莓状黄铁矿普遍分布于富有机质灰泥中(4~7层)(图 4-D)。除少量海绵骨针、介形虫及有孔虫等实体化石外, 遗迹化石丰富, 如Chondrites type-B常在层面构成分枝且相互具有交切和叠伏等特征(图 4-E), 潜穴充填物周围颗粒堆积并呈现出明显的涡状排列(图 4-F), 指示潮下深水陆棚环境。
 | 图 4 重庆巫溪红池坝剖面Clarkina changxingensis带沉积特征Fig.4 Sedimentary characteristics of Clarkina changxingensis zone in Hongchiba section of Wuxi County, Chongqing |
4)大隆组中部硅质页岩
位于Clarkina yini带内, 有机质丰富(图 5-A), 化石以放射虫及少量薄壳介形虫为主(8层), 缺乏生物扰动, 为深水缺氧盆地环境。
 | 图 5 重庆巫溪红池坝剖面Clarkina yini带内沉积特征Fig.5 Sedimentary characteristics of Clarkina yini zone in Hongchiba section of Wuxi County, Chongqing |
5)大隆组上部泥质灰岩
垂向上方解石/黏土矿物总体升高, 构成多套向上变浅的沉积序列(9层)。单个旋回中, 下部缺乏生物扰动层, 放射虫及少量海绵骨针大多定向排列(图 5-B), 形成毫米级纹层(图 5-C); 上部则生物扰动强烈, 包括Planolites montanus, Planolites beverleyensis及Cochlichnus anguineus。潜穴类型多样, 指示水循环良好的深水陆棚环境。
6)大隆组顶部含生屑泥晶灰岩
剖面10层下部生屑含量较高, 介于10%~20%之间, 常见放射虫和海绵骨针, 以后者最为丰富, 垂向上发育微弱的正粒序及冲刷面(图 5-D)。正粒序上段生物造迹现象明显, 包括潜穴管相对较粗的Palaeophycus striatus、Palaeophycus wutingensis、Taenidium satanassi及Thalassinoides suevicus在层面广泛分布。10层上部生物造迹渐少, 潜穴直径逐渐变细直至最终消失。
7)大冶组底部黏土岩及泥晶灰岩
黏土岩位于大冶组底部, 分别发育于12层(Clarkina meishanensis带)与14层下部, 间夹薄层富莓状黄铁矿泥晶灰岩, 黏土岩单层厚6~8icm; 泥晶灰岩层中发育水平纹层, 含有少量石英颗粒, 未见生物化石或生物扰动构造, 反映了生物灭绝期深水陆棚环境的萧条面貌。
红池坝剖面大隆组产出极其丰富的深水相遗迹化石, 对讨论长兴中— 晚期深水盆地— 陆棚环境中的生物分布参数具有至关重要的作用。
3.2.1 Clarkina changxingensis带 红池坝剖面Clarkina changxingensis带内依次发育Palaeophycus tubularis、Palaeophycus hebert、Chondrites type-B、Treptichnus bifurcus等典型遗迹化石分子。其中Palaeophycus tubularis及Palaeophycus hebert生物扰动量占10%~20%, 生物造迹周期性出现, 总体与沉积界面斜交, 向上开口为主以保持水道和空气流通为特征(图 6-A, 6-B), 是浊流事件后底栖生物短期觅食的结果, 属深水盆地贫氧型机遇分子。
 | 图 6 重庆巫溪红池坝剖面Clarkina changxingensis带主要遗迹化石类型Fig.6 Main types of trace fossils of Clarkina changxingensis zone in Hongchiba section of Wuxi County, Chongqing |
