



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
叠层石中的海绿石化和黄铁矿化:以天津蓟县中元古界铁岭组为例*
[梅朝佳 ]
]
]
|
|

作者简介 梅朝佳,女,1994年生,中国地质大学(北京)博士研究生,主要从事微生物碳酸盐岩和层序地层等方面的研究。E-mail: mei_chaojia@163.com。
作为微生物碳酸盐岩的主要类型之一,叠层石是微生物席的主要建造物已成为共识。天津蓟县中元古界铁岭组二段叠层石生物礁灰岩发育,其中的细粒叠层石被前人解释为微生物席捕获碳酸盐泥的微生物建造物,使得其既不同于现代叠层石,也不同于显生宙尤其是寒武纪的叠层石。更为特殊的是,这些叠层石中的海绿石和黄铁矿代表着 2种特殊的矿化作用,其中研究区普遍产出的黄铁矿,作为硫酸盐还原细菌的产物,是了解古代微生物的窗口;而发育在高能浅海的海绿石,产出环境不同于现代海绿石,不能作为慢速沉积环境的指示矿物,亦不具有沉积间断的地质意义。 2种矿化作用表明铁岭组叠层石是由沉淀作用而非捕获碳酸盐泥形成,这为了解中元古代叠层石的形成和特征提供了一些有益的线索。
As the main type of microbial carbonates,it has become the consensus that stromatolites are mainly constructed by microbial mats. The stromatolitic bioherm of the Second Member of the Tieling Formation in Jixian section,Tianjin,are made of fine-grained stromatolites, interpreted as a microbial structure formed by trapping carbonate muds on microbial mats; they are different from the modern stromatolites and most of the Phanerozoic stromatolites. Particularly, the glauconite and pyrite preserved in the stromatolites represent two types of special mineralization. As a result of sulfate-reducing bacteria,pyrite commonly produced in the study area,is a window to understand the ancient microorganisms. Glauconite that is developed in the high-energy shallow sea is different from the modern glauconite in the production environment; it may not be used as the indicator mineral of the slow depositional rate,nor has the geological significance of sedimentary discontinuity. The two types of mineralization directly suggest that the Tieling stromatolites are formed by precipitation,rather than the capture of carbonate muds. This study provides some useful clues for understanding the characteristics and formation of the Mesoproterozoic stromatolites.
