











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
微生物岩的概念和重庆老龙洞剖面P-T界线地层微生物岩成因*
[吴亚生1, 2, 3  , 姜红霞
, 姜红霞4 , 虞功亮5 , 刘丽静6 ]
, 姜红霞|
|

第一作者简介 吴亚生,男,1963年生,1984年毕业于南京大学,1987年在中国科学院地质与地球物理研究所获硕士学位,2002年获中国科学院南京地质古生物研究所博士学位,现为中国科学院地质与地球物理研究所副研究员和中国科学院大学岗位教授,主要从事碳酸盐岩古生物和沉积相、生物礁、微生物沉积研究。E-mail: wys@mail.igcas.ac.cn。
微生物岩的原始定义指底栖微生物主导形成的沉积体或岩石体。笔者对该概念进行了扩充,认为微生物岩除了包括叠层石、凝块石、纹层石、核形石、均一石之外,还应该包括微生物骨架岩、微生物粘结岩、非钙化浮游或漂浮微生物形成的模铸岩、矿化浮游或漂浮微生物形成的颗粒岩和泥粒岩。 P-T界线地层微生物岩的特征是具有由较粗矿物晶体(主要是方解石,其次是白云石)组成的斑点状、树枝状、网状结构的灰岩。这 3种结构分别称为斑点状体、树枝状体、网状体,在露头上呈暗色,在薄片中呈浅色,一般由无定形的亮晶充填体和其间的微亮晶组成。亮晶充填体是指无定形的孔洞被不同成岩期形成的矿物充填形成的结构体,因成岩作用各异,造成不同层位、不同地点的亮晶充填体内部的矿物类型和充填顺序存在差异;所有的亮晶充填体都不具有壁,故不是钙化化石。通过形态、大小和生态比较,以及形成演化分析,认为亮晶充填体的前身是漂浮蓝细菌微囊菌,胶鞘是微囊菌形成模铸化石的关键因素。亮晶充填体是表层水漂浮生活的微囊菌沉入海底后,被泥晶沉积物掩埋或者被早期海底胶结物胶结,在泥晶沉积物半固结或固结之后腐烂留下的孔洞被后期成岩作用形成的矿物充填形成的。 P-T界线地层微生物岩段顶部遭受成岩作用程度高,树枝状体和网状体中的亮晶充填体的轮廓基本都被破坏,变成微亮晶和亮晶,以前被学者解释为凝块石;但斑点状体、树枝状体、网状体是成岩流体沿着亮晶充填体或其他化石丰富的地方运移形成较粗的晶体而造成的,并不符合凝块石的定义。同时,少数学者把树枝状体本身当成底栖生物,也是没有充分认识树枝状体的矿物组成而做的解释。该微生物岩段含有钙化的小球状化石和同心层状化石,但它们不是组成微生物岩的主体。
About the first author Wu Ya-Sheng,born in 1963,graduated and obtained his Bs,Ms,and Ph.D. degrees from Nanjing University,Institute of Geology,Chinese Academy of Sciences(CAS),and Institute of Geology and Paleontology,CAS in 1984,1987 and 2002,respectively. Now he is an associate professor in Institute of Geology and Geophysics,CAS and adjunct professors of University of CAS,with main research interests in paleoecology and sedimentology of microbialites,reefs,and other carbonates. E-mail: wys@mail.igcas.ac.cn.
Microbialites are carbonate bodies formed by microbial actions of benthic microbial communities or mainly composed of skeletons of planktonic microbes,and include the old types such as stromatolites,thrombolites,microbial laminated rocks,oncolites,leiolite,and the new types: Microbial framestones,microbial bindstone,and microbial caststone mainly composed of cast fossils of microbes,microbial grainstone,and microbial packstone. The Permian-Triassic boundary microbialites in South China include three types: Those with patchy coarse-mineral structures,those with dendroid coarse-mineral structures,and those with net-like coarse-mineral structures. The three types of structures are composed of spar-filled structures and the microsparry filling the space between the spar-filled structures. The spar-filled structure is defined as caves left by decay of some non-calcified organisms and filled by various minerals formed in diagenesis,and varies in inner fillings and fabric due to difference in diagenetic history. The rocks between the coarse-mineral structures are micrites with few fossils and scattered spar-filled structures. The shapes of the spar-filled structures in the PTB microbialites are very similar to those of the colonies of Microcystis,an extant cyanobacterial genus,in shape and size and are the cast fossils of the latter. The PTB microbialites are not thrombolites. As colonies of Microcystis sank to the floor and were buried by micrites,their sheaths did not decay until the micrites were slightly lithified. After their decay,the caves they left were firstly cemented by needle-like aragonite,and then altered by dolomitization forming dolomites,which mostly were altered to calcite and were dissolved by fresh water in surface environment. The caves finally were filled by blocky calcite. Diagenesis has destroyed the outlines of most spar-filled structures,making it difficult to recognize. Diagenetic fluids migrated along the sediments with abundant spar-filled structures,destroying them,forming coarse minerals,which resulted in the patchy,dendroid and netlike appearance of the PTB microbialites. Since the formation of the patchy,dendroid and netlike bodies is controlled by diagenesis rather than microbes,and are not composed of micrites but coarse minerals,they are not thrombolites. There is no calcified wall in the spar-filled structures,and they are not calcimicrobes. Small coccoid fossils occur in the upper part of the microbialite interval,but not the lower part,in small content,and is not responsible for formation of PTB microbialites.
微生物岩既是一种重要的碳酸盐沉积, 又是重要的油气储集层。微生物指蓝细菌、细菌等需要借助显微镜才能观察的微小生物, 其主要以矿化、诱导矿化、捕获粘结等方式形成矿物沉积, 以碳酸盐沉积为主。在中奥陶世以前, 特别是在漫长的元古代, 微生物形成的碳酸盐沉积具有相当大的规模和数量, 不仅记录了古海洋和古大气圈的重要信息, 而且是重要的油气储集层和烃源岩, 所以微生物岩研究既具有重要的科学意义, 又对国民经济发展具有重要的应用前景。然而, 由于对微生物的认知有限, 特别是以往的研究主要集中在常见的有机和无机成因碳酸盐沉积方面, 导致微生物岩的研究开展较少, 总体研究程度非常薄弱。除了叠层石研究之外, 凝块石和其他类型微生物岩的研究虽然近年来开展较多, 但是研究的程度往往不够深入。造成这种状况的主要原因在于一些相关的概念在现有文献中尚不是很明晰, 对大部分学者来说不好把握。例如, 凝块石的定义从文字而言并不复杂, 但是实际应用时却困难很大, 故文中首先讨论一下凝块石和微生物岩的有关概念。
众所周知, 二叠纪末发生了显生宙中最大的一次生物集群灭绝事件(e.g., Shen and Bowring, 2014), 其后在华南浅海地区广泛发育了一套树枝状的微生物岩。对这套微生物岩的研究始于1999年(Kershaw et al., 1999), 针对其形成机制和包含的古环境信息, 最主要的认识是树枝状岩石为凝块石(e.g., Ezaki et al., 2003)。但姜红霞和吴亚生(2007)研究后发现, 树枝状特征主要是成岩作用形成的, 其中的不规则囊状体(即本文的亮晶充填体)是某种菌落形成的模铸化石; Wu 等(2014a)通过进一步的研究提出, 树枝状微生物岩是当时的蓝细菌赤潮形成的沉积受成岩作用改造形成的模铸化石。虽然模铸化石这一认识目前已被多数学者接受(Luo et al., 2014; Deng et al., 2015; Shen et al., 2015; Yang et al., 2015; Liao et al., 2016; Zhang et al., 2016), 但仍有少数学者坚持认为其为凝块石(Wu et al., 2017), 甚至提出其是钙质微生物成因(Kershaw et al., 2018)。限于期刊的篇幅限制, Wu 等(2014a)的文章中未能就微生物岩剖面的岩石学特征进行细致描述, 因此为了进一步澄清认识, 促进对这套微生物岩的认识, 文中对树枝状微生物岩研究最经典的重庆北碚老龙洞2个剖面进行详细的岩石学描述, 在此基础上对这套微生物岩的形成机制做进一步的解释。
微生物岩研究的开展, 比微生物岩定义的出现要早很多, 如叠层石这一术语, 早在1908年就被提出来了(Kalkowsky, 1908)。虽然叠层石是最经典的微生物岩, 但微生物岩不是只包括叠层石。Aitken(1967)定义了微生物岩的另一个重要类型— — 凝块石。凝块石的初始定义很简单, 是指与叠层石外形和大小相似但内部不具有纹层构造的岩石, 其具有凝块状或海绵状结构(a macroscopic clotted or spongy fabric)。然而, 这是一个很难被应用的定义, 因为岩石中的不均匀结构很多, 究竟什么样的结构是凝块状或海绵状并不好把握; 同时, 不仅沉积作用可以形成不均匀结构, 成岩作用也可以形成, 如果两者不加区分, 将使这一术语失去意义。
此后, 很多学者认为凝块石是叠层石受到生物扰动破坏形成的。Pratt和James(1982)在使用凝块石术语时出现了重大歧义, 他们将薄片上见到的1~3 cm宽的暗色斑块叫做凝块石, 而这实际上是把Aitken(1967)定义的凝块石内部的凝块当成了凝块石。这一用法仅少数学者认同(边立曾等, 1996)。
Kennard和James(1986)指出了Pratt和James(1982)使用凝块石术语的错误, 否定凝块石是由叠层石内部构造被生物扰动破坏形成的观点, 主张凝块石是一种独立的微生物沉积类型, 以具有凝块结构为特征, 其中凝块是毫米级到厘米级大小的、暗色的、泥晶或者微晶组成的、形状和分布不规则的、被泥晶或砂级沉积物或亮晶分隔开的构造, 内部结构包括叶状的、细胞状的、微球状的、凝块状的、微粒状的、球粒状的、蠕虫状的、斑点状的、块状的等, 或者由肾形藻、葛万藻、努亚藻等钙化微生物组成, 是不连续分布的小球状微生物群落(主要是蓝细菌)原地钙化形成的。在这一定义中, 凝块被认为是以泥晶为主的原地沉积, 形成岩石的格架, 凝块之间是沉积物或亮晶胶结物, 这为凝块石的识别提供了基本依据; 但不足之处在于其把原地钙化微生物骨骼当成凝块, 同时对凝块成因的解释不够全面。笔者认为, 以原地钙化微生物骨骼为主形成的岩石应属于生物礁, 可以归入到微生物岩的范畴, 但不应该归入到凝块的范畴, 同时小球状微生物的钙化不是全部凝块的成因, 甚至不是主要成因。从Kennard和James(1986)文章中的照片看, 凝块是暗色的部分, 凝块之间为灰色的泥晶沉积物和白色的亮晶沉积物, 由于泥晶和亮晶是充填在暗色凝块之间的, 所以形成时间比暗色凝块晚; 限于照片分辨率不够, 笔者看不清凝块究竟是化石还是泥晶灰岩溶蚀的残余。