Chondrites type-B构成丰度极高的单一微小潜穴系统, 层面扰动量大于50%, 以水平或近水平分布为主, 在暗色寄主层面呈椭圆状、枝状、羽状分枝, 交切复杂, 级别2~3级(图 6-C)。指示该类造迹生物殖居期间, 深水陆棚环境长期处于底层水贫氧且孔隙水缺氧条件(马会珍等, 2010)。
Clarkina changxingensis带顶部见Treptichnus bifurcus等遗迹分子分布, 具扰动深度浅、扰动指数较低等特征(图 6-D), 反映沉积底质仍处于贫氧界面附近。
3.2.2 Clarkina yini带 红池坝剖面Clarkina yini带内分别发育Planolites montanus(图 7-A)及Palaeophycus striatus等遗迹化石分子。其中Palaeophycus striatus表现为垂直或与层面高角度斜交潜穴(图 7-B), 伴生有多样具不规则斜纹Palaeophycus wutingensis(图 7-C)及复杂三维潜穴系统Thalassinoides suevicus(图 7-D), 但上述遗迹化石扰动强度均相对较低, 垂向不连续的潜穴变化说明造迹生物殖居窗较小。Clarkina yini带顶部毫米级泥质纹层灰岩发育, 见大量黄铁矿, 逐渐替代原有潜穴发育层, 指示原富氧深水陆棚含氧水平逐渐降低, 并最终转向缺氧。
 | 图 7 重庆巫溪红池坝剖面Clarkina yini带主要遗迹化石类型Fig.7 Main types of trace fossils of Clarkina yini zone in Hongchiba section of Wuxi County, Chongqing |
晚二叠世, 区内在浅水台地上形成了具伸展性质的开江— 梁平海槽、城口— 鄂西海等(马永生等, 2006; 王一刚等, 2006; 毛黎光等, 2011; 肖安成等, 2011; 王学军等, 2015)(图8), 但关于该套深水沉积的时代尚存在较多争议。牛志军等(2001)认为城口— 鄂西海在晚二叠世吴家坪期率先发育大隆组, 并表现出自盆地中心向边缘渐次升高的特征; 肖安成等(2011)进一步提出, 城口— 鄂西海深水盆地发育时期早于开江— 梁平海槽, 两者具有由东向西扩展的次序规律(毛黎光等, 2011); 赵宗举等(2012)则指出, 上述深水盆地大隆组与长兴组为典型的“ 同期异相” 关系(杜金虎等, 2010), 对应于长兴阶。
 | 图 8 扬子地块北部地区晚二叠世岩相古地理图(据肖安成等, 2011, 有修改)Fig.8 Lithofacies palaeogeography map of the Late Permian in northern Yangtze Block (modified from Xiao et al., 2011) |
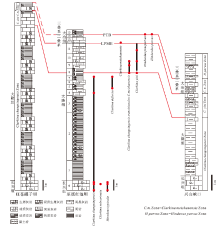
依据红池坝剖面牙形刺化石地层分布, 与广元旺苍罐子坝剖面(房强等, 2012)、宜昌兴山峡口剖面(Shen et al., 2014)生物地层对比表明, 区域大隆组层位普遍高于长兴组, 大隆组底界位于Clarkina changxingensis带内(图 9), 大致与长兴中期相当(吴亚生, 邮件交流)。大隆组底部深水沉积与下伏长兴组生物碎屑灰岩沉积相序变化明显, 表明区域在长兴中期曾经历显著的岩相分异, 反映了同时期系列深水盆地快速沉降过程。