微生物席是形成大多数叠层石和其他大规模底栖微生物碳酸盐岩加积表面的复杂结构(Riding, 2000)。对现代叠层石多样化构成的研究也表明, 叠层石是以蓝细菌为主的微生物席的建造物(梅冥相和孟庆芬, 2016)。基于Kalkowsky(1908)构建并定义的“ 叠层石” 术语, 叠层石被广泛认为是早期石化的纹层状生物沉积构造(Awramik and Sprinkle, 1999; Riding, 2000, 2011a; 梅冥相和孟庆芬, 2016; Tosti and Riding, 2017)。在微生物碳酸盐岩尤其是叠层石的形成过程中, 颗粒的捕获在局部是重要的, 关键的过程还是沉淀, 因为这个过程不仅产生了钙化微生物的生物礁聚集, 而且提高了微生物席的堆积和保存(Riding, 2000; Dupraz et al., 2009)。Riding(2011b)强调, 细粒叠层石主要是同沉积微生物沉淀作用的产物, 并极有可能是由细胞外聚合物(EPS)或其他细胞产物的异养细菌钙化作用所产生, 如异化的硫酸盐还原作用。然而, Tosti 和 Riding(2017)将天津蓟县铁岭组二段生物礁灰岩中的细粒叠层石解释为微生物席捕获碳酸盐泥形成的微生物建造物, 但该捕获理论似乎不能用于解释天津蓟县铁岭组叠层石中亮暗纹层的形成。
发育在铁岭组二段叠层石生物礁灰岩中的海绿石(Glauconite)(K, Na, Ca)(Fe, Al, Mg, Mn)2(Si, Al)4O10(OH)2, 是一种富钾、富铁的含水层状铝硅酸盐矿物, 常呈颗粒状或碎屑产出, 易被搬运形成异地海绿石(Ordin and Fullagar, 1988); 反之, 若其呈薄膜状或结壳层等其他的形式发育在不整合面或硬底面上, 就应该属于原地海绿石(梅冥相等, 2008)。与现代大量发育在水深200~300im、温度为15~20i℃的弱碱性水体以及弱氧化至弱还原环境中(赵全基等, 1992; 陈丽蓉, 1994; Kitamura, 1998; Chafetz and Reid, 2000; 梅冥相等, 2008; 汤冬杰等, 2016)的海绿石不同, 研究区的海绿石发育在前寒武纪的高能浅海环境中(孟晓庆, 2006; 梅冥相等, 2008; 周锡强等, 2009; 汤冬杰等, 2016)。
海绿石常被作为海洋环境中低沉积速率的标志, 其中显生宙中钾含量高的海绿石更被认为是有代表意义的凝缩段(Baum and Vail, 1988; Amorosi, 1995; 葛铭等, 1995; Harris and Whiting, 2000; 陈淑慧等, 2014; Banerjee et al., 2016)。值得注意的是, 前寒武纪中的海绿石与显生宙海绿石在沉积环境上存在明显不同, 前者90%以上集中分布在具较高沉积速率的潮间带至浅潮下带环境中, 而后者70%以上集中分布在具较低沉积速率的中陆棚至上斜坡较深水环境中(Banerjee et al., 2016)。因此, 集中形成于浅水环境, 是前寒武纪海绿石不同于显生宙海绿石的一个特殊现象(Chafetz and Reid, 2000; 梅冥相等, 2008; 周锡强等, 2009; Banerjee et al., 2015, 2016)。