通过大量的薄片观察, 笔者发现潮坪鸟眼灰岩和窗格灰岩的鸟眼孔和窗格孔往往遭受过溶蚀作用, 且在溶蚀作用中有泥晶形成, 并沉积在鸟眼孔和窗格孔的底部; 而鸟眼孔和窗格孔的上部, 常被后期的亮晶方解石胶结物充填, 形成示顶底构造。实际上, 以前学者定义凝块石时, 使用的一些照片就是鸟眼灰岩的; 但这些学者的解释不准确, 他们误把凝块当成骨架。实际上, 鸟眼孔和窗格孔是沉积期微生物占据的地方, 在沉积时微生物才是真正的骨架, 它们之间是被吸附或障积的泥晶。所以, 从本质上说, 凝块石的定义正好把骨架与沉积物搞反了。若按照以前学者的定义, 凝块是小球状微生物群落钙化形成的, 那么它就应该是化石; 而如果它是化石, 那么这种岩石就应该是钙化微生物形成的生物礁。所以, 仅就定义而言, 认为其是钙化微生物形成的这种认识是不妥的。
Riding(2000)对叠层石和凝块石的定义与前人相同, 并将凝块石进一步分为钙化微生物凝块石、粗糙粘结凝块石、树形凝块石(arborescent thrombolite)、泉华凝块石和沉积后凝块石共5种。 钙化微生物凝块石是由树枝状钙化蓝细菌形成骨架, 这种骨架在薄片中状似凝块, 但不是典型的钙质微生物化石(Riding, 2000, Fig.10), 而是特殊的钙化微生物(Tarthinian); 粗糙粘结凝块石指凝块石中包含较粗的沉积物颗粒, Riding未提供显微照片, 因而特征不好把握; 泉华凝块石即具有凝块结构的泉华; 树形凝块石指有十几厘米大小的树形结构的凝块石, Riding未提供显微照片; 沉积后凝块石指准同生期或者成岩期形成的凝块石。树形石(dendrolite)指由厘米级的、灌木状钙化微生物(表附藻、肾形藻等)形成的微生物岩, 是新定义的一个微生物岩类型, 但名称与树状凝块石容易混淆, 笔者认为其实际上是钙质微生物形成的骨架岩。之后的研究中, Riding(2011)把表附菌、肾形菌、葛万菌形成的岩石既归入到树形石中, 又归入到钙质微生物凝块石(Calcified microbe thrombolites)中, 造成了矛盾。
另外, Shapiro(2000)指出, Aitken(1967)对凝块(clot)的原始定义就不清楚, 其就是等同于凝块石的, 但实际上凝块石是宏观构造, 凝块是构成凝块石的中观构造。而Ezaki等(2017)仍然把表附菌形成的岩石称为树形凝块石(dendritic thrombolites)。
尽管叠层石、核形石、凝块石都是微生物岩, 但在1987年之前还没有微生物岩这一名词。Burne和Moore(1987)定义了微生物岩这一术语, 指底栖微生物群落主导形成的碳酸盐沉积, 并且把微生物岩分为3类: 微生物绑结岩、微生物泉华和微生物骨架岩。微生物绑结岩, 是丝状蓝细菌、小球状蓝细菌、硅藻以及少数真核微生物借助体表黏液吸附、固着沉积物而形成的, 典型代表是叠层石。微生物泉华, 是微生物在无机成因泉华的形成过程中被包裹进去形成的, 所以把这种泉华归入微生物岩的范畴; 且由于微生物群落的存在, 也造成了泉华具有一些特有结构。微生物骨架岩, 指丝状蓝细菌等微生物矿化作用形成的岩石, 它不具有明显可鉴定的微生物化石形态, 因此应该另外命名, 因为传统意义的骨架岩是由可鉴定的原地的钙化微生物构成的; 而在后来学者的文章中, 属于Burne和Moore(1987)的微生物骨架岩范畴的岩石, 一般都被归入凝块石中。在这3种分类中, 尽管第3种的名称不妥, 但第1种的名称还是比较合理的; 另外, 将含有微生物引起的构造的泉华归入微生物岩, 也被后来的学者认可。Burne和Moore(1987)还用大量的实例, 说明把叠层石和凝块石简单地认为分别是丝状蓝细菌和小球状蓝细菌形成的认识是不正确的, 他们把微生物岩分为5种: (1)叠层石, 具有纹层构造; (2)凝块石, 具有凝块结构; (3)核形石, 由同心状结构的纹层组成; (4)隐结构微生物岩, 具模糊的斑点或斑状结构; (5)球粒状微生物岩, 是球粒状集合体。在前人研究的基础上, Riding(2000)将微生物岩分为叠层石、凝块石、树枝石和均一石共4种。
中国学者也对微生物岩分类进行初步探索。如梅冥相(2007)提出将核形石和纹理石作为微生物岩的新类型。实际上, Burne和Moore(1987)的微生物岩分类已经包含了核形石, 而纹理石如果是微生物成因的, 就应当属于面状的叠层石。鉴于叠层石一般都是柱状的形态, 所以将面状叠层石给予一个专有名称“ 纹理石” , 是很有意义的, 不过需要同无机成因的纹层构造区别开来。韩作振等(2009)提出, 表附菌形成的微生物岩不宜归入现有的微生物岩分类系统中, 而应归入生物礁岩分类体系中, 称为骨架岩。王月等(2011)也讨论了微生物岩的分类问题。罗平等(2013)把纹理石改叫层纹石, 亦主张将表附菌灰岩称作骨架岩。
Wu等(2007)描述了一类主要由小球状微生物化石构成的岩石, 这种小球状化石在西藏姜叶玛的P-T界线地层也有分布(Shen et al., 2010), 推测是漂浮生活的。根据定义, 微生物岩只包括底栖微生物及其作用形成的岩石, 但漂浮生活的微生物形成的岩石也是微生物成因的岩石类型, 把它们排斥在微生物岩之外并不合理。所以, 笔者提出对微生物岩分类进行扩充(表1)。
| 表1 微生物岩的新分类方案 Table 1 Scheme for microbialites classfication |
微生物包括底栖微生物和浮游/漂浮微生物2大类, 就数量而言, 浮游/漂浮微生物数量更大。底栖微生物和浮游/漂浮微生物形成的沉积都为微生物沉积, 形成的岩石均属于微生物岩的范畴。微生物分为钙化(矿化)的和非钙化(矿化)的, 但矿化的底栖微生物形成的沉积物既属于微生物岩的范畴, 也属于生物礁岩的范畴。浮游/漂浮的矿化微生物形成的岩石, 如果矿化化石数量达到一定的含量, 也应当归入微生物岩范畴。笔者建议, 矿化化石在岩石中的含量达到泥粒岩和颗粒岩的范畴或者含量大于50%, 就可以当作微生物岩。例如, 放射虫含量达到泥粒岩的范畴, 叫放射虫泥粒岩。如果浮游/漂浮微生物不是矿化的, 但形成模化石或铸体化石, 且模化石或铸体化石含量达到岩石体积的50%以上, 也属于微生物岩范畴, 可以在化石名称之后加上模铸岩命名, 如微囊菌模铸岩。对于非钙化底栖微生物形成的沉积物, 如果具有纹层构造, 那么面状的叫层纹石, 柱状的和丘状的叫叠层石, 同心状的叫核形石; 如果不具有纹层构造, 那么内部结构不均一的属于凝块石的范畴, 内部结构均一的属于均一石的范畴。
P-T界线地层指晚二叠世末期到早三叠世的地层, 既包括了大灭绝之前的一段地层, 也包括了大灭绝之后的一段地层。大灭绝之前的地层, 以含有大灭绝前的生物群为特征, 最典型的生物是䗴和柯兰尼虫。大灭绝之后的地层, 在浅水相区包括一套微生物岩和其上的一段地层。微生物岩是一套在露头上就可以看到树枝状、网状或者斑点状特征的岩石。在薄片中可以看到, 树枝状、网状和斑点状特征是由特殊的内部组成和结构造成的。在浅水区, 这套微生物岩之上是含小腹足类的泥晶灰岩, 如在重庆北碚老龙洞剖面。再向上出现没有生物的泥质泥晶灰岩或泥灰岩。根据岩性特征和生物面貌, 把P-T界线地层限定为这段地层。P-T界线地层之上的下三叠统中还出现多套微生物岩, 但那已经不属于界线地层的范畴。
P-T界线是根据浙江长兴煤山剖面的牙形刺化石带定义的, 以主齿极其粗壮的牙形刺Hindeodus parvus的首现作为三叠纪的开始。二叠纪生物大灭绝的主幕发生在煤山剖面的24e层之顶(Yin et al., 2000; Shen et al., 2014), 即牙形刺Clarkina yini带(Yin et al., 2001; Wu, 2005)之顶。Hindeodus parvus首现于煤山剖面的27c层。Clarkina yini带与Hindeodus parvus带之间自下而上还存在Clarkina meishanensis带和Hindeodus changxingensis带2个牙形刺带(Wu, 2005)。煤山剖面的28层属于牙形刺Isarcicella staeschei带。就牙形刺化石带而言, P-T界线地层一般应自下而上包括这5个牙形刺化石带: C.yini, C.meishanensis, H.changxingensis, H.parvus, I.staeschei。
一些学者认为, H.parvus在浅水相区出现在微生物岩的底部(刘建波等, 2007; 王海峰等, 2012)。但刘建波等(2007)一文图中的H.parvus, 有一些标本不具有H.parvus正模的典型特征。在江西修水P-T界线地层剖面上, 典型的H.parvus首现在微生物岩的中部(Wu and Jiang, 2014), 微生物岩的下伏地层是含有典型H.changxingensis的生屑颗粒灰岩, 微生物岩之上的泥晶灰岩和颗粒灰岩中含有H.parvus, 所以, P-T界线地层的微生物岩只包括了H. changxingensis带的上部和H.parvus带的下部。
杨浩等(2006)认为, 湖北崇阳界线地层剖面中H.parvus出现在微生物岩的顶部。根据该文中的图4, 鉴定为H.parvus的照片1和照片2与标准的H.parvus是不同的, 实际上是层位偏上的类似H.parvus, 虽然它们一般也被归入到H.parvus, 但实际上是与H. parvus正模有区别的。Wu和Jiang(2009)提出将这类化石归入Hindeodus crenatusWu(2009)。H.crenatus的模式标本见Wu和Jiang(2009), 以侧面更强的膨大和细齿排成弧线而不是直线为特征。H.parvus是张克信等(1995)首次报道的。1995年以前, P-T界线是用菊石、双壳类等化石确定的, 精度较低。目前, 在缺乏牙形刺化石的情况下, 碳同位素曲线是一个比较有效的替代手段。
二叠纪末生物大灭绝事件发生在二叠系— 三叠系界线之下。在浅水相区, 大灭绝之前的地层一般是海百合灰岩(例如, 在重庆北碚老龙洞的剖面上), 海百合灰岩之上就是微生物岩。因微生物岩中不见大化石, 所以在露头和薄片上其与海百合灰岩很容易区分。在深水相区的P-T界线地层中, 不发育微生物岩。浅水相区微生物岩的发育不限于P-T界线地层, 在P-T界线地层之上的下三叠统中也有微生物岩。限于篇幅, 文中不讨论P-T界线地层之上的微生物岩。
P-T界线地层微生物岩的第1例报道出现于1999年, 研究对象包括四川华蓥县的老龙洞、百竹园、涧水沟、文星场4个剖面(Kershaw et al., 1999), 微生物岩的特征为: 下伏地层为二叠纪生物礁之上的海百合灰岩, 上覆地层为三叠系飞仙关组纹层状泥晶灰岩和页岩; 呈指状(digitate)、局部凝块石状(thrombolitic), 有残余的叶状构造, 像微生物岩, 但大多数因重结晶而不能确定是微生物成因。在Kershaw 等(1999)一文中: 图2-a, 2-b照片被标注为指状, 这种外貌(也是内部构造)在后来的文章被广泛称作树枝状(dendroid); 图2-c是素描图, 斑状的暗色部分被作者标注为凝块石状(thrombolitic), 这种形态的构造后来被叫做斑点状的构造(王永标等, 2005)。Kershaw等(1999)把斑点状的岩石视为凝块石状, 而不认为树枝状的岩石是凝块石状。凝块石属于微生物岩, Kershaw等(1999)说斑点状的岩石是凝块石状的, 并不是说就是凝块石, 也未敢确定这套岩石是微生物岩, 只是认为像微生物岩。之后, Kershaw等对此又进行了深入研究:Kershaw等(2007)对华南P-T界线地层微生物岩再次进行描述, 认为微生物岩主要是树形石(dendrolites), 还有一些凝块石状的层, 其形成环境是缺氧的、分层的浅海陆架; Kershaw等(2012)提出华南的微生物岩是肾形藻类钙质微生物形成的凝块石, 中东和土耳其的微生物岩包括叠层石和凝块石两类; 最近, Kershaw等(2018)提出P-T界线地层的微生物岩是钙质微生物形成的。
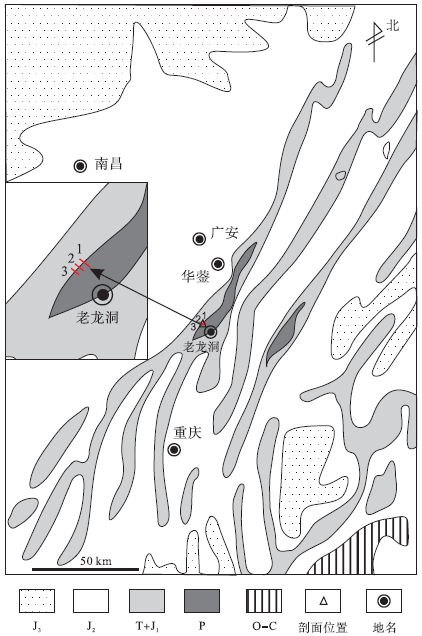 | 图1 重庆老龙洞P-T界线地层1、2、3号剖面地理位置Fig.1 Location of sections 1, 2, and 3 at Laolongdong area, Chongqing |
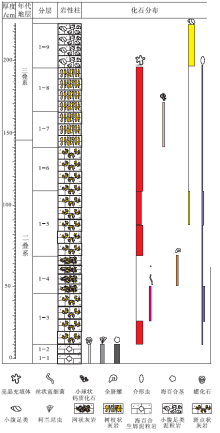
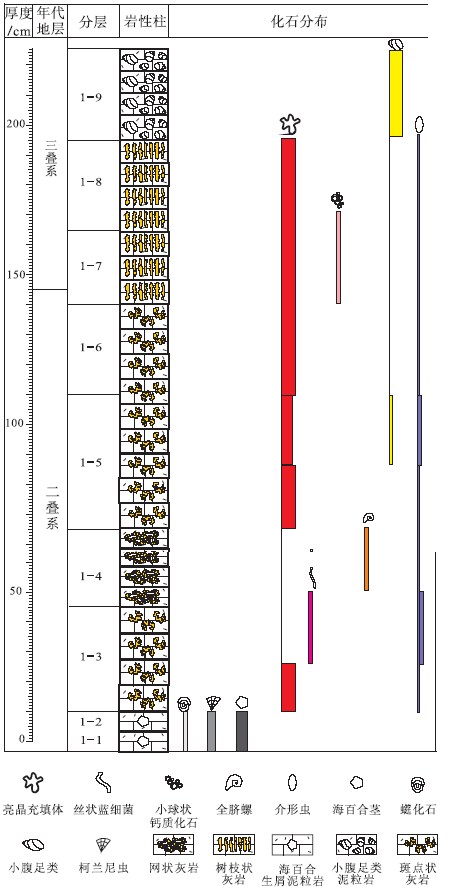 | 图2 重庆老龙洞1号剖面P-T界线地层柱状图Fig.2 Comprehensive column of the Permian-Triassic boundary microbialite sequence of section 1 at Laolongdong, Chongqing |