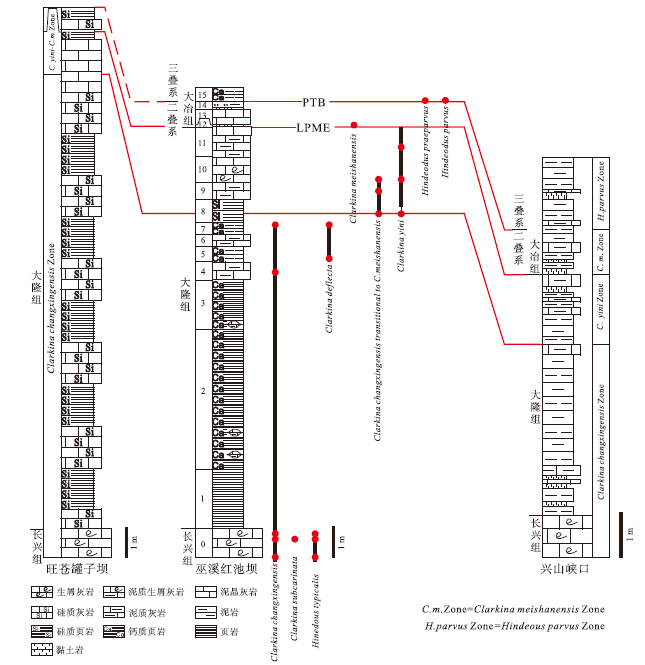 | 图 9 扬子地台旺苍罐子坝— 巫溪红池坝— 兴山峡口牙形刺带对比Fig.9 Correlation of conodont zones from different sections(Guanziba section of Wangcang County, Hongchiba section of Wuxi County and Xiakou section of Xingshan County)in Yangtze Plate |
红池坝剖面深水陆棚相在垂向主要发育于Clarkina changxingensis带上部与Clarkina yini带上部。前者Chondrites type-B等遗迹分子显示底栖造迹生物在形态和生理上均发生了特殊的变异, 结合有机质富集背景表明, 该时段深水陆棚环境长期处于特殊贫氧环境。
相比较之下, Clarkina yini带上部沉积记录反映了二叠纪末海洋化学状态的频繁波动过程。早期多样性的遗迹化石类型、复杂三维潜穴系统及低有机质含量均反映了深水陆棚经历了水体含氧量迅速升高的过程, 自由氧含量的增高指示原有贫氧带的萎缩, 后者与二叠纪末海平面下降事件相互印证(Yin et al., 2007)。随后细粒纹层沉积层与潜穴沉积层交替出现则暗示水体氧气含量的富集可能缺乏足够的稳定。Clarkina yini带顶部, 正常的含氧底质水不断被长时间尺度的缺氧事件打断, 生态环境频繁趋于恶化。
值得注意的是, 红池坝剖面Clarkina yini带上部由缺氧事件造成的底栖造迹生物丰度震荡变化并非孤立事件。煤山剖面化石记录显示, 珊瑚、及菊石等生物丰度及多样性在Clarkina yini带顶部均出现不同程度的失稳及衰退现象(Yin et al., 2007), 地球化学及生物标志化合物等也有相应的证据(Grice et al., 2005; Algeo et al., 2007), Huang等(2007)根据煤山剖面绿硫细菌的AIR和Pr/Ph观察到第24层内部存在波动明显的氧化还原条件变化过程。二叠纪末深水陆棚可能处于水体缺氧不断扩张的过程(Algeo et al., 2007), 而红池坝剖面深水沉积记录则从底栖生物的应对策略角度映射了生物危机事件的发展过程。
1)重庆巫溪红池坝剖面上二叠统顶部— 下三叠统底部地层发育完整。通过牙形刺化石研究, 建立了5个牙形刺化石带, 分别为Clarkina changxingensis带, Clarkina yini带, Clarkina meishanensis带, Clarkina yini带, Hindeodus parvus带。
2)红池坝剖面大隆组沉积相类型包括深水盆地相与深水陆棚相, 前者为具远源浊流的碳质/钙质泥页岩、富含有机质硅质页岩等沉积组合; 后者主要由产多样遗迹化石的泥质灰岩/钙质泥岩、含生屑泥晶灰岩组成。
3)大隆组/长兴组界线位于Clarkina changxingensis带内。该时期长兴组缓坡沉积与大隆组深水盆地垂向沉积相序发生突变。剖面对比显示, 区域大隆组/长兴组界线显著岩相分异, 是系列伸展背景下地堑区快速沉降的结果。
4)红池坝剖面在Clarkina changxingensis带上部与Clarkina yini带上部均处于深水陆棚相背景。遗迹化石分布及特征揭示, 前者底质水体长期处于稳定的贫氧— 缺氧环境; 后者水体氧气含量变化明显, 特别是后期随着富黄铁矿纹层逐渐代替Palaeophycus、Thalassinoides suevicus等多样潜穴分布层, 水体氧化还原条件呈频繁波动变化趋势。红池坝剖面的沉积记录见证了二叠纪末生物与环境的协同演化过程。
致谢 中国科学院地质与地球物理研究所吴亚生副研究员为牙形刺鉴定提供了有益的帮助和建议, 杜翔及张静同学在野外露头及室内资料整理中给予了很大帮助, 在此表示衷心的感谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|