黄铁矿作为沉积岩中常见的自生矿物, 在不同的环境中形成模式也不尽相同(常华进和储雪蕾, 2011), 但主要都来源于硫酸盐还原细菌(SRB)。沉积硫化物占地球表面硫化物的95%以上, 但由于Fe的大量富集, 致使沉积物中的硫化物几乎全以黄铁矿的形式产出(Rickard et al., 2017), 并可作为了解微生物历史窗口的矿物(Schieber, 2002)。天津蓟县铁岭组二段叠层石柱体及柱间泥晶中广泛分布的黄铁矿, 无疑是造成微生物席及其构成的EPS降解的异养细菌SRB的残余物。
文中通过对天津蓟县中元古界铁岭组二段特殊海绿石和黄铁矿的产出环境进行讨论, 探索其成因及地质意义, 同时揭示铁岭组二段细粒叠层石的形成机制, 以了解前寒武纪的海水性质以及大气环境。
天津蓟县剖面铁岭组主要由海相碳酸盐岩组成。基于前人对研究区铁岭组地层学及沉积学的研究成果(宋天锐等, 1991; 高林志等, 1996; 梅冥相等, 2000; 霍勇等, 2012; 曲永强等, 2012),
并根据铁岭组中部和顶部发育的不整合面分别对应于区域构造运动“ 铁岭上升” 和“ 芹峪上升” , 将铁岭组划分为2个三级层序。这2个层序大体上与铁岭组的2个亚组对应, 即代庄子亚组(铁岭组一段)和老虎顶亚组(铁岭组二段)。各层序界面清晰, 其中铁岭组二段顶面为区域平行不整合面, 底面铁质风化壳可作为暴露层序面的典型, 而一段底面为岩性转换界面(霍勇等, 2012)。
铁岭组一段(DS1)以含锰白云岩为主, 中厚层含锰白云岩与薄层白云质泥页岩组成环潮坪型碳酸盐米级旋回(Schwarzacher, 1993; 梅冥相等, 2000; 李怀坤等, 2014)。梅冥相等(2000)将此三级层序划为上潮间坪含锰白云岩构成的海侵体系域(TST)、下潮间坪厚层块状含锰叠层石质白云岩构成的早期高位体系域(EHST)以及向上变浅至暴露相的晚期高水位体系域(LHST), 此为典型的退积副层序堆叠。铁岭组二段(DS2)以白云岩或白云质灰岩为主, 中部发育巨厚叠层石礁灰岩夹盆屑质块状灰岩。其海侵体系域(TST)由潮间坪灰质白云岩到潮下坪叠层石灰岩这样一种向上变深的退积序列所构成; 高位体系域(HST)表现为潮下坪相叠层石质灰岩转变为潮上坪相泥晶白云岩, 是一个向上变浅至暴露的进积序列, 与构成TST单元的序列正好相反。文中所研究的薄膜状海绿石以及黄铁矿主要产于该段的EHST中。
虽然早期根据海绿石K-Ar年龄推测铁岭组形成时限不超过1200iMa(王松山, 1995; 李明荣和王松山, 1996), 但后期测试得出下马岭组凝灰岩锆石的U-Pb同位素年龄约为1380iMa(高林志等, 2007; 苏文博等, 2010; 李怀坤等, 2014), 铁岭组二段底部的斑脱岩测年结果约为1440iMa(苏文博等, 2010; 李怀坤等, 2014), 故推测蓟县剖面及整个华北北缘的下马岭组与铁岭组分界处时限大约为1400iMa。
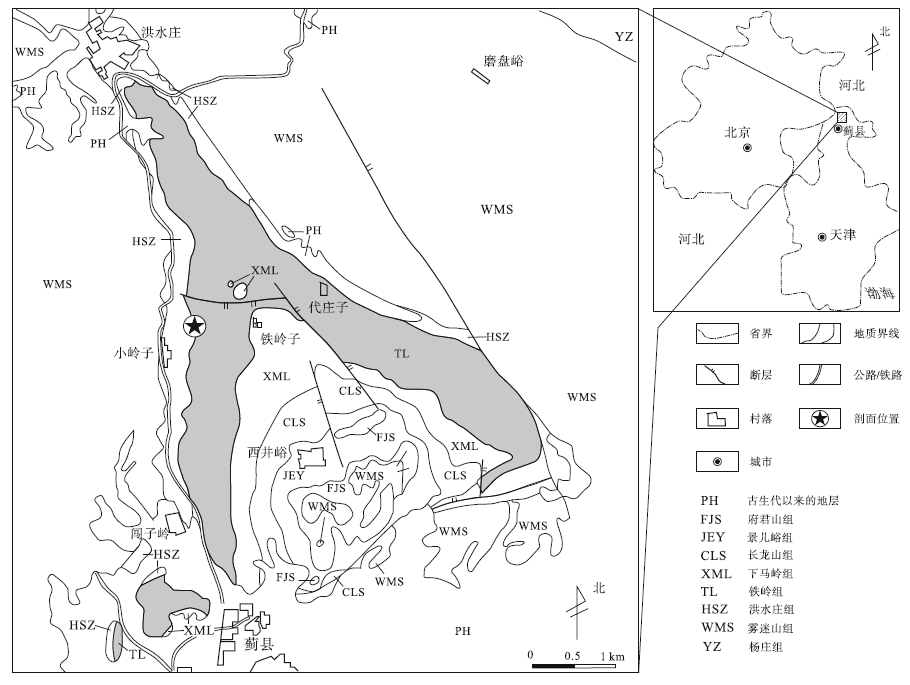 | 图 1 天津蓟县小岭子村— 洪水庄地区地质概图①Fig.1 Geological map of Xiaolingzi Village-Hongshuizhuang area, Jixian County, Tianjin |
叠层石作为地球早期生命历史的代表物, 横跨了从古代到现代3.5× 109年的地质年代记录, 产出形态及环境更是多种多样。高振西等(1934)首次在华北地台铁岭组中识别出大量的叠层石, 并将之描述为Collenia chihsienensis; 在此基础上, 曹瑞骥和梁玉左(1974)标识出Anabaria和Baicalia, 同时创建新的属Pseudotielingella 和 Tielingella。