Lehrmann(1999)报道了贵州边阳的微生物丘(calcimicrobial mounds): 三叠系最下部是由骨架岩组成的生物层(biostromes), 具有钙质微生物构成的坚固骨架(a rigid calcimicrobial framework), 骨架由不规则到丛状的、具房室结构或凝块状泥晶结构的、可归入钙化蓝细菌肾形菌的块体组成(The framework is made of irregular-to-tufted masses of chambered-to-clotted micrite structures referable to Renalcis, a calcified coccoid cyanobacteria); 骨架岩生物层之下是含有䗴Palaeofusulina的长兴组灰岩, 代表大灭绝之前的地层; 与骨架岩生物层互层的颗粒灰岩中, 含有早三叠世的牙形刺Isarcicella isarcica。根据煤山剖面的研究, Isarcicella isarcica位于H. parvus之上的Isarcicella staeschei之上。Lehrmann(1999)未提供牙形刺的照片, 所以无法判断其牙形刺鉴定是否正确。如果鉴定正确, 那么, 这套微生物岩的顶界高于其他剖面。这有待于进一步的研究去确定。Lehrmann(1999)一文中的图3-A、3-B(本文的图15-B是他们的图3-B)的薄片照片所示化石确实与肾形菌相似, 但是并不典型, 而且照片分辨率低、不清晰, 且此后所有其他学者都没有在P-T界线地层微生物岩中发现肾形菌。所以, 要么Lehrmann(1999)的骨架岩生物层与Kershaw等(1999)、笔者以及其他作者研究的P-T界线地层微生物岩的层位不同, 要么Lehrmann(1999)的材料太特殊, 仅代表局部和特殊的情况, 而作者以偏概全, 把局部现象当成P-T界线地层微生物岩的一般情况。Ezaki等(2003)认为这套生物层中的肾形菌可能是成岩构造。Lehrmann(1999)没有把这套岩石归入微生物岩, 而是归入生物礁, 并且提出, 二叠纪的浅海生物礁生态系没有灭绝。这一观点意义重大, 可惜没有得到后来任何研究的证实。
 | 图3 重庆老龙洞1号剖面P-T界线地层微生物岩露头特征 A— 1-3层(即1号剖面第3层, 全文同), 斑点状, 棕色斑点分布于灰色斑点之间, 是由微生物模铸化石和成岩作用共同形成的, 是曾经含有较多微生物遗体的地方; 灰色的是普通的泥晶灰岩, 是沉积期含微生物遗骸少、受成岩作用影响小的地方; B— 1-4层, 网状, 棕色网状体是由微生物模铸化石和成岩作用共同形成的, 网眼中的灰色泥晶灰岩, 是微生物模铸化石少、受成岩作用影响小的地方; C— 1-5层, 斑点状, 棕色斑点状岩石分布在灰色斑点状岩石之间; D— 1-6层, 薄片中为斑点状灰岩, 但露头上不清晰; E— 1-7层上部, 树枝状, 露头为大致平行于层面的方向; F— 1-7层下部, 树枝状, 露头垂直于层面; G— 1-8层, 树枝状, 露头大致平行于层面; H— 1-9层, 树枝状, 露头大致平行于层面。全部为露头照片Fig.3 Outcrop appearance of the Permian-Triassic boundary microbialites of beds 1-3 to 1-9 of section 1 at Laolongdong, Chongqing |
Ezaki等(2003)对华蓥县老龙洞(注: 即笔者所说的重庆北碚老龙洞)P-T界线地层的树枝状岩石进行研究后, 首次将这套岩石归入凝块石。凝块石当然属于微生物岩的范畴。在Ezaki等(2003)一文中, 图4-C、4-D和图5、图6、图7的露头和光面照片均具树枝状特征, 其中在薄片中呈浅色的树枝状部分被标示为凝块石(thrombolite); 图8-A、8-B的薄片照片中, 宽1~5imm的树枝状部分被标示为亮晶凝块(sparitic mesoclots)。之后的研究中, Ezaki等(2008)仍然将树枝状微生物岩解释为凝块石。
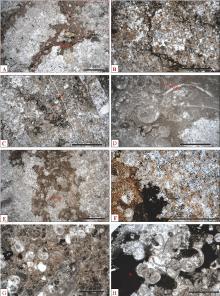
 | 图4 重庆老龙洞1号剖面第1-3~1-4层微生物岩显微特征 A— 1-3层(即1号剖面第3层)下部, 具斑点状体(红色虚线内)的泥晶灰岩, 斑点状体内部有大块状方解石(c)、泥晶灰岩残余(m); B— 1-3层, 1个斑点状体的局部放大, 可见大块状方解石(c)中的漂浮状暗色菱形方解石(r)具环带构造; C— 1-3层中部, 含斑点状体(红色虚线内)的泥晶灰岩, 斑点状体由块状亮晶方解石和其间的泥晶(m)组成, 斑点状体之间的泥晶中有很多不规则形状的亮晶充填体(指某种生物腐烂消失后留下的孔洞后来被矿物充填形成的结构体; 黄色虚线内)和少数介形虫(o); D— 1-3层中部, 1个斑点状体内的亮晶充填体内有少数漂浮状菱形方解石和少数针状方解石(nc); E— 1-3层中部, 斑点状体(红色虚线内)之间的泥晶中有较多的丝状蓝细菌(f)和介形虫化石; F— 1-3层中部, 斑点状体之间的泥晶局部放大, 丝状蓝细菌有一端弯曲现象; G— 1-4层, 具网状体(照片上半部分)的岩石, 网状体之间的泥晶(照片下半部分)中有亮晶充填体(黄色虚线内); H— 1-4层, 1个网状体的局部放大, 见1个螺旋状的小腹足类化石(g)、很多大块状方解石亮晶(c)、一些黄色的粒状方解石(gc)、少数菱形方解石晶体(r)和一些泥晶斑点(m)Fig.4 Microscopic features of the microbialites of beds 1-3 and 1-4 of section 1 at Laolongdong, Chongqing |
 | 图5 重庆老龙洞1号剖面第1-5~1-6层的微生物岩的显微特征 A— 1-5层, 具斑点状体(红色虚线内)的泥晶灰岩, 斑点状体由丰富的亮晶充填体(例如, 黄色虚线内2个)和微亮晶(例如, 紫色虚线内)组成; B— 1-5层, A图的局部放大, 1个不规则形状的亮晶充填体(黄色虚线内)边缘由黄色白云石(yd)组成, 内部由白色粒状方解石(gc)组成; C— 1-5层, 1个不规则形状的亮晶充填体(黄色虚线内), 内部边缘有少数朝内生长的针状方解石(nc)、黄色的他形粒状白云石晶体(yd)、白色块状方解石胶结物(c); D— 1-5层上部, 斑点状体(浅色区域, 被红色虚线包围的部分)和其间的泥晶(暗色区域), 泥晶中有亮晶充填体(s1所指黄色虚线内), 斑点状体由亮晶充填体和其间的微亮晶(如紫色虚线包围的部分)组成, 照片之左半部分的斑点状体内的亮晶充填体大多数可以勉强辨认(s2所指黄色虚线内), 边缘的亮晶充填体(s3所指黄色虚线内)轮廓更为明显; 右半部分可能也有很多亮晶充填体, 但轮廓无法辨认, 仅标出1个(黄色虚线内); E— D图的局部放大, 可见构成斑点状体的微亮晶(紫色虚线内)、块状方解石(c)和亮晶充填体(黄色虚线内); F— 1-6层, 主要由斑点状体组成的灰岩, 斑点状体(红色虚线包围的区域)由块状方解石(c)、暗色粒状方解石(白色虚线内)、少数泥晶(混杂在粒状方解石之间, 不易辨认)组成, 斑点状体之间为泥晶, 其中有少数轮廓分明的亮晶充填体(s1, 黄色虚线内); G— 1-6层, 左下角是1个斑点状体的局部, 中部是斑点状体之间泥晶灰岩中的几个亮晶充填体, 它们内部为块状方解石, 其中有一些暗色的菱形方解石(r), 边缘有少数残留的针状方解石(nc); H— 1-6层, 主要由 斑点状体组成的灰岩, 斑点状体之间为泥晶, 泥晶中有一些小腹足类(g)和一些亮晶充填体(s)Fig.5 Microscopic features of microbialites of beds 1-5 and 1-6 of section 1 at Laolongdong, Chongqing, China |
 | 图6 重庆老龙洞1号剖面第1-6~1-8层微生物岩的显微特征 A— 1-6层上部, 具斑点状体的灰岩, 斑点状体(红色虚线内)由大块状亮晶方解石(如白色虚线内是1个, 可见解理面)组成, 其中含有漂浮状菱形方解石晶体(r)和泥晶灰岩的残余(m); B— 1-7层, 主要由树枝状体组成的岩石, 树枝状体(4个, 红色虚线内)由丰富到密集的亮晶充填体(例如, 黄色虚线内)和其间的微亮晶(紫色虚线内)组成, 亮晶充填体一般有黄色粒状白云石组成的边缘, 但去白云石化使一些亮晶充填体不具黄色边缘, 因而难以识别(蓝色虚线内); C— 1-7层, 1个树枝状体的局部放大, 由密集的亮晶充填体(黄色虚线内的)和其间的微亮晶(紫色虚线内)组成, 一些亮晶充填体(s1)几乎全部由黄色粒状白云石组成, 另外一些(如, s2)边缘由黄色粒状白云石(yd)组成, 内部由白色粒状方解石(gc)组成; D— 1-7层上部, 树枝状体内部的一些亮晶充填体被大块状方解石充填(c, 白色虚线内是1个晶体, 正交偏光下整体消光), 这些大块状方解石中一般含有漂浮状的暗色菱形方解石晶体(r); E— D图的局部放大, 见1个亮晶充填体内部被大块状亮晶方解石充填, 边缘有少数黄色的针状方解石(nc)和黄色粒状白云石(yd), 中央有2个微亮晶充填的地方(紫色虚线内), 代表前身生物体有洞(是蓝细菌微囊菌属的群体的一个特征); F— 1-8层, 主要由树枝状体组成的灰岩, 树枝状体占到岩石体积的70%以上, 树枝状体(红色虚线内)由黄色白云石和白色方解石组成, 亮晶充填体轮廓难以辨认; G— 1-8层, 树枝状体局部放大, 边缘的亮晶充填体(黄色箭头s2所指黄色虚线内)尚可辨认, 由黄色半自形粒状白云石组成(yd), 内部由白色粒状方解石组成(gc); 内部的亮晶充填体大多被成岩作用破坏, 现由块状方解石(c)、斑状分布的黄色粒状白云石、斑状分布的白色粒状方解石(gc)、少数针状方解石(nc)和少数斑状分布的泥晶组成; 树枝状体边缘有少数轮廓保存较好的亮晶充填体(黄色箭头s1所指黄色虚线内), 其内部由黄色白云石晶体组成; H— 1-8层, 树枝状体之间的泥晶中有一些轮廓保存较好的亮晶充填体(例如, 黄色虚线内), 其边缘由黄色粒状白云石组成, 内部由白色粒状方解石组成Fig.6 Microscopic features of microbialites of beds 1-6 to 1-8 of section 1 at Laolongdong, Chongqing |
 | 图7 重庆北碚老龙洞1号剖面第1-9层非微生物岩和3号剖面第3-1~3-4层微生物岩显微特征 A— 1-9层, 具树枝状体的生屑泥粒岩, 树枝状体由小腹足类壳和介形虫壳(箭头所指处)和其内、其间的亮晶方解石(c)组成; B— 1-9层, 树枝状体局部放大, 见多个小腹足类壳(g)和少数介形虫壳(o), 所有箭头指处是壳壁, 壳内由块状方解石(c)、白色粒状方解石(gc)充填, 块状方解石包含有少数黑色菱形方解石晶体(r); C— 3-1层(即3号剖面第1层), 具斑点状体的灰岩, 斑点状体(红色虚线内)由白色微亮晶(例如, 粉色虚线内)、粒状方解石、黄色粒状白云石(yd)、少数块状方解石(c)组成, 不均匀分布的黄色粒状白云石、白色粒状方解石、微亮晶是亮晶充填体受成岩作用改造形成的; D— C图中1个亮晶充填体(黄色虚线内)的放大, 内部见少数针状胶结物(nc); E— 3-2层, 具斑点状体的灰岩, 斑点状体(红色虚线内)由亮晶充填体和其间的微亮晶(例如, 绿色虚线内)组成, 少数位于斑点状体边缘的亮晶充填体轮廓清晰(例如, 黄色虚线内4个), 但大多数亮晶充填体的轮廓被成岩作用破坏, 无法分辨; 轮廓清晰的亮晶充填体边缘为黄色粒状白云石(e)、内部为白色粒状方解石, 或内部全部被黄色的粒状白云石充填(例如, 黄色虚线内的那些亮晶充填体); F— E图的局部放大, 可见亮晶充填体(黄色虚线内)由黄色的粒状白云石(yd)组成, 亮晶充填体表面局部有暗色泥晶(dm), 可能是沉积期吸附在亮晶充填体表面的泥晶, 厚度不均匀, 在很多地方缺失, 所以不是钙化壁; 见到亮晶充填体之间的白色的针状胶结物(白色虚线内), 可能是沉积期或准同生期在亮晶充填体之间形成的针状文石胶结物, 后来转变成方解石; G— 3-3层, 具斑点状体的灰岩, 一些亮晶充填体内部可见3种胶结物: 边缘的1层黄色粒状白云石(yd)、垂直于亮晶充填体边界朝内生长的一些黄色针状方解石(nc)、内部的大块状方解石胶结物(c); H— 3-4层, 具斑点状体的灰岩, 斑点状体由大块状方解石(c)、小腹足类(g)、微亮晶 (例如, 紫色虚线内)组成Fig.7 Microscopic features of dendoid bioclastic grainstone of Bed 1-9 of section 1 and those of microbialites of beds 3-1 to 3-4 of section 3 at Laolongdong, Chongqing, China |
 | 图8 重庆老龙洞3号剖面P-T界线地层微生物岩柱状图Fig.8 Comprehensive column of P-T boundary microbialite at section 3 in Laolongdong, Chongqing |
王永标等(2005)根据对多个剖面的研究, 认为P-T界线地层的微生物岩是由微晶基质和中、粗晶方解石组成的指状体或斑点组成的, 指状体和斑点的发育程度不同:指状体即本文中的树枝状体, 长度一般为6~15 cm, 宽为0.4~2 cm, 本身也分叉, 而斑点大小一般为0.2~1.5 cm; 指状体长轴方向多与地层层面近垂直, 而斑点则较均匀地散布在微晶灰岩基质中, 没有明显的方向性。杨浩等(2006)描述了湖北崇阳的花斑状微生物岩, 实际上就是斑点状微生物岩。姜红霞和吴亚生(2007)提出P-T界线地层微生物岩中的树枝状体是成岩流体改造形成的, 受到不规则囊状化石的控制; 微生物岩主要由不规则囊状的模铸化石形成的, 而不规则囊状的模铸化石可能是某种细菌菌落形成的。何磊等(2010)认为华南的微生物岩是底栖蓝细菌形成的, 包括层纹状微生物岩、叠层石微生物岩、花斑状微生物岩、树枝状微生物岩及穹窿状微生物岩5种类型。Yang等(2011)对微生物岩中的生物化石进行了系统研究, 将树枝状微生物岩解释为凝块石。Tang等(2017)研究了四川鱼洞子剖面P-T界线地层微生物岩, 将斑点状岩石描述为凝块石, 并对微生物岩的形成环境进行了不同于前人的解释。Wu等(2017)提出, 江西修水的P-T微生物岩是凝块石和树枝石。
Wu等(2014a)提出, P-T界线地层微生物岩是由蓝细菌门微囊菌属的群体形成的模铸化石组成的; 微囊菌是一种浮游蓝细菌, 由于当时浅海地区发生海水富营养化, 在水体表层发生了微囊菌赤潮, 赤潮造成底层水缺氧。笔者认为Wu等(2014a)已经对P-T界线地层微生物岩的形成机制给出了比较合理的解释, 引起了很多学者的关注, 但还有少数学者(Wu et al., 2017; Tang et al., 2017; Kershaw et al., 2018)持不同意见。笔者认为, P-T界线地层微生物岩研究的困难在于形成微生物岩的微生物是古生物学家未曾见过、古生物学教科书和文献中未曾涉及过的特殊生物: 一种形状不规则的微生物。此前, 古生物学家研究的都是形状规则的化石, 并且主要是根据形状、形态特征来鉴定古生物的; 面对形状不规则的东西, 一时无法想象它们不是非生物构造。另一方面, 地质现象变化万千, 在研究任何一种地质现象的时候, 需要有全局意识, 要分析主要现象。也就是说, 对一个剖面、一套地层, 要进行全面的观察, 而不是局部的; 要抓住主要现象、而不是局部现象; 切忌以局部现象代替主要问题, 切忌以偏概全。限于篇幅, Wu等(2014a)未对老龙洞剖面的微生物岩特征做详细描述。本文的主要目的之一就是对老龙洞剖面P-T界线地层微生物岩的岩石学和古生物学特征做全面描述, 以展示P-T界线地层微生物岩的完整、真实面貌, 为理解微生物岩的成因提供基础。
P-T界线地层微生物岩的下伏地层是上二叠统长兴组含海百合、䗴、大有孔虫的厚层或块状灰岩; 上覆地层是下三叠统灰色薄板状泥质灰岩。P-T界线地层微生物岩本身不含大化石, 厚度一般只有2~10 m或3~15 m(王永标等, 2005), 包括牙形刺化石带Hindeodus changxingensis带的上部和H. parvus 带的下部。
根据笔者迄今对重庆北碚老龙洞、江西修水、四川宣汉、湖北利川等地7个剖面的露头观察和薄片研究以及所有相关文献, P-T界线地层微生物岩是一套形成于晚二叠世末生物大灭绝之后, 以在露头上具有树枝状、网状、斑点状花纹为特征的特殊岩石。树枝状、网状、斑点状花纹系由暗色(一般是棕色)和浅色(一般是灰色)2种岩石组成, 既可以看成浅色的树枝状岩石存在于暗色的岩石之中, 也可以看作暗色的树枝状岩石存在于浅色的岩石之中, 即2种颜色的岩石都像树枝状; 斑点状的岩石也如此。但薄片研究发现, 浅色的那部分岩石是一般的泥晶灰岩; 暗色的那部分岩石成分和结构比较复杂, 一般由较粗的晶体组成, 因较粗的晶体透光性更好, 在露头上会吸收更多的阳光, 导致其反射的阳光少, 表现为暗色。目前, 所有研究的重点都是关于暗色的那部分岩石。为了述说方便, 笔者把暗色的树枝状、网状、斑点状岩石部分称为树枝状体(dendroid bodies)、网状体(net-like bodies)、斑点状体(patchy bodies)。斑点状体之间、树枝状体之间及网状体的网眼内是泥晶灰岩。泥晶灰岩中通常有一些形状不规则、内部被各种较粗晶体胶结物充填的结构, 本文称之为亮晶充填体。斑点状体、树枝状体、网状体内部含有丰富的亮晶充填体, 但很多情况下都被成岩作用破坏, 不易识别。P-T界线地层微生物岩研究的关键问题是确定树枝状体、网状体、斑点状体和亮晶充填体的形成机制。
重庆北碚(原属于四川华蓥)老龙洞剖面长兴组顶部是厚层块状的生屑粒泥岩和泥粒岩, 含柯兰尼虫(Colaniella)、棘皮类(以海百合为主)、古䗴(Palaeofusulina)、管壳石(Tubiphytes)、腕足类等, 是大灭绝之前的沉积。这套灰岩之上即是微生物岩, 岩性为灰岩和白云岩, 厚2.2im, 薄层状, 下部层面比较平坦、上部层面波状起伏。下部具有斑点状、上部具有树枝状结构, 不再含有海百合、柯兰尼虫、古䗴等。之所以称这套地层为微生物岩, 是因为前人将其描述为微生物岩, 实际上是否为微生物岩, 需要通过研究去进一步确定。目前, 暂且以是否含有斑点状体、树枝状体、网状体作为其是否为微生物岩的判断标志。
在重庆北碚天府镇天府煤矿后山露头上, 笔者实测了3条剖面(即老虎洞剖面), 分别叫1、2、3号剖面(图1)。3个剖面之间相距不到3im, 1号剖面包括了大灭绝前的地层、微生物岩、微生物岩之上的地层, 2号、3号剖面只包括微生物岩的中上部和微生物岩之上的地层, 微生物岩的下部因植被茂盛未测量。文中以1号和3号剖面为主, 首先描述老龙洞剖面的岩石和古生物特征, 然后做出解释。
老龙洞1号剖面全长2.25 m。在野外工作中, 将1号剖面分为9层(1-1~1-9)(图2)。尽管笔者做了大量的牙形刺化石处理工作, 但在该剖面中并未发现Hindeodus parvus, 因而P-T界线不能根据牙形刺确定。在江西修水剖面, 笔者做了牙形刺化石和碳同位素研究(Wu et al., 2014b), 发现代表三叠纪开始的牙形刺H. parvus的首现面, 正好在碳同位素曲线首次下降到最低点的地方, 处于微生物岩段的中部偏下。在湖南慈利P-T界线地层剖面上, 碳同位素曲线的初始最低点也在微生物岩的中部(Luo et al., 2014; Wang et al., 2016)。Wu 等(2017)也研究了修水剖面P-T界线地层微生物岩的碳同位素和牙形刺化石, 该剖面中的碳同位素曲线初始最低点位于微生物岩段的中部偏上, 距微生物岩段的底界约1 m。Mu等(2009)研究了老龙洞P-T界线地层剖面的碳、氧同位素, 发现碳同位素曲线的初始最低点(其标本T6-2)在微生物岩底界之上1.45im处, 即在微生物岩段的中部偏上处。笔者以上述地层分界作为P-T界线的位置(图2)。
1-1~1-2层: 生屑泥粒岩, 不含斑点状体、树枝状体、网状体。生屑包括古䗴(Palaeofusulina)、柯兰尼虫、有孔虫和苔虫等。
1-3层: 含斑点状体的泥晶灰岩。露头上(图3-A)和薄片中都可见到斑点状体。斑点状体(图4-A, 4-C: 红色虚线内)形状不规则, 宽一般为3~8 mm, 由大块状方解石亮晶(图4-A: c)和其间的泥晶(图4-C: m)组成。斑点状体之间为泥晶, 其中有较多的形状不规则、内部被各种胶结物充填的亮晶充填体(图4-C: 黄色虚线内)和一些介形虫(图4-C: o)。泥晶中的亮晶充填体(图4-C: 黄色虚线内)内部由白色大块状方解石组成, 边缘有少数白色针状方解石。斑点状体内部的大块状方解石亮晶, 一般都含有漂浮状的暗色菱形方解石晶体以及少数针状方解石(图4-D: nc), 部分菱形方解石晶体具环带构造(图4-B: r)。亮晶充填体没有壁, 形状不规则, 无定向性。该层中部含有较多的丝状钙化蓝细菌(图4-E, 4-F: f), 丝状蓝细菌有一端弯曲的现象(图4-F: f)。
解释: (1)沉积物不可能是漂浮状。本层斑点状体内部大块状方解石中的菱形方解石晶体是漂浮状的, 说明菱形方解石晶体是溶蚀残余的, 原岩被溶蚀成蜂窝状, 大块状方解石充填在蜂窝状溶蚀孔洞内。(2)环带状菱形方解石不是原生的。只有白云石才会有环带状构造, 所以环带状方解石是环带状白云石通过去白云石化形成的, 说明大块状方解石形成之前不仅发生了溶蚀作用, 还发生了去白云石化作用; 去白云石化作用和溶蚀作用可能是同源淡水流体造成的, 因为它们都需要淡水; (3)针状方解石一般是针状文石转变来的, 针状文石是最早的海底胶结物; (4)可以恢复亮晶充填体的成岩历史。泥晶中的亮晶充填体的成岩历史为: 针状文石形成→ 针状文石转变为针状方解石→ 淡水溶蚀作用形成孔洞→ 大块状方解石胶结物充填所有溶蚀孔洞; 斑点状体中的亮晶充填体的成岩历史为: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 淡水造成去白云石化使菱形白云石变成菱形方解石→ 淡水溶蚀作用使岩石成蜂窝状→ 大块状方解石胶结物充填所有溶蚀孔洞。斑点状体内部的亮晶充填体经历了更多的成岩作用, 包括白云石化作用、去白云石化作用和更强的溶蚀作用。斑点状体内部亮晶充填体之间的泥晶也遭受成岩作用影响, 晶体有一定程度的增大。
因此, 斑点状体的形成主要是成岩作用的结果。斑点状体内部的亮晶充填体内部包括了6种成岩作用形成的3种胶结物。如果没有这些胶结物, 亮晶充填体就是一个个孔洞, 岩石就是孔洞状的泥晶灰岩。是胶结物, 特别是大块状方解石, 赋予了岩石斑点状特征。大块状方解石形成的斑点状构造, 除了姜红霞和吴亚生(2007), 以前的学者都没有认识到。
有人把丝状蓝细菌解释为有孔虫, 对此笔者不认同。笔者认为, 厚度不均匀的泥晶钙化壁、一端弯曲是蓝细菌胶须菌的一个特征。本层的丝状蓝细菌可能是钙化的胶须菌。
1-4层: 主要由网状体组成的次生灰岩。在露头上, 网状体呈棕色, 灰色泥晶呈网眼状。在薄片中, 网状体由大块方解石(图4-H: c)和漂浮于其中的黄色粒状方解石(茜素红染色变红)、泥晶(图4-H: m, 几个很小的晶体)、菱形方解石晶体(图4-H: r, 颜色浅)、一些全脐螺类和小腹足类(图4-H: g, 螺旋形)或虫管化石组成。网眼中为泥晶, 有少数轮廓完好的亮晶充填体(图4-G: 黄色虚线内)。
解释: (1)菱形方解石晶体的存在, 说明存在过菱形白云石晶体和发生过去白云石化作用; 大块状亮晶方解石以及其中漂浮状溶蚀残余的存在, 说明曾经发生过强烈溶蚀作用使网状体内岩石呈蜂窝状。根据网眼中的泥晶中存在亮晶充填体以及与层1-3的类比, 推测本层网状体内部曾经有很多的亮晶充填体, 但它们的轮廓已经被强烈的溶蚀作用破坏了; 本层亮晶充填体的成岩历史与1-3层斑点状体内的亮晶充填体的相似, 但本层中针状方解石几乎全部被溶蚀, 未见残余; (2)本层的螺旋状化石壳壁具有纹层状结构, 为隐晶方解石质的, 可能属于腹足动物门全脐螺类的(姜红霞等, 2010); 有些拉长螺旋形的化石, 与其他学者鉴定的虫管(Yang et al., 2015)很相似, 所以可能2种化石都有。本层与1-3层的区别是小腹足类化石局部较丰富, 网状体内成岩作用更强烈, 导致亮晶充填体无法辨认。
1-5层(下): 具斑点状体的泥晶灰岩。露头上(图3-C)和薄片中都可见到斑点状。在露头上, 棕色的斑点状体占50%以上; 在薄片中, 斑点状体(图5-A: 红色虚线内是一个斑点状体的局部)由丰富的亮晶充填体(图5-A: 有50个以上, 其中2个用黄色虚线标出轮廓)、大块状方解石(图5-A: c)及其间的微亮晶(图5-A: 如紫色虚线内)组成。亮晶充填体呈不规则状, 大小一般为0.1~1 mm, 边缘被黄色白云石(图5-B, 5-C: yd)充填, 内部被白色粒状方解石(图5-B: gc)充填, 黄色白云石和白色粒状方解石粒径为0.05~0.08 mm, 或内部被大块状方解石(图5-C: c)充填, 且含有针状方解石(图5-C: nc)、白色菱形方解石(图5-C: r)。有少数介形虫, 两壳瓣分离或不分离。无论是斑点状体内部还是之间, 亮晶充填体都不具有可以识别的生长方向, 是随机分布的。
解释: 本层的特点是斑点状体内的亮晶充填体有50%保留轮廓, 20%保留部分轮廓, 30%轮廓不好识别, 说明溶蚀作用不强烈。黄色的边缘是识别亮晶充填体的重要依据。本层“ 黄色白云石+白色粒状方解石” 型亮晶充填体的成岩历史是: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 亮晶充填体内部去白云石化形成白色粒状方解石。“ 针状方解石+菱形方解石+大块状方解石” 型亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 去白云石化使菱形白云石变成菱形方解石→ 部分亮晶充填体内部发生淡水溶蚀作用→ 大块状方解石胶结物充填所有溶蚀孔洞。
1-5层(上): 主要由斑点状体组成的灰岩。露头上可见棕色的斑点状体分布在不规则状的灰色泥晶之中, 斑点状体(图5-D: 红色虚线内; 5— E: 照片的浅色区域)占岩石体积50%以上。斑点状体之间是泥晶(图5-D, 5-E: 暗色区域), 含有一些轮廓清晰的亮晶充填体(图5-D: s1, 黄色虚线内)。斑点状体由亮晶充填体(图5-D, 5-E: 黄色虚线内)和其间的微亮晶(图5-D, 5-E: 粉色虚线包围的部分)组成。斑点状体内的亮晶充填体大多数轮廓不清晰(图5-D: s2, 黄色虚线内), 但斑点状体边缘的亮晶充填体至少保存了50%的轮廓(图5-D: s3, 黄色虚线内)。推测斑点状体内部有些亮晶充填体已经被完全改造为块状亮晶方解石, 因而无法辨认。
解释: 推测斑点状体内部的亮晶充填体本来很丰富, 但是由于成岩作用改造, 一部分已经无法识别。微亮晶可能是亮晶充填体之间的泥晶重结晶形成的, 也可能是亮晶充填体被成岩作用改造形成的。亮晶充填体的成岩历史可能与1-5层(下)相同。
1-6层: 主要由斑点状体组成的灰岩。露头上不清晰(图3-D), 薄片上可见斑点状体(图5-F: 红色虚线内)由块状亮晶方解石(图5-F: c)、暗色粒状方解石(图5-F: 白色虚线内)和溶蚀残余的泥晶(图6-A: m)组成。块状方解石中有漂浮状的暗色菱形方解石晶体(图5-G: r; 图6-A: r)。斑点状体之间为泥晶, 泥晶中有少数轮廓分明的亮晶充填体(图5-F: s1, 黄色虚线内)、少数介形虫、小腹足类, 介形虫多数壳瓣分离。斑点状体内部的亮晶充填体已经无法辨认, 边缘的亮晶充填体(图5-F: s2, 黄色虚线内)还保留部分轮廓。
解释: (1)斑点状体内部成岩作用强烈, 使得亮晶充填体无法识别, 但是其边缘残余的亮晶充填体形态以及斑点状体之间泥晶中亮晶充填体的存在, 说明斑点状体内部曾经含有丰富的亮晶充填体, 而溶蚀残留的泥晶, 是原来亮晶充填体之间的泥晶; (2)根据针状方解石、菱形方解石、块状亮晶方解石的存在, 恢复亮晶充填体的成岩历史为: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 淡水造成去白云石化使菱形白云石变成菱形方解石和粒状方解石→ 部分亮晶充填体内部发生淡水溶蚀作用→ 大块状方解石胶结物充填所有溶蚀孔洞。