基于前人大量的研究成果, 可将天津蓟县铁岭组叠层石认为是由丰富多样的柱状叠层石组合形成的“ 巨厚层大型生物丘” , 具体类型有: Baicalia cf. baicalica, Chihsienella chihsienensis, Conicodomenia longotenuia, Conophyton luotuolingens, Tielingella tielingensis, Paraconophyton inconspicum, Scopulimorpha regularis, Pseudochihsienella inconspicua。 由于该叠层石相互渐变、连续交替组合, 形成了不同的叠层石组合, 因而被列为中国5个各具特色的中— 晚元古代叠层石组合之一(Chen et al., 1981; 曹瑞骥和袁训来, 2003; 梅冥相等, 2008)。
铁岭组叠层石在垂向上以底部为水平状叠层石, 向上变化为柱状、锥状或墙状叠层石为特征, 主要为Anabaria chihsienensis, Baicalia cf. baicalica等截面为圆形或卵形的柱状叠层石。叠层石生物层的单层厚度多为数米, 少数能达到20im左右, 表明该叠层石的产出环境为潮下高能动荡环境(梅冥相等, 2008)。另外, 图 2-d中显示的铁岭组柱状叠层石在平面上呈拉长状, 大小为1.5icm× 2.6icm~5.6icm× 20icm, 暗示可能存在一个之前被忽略但是应该普遍关注的现象, 即元古代柱状叠层石具有在平面上延伸的特点(Tosti and Riding, 2017)。
与现代多数产于慢速、较深水、弱还原沉积环境中的海绿石(黄杏珍, 1982; Odin, 1988; Chafetz and Reid, 2000; Banerjee et al., 2016)不同的是, 文中所研究的古老叠层石中发育的海绿石, 产于天津蓟县剖面中元古界铁岭组二段高能叠层石生物层灰岩中, 其以胶体薄膜状的形式发育在叠层石柱体间的碳酸盐岩泥晶的边界上, 体现了原地海绿石的特点(梅冥相等, 2008; 周锡强等, 2009; 汤冬杰等, 2016)。同时, 因为其产于高能浅海叠层石柱体的边界, 故也明显不同于现代海洋环境中产出的海绿石(梅冥相等, 2008)。
 | 图 2 天津蓟县中元古界铁岭组二段叠层石灰岩宏观特征Fig.2 Macroscopic characteristics of stromatolitic limestone in the Second Member of Mesoproterozoic Tieling Formation at Jixian section, Tianjin |
与产于较深水环境、呈颗粒状产出的海绿石不同, 研究区海绿石产出于天津蓟县铁岭组二段叠层石岩礁灰岩中, 从宏观上看其主要呈薄膜状产出于柱状叠层石(以Tielingella tielingensis、Chihsienella chihsienensis和Baicalia cf. baicalica为主)外壁上(图 3)。而在其他类型的叠层石中, 如丘状、锥状及墙状叠层石中, 未发现发育海绿石。
从图3-a可以看出, 柱状叠层石部分轻微分叉, 叠层石柱体及充填的碳酸盐泥晶基本保持宽度不变, 海绿石主要以不足1imm的墨绿色薄膜产于柱状叠层石外壁上。
 | 图 3 天津蓟县中元古界铁岭组二段叠层石中海绿石的宏观特征Fig.3 Macroscopic characteristics of glauconite in stromatolite in the Second Member of Mesoproterozoic Tieling Formation at Jixian section, Tianjin |
与图 3-a不同的是, 图 3-b为底部发育大型丘状叠层石、向上以1条缝合线为界(图 3-b黄色虚线所示)突变为柱状叠层石的组合体。在底部的大型丘状叠层石中没有海绿石的发育, 相反在上部柱状叠层石中, 其外壁大量发育墨绿色海绿石薄膜, 同时在碳酸盐泥晶的局部发育经过风化作用和后期改造的淡绿色海绿石。