1-7层: 主要由树枝状体组成的岩石。露头上可见明显的暗色树枝状体(图3-E, 3-F), 占岩石体积的70%以上, 主延伸方向垂直于地层层面。在薄片上, 树枝状体(图6-B: 红色虚线内, 4个)由丰富到密集的亮晶充填体(图6-B, 6-C: 黄色虚线内)和其间的微亮晶(图6-B, 6-C: 紫色虚线内)组成。一些亮晶充填体全部由黄色半自形白云石晶体组成(图6-C: 黄色箭头s1所指黄色虚线内), 更多的亮晶充填体(图6-C: 黄色箭头s2所指黄色虚线内)的边缘由黄色半自形粒状白云石组成(图6-C: yd), 内部由白色他形粒状方解石组成(图6-C: gc)。大多数亮晶充填体保留了轮廓(图6-B: 例如, 黄色虚线内), 但一些可能因为去白云石化作用比较强烈, 已经没有了黄色边缘, 因而识别困难(图6-B: 蓝色虚线内), 只有尚存的少数白色菱形方解石晶体是识别的依据。树枝状体之间是泥晶。本层上部, 树枝状体内的一些亮晶充填体被大块状白色方解石(图6-D: c, 白色虚线内是1个晶体, 正交偏光下整体消光)充填, 这些大块状方解石中一般含有漂浮状的暗色菱形方解石晶体(图6-D: r), 边缘有少数针状方解石(图6-E: nc)和黄色自形白云石晶体(图6-E: yd)。树枝状体之间的泥晶中有少数介形虫壳(图6-D: o)和亮晶充填体(图6-D: 黄色虚线内)。
解释: (1)与1-5层和1-6层相比, 1-7层的树枝状体含量最高。(2)树枝状体由密集的亮晶充填体组成。(3)亮晶充填体大多数轮廓保存较好。(4)亮晶充填体包括3种类型; 类型1(全部由黄色白云石晶体组成)的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石晶体; 类型2(黄色粒状白云石+白色粒状方解石)的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石晶体→ 淡水造成亮晶充填体内部去白云石化使黄色粒状白云石变成白色粒状方解石; 类型3(针状方解石+黄色白云石+大块状方解石)的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石晶体→ 淡水造成去白云石化使黄色白云石变成菱形方解石和粒状方解石→ 淡水溶蚀作用使亮晶充填体内部变成蜂窝状→ 大块状方解石胶结物充填所有溶蚀孔洞。(5)一些亮晶充填体内部有微亮晶, 说明其前身生物内部有孔洞, 内部有孔洞的生物如现生蓝细菌门微囊菌属的群体。(6)所有的亮晶充填体没有定向性。
1-8层: 主要由树枝状体组成的岩石。露头上树枝状特征明显(图3-G), 体积占到岩石的70%以上。在薄片中, 树枝状体(图6-F: 红色虚线内)边缘的亮晶充填体(图6-G: 例如, 黄色虚线内)尚可辨认, 其边缘由黄色半自形粒状白云石组成(图6-G: yd), 内部由白色粒状方解石组成(图6-G: gc)。树枝状体内部的亮晶充填体大多被成岩作用破坏, 轮廓无法识别, 现由块状方解石(图6-G: c)、斑状分布的黄色粒状白云石(图6-G: yd)、斑状分布的白色粒状方解石(图6-G: gc)、少数针状方解石(图6-G: nc)、少数斑状分布的泥晶(图6-G: m)组成。树枝状体边缘有少数轮廓保存较好的亮晶充填体(图6-G: 黄色箭头s1所指的黄色虚线内), 完全由黄色粒状白云石组成。树枝状体之间是泥晶, 含有一些轮廓保存较好的亮晶充填体(图6-H: 黄色虚线内)。
解释: (1)本层树枝状体内部的亮晶充填体大多数已被成岩作用破坏, 轮廓无法识别。但黄色粒状白云石、白色粒状方解石、针状方解石、大块状亮晶方解石这些矿物成分与前面描述的保存有轮廓的亮晶充填体中的一样, 所以属于被破坏的亮晶充填体的; 树枝状体边缘以及树枝状体之间的泥晶中有保存较好的亮晶充填体, 说明树枝状体内部被破坏的是亮晶充填体; (2)树枝状体内部“ 针状方解石+粒状方解石+块状方解石” 型亮晶充填体经历的成岩作用: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 淡水造成去白云石化使菱形白云石变成菱形方解石和粒状方解石→ 淡水溶蚀作用使亮晶充填体成蜂窝状→ 大块状方解石胶结物充填所有溶蚀孔洞; “ 黄色粒状白云石+白色粒状方解石” 型亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石晶体→ 淡水造成去白云石化使菱形白云石变成菱形方解石和粒状方解石; 只由黄色粒状白云石组成的亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石晶体。
1-9层: 具树枝状体的生屑泥粒岩。在露头上, 树枝状特征明显(图3-H), 由棕色树枝状体和其间的浅灰色泥晶灰岩组成, 垂直于层面延伸。在薄片中, 树枝状体(图7-A: 右边一半, 浅色)由大块状亮晶方解石(图7-A: c)和其间的微亮晶(图7-A: 紫色虚线内)组成。树枝状体内含有丰富的小腹足类(图7-A: 箭头所指; 7-B: g)、介形虫等的壳(图7-A: 箭头所指的; 7-B: o)。树枝状体之间(图7-A: 左边一半, 深色)含有丰富的小腹足类等化石, 属于生屑粒泥岩— 泥粒岩。小腹足类和介形虫壳内的充填物包括块状亮晶方解石(图7-B: c)、白色粒状方解石(图7-B: gc)。块状方解石中有少数漂浮状的黑色菱形方解石晶体(图7-B: r)。树枝状体内小腹足类和介形虫的壁的形态都可以分辨(图7-B: 红色箭头所指)。
解释: (1)虽然本层在露头上树枝状构造明显, 但其树枝状体是由充填生物壳的块状方解石、粒状方解石以及生物壳之间的微亮晶形成的。由于微亮晶是由泥晶重结晶形成的, 所以本层实际上不是微生物岩, 而是一般的生屑泥粒岩。这说明, 有树枝状构造, 不一定是微生物岩。(2)生物壳内部经历的成岩历史包括: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石晶体→ 淡水造成去白云石化使菱形白云石变成菱形方解石和粒状方解石→ 淡水溶蚀作用使大多数粒状方解石溶蚀掉→ 块状方解石胶结物充填所有溶蚀孔洞。(3)本层树枝状体内的生物化石的壳都保存较好, 这说明如果有钙化的化石, 其钙化壁应当或多或少地被保存下来。
老龙洞3号剖面位于1号剖面之南, 相距5 m。野外分为20层(3-1~3-20)(图8)。3-1~3-7层露头上具斑点状外貌(图9-A), 3-8~3-11层露头上具树枝状外貌(图9-C), 3-12层以上无树枝状外貌(图9-D)。笔者做了大量牙形刺化石处理, 但未获得牙形刺化石Hindeodus parvus。所以, P-T界线不能依靠牙形刺化石确定。由于该剖面与1号剖面距离只有5im, 所以参照1号剖面, 把P-T界线位置放在3-3层之上。与1号剖面相同, P-T界线都是位于斑点状岩石的上部, 也就是小球状化石(Wu et al., 2016)开始出现的地方。
 | 图9 重庆老龙洞3号剖面P-T界线地层微生物岩露头特征 A— 3-3层~3-7层, 斑点状; B— 3-7层~3-12层, 树枝状; C— 3-8层~3-12层, 树枝状; D-3-13~3-17层。L3-3指示标本L3-3取样位置Fig.9 Outcrop features of the P-T boundary microbialite interval of section 3 at Laolongdong, Chongqing |
3-1层: 在露头上, 看不清斑点状特征, 但在薄片中, 其为具斑点状体的灰岩。斑点状体形状不规则(图7-C: 红色虚线内), 由白色微亮晶(图7-C: 粉色虚线内)、白色粒状方解石、黄色粒状白云石(图7-C: yd)和少数块状方解石(图7-C: c)组成。不均匀分布的黄色粒状白云石、白色粒状方解石、微亮晶是亮晶充填体遭受成岩作用改造后形成的(图7-C: 黄色虚线)。尽管亮晶充填体的轮廓已经无法辨认, 但隐约的黄色晶体(图7-C: yd), 代表了曾经的亮晶充填体的黄色白云石的边缘(如上文所述)。在斑点状体边缘, 有一些亮晶充填体的轮廓尚可识别, 特别是黄色的亮晶白云石晶体尚可分辨(图7-C: s1)。2个亮晶充填体的内侧(图7-C: a所在的位置)都变成了微亮晶, 说明微亮晶可能既来自于亮晶充填体之间的泥晶重结晶, 也可能是亮晶充填体本身去白云石化。斑点状体之间为泥晶, 其中有少数轮廓清晰的亮晶充填体(图7-C: s2)。这些亮晶充填体形状不规则, 内部见针状胶结物(图7-D: nc)。斑点状体之间有很少的介形虫(图7-C: o), 它们的2个壳瓣多不分离。
3-2~3-3层: 具斑点状体的灰岩。斑点状体(图7-E: 红色虚线内)由亮晶充填体和其间的微亮晶(图7-E: 绿色虚线内)组成。少数位于斑点状体边缘的亮晶充填体轮廓清晰(图7-E: 黄色虚线内4个), 但大多数亮晶充填体的轮廓已被成岩作用破坏, 无法分辨(图7-E)。轮廓清晰的亮晶充填体的边缘呈黄色(图7-E: e), 或内部全部被黄色的粒状白云石充填(图7-E: 黄色虚线内)。在一些亮晶充填体表面, 局部有暗色泥晶聚集体(图7-F: dm), 它们厚度很不均匀, 如图7-E中黄色箭头所指的亮晶充填体, 左侧有一个疙瘩状的泥晶聚集体(ag), 右侧黄色箭头所指处却没有泥晶聚集体; 局部见到亮晶充填体之间的白色针状胶结物(图7-F: 白色虚线内)。而亮晶充填体内部, 一些完全被块状的亮晶方解石充填(图7-E: c); 一些可见3种胶结物: 边缘1层黄色粒状白云石(图7-G: yd)、针状方解石(图7-G: nc)、内部的大块状方解石胶结物(图7-G: c)。黄色针状方解石是垂直于亮晶充填体的边界朝内生长的。
解释: (1)本层具有该剖面保存最好的亮晶充填体, 不仅保留了内部的黄色白云石晶体, 还保留了外表吸附的泥晶聚集体; (2)泥晶聚集体厚度很不均一, 甚至有些地方缺失, 因此它不是生物的体壁, 亮晶充填体不是钙化生物; 另外, 钙化蓝细菌, 如肾形菌的钙化壁是隐晶质的, 方解石晶体要比泥晶细一些; (3)多数情况下, 亮晶充填体之间是泥晶, 但本层见到亮晶充填体之间是胶结物的情形, 说明2种情况都是存在的; (4)本层有3种结构的亮晶充填体: Ⅰ . 全部被黄色粒状白云石充填的, 其成岩历史是: 针状文石形成→ 针状文石转变为针状方解石→ 针状方解石发生白云石化形成黄色粒状白云石; Ⅱ . 边缘由黄色粒状白云石组成, 内部由白色粒状方解石组成的, 其成岩历史是: 针状文石形成→ 针状文石转变为针状方解石→ 针状方解石发生白云石化形成黄色粒状白云石→ 去白云石化使内部变成白色粒状方解石; Ⅲ . 边缘由黄色粒状白云石组成, 还有少数针状方解石, 内部被大块方解石充填的, 其成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 针状方解石发生白云石化形成黄色粒状白云石→ 去白云石化使内部变成白色粒状方解石→ 溶蚀作用消灭所有的粒状方解石, 形成溶蚀孔洞→ 大块状亮晶方解石充填所有溶蚀孔洞。
3-4层: 具斑点状体的灰岩(图9-A)。斑点状体由大块状方解石(图7-H: c; 图10-B: c)、小腹足类(图7-H: g)、微亮晶(图7-H: 紫色虚线内)组成。斑点状体之间是泥晶, 有较多的介形虫(图10-A: o)、小腹足类(图10-A: g)、全脐螺或虫管化石(图10-A: g1)。此层应该与1号剖面的含小腹足类层是同一层。介形虫多数壳瓣分离, 少数壳瓣不分离。小腹足类有高塔形的(图10-A: g)。斑点状体内的大块状方解石包含有针状方解石(图10-B: nc)和黑色菱形方解石晶体(图10-B: r)。
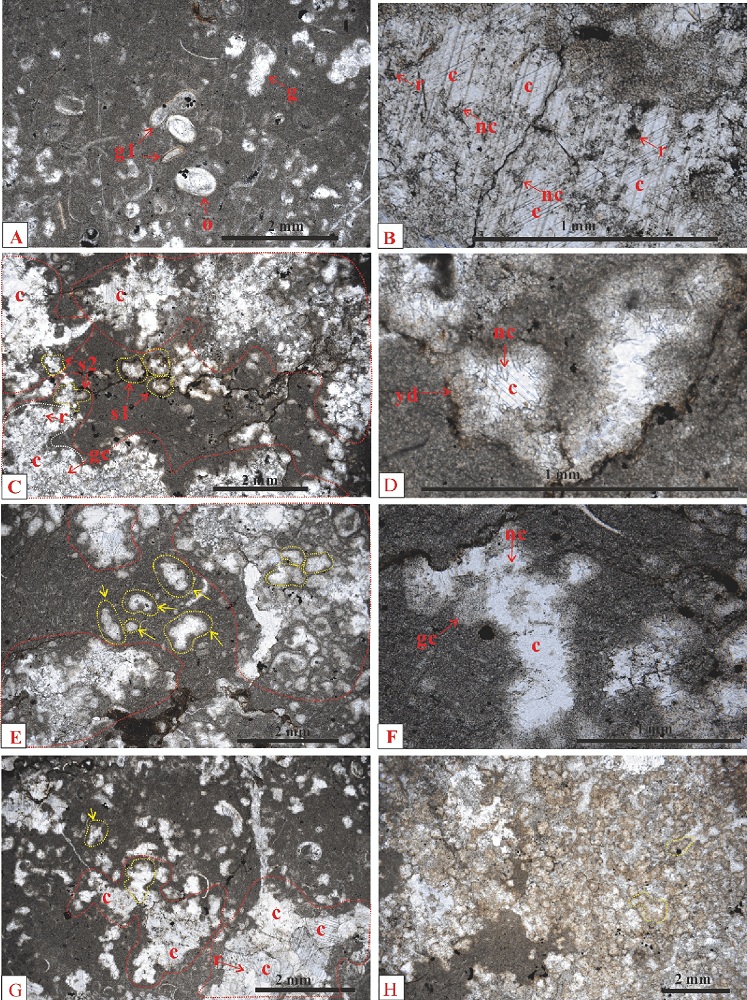 | 图10 重庆老龙洞3号剖面第3-4~3-8层微生物岩显微特征 A— 3-4层, 具斑点状体的灰岩, 斑点状体之间的泥晶中有较多的介形虫(o)、小腹足类(g)、全脐螺或虫管化石(g1), 小腹足类有高塔形的(g); B— 3-4层, 斑点状体局部放大, 主要由大块状方解石(c)组成, 其中含有漂浮状针状方解石(nc)和个别黑色菱形方解石晶体; C— 3-5层, 具斑点状体的灰岩, 斑点状体(红色虚线内)由亮晶充填体和其间的微亮晶组成, 斑点状体内部多被块状方解石(c)充填, 轮廓多被破坏(例如, 白色虚线内); 有些块状方解石很大(例如, 白色虚线内是1个连晶方解石晶体, 正交偏光下整体消光), 其中有斑状分布的暗色粒状方解石(gc), 偶见黑色菱形方解石(r); 斑点状体边缘的一些亮晶充填体虽然轮廓被保存(s2), 但一般不具有黄色粒状白云石组成的边缘; 斑点状体之间为泥晶, 其中有一些亮晶充填体轮廓保存完好(例如, s1); D— 图C中的1个亮晶充填体的放大, 边缘有黄色粒状白云石(yd)和针状方解石(nc), 内部由大块状方解石(c)充填; E— 3-6层, 具斑点状体的灰岩, 斑点状体(红色虚线内)由丰富的亮晶充填体和大块状方解石组成, 内部的亮晶充填体有一半以上保存了轮廓(例如, 右边黄色虚线内); 大块状方解石是对1个到多个亮晶充填体的溶蚀孔洞的充填, 亮晶充填体轮廓被破坏, 无法识别; 斑点状体之间为泥晶, 有较多的亮晶充填体(例如, 黄色箭头所指黄色虚线内); F— 3-6层, 1个亮晶充填体的放大, 其边缘有暗色粒状方解石(gc), 边缘和内部有少数针状方解石(nc), 内部是大块状方解石(c); G— 3-7层, 具少数斑点状体的粒泥岩, 斑点状体由亮晶充填体(例如, 黄色虚线内是1个亮晶充填体)和大块状方解石(例如, c)组成; 大块状方解石包含了多个亮晶充填体, 破坏了它们的轮廓, 内部有漂浮状的暗色菱形方解石(r); 一些地方只看到大块状方解石, 亮晶充填体的轮廓已经被完全破坏(右下角红色虚线内); 粒泥岩的粒是亮晶充填体(例如, 黄色箭头所指黄色虚线内)和介形虫; H— 3-8层, 主要由树枝状体组成的岩石, 树枝状体(整个照片是1个树枝状体的局部放大)呈浅色, 由密集的亮晶充填体(黄色虚线内的2个)和其间的微亮晶组成; 亮晶充填体一般边缘由黄色白云石晶体组成, 内部由白色粒状方解石组成Fig.10 Microscopic features of microbialites of beds 3-4 to 3-8 of section 3 at Laolongdong, Chongqing |
解释: 本层斑点状体内的亮晶充填体轮廓已经被成岩作用破坏, 无法识别。但是, “ 针状方解石+黑色菱形方解石+大块状方解石” 的矿物组成是亮晶充填体曾经存在的证据, 它们经历的成岩历史为: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石→ 去白云石化作用形成黑色菱形方解石→ 溶蚀作用使亮晶充填体成为蜂窝状→ 大块状方解石充填所有溶蚀孔洞。
3-5层: 具斑点状体的灰岩(图9-A)。斑点状体(图10-C: 红色虚线内)由亮晶充填体和其间的微亮晶组成。斑点状体内部的亮晶充填体大多数被形状不规则的块状亮晶方解石(图10-C: c)充填, 轮廓大多数被破坏(图10-C: 白色虚线内)。有些块状方解石很大(图10-C: 白色虚线内是1个连晶方解石晶体), 其中有斑状分布的暗色粒状方解石(图10-C: gc), 偶见黑色菱形方解石(图10-C: r)。斑点状体边缘的一些亮晶充填体虽然轮廓被保存下来(图10-C: s2), 但一般不具有黄色粒状白云石组成的边缘。斑点状体之间为泥晶, 其中有一些分散保存的亮晶充填轮廓保存完好(图10-C: 例如, s1), 一般有黄色粒状白云石组成的边缘的残留(图10-D: yd), 有一些针状方解石(图10-D: nc), 内部被大块方解石(图10-D: c)充填。
解释: (1)斑点状体内部的亮晶充填体因经历强烈溶蚀作用, 轮廓无法识别, 但有粒状方解石和少数黑色菱形方解石的残余; 斑点状体边缘的亮晶充填体轮廓尚可识别; 斑点状体之间的泥晶中的亮晶充填体轮廓保存较好; (2)亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石→ 去白云石化作用形成黑色菱形方解石→ 溶蚀作用使亮晶充填体成为蜂窝状→ 大块状方解石充填所有溶蚀孔洞。
3-6层: 具斑点状体的灰岩(图9-A)。斑点状体占岩石体积的30%~40%。斑点状体(图10-E: 红色虚线内)由丰富的亮晶充填体和大块状亮晶方解石组成。斑点状体内部的亮晶充填体有一半以上保存了轮廓(图10-E: 右边黄色虚线内)。大块状方解石是对1个到多个亮晶充填体的溶蚀孔洞的充填, 使其中的亮晶充填体轮廓被破坏, 无法识别(图10-E)。斑点状体之间为泥晶, 有较多的亮晶充填体(图10-E: 黄色箭头所指的黄色虚线内)。大块状方解石的边缘有暗色的粒状方解石(图10-F: gc), 边缘和内部还有少数针状的方解石(图10-F: nc)。
解释: 本层的斑点状体中保存了轮廓的亮晶充填体较多, 反映溶蚀作用不强。亮晶充填体内部的矿物特征与3-4层相似, 所以有相似的成岩经历。
3-7层: 具少数斑点状体的粒泥岩(图9-A)。斑点状体占岩石体积的30%。斑点状体由亮晶充填体(图10-G: 黄色虚线内)和大块状亮晶方解石(图10-G: c)组成。大块状方解石包含多个亮晶充填体, 破坏了它们的轮廓, 内部有漂浮状的暗色菱形方解石(图10-G: r)。部分区域只看到大块方解石, 亮晶充填体的轮廓已经被完全破坏(图10-G: 右下角红色虚线内)。粒泥岩的粒是亮晶充填体(图10-G: 黄色箭头所指黄色虚线内)和介形虫, 50%的介形虫壳瓣不分离。
解释: (1)较强的溶蚀作用使得很多亮晶充填体无法识别; (2)成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成菱形白云石→ 去白云石化作用形成黑色菱形方解石→ 溶蚀作用使亮晶充填体成为蜂窝状→ 大块状方解石充填所有溶蚀孔洞; (3)露头上, 3-7层呈楔形, 说明3-7层可能遭受了剥蚀作用。
3-8层: 主要由树枝状体组成的岩石(图9-B, 9-C)。树枝状体在露头上很明显, 暗色, 垂直于地层层面延伸, 大约占岩石体积的70%以上。在薄片中, 树枝状体(图10-H:整个照片是1个树枝状体的局部放大)呈浅色, 由密集的亮晶充填体(图10-H: 黄色虚线内的2个)和其间的微亮晶组成。亮晶充填体一般边缘由黄色白云石晶体组成, 内部由白色粒状方解石组成, 轮廓一般保存较好, 形状多样。亮晶充填体本身没有任何定向性, 不具有附着生活的特征。磨片缺少3-9层。
解释: 本层树枝状特征明显, 树枝状体内亮晶充填体密集, 亮晶充填体未经历强烈溶蚀作用, 所以大多数保存了轮廓。大多数亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石→ 亮晶充填体内部去白云石化作用→ 少数亮晶充填体内部发生溶蚀作用→ 大块状方解石充填溶蚀孔洞。
3-10层: 主要由树枝状体组成的岩石(图9-C)。在露头上, 树枝状体垂直于地层延伸。在薄片上, 树枝状体(图11-A: 红色虚线内)由斑状分布的微黄色的粒状白云石和其间的白色粒状方解石混杂组成。树枝状体之间为泥晶, 有少数亮晶充填体(图11-A: 黄色箭头所指黄色虚线内)轮廓较好, 边缘由黄色粒状白云石组成。
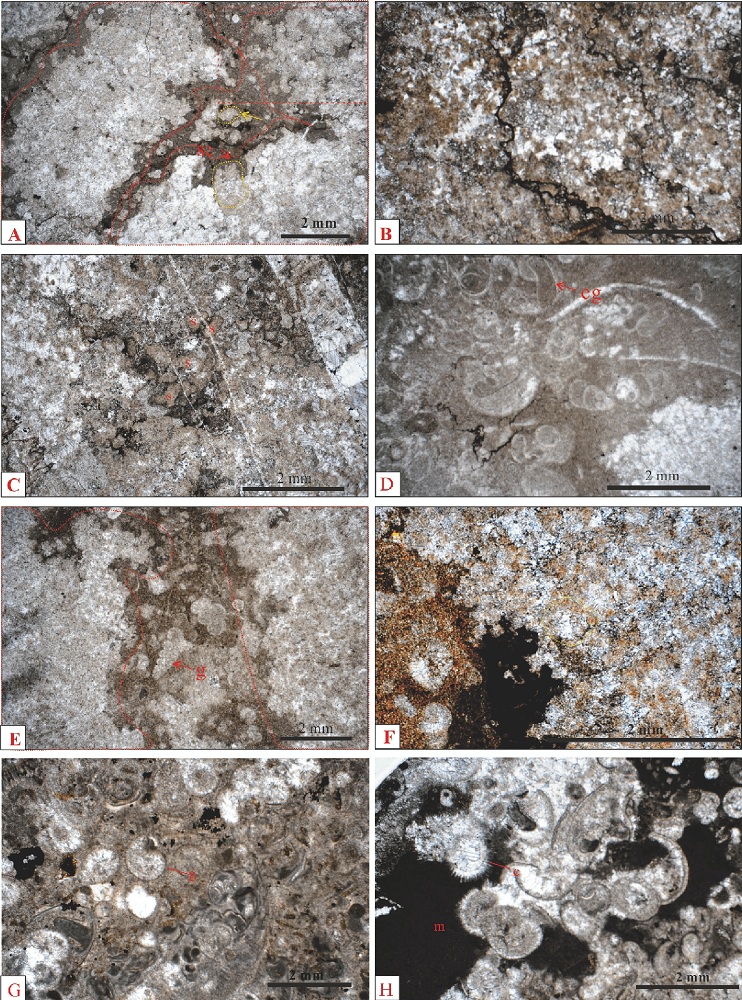 | 图11 重庆老龙洞3号剖面第3-8~3-15层显微特征 A— 3-10层, 主要由树枝状体组成的岩石; 树枝状体(红色虚线内)由斑状分布的微黄色的粒状白云石和其间的白色粒状方解石混杂组成; 树枝状体之间为泥晶, 其中有少数亮晶充填体(例如, 黄色箭头所指黄色虚线内)轮廓较好, 亮晶充填体的边缘由黄色粒状白云石组成; B— 3-11层, 主要由树枝状体组成的岩石; 树枝状体(整个照片是1个树枝状体的局部)由密集的亮晶充填体和其间的微亮晶组成; 大多数亮晶充填体主要由黄色白云石晶体组成, 仅中央由白色粒状或块状方解石组成; C— 3-12层, 树枝状体的局部放大, 全部由亮晶充填体和其间的微亮晶组成; 一部分亮晶充填体全部由黄色粒状白云石组成; 由于过于密集, 亮晶充填体之间的界线较难确定, 但少数轮廓仍然可识别(s); D— 3-13层, 为小腹足类泥粒岩; 小腹足类一般为塔形螺; 有全脐螺或虫管(eg); E— 3-14层, 具典型树枝状体的岩石; 树枝状体(红色虚线内)由斑状分布的黄色白云石晶体和白色粒状方解石晶体混杂而成; 亮晶充填体轮廓大多难以确定; 树枝状体之间一般为泥晶, 局部含小腹足类, 特别是高塔形的小腹足类(g); F— 3-14层, 照片右上部是1个树枝状体的局部放大, 由斑状分布的黄色白云石晶体和白色粒状方解石晶体混杂而成, 局部可见亮晶充填体的黄色轮廓(例如, 黄色虚线内)比较模糊和难以辨认; G— 3-15层, 小腹足类颗粒灰岩, 生屑主要是小腹足类壳(g); H— 3-15层, 局部为泥晶(m), 一些小腹足类的壳被大块状方解石充填Fig.11 Microscopic features of microbialites of beds 3-8~3-15 of section 3 at Laolongdong, Chongqing |
解释: 本层树枝状特征典型。树枝状体内的亮晶充填体轮廓不清晰, 但有一定程度的保存。大多数亮晶充填体的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石→ 亮晶充填体内大多数白云石晶体发生去白云石化作用→ 少数亮晶充填体内部发生溶蚀作用→ 大块状方解石充填溶蚀孔洞。
3-11~3-12层: 主要由树枝状体组成的岩石(图9-C)。树枝状体占岩石体积的70%。在露头上, 树枝状特征明显, 树枝状体垂直于地层层面延伸。在薄片上, 树枝状体由密集的亮晶充填体和其间的微亮晶组成(图11-B)。大多数亮晶充填体主要由黄色白云石晶体组成, 仅中央由白色粒状或块状方解石组成。一部分亮晶充填体全部由黄色粒状白云石组成。由于过于密集, 亮晶充填体之间的界线较难确定, 但少数轮廓仍然可识别(图11-C: s)。
解释: (1)本层一部分亮晶充填体全部由黄色白云石组成, 它们的成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石; (2)本层大多数亮晶充填体边缘由黄色白云石晶体组成, 内部由白色粒状方解石或块状方解石组成, 成岩历史: 针状文石形成→ 针状文石转变为针状方解石→ 白云石化作用形成黄色粒状白云石→ 亮晶充填体内部去白云石化作用→ 亮晶充填体内部发生溶蚀作用→ 大块状方解石充填溶蚀孔洞。
3-13层:为小腹足类泥粒岩(图11-D)。一般为塔形螺, 有全脐螺或者虫管化石(图11-D: eg)。
3-14层: 主要由树枝状体组成的岩石。树枝状体占岩石体积的70%以上, 形状典型, 垂直于地层层面延伸。在薄片上, 树枝状体(图11-E: 红色虚线内)由斑状分布的黄色白云石晶体和白色粒状方解石晶体混杂而成。亮晶充填体轮廓大多难以确定, 局部可见亮晶充填体的轮廓(图11-F: 黄色虚线内)。树枝状体之间一般为泥晶, 局部含小腹足类, 特别是高塔形的小腹足类(图11-E: g)。
解释: 本层的树枝状形态最典型, 是以前的微生物岩研究者关注的重点, 无论Kershaw等(2007)还是Ezaki(2003)的文章, 照片都是本层这种特征的。由于受成岩作用影响, 本层的树枝状体内的亮晶充填体的轮廓难以辨认, 但局部还是可以识别。斑状分布的黄色粒状白云石是亮晶充填体的边缘, 它们很密集, 代表亮晶充填体很密集。
3-15层: 为小腹足类颗粒灰岩。小腹足类几乎全部是低塔形的。
限于文章篇幅, 对3-15层之上的地层和2号剖面不做描述。
5.1.1 斑点状、网状、树枝状特征是成岩作用形成的
露头上的斑点状、网状、树枝状特征是P-T界线地层微生物岩的最显著特征。分析暗色的斑点状体、树枝状体、网状体的形成机制, 就成了认识这套岩石的关键。Ezaki等(2003)把树枝状体解释为凝块石, 所以把这套地层归入微生物岩的范畴。但是, 从上文对2条剖面的描述可以看出, 树枝状体不一定是微生物岩: 1号剖面上具有斑点状体、树枝状体、网状体的地层包括1-3~1-9层, 以1-7、1-8、1-9层的树枝状体最典型。虽然1-7层和1-8层看不到多少熟悉的生物化石, 但1-9层却是由熟悉的生物化石组成的, 如小腹足类和介形虫类等化石。1-9层中的树枝状体内部的化石类型和数量与其间的相似, 但树枝状体内部有较多的大块状方解石亮晶, 一个晶体的大小可以包含几个化石以及化石之间的地方。在大晶体之间的地方, 泥晶重结晶形成微亮晶。大的块状亮晶与它们之间的微亮晶, 共同构成了树枝状体。大块状亮晶方解石和微亮晶的形成, 是成岩作用的结果。所以, 该层的树枝状体完全是成岩作用形成的, 与微生物无关。也就是说, 树枝状岩石不一定是微生物岩。
1-3~1-8层的斑点状体、网状体、树枝状体一般很少含有熟悉的化石, 但含有丰富的亮晶充填体, 而且分3种不同的情况: 第1种情况是亮晶充填体的轮廓保存较好的第1-7层, 亮晶充填体一般都具有黄色粒状白云石组成的边缘和白色粒状方解石组成的内部; 第2种情况是亮晶充填体轮廓大多数难以辨认的, 如第1-5层和第1-8层。在第1-5层中, 亮晶充填体被较小的块状亮晶方解石充填, 不具有黄色的边缘, 因而与亮晶充填体之间的微亮晶没有明确的界限, 所以轮廓难以确定, 但有一些亮晶充填体的轮廓被保存下来了。在第1-8层中, 树枝状体内部的亮晶充填体的黄色边缘大多数被破坏, 仅有局部残余, 一些相邻的亮晶充填体被同一大块状亮晶方解石充填, 说明溶蚀作用把2个相邻的亮晶充填体之间的泥晶也溶蚀了, 所以, 亮晶充填体的轮廓不完整, 难以识别。第3种情况是, 树枝状体由很大的块状亮晶方解石组成, 看不到亮晶充填体, 只有少数暗色菱形方解石、针状方解石、泥晶残余呈漂浮状存在于块状亮晶方解石之中, 如第1-3层、1-6层, 代表曾经发生了强烈的溶蚀作用, 使大多数亮晶充填体以及它们之间的泥晶溶蚀殆尽。原来亮晶充填体内部的针状方解石和菱形方解石只剩丝缕状残余, 岩石呈蜂窝状, 大块亮晶方解石以连晶方式充填溶蚀孔洞, 一个晶体包含了原来若干个亮晶充填体的空间。这3种情况反映了溶蚀作用强度的不同。
5.1.2 斑点状体、网状体、树枝状体是成岩流体自上而下的渗流造成的
就目前笔者所研究的剖面看, 斑点状和网状的微生物岩出现在微生物岩段的下部, 树枝状的微生物岩出现在微生物岩段的上部。在老龙洞1号剖面上, 斑点状岩石厚度为1.3 m, 位于下段; 树枝状岩石只有0.8 m, 位于上段。在江西修水剖面上, 斑点状微生物岩厚1.5 m, 在下段; 树枝状微生物岩厚0.5 m, 在上段(Wu et al., 2014b)。斑点状体在岩石中的体积含量少于网状体和树枝状体的含量。这种下部含量少、上部含量多的分布, 反映了形成机制的影响力自上而下减弱。一般来说, 自上而下渗透的成岩流体具有这样特点。在所有的剖面上, 树枝状体出现在微生物岩段的顶部或上部, 不出现在中下部。
笔者对此的解释是: 流体向下的运移路径受到重力作用和沉积物结构的影响。重力作用使流体向下运移。沉积物的结构, 主要是原始孔隙的多少, 能够使流体顺着孔隙高的方向运移。原始孔隙主要受沉积物粒度和化石分布控制。由于沉积物都是泥晶, 没有颗粒, 因此沉积物的孔隙主要受化石的分布控制。由于化石一般有体腔孔, 因此化石丰富的地方原始孔隙度就高。笔者认为, 亮晶充填体的前身是某种化石。这种前身化石丰富的地方, 就是原始孔隙度高的地方。如果化石的分布是随机的, 那么, 流体将倾向于沿着化石丰富的地方运移。当流体的数量较大的时候, 重力的影响起主要作用, 流体的运移路径以自上而下的垂向运移为主。当流体的数量较少的时候, 流体的运移路径主要受沉积物原始孔隙控制, 即主要受化石分布控制, 作随机的运移。当流体自上而下运移的时候, 流体的数量必然自上而下减少。所以, 不难理解会出现上部以树枝状运移路径为主, 下部以随机的斑点状或网状运移路径为主的现象。
流体沿着前身化石丰富的地方运移, 对前身化石进行改造, 使其轮廓被破坏, 或完全被破坏, 从而难以识别。笔者观察到, 与微生物岩段上部相比, 亮晶充填体的形态在微生物岩段下部的斑点状体内和斑点状体之间有更好的保存, 而在微生物岩段最上部的树枝状体中保存最差、难以识别, 最上部的树枝状体形态最典型。这些特征都说明了是自上而下的流体作用形成了斑点状体、网状体和树枝状体。
5.2.1 亮晶充填体的前身是某种生物腐烂后留下的孔洞
如果研究者只观察微生物岩段最上部形状典型的树枝状体, 就无法看清其中的亮晶充填体, 只能在局部、特别是在树枝状体之间看到亮晶充填体轮廓的残留。根据这些残留, 研究者很难恢复亮晶充填体的形态和特征。但如果研究者对整个微生物岩段的斑点状体、网状体、树枝状体进行全面地观察, 就会发现: 所有的斑点状体、网状体、树枝状体中的亮晶充填体, 是大小、形状都相似的东西, 但内部充填的物质不一样。微生物岩段最下部斑点状体中的亮晶充填体, 特别是斑点状体之间泥晶中分散保存的亮晶充填体, 形状保存较好, 越往上部, 亮晶充填体保存越差, 树枝状岩石顶部的保存最差。