在平面上, 叠层石都呈现出不规则的圆, 如在图 3-c中可以清楚地识别出叠层石柱体、碳酸盐泥晶, 以及产出于叠层石柱体和碳酸盐泥晶边界上呈1.5imm厚的薄膜状海绿石, 其呈环形发育在柱体四周。除以薄膜状发育在叠层石柱体外壁以外, 海绿石还以宽度约1.5icm的带状发育在整个碳酸盐泥晶中(如图 3-d中黄色虚圈所示), 为早期成岩作用胶结的产物。
天津蓟县铁岭组叠层石在宏观上存在多样的产出形态, 但其本质都是由具有一定起伏的基本层堆叠起来的, 是具有清晰界限的细粒纹层集合体。叠层石主要由较暗的隐晶质碳酸盐泥晶构成, 柱间则为泥晶灰岩, 并因两者颜色上的差异形成交替发育的亮暗纹层, 但本质都为方解石晶体。在手标本的显微照片中, 能观察到叠层石呈细粒沉积, 由不连续的亚毫米级(通常为50~200iμ m)、不透明且不规则的小透镜体和隐晶质碳酸盐泥晶组成, 并穿插相似的以微亮晶为主的薄层(图 4)。叠层石的暗色条纹具规律性变化, 从颜色较浅的不连续似透镜体或凝块定向排列, 到中部颜色较深、连续性好的暗色条纹与相间的浅色亮晶形成的交替纹层, 再到大量以暗色泥晶为主夹少量亮晶的似条纹状结构(图4-a), 显示出造叠生物对叠层石发育的定向控制。同时可观察到, 叠层石是由某个生长点或面开始生长的(图 4-b中“ A” 为生长点), 在空间上呈球面向外生长, 并由于造叠生物、环境以及沉积物的供应等因素影响使得其形状发生改变。发育在叠层石柱体与充填叠层石柱体间的碳酸盐泥晶交界处的海绿石宏观上呈薄膜状产出, 在碳酸盐泥晶中局部也发育弥散状或密集状薄膜。因此, 对天津蓟县铁岭组二段叠层石灰岩中海绿石的显微组构的研究表明, 其分布状况与宏观表现一致。
 | 图 4 天津蓟县中元古界铁岭组二段叠层石及海绿石的微观特征Fig.4 Microscopic characteristics of stromatolite and glauconite in the Second Member of Mesoproterozoic Tieling Formation at Jixian section, Tianjin |
可将天津蓟县铁岭组二段叠层石灰岩中的海绿石依其发育位置的不同划分为两类。
类型一为发育在叠层石柱体和柱间充填物交界处的海绿石(图 4-c, 4-d)。其中墨绿色薄膜状的海绿石以富含较多残留有机质的黑色叠层石柱体边(图 4-c中“ b” 所示)为界, 主要发育在柱间泥晶灰岩中; 局部发育颗粒状海绿石(图 4-c中“ g2” 表示), 粒径范围较大, 为0.05~0.15imm; 下部的胶体状海绿石中同时发育黄铁矿聚集体(图 4-c中“ p” 表示)。与以叠层石为界发育在泥晶灰岩中的海绿石不同, 图 4-d中的海绿石以胶体形式呈弥散状或密集状发育在叠层石与泥晶灰岩交界处, 其带宽可达到1.5imm, 同时在泥晶灰岩中偶见长条状海绿石聚集体(图 4-d中箭头所示)。在柱状叠层石纹层中均未发育海绿石。
类型二中的海绿石主要以叠层石外壁为界, 呈不规则胶体形式弥散状或聚集状发育在泥晶灰岩中, 大量发育在方解石晶体附近, 其形态受到亮晶方解石晶形的影响和改造。弥散状海绿石的分布似乎没有规律, 连接成不规则片状的海绿石常发育在富含大量残余有机质的边界附近(图 4-e)。
图 4-f充分证实了大量海绿石发育在叠层石柱体边缘。颗粒状海绿石大量发育在亮晶方解石附近, 粒径40~150iμ m, 变化较大, 形状受限于方解石晶形。海绿石以复合薄膜构成不规则片状覆盖于亮晶方解石之上, 部分颗粒还隐约保留有海绿石颗粒边(图 4-f中箭头所示), 显示出溶蚀的特点。