在1号剖面第1-3层, 即微生物岩的最底部, 亮晶充填体的典型组成是: 边缘有少数尖端指向亮晶充填体中心的针状方解石, 它们被包含在1个或多个大块状的白色亮晶方解石胶结物中(图4-A至4-E)。这种针状的方解石胶结物是在沉积期和准同生期由针状文石转变来的。而针状文石, 在浅海环境中, 或在沉积物孔隙中, 一般是在沉积期和准同生期形成的, 是最早的胶结物。其他环境中均不形成这样的文石胶结物。由于文石矿物的不稳定性, 它们一般在准同生期或者成岩早期就会转变为方解石, 但针状的形态会保留下来。由于针状方解石的粗端直接生长在泥晶之上, 亮晶充填体没有任何生物成因的“ 壁” 或“ 壳” 之类的构造, 所以, 笔者认为针状文石是生长在某种孔洞的壁上的, 即亮晶充填体的前身是一个孔洞。针状文石很可能没有完全充填满原来的孔洞。由于现今亮晶充填体内部都只见到少数针状方解石, 推测发生了溶蚀作用, 使大多数的针状方解石溶蚀掉了。在溶蚀作用之后, 发生了块状方解石的胶结作用, 使孔隙被充填满。
亮晶充填体不是钙化生物。微生物岩段含有少数钙化生物, 主要是一些小腹足类、介形虫类等。它们主要分布在斑点状体、网状体、树枝状体之间的泥晶中, 在1-9层的树枝状体中有密集的分布。无论在泥晶中, 还是在1-9层树枝状体中, 这些生物化石的壳壁都得到较好的保存, 并没有被溶蚀掉。在1-3~1-8层的斑点状体、网状体、树枝状体中很少见到这些种类的化石, 推测要么是这些种类的化石本来就没有, 要么是被成岩流体溶蚀了。但成岩流体却没能够溶蚀掉泥晶中的这些种类的化石, 而泥晶中的亮晶充填体没有壳壁, 所以推测亮晶充填体本来就是没有壁的, 不是钙化生物。在笔者观察的数以千计的亮晶充填体中, 只有3号剖面3-2层的一个薄片局部出现特殊情况, 亮晶充填体表面有不规则分布的泥晶。蓝细菌的钙化壁是泥晶的, 那么3-2层的这些亮晶充填体的泥晶是不是钙化的?首先, 它们厚度不规则; 第二, 它们不是在亮晶充填体表面到处都有, 而是在很多地方不存在, 例如图7-F中黄色虚线包围的那个亮晶充填体, 泥晶在左侧呈疙瘩状分布。Kershaw等(2018)根据与本文图7-E、7-F相似的材料, 提出本文解释的亮晶充填体是整个由泥晶组成的钙化生物。一般来说, 泥晶组成的生物是比较稳定的, 不容易遭受成岩作用影响。这也是为什么很多研究碳同位素的学者选择岩石泥晶组分的原因。泥晶组成的钙化生物在老龙洞1号剖面的1-3层局部很丰富(图4-F), 为丝状蓝细菌, 未受成岩作用改造。把亮晶充填体解释为泥晶钙化生物的做法, 是没有依据的。
由于成岩作用影响的不同, 不同层位、不同地点的亮晶充填体的内部充填物很不相同。根据观察到的所有情况, 亮晶充填体经历的成岩作用依次包括: 针状文石形成、针状文石全部转变为针状方解石、白云石化、去白云石化、溶蚀、大块方解石胶结物形成。针状文石不存在。针状方解石只有少数残留, 一般被包含在大块状方解石中。从未见到亮晶充填体全部为针状方解石的情况, 说明白云石化作用的影响比较普遍, 使大多数亮晶充填体的针状方解石都转变成为白云石; 或者, 即使针状方解石没有全部转变为白云石, 在溶蚀作用中, 几乎全部被溶蚀了, 只剩下少数残留。白云石化作用形成的白云石是半自形到自形的, 粒状, 往往呈黄色。大多数亮晶充填体中的粒状白云石已经转变为粒状方解石, 并且大多数亮晶充填体的粒状方解石被溶蚀了。或者, 粒状白云石没有转变为粒状方解石的情况下, 也在溶蚀阶段大多数被溶蚀了。但有少数亮晶充填体中的粒状白云石被保留下来, 这就是图7-E所示3号剖面3-2层的局部现象。无论是粒状白云石组成的亮晶充填体, 还是粒状白云石发生去白云石化变成粒状方解石的亮晶充填体, 在溶蚀阶段都遭受到不同程度的溶蚀, 形成的溶蚀孔洞在其后的胶结阶段全部被大块状亮晶方解石充填。在一些大块状方解石中有漂浮状的暗色的菱形方解石, 有的还具有环带构造, 而环带构造是交代白云石化作用中形成的, 是白云石的典型特征, 所以漂浮状的菱形方解石是去白云石化作用的关键证据。假定一个非钙化的微生物体, 腐烂后留下孔洞, 那么孔洞经历了针状文石形成、针状文石全部转变为针状方解石、白云石化、去白云石化、溶蚀、大块方解石胶结物形成这6种成岩作用之后, 会形成多少种类型的亮晶充填体?本文列出了8种(图12), 但实际情况可能更多。
 | 图12 由某种生物腐烂形成的孔洞, 经历6种成岩作用形成的亮晶充填体的可能类型的推演Fig.12 Possible types of spar-filled structures, pores filled by calcites and dolomites during 6 successive diagenetic processes |
如1号剖面第1-5层中, 亮晶充填体由外缘的黄色粒状白云石(图5-C: yd)和内部的白色块状方解石(图5-C: c)组成, 有少数针状方解石(图5-C: nc)和他形粒状方解石(图5-B: gc), 属于推演的情况D(图12-D)。在1号剖面1-7层中, 一些亮晶充填体几乎全部由黄色粒状白云石组成(图6-C右下角, s1箭头所指黄色虚线内的), 属于推演的情况F(图12-F); 另外一些亮晶充填体由边缘的黄色粒状白云石和内部的白色粒状方解石组成(图6-C右下角, s2箭头所指的黄色虚线内), 属于推演的情况C(图12-C)。在3号剖面第3-1层中, 亮晶充填体是“ 少数针状方解石+块状方解石” 结构的(图7-D), 属于推演的情况E(图12-E)。在3号剖面的第3-2层, 部分亮晶充填体全部由黄色粒状白云石组成(图7-F), 属于推演的情况F(图12-F)。文中对1号和3号剖面每一层的亮晶充填体的结构进行了详细描述, 这些类型的亮晶充填体绝大多数都可以归入到图12的推演类型之中。如果对P-T界线地层的亮晶充填体缺乏正确认识, 那么正确解释其形成机制几乎是不可能的。
5.2.2 亮晶充填体是某种生物的模铸化石
通过以上的分析, 已经得出这样的认识: 斑点状体、网状体、树枝状体形成之前, 它们所在的地方与它们之间的泥晶沉积物的唯一区别, 是亮晶充填体的数量更加丰富。也就是说, 在斑点状体、网状体、树枝状体形成之前, 沉积物都是由泥晶和亮晶充填体组成的。但亮晶充填体分布不均匀, 有些地方丰富、有些地方稀少。后来成岩流体沿着亮晶充填体丰富的地方运移, 形成粗的白云石和方解石晶体, 导致形成了斑点状体、树枝状体、网状体。
毫无疑问, 亮晶充填体的前身是某种成因的孔洞。但究竟是什么成因的孔洞?首先, 不是原始的粒间孔洞。其次, 不大可能是溶蚀孔洞。这种孔洞, 是在针状方解石、也就是针状文石形成之前就已经存在的孔洞。针状文石是碳酸盐沉积物中最早期的胶结物。所以, 亮晶充填体的前身孔洞是很早期形成的, 即准同生期或早成岩期。由于斑点状体、网状体、树枝状体之间是泥晶, 斑点状体、网状体、树枝状体内部亮晶充填体之间一般也都是泥晶, 尽管泥晶可能重结晶成微亮晶, 或者被溶蚀残留呈漂浮状存在于大块状亮晶方解石中。因此, 没有理由认为亮晶充填体是泥晶中无缘无故的化学溶蚀作用形成的。如果不是有某种丰富的化石, 那么泥晶沉积物是均一的, 溶蚀作用无法形成如此密集而形状不规则的孔洞。亮晶充填体大小0.1~3.3 mm, 如果是纯粹无机溶蚀作用, 范围会更大, 特别是可能形成较大的溶洞。这种大小有一定范围、分布密集的孔洞, 可能是某种微生物体腐烂形成的。目前尚无化学溶蚀作用形成这种特征的孔洞的实例。所以, 笔者推测这类孔洞是某种微生物腐烂留下的。
地质历史时期的生物被保存下来叫化石, 可分为5种: 第1种是实体化石, 如蓝细菌被硅化形成的化石, 细胞结构都能保存下来; 第2种是骨骼化石, 如恐龙化石, 只保存骨骼部分; 珊瑚、贝壳也如此; 第3种是碳质化石, 指植物、动物经过脱水和轻度降解以后被以压扁的有机质保存下来; 第4种是印痕化石, 指生物仅仅在细粒沉积物的层面上留下形状和痕迹; 第5种是模型化石和铸体化石, 前者是如果生物体被沉积物掩埋, 沉积物在生物体腐烂之前半固结或固结, 那么, 生物体腐烂之后留下的孔洞就保留了生物体的形状; 后者是如果模化石后来被某种物质充填, 这个充填的构造就具有的生物体形状。所以, 笔者认为, 亮晶充填体是某种生物腐烂后被各种胶结物充填形成的模铸化石。
5.2.3 亮晶充填体的前身(孔洞)是蓝细菌门色球菌纲微囊菌属的群体
日常所知的生物, 都是有一定形状的, 但变形虫除外。变形虫最大的才200~600 μ m, 而且形态的变化没有亮晶充填体大。所以, 亮晶充填体的前身不可能是变形虫。
日常所知的生物, 都是有一定形状的, 但变形虫除外。变形虫最大的才200~600 μ m, 而且形态的变化没有亮晶充填体大。所以, 亮晶充填体的前身不可能是变形虫。
什么生物的形状是不规则的, 或者说是没有固定形状的?生物界分为5个界。原核生物、原生生物、植物、动物、真菌。后生动物和后生植物都是有固定形状的。原生生物和真菌不具有亮晶充填体这样的形态。所以应当从原核生物中寻找是哪一类生物。笔者文献调研发现, 细菌菌落的形状是不规则的, 于是提出亮晶充填体可能是某种细菌菌落的化石(姜红霞和吴亚生, 2007)。但进一步的思考后认为, 细菌菌落的形态与亮晶充填体的相似性还是不够, 亮晶充填体是细菌菌落的可能性不大。然后笔者检查了所有现生的蓝细菌的属的特征, 发现现生蓝细菌的一个叫微囊菌(Microcystis Kü tzing, 1833)的属, 形态与亮晶充填体有很大的相似性。
微囊菌属于原核生物界蓝细菌门(以前叫蓝藻门)色球菌纲色球菌目(Chroococcales)微囊菌科(Microcystaceae)(Komá rek and Komá rková , 2002; 虞功亮等, 2007), 模式种为Microcystis aeruginosa Kü tzing, 一般呈多细胞的群体出现。群体微小或大型, 自由漂浮, 或附着于水中的各种基质上; 形态为球形、椭圆形、不规则分叶状或长带状, 某些种类为不规则树枝状; 通常由细胞聚集组成或由细胞聚集成亚群体, 再组成群体; 群体外面一般都包裹有一层透明的粘性的胶质, 称胶鞘。现代微囊菌是一种主要生活在淡水中的漂浮生物, 由于细胞中有气囊, 可以根据需要调节浮力, 从而达到上浮和下沉的目的。微囊菌有个独特习性: 当水温大于10 ℃, 就上浮到水面, 繁殖。当水温下降到10 ℃以下, 就下沉到水底(Š ejnohova and Marsalek, 2012)。所以, 春夏秋三季是微囊菌繁盛的时候, 冬季就较少看到它们。当水体富营养化、温度较适宜的时候, 微囊菌在表层水大量繁殖, 把水染绿, 称为水华。微囊菌繁盛会导致水体中有机质含量增加, 其降解会消耗大量氧气, 从而会造成下部水体的贫氧或缺氧。另外有些微囊菌能够分泌毒素, 可以造成水生生物的疾病和死亡。
在类比的过程中, 首先需要确定微囊菌能不能形成模铸化石。根据法国学者(Latour et al., 2007)对Grangent水库沉积物的研究, 其中的微囊菌的胶鞘可以保存14年之久而不腐烂。根据现代碳酸盐沉积学的成果, 在现代热带海洋的海滩上, 碳酸盐沉积物的胶结作用可以在几个月内发生。由于老龙洞剖面在二叠纪— 三叠纪之交属于低纬度地区, 且下伏的长兴组是生物礁, 笔者有理由认为当时老龙洞地区属于热带亚热带浅海地区, 碳酸盐沉积物的早期胶结作用应该可以发生, 所以, 在微囊菌被泥晶沉积物埋藏之后, 泥晶沉积物在微囊菌的胶鞘腐烂之前发生固结或半固结、微囊菌的胶鞘腐烂之后留下的孔洞保留了微囊菌的形状, 是完全可能的。
其次, 比较微囊菌在形态和大小上与亮晶充填体的相似性。现生微囊菌的群体形状以不规则为特征, 而亮晶充填体的形状也以不规则为特征。如何比较2种形状不规则的东西的相似性, 这是个缺乏先例的事情。笔者想到的是, 对现代微囊菌群体和老龙洞剖面的亮晶充填体分别进行形状分析, 找出一些特殊的形状进行比较。 根据发表的文献的照片, 以及笔者对现代微囊菌群体的照相, 发现现代微囊菌群体除了一般的不规则形状之外, 还可以归纳出如下特殊形状(图13), 按照由小到大、由简单到复杂的顺序, 有: (1)近球形, (2)花生形, (3)三叶形, (4)胚胎形, (5)树形, (6)不规则网状。微囊菌群体大小一般在0.1~3 mm。根据对所有薄片的观察, 发现亮晶充填体除了大量不规则形状外, 还有一些特殊的形状(图14), 包括: (1)近球形, (2)花生形, (3)三叶形, (4)胚胎形, (5)简单树形, (6)复杂树形, (7)不规则网状。亮晶充填体的大小从小于0.5 mm 到大于3 mm。通过比较不难发现, 亮晶充填体的大小和形状与现代微囊菌群体有高度相似性。
 | 图13 现代微囊菌群体的形状和大小 现代微囊菌群体的形状包括: 近球形(I)、花生形(H)、三叶形(I中部)、胚胎形(G、J)、简单树形(E)、复杂树形(A、B)、不规则网状(C、D)、圆环形(F)等。群体表面都有1层胶鞘, 在C、D、G、I中都可比较清晰看到胶鞘。J中可以看到群体内部有一些洞。图中A-I标尺为0.1 mm, J中为1 mmFig.13 Colonial forms of extant cyanobacterium Microcystis |
 | 图14 重庆老龙洞1、3号剖面P-T界线地层微生物岩中亮晶充填体的大小和形状 亮晶充填体的形状包括: 近球形(L)、花生形(G、K)、腰果形(I、J)、三叶形(H)、胚胎形(E)、简单树形(F)、复杂树形(B、C、D)、不规则网状(A)等。所有标尺长度均为0.5immFig.14 Forms of the cement-filled structures comprising the patchy and dendroid or net-like structures in the P-T boundary microbialites |
最后, 笔者分析了所有其他现代蓝细菌以及低等绿藻的单体和群体形状, 发现没有任何其他属的单体或群体在形状和大小方面与亮晶充填体有微囊菌这样的相似性。