与其他区域产出的叠层石相比, 除发育在叠层石柱体与柱间碳酸盐泥晶边界以及碳酸盐泥晶中的薄膜状或颗粒状海绿石外, 使得天津蓟县铁岭组二段叠层石礁灰岩成为特殊地质现象的原因还包括叠层石中普遍发育的不规则胶体状和规则颗粒状黄铁矿。
在柱状叠层石中发育大量黑色斑状体, 其粒径变化较大, 为0.04~0.2imm, 普遍发育于叠层石亮暗纹层中(如图 5-a箭头所示), 多数尺寸较大的黄铁矿较集中发育在暗色纹层附近。同样, 在叠层石柱体及柱间泥晶灰岩边界, 在发育海绿石的同时, 也发育大量的黑色黄铁矿不规则集合体(图 5-b)。这些黄铁矿集合体在海绿石周围或之中发育, 粒径大小较一致, 约0.15imm。此外, 在叠层石和泥晶灰岩中也有发现黄铁矿, 相比较之下, 其粒径较小, 呈星散状分布, 且没有发现明显的顺层发育, 多数为单个黄铁矿晶体, 显示出原生的特点。图 4-f为图 5-b的局部(黄色虚线所示区域“ B” )放大, 通过对图 4-f的进一步观察发现, 在泥晶灰岩中发现粒径不超过0.04imm黄铁矿晶体, 表明黄铁矿在泥晶灰岩中较普遍。
 | 图 5 天津蓟县中元古界铁岭组二段叠层石中黄铁矿的微观特征Fig.5 Microscopic characteristics of pyrite in stromatolite in the Second Member of Mesoproterozoic Tieling Formation at Jixian section, Tianjin |
叠层石中的原生黄铁矿, 大多数不是以自形晶形态产出(图 5-c, 5-d), 而是以胶体状的形式复合发育在叠层石中, 集合体粒径最大可达1imm。这种不规则的黄铁矿晶体的大量产出, 表明其与硫酸盐还原反应有关, 为SRB降解腐烂的微生物席及其构成的EPS后形成的残余物。
天津蓟县铁岭组二段中的海绿石, 宏观上以薄膜状(图 3-a, 3-b, 3-c)附于叠层石柱体外壁上, 部分充填在整个柱间泥晶中(图 3-d), 微观上呈胶状体, 局部呈颗粒状产于叠层石柱体与柱间泥晶交界以及柱间泥晶灰岩中(图 4), 显示出原地海绿石以及“ 早期溶蚀作用和晚期成熟作用” 的特点。具体表现为, 早期充填在碳酸盐泥晶的微孔隙中, 随着碳酸盐泥晶的溶蚀作用以及更加强烈的海绿石化作用, 逐渐聚合成不规则形态的片状体(图 4-f)。由于沉积物的供给速率大于叠层石生长速率, 造成叠层石柱体的生长截止(如图 3-b)(Tosti and Riding, 2017), 同时柱间泥晶不断充填, 形成有利于海绿石生长的微环境, 从而形成了海绿石带。
Hower(1961)提出的层形点阵(Layer lattice theory)模式, 显然不适用于天津蓟县铁岭组高能叠层石灰岩中的海绿石的形成, 相比较之下, 研究区海绿石显示出的“ 早期溶蚀和晚期成熟作用” 的特点, 似乎更符合Odin和Matter(1981)提出的颗粒变绿理论(Verdissement of grains)。海绿石的不同颜色对应于不同的成熟度, 研究区墨绿色的海绿石(图 3, 图 4)显示出高成熟特点, 梅冥相等(2008)和汤冬杰等(2016)对研究区海绿石的组分研究也证明了其为高成熟海绿石。研究认为, 高成熟海绿石晶粒的形成时间估计为105~106年, 常被认为是沉积明显间断的标志(Odin and Matter, 1981; Odin and Fullagar, 1988; Amorosi et al., 2012)。相比较之下, 发育于铁岭组高能叠层石礁灰岩中的高成熟海绿石明显不同, 不具有能指示沉积间断的地质意义。因此, 古老岩石记录中的海绿石矿物, 不能作为中陆架和深水环境或者低沉积速率环境的指示矿物, 这个观点在对美国西南部奥陶纪产在高能浅海地层中的海绿石的研究中也得到了证实(Chafetz and Reid, 2000)。同时, 海绿石的发育也表明, 天津蓟县中元古代的浅海环境可能还处于氧气不足的阶段。
通过组分分析, 汤冬杰等(2016)指出天津蓟县铁岭组二段叠层石礁体缺乏陆源碎屑, 因此, 研究区内呈星散状普遍发育在柱状叠层石纹层和海绿石中的黄铁矿为原生黄铁矿。作为SRB降解腐烂的微生物席沉淀形成的残余物, 原生黄铁矿的产出表明天津蓟县铁岭组二段灰岩中柱状叠层石主要是由沉淀作用形成。在地质演化中, 尽管微生物岩的有机格架仍然存在, 但是由于席状体复杂的细节常随有机物死亡而快速消失(Dé farge et al., 1996; Riding, 2000), 使得虽然钙化蓝细菌在现存大多数微生物碳酸盐岩中保存得很好(Riding, 2000), 但在现代和化石海洋环境中却很少发现(Grotzinger and Knoll, 1999), 这表明可能有包括SRB在内的其他新陈代谢组分使得这些环境中的微生物席发生石化(Baumgartner et al., 2006)。
梅冥相等(2008)描述了天津蓟县铁岭组二段叠层石岩礁灰岩中的黄铁矿, 指出“ 草莓状” 黄铁矿的产出可能代表了SRB在形成叠层石和柱间泥晶灰岩活动时的痕迹。包括叠层石在内的大量微生物碳酸盐岩中黄铁矿的产出, 亦表明了微生物碳酸盐岩的形成与硫酸盐还原反应存在密切联系(Schieber, 2002; Baumgartner et al., 2006; 曹瑞骥和袁训来, 2006; 梅冥相等, 2008, 2011, 2015; Bartley et al., 2015)。Baumgartner等(2006)将这种关系理解为对微生物席的石化和保存过程的重要作用。
通过前人的研究发现, 在叠层石的形成中沉淀作用占据主要地位, 而颗粒的捕获只在局部是重要的(Riding, 2000; Dupraz et al., 2009)。与寒武纪以来发育的粗糙纹层叠层石以及现代产出的粗粒叠层石不同, 天津蓟县铁岭组发育具良好纹层的细粒叠层石。Riding(2011b)强调, 细粒叠层石主要是同沉积微生物沉淀作用的产物, 微生物席下部缺氧层中的硫酸盐还原细菌等对有机质的降解促进了碳酸钙的沉淀, 异养细菌集中降解腐烂的有机质(Bartley, 1996)及细胞外基质内有机质腐烂驱动的泥晶化作用, 产生并保存了较细粒的泥晶纹层, 并主要形成于强烈的硫酸盐还原反应的区域内(Bosak et al., 2013)。研究区中普遍发育的黄铁矿作为SRB降解腐烂的微生物席沉淀形成的残余物, 证明由沉淀作用主导产生了柱状叠层石, 同时早期成岩作用胶结形成的海绿石也间接证实了该观点。
以微晶结构为主导的许多微生物碳酸盐岩, 其明显的均一性掩盖了一系列可能的起源, 同时也增加了对微生物碳酸盐岩形成研究的困难程度。天津蓟县中元古界叠层石礁灰岩中2种特殊的矿化现象, 赋予该柱状叠层石不同于现代及显生宙尤其是寒武纪以来的叠层石的特殊意义。铁岭组二段叠层石边界及柱间泥晶中发育的海绿石, 由于产在高能浅海环境中而不能指示低速率沉积环境, 因而不具有指示沉积间断的地质意义, 也表明当时的大气圈氧气还不充分。同时, 普遍发育的黄铁矿, 作为SRB降解腐烂有机质沉淀形成的残余物, 表明该叠层石主要是由沉淀作用而不是捕获泥晶形成; 海绿石的发育说明叠层石生长期间内在柱体间形成了一种特殊的还原环境, 也间接地论证了叠层石的形成是沉淀作用的结果。不可否认叠层石的形成与SRB之间存在密切的联系, 但主导叠层石形成的具体微生物席及形成机制还有待进一步研究。
致谢 在成文过程中, 梅冥相教授提出了富有建设性的意见和建议, 使得文章增色不少; 在野外资料采集时得到了同门师兄赵新伟的帮助, 以及师兄王龙、肖恩照及师姐张瑞的帮助, 笔者在此一并表示由衷的感谢!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|