因此, 亮晶充填体的前身(孔洞)是蓝细菌门色球菌纲微囊菌属的群体。
5.2.4 亮晶充填体的前身生物是漂浮的
如果亮晶充填体的前身生物是底栖的, 会有2种情况: 第1种, 水体较深因而较为安静, 亮晶充填体的前身生物微囊藻在被沉积物掩埋后会保持生长状态, 即生物体具有向上生长的形态, 并且保持向上生长的这种形态, 但是, 笔者在所有的薄片中未见到这样的情形。第2种, 水体较浅, 受到破浪的搅动, 底栖的微囊藻会搅动, 保存在沉积物中应该是各个方向的。但是, 既然是底栖生物, 就应该有向上的生长形态。在具有向上的生长形态的情况下, 即使被搅动、倒置或者躺倒保存的, 仍然应该看得出生长形态, 但这种情况并没有在薄片中见到。
现生微囊藻是漂浮的, 所以, 假定亮晶充填体的前身微囊藻也是漂浮的, 似乎是很自然的事情。假如亮晶充填体的前身微囊藻是漂浮生活的, 那么, 会出现3种情况: 第1种, 如果微囊藻群体密度比较小, 那么它们可以在水体中自由滚动。当温度下降、表层水不适宜它们生活时, 它们会通过调节细胞中的气囊而下降。在下降的过程中, 海水的波动会造成它们的翻滚, 因此它们沉降到海底后, 位置会各不相同, 不会有定向性。第2种, 如果微囊藻群体很密集, 那么由于它们之间可能粘连, 海水的小波动不会造成单个群体的翻滚, 而当温度下降、或者由于表层有太多的泥质沉积物使它们承受不了重量时, 它们会像一个巨型生物体那样整体下降, 沉降到海底之后, 大部分群体保持了原来在表层水的大致生长方向。第3种, 即使微囊藻群体很密集, 当风浪较大时, 它们很难保持固定的生长方向, 在由于温度或者其他原因下沉到海底的过程中, 大多数群体发生了翻滚, 导致在沉积物中保存下来的化石没有统一的生长方向。
现生微囊藻是漂浮生活的, 即使如此, 也有一些呈树形的, 即是有生长方向的。所以, 不能因为有树形的群体, 就认为这种生物一定是底栖生活的。
此外, 附着生活的生物往往有附着的构造, 如假根。即使是单细胞的蓝细菌, 附着生活的形态是有极性的, 一般是附着的一端小, 生长端粗。但笔者在薄片中未看到附着生活的构造或者生物体有极性的证据。
通过以上研究, 笔者确定亮晶充填体与现代微囊菌群体在大小和形状以及生活方式方面非常相似, 所以认为亮晶充填体是当时的微囊菌形成的模铸化石。
Kershaw 等(1999)首次描述了四川华蓥涧水沟P-T界线地层的“ 指状” (digitate)岩石(他们的图5-A, 本文的图15-A), 即后来学者描述的树枝状岩石; 他们认为指状岩石像凝块石, 但由于重结晶, 不能肯定是凝块石成因的。
 | 图15 P-T界线地层文献中的照片、原作者的解释和我们的解释(Ⅰ )Fig.15 Microphotograph of the P-TB microbialites in previous documents, interpretations on dendroid bodies and spar-filled structures by previous authors and us(Ⅰ ) |
Ezaki等(2003)首次明确地把四川华蓥东湾剖面的树枝状岩石归入凝块石, 这一观点后来被很多学者接受。东湾剖面在本文研究的老龙洞剖面的北偏东方向50 km处。东湾剖面微生物岩的露头呈典型的树枝状(Ezaki等(2003)一文中的图5、图6), 与老龙洞剖面的树枝状岩石非常相似。该文中的图8-C(本文的图15-C)是一个薄片照片, 他们把红色虚线(本文所加)内的部分(笔者认为就是本文所说的树枝状体)解释为凝块状的(The clotted portion), 即他们将树枝状体解释为凝块石。他们未对其中的亮晶充填体进行描述, 但笔者看到, 其中有大量轮廓显著的亮晶充填体, 如黄色虚线内的2个。Ezaki等(2003)一文的重要成果之一是描述了小球状钙化化石(他们的图8-G, 本文的图15-D), 但没有进行属种鉴定。笔者后来(Wu et al., 2016)对这种小球状化石进行了专门研究, 鉴定为蓝细菌门色球菌纲的Stanieria属, 并统计了含量, 认为其是很次要的生物, 而且只分布在微生物岩段的上部, 在下部没有出现。通过研究, 笔者确定Stanieria是附着在微囊菌上生活的。
Kershaw 等(2007)描述了四川华蓥百竹园剖面P-T界线地层的微生物岩, 把笔者定义的树枝状体叫凝块石(他们的图3-C, 本文的图15-E)。从其照片看, 树枝状体主要由粒状方解石组成, 树枝状的特征很典型, 去白云石化程度高, 难以分别其中的亮晶充填体, 与本文描述的老龙洞剖面P-T微生物岩段上部的树枝状岩石相似。
刘建波等(2007)描述了贵州罗甸大文剖面P-T界线地层的微生物岩, 他们称为“ 网状凝块石” 的就是本文所说的网状体, 他们称为“ 球状体” 的就是本文所说的亮晶充填体(他们的图4-E, 本文的图15-F)。
Kershaw等(2012)描述了四川华蓥涧水沟剖面P-T界线地层的树枝状微生物岩, 该文中的图6-A、6-B是典型的树枝状微生物岩, 他们解释为钙质微生物生长形成的凝块石。笔者认为, 他们说的凝块石就是本文定义的树枝状体, 其去白云石化作用程度高, 破坏了亮晶充填体的轮廓, 使其内的大多数亮晶充填体无法辨别, 只在边缘有少数亮晶充填体的轮廓可以勉强识别(他们的图6-B, 本文的图15-G)。
以上学者主要关注微生物岩段顶部成岩作用程度高的树枝状岩石。这部分微生物岩因为成岩作用程度高, 现在主要由粒状方解石和微亮晶组成。如本文前面所述, Aitken(1967)、Kennard和James(1986)的凝块石定义以及大家使用的凝块石概念都是: 由微生物的作用形成的、主要由泥晶组成的不均一的沉积体, 其中的泥晶部分叫凝块, 被认为是岩石的骨架。Ezaki等(2003)、刘建波等(2007)、Kershaw等(2007, 2012)把P-T界线地层微生物岩中的斑点状体、树枝状体 、网状体当作凝块石, 但本文的研究表明, 这3类结构体是成岩作用对原始沉积物改造形成的次生结构, 主要由多期成岩作用形成的多种形态的、较粗的方解石和白云石晶体组成, 包括组成亮晶充填体的针状方解石、黄色粒状白云石、白色粒状方解石、暗色菱形方解石、大块状白色方解石和亮晶充填体之间的微亮晶。把这3种结构归入凝块石完全不符合凝块石的原始和通用定义。
姜红霞和吴亚生(2007)确定江西修水剖面P-T界线地层微生物岩中的斑点状体、树枝状体是成岩作用形成的, 其中的不规则囊状体(即本文定义的亮晶充填体)是某种菌落形成的模铸化石。在对重庆北碚老龙洞3个剖面的P-T界线地层微生物岩进行大量研究、特别是与现代蓝细菌等微生物进行对比研究之后, Wu 等(2014a)确定P-T界线地层微生物岩中的亮晶充填体是蓝细菌门微囊菌群体形成的模铸化石。该成果得到同行学者的广泛关注(Luo et al., 2014; Deng et al., 2015; Shen et al., 2015; Yang et al., 2015; Liao et al., 2016; Zhang et al., 2016)。但其后仍然有少数学者持不同观点:
1)Wu等(2017)研究了江西修水清水岩剖面的P-T界线地层微生物岩之后, 将其中的树枝状体(他们的图5-B, 本文的图15-H)归入树形石(dendrolite)。
前文已经介绍, 树形石(dendrolite)是Riding(2000)定义的一种微生物岩类型, 指由厘米级的、灌木状钙化微生物(表附藻、肾形藻等)形成的微生物岩。本文的研究表明, 树枝状体是多期成岩作用形成的结构, 其中除了在下部一个层位含有丝状蓝细菌、上部含有数量很少的小球状化石之外, 不含其他钙化化石, 更不含灌木状钙化微生物。虽然1号剖面第1-9层的生物很丰富, 但都是腹足类、介形虫类等。灌木状微生物指的是完全由泥晶组成的真正的钙化生物, 而P-T界线地层微生物岩中的树枝状体, 仅仅形状像树枝状, 实际上不是灌木状生物; 而且树枝状岩石仅仅发育在微生物岩段的上部, 中下部都是斑点状的和网状的, 而树枝状、斑点状、网状的岩石内部的结构和成因都是一样的, 都是由不同期成岩作用形成的多种类型的较粗的方解石、白云石晶体和微亮晶组成的, 特别是晚期形成的大块状方解石胶结物对斑点状、网状、树枝状特征的形成起了关键作用。所以, 将树枝状体归入树形石是难以理解的。笔者注意到, Wu等(2017)一文中图5-B中的树枝状体, 尽管因为去白云石化作用的破坏造成亮晶充填体难以辨认, 但还是能识别出一些(本文图15-H: 黄色虚线内)。
2)Tang等(2017)描述了四川鱼洞子剖面P-T界线地层的微生物岩, 包括3部分: 下段厚3.1 m, 露头上呈斑点状(该文中的图5-B); 中部厚1.3 m, 露头上呈树枝状(该文中的图7-B); 上段厚6.8 m, 露头上呈网状(该文中的图9-B)。他们把斑点状体、树枝状体、网状体都描述为凝块。虽然照片质量不好, 笔者仍然可看到斑点状体(他们的图6-C, 本文的图15-I: 红色虚线内), 且斑点状体内亮晶充填体的轮廓几乎全部被成岩作用破坏了, 而大块状方解石(他们的图6-C, 本文的图15-I: c)应该是表生成岩期之后充填溶蚀孔洞的最后一期胶结物; 斑点状体之间的泥晶中有一些介形虫(他们的图6-C, 本文的图15-I: o)和轮廓保存较好的亮晶充填体(他们的图6-C, 本文的图15-I: 如黄色虚线内的2个)。他们的树枝状和网状微生物岩的薄片照片质量不好, 笔者难以做出解释。
Kershaw(2017)提出P-T界线地层微生物岩类型具有古地理分异: 以中国华南为代表的东特提斯地区, 以钙质微生物组成的微生物岩为主; 以土耳其为代表的西特提斯地区, 以叠层石等的发育为特征。Kershaw(2017)一文中图1右边的照片(本文图15-J, 黄色虚线系本文加), 被用来说明华南钙质微生物型微生物岩。显然, 黄色虚线内的结构体被当成钙质微生物。
之后, Kershaw等(2018)对重庆北碚老龙洞剖面的P-T界线地层微生物岩(他们的3-A; 本文的图15-I, 黄色虚线系本文所加)和四川华蓥百竹园剖面的P-T界线地层微生物岩(他们的图4-A)进行重新解释。笔者注意到, 这2个照片中有很多轮廓明显的亮晶充填体(本文的图15-J: 黄色虚线内), 但都被他们解释为泥晶组成的钙质微生物(calcimicrobe)。
如上文所述, 如果是泥晶的钙化生物, 其应该大量被保存下来。因为1号剖面第1-3层的钙化蓝细菌都被很好地保存下来。笔者对数以千计的亮晶充填体进行观察, 没有发现1个有泥晶壁的。Kershaw 等(2018)解释为钙质微生物的亮晶充填体(本文的图15-I, 15-J)内部, 根本不是泥晶质的, 而是粒状的白云石和方解石。同样的亮晶充填体, 见于老龙洞3号剖面第3-2层(本文的图7-E)。这些亮晶充填体, 是笔者观察研究的数以千计的亮晶充填体中的少数特例: 亮晶充填体内部的粒状白云石基本全部被保存下来, 而且一些亮晶充填体的表面有附着的泥晶斑点。既然这些附着的泥晶斑点被保存下来了, 那么, 如果亮晶充填体原来是泥晶组成的钙质微生物的话, 它们泥晶的钙质壁为什么没有被保存下来?钙质微生物应当有一定结构的壁或者壳, 应当有一定的形态构造。但根据笔者对研究剖面微生物岩段数以千计的亮晶充填体的观察, 发现它们都没有壁或壳, 而且没有一定的形态构造, 所以根本不是钙质微生物。
笔者认为, Karshaw(2017)一文中图1的右上照片(本文的图15-J: 黄色虚线内)中, 实际上都是保存很好的亮晶充填体, 其内部边缘的黄色粒状晶体是白云石, 内部中央的浅色粒状晶体应该是方解石, 即属于“ 黄色粒状白云石+白色粒状方解石” 型的亮晶充填体。Karshaw等(2018)一文中的图4-A, 可能是与Kershaw(2017)同一薄片的照片, 其中, 既有全部由黄色粒状白云石组成的亮晶充填体(图15-K: 紫色虚线内多个亮晶充填体), 又有边缘是黄色粒状白云石、内部是白色粒状方解石的亮晶充填体(图15-K: 黄色虚线内), 还有边缘是黄色粒状白云石(图15-L: yd)、内部是大块状方解石(图15-L: c)、还含有针状方解石(图15-L: nc)的亮晶充填体。所以这是与老龙洞1号剖面第1-5层的“ 黄色粒状白云石+针状方解石+大块状方解石” 型亮晶充填体一样的结构体。
Tang等(2017)、Wu等(2017)和Kershaw等(2018)的解释失误, 不仅说明P-T界线地层微生物岩组成和结构的复杂性、研究的艰巨性, 还说明微生物岩的研究任重道远。在今后的微生物岩研究中, 需要提高观察的细致性和全面性, 结合多学科的知识, 进行综合分析。只有这样, 才能透过复杂的现象看到本质, 得出正确的认识。
尽管笔者(Wu et al., 2014a)根据薄片观察的现象, 认为P-T界线地层微生物岩亮晶充填体的前身微囊藻是漂浮生活的, Kershaw 等(2018)仍然坚持认为, 形成这种微生物岩的微生物是底栖生活的。其列举的理由包括: (1)它们形成生物层; (2)形成向上生长的分枝状岩石骨架, 骨架之间有孔洞, 孔洞之中有示顶底构造; 漂浮生活不可能形成向上生长的骨架。
众所周知, 生物层是原地钙化底栖生物形成的。首先, 由于P-T界线地层的微生物岩不具有原地钙化生物, 不是原地钙化生物形成的, 所以, 不是生物层。Kershaw 等(2018)不能用这套岩石是生物层来作为它们是底栖生物形成的理由, 否则就犯了因果倒置的逻辑错误; 第二, 这套岩石不存在分枝状底栖生物形成的骨架。诚然, 微生物岩段上部的树枝状体像某种底栖生物, 但本文已经详细阐述了, 它们是成岩作用形成的结构, 在成岩作用将它们改造成树枝状体之前, 它们与其间的泥晶沉积没有多大区别, 唯一区别是它们之中有更多的亮晶充填体。任何人只要观察一下1号剖面第1-9层的露头和薄片(图7-A, 7-B), 就一定会理解树枝状体的形成与微生物无关, 主要是一种成岩现象。地球科学研究, 不能因为某某像什么就归入什么, 而要通过研究, 确定确实是什么才能归入什么, 否则就是简单化的做法, 而不是科学研究。
Wu等(2017)认为江西修水P-T界线地层微生物岩中钙质微生物很丰富, 对微生物岩的形成具有重要作用, 但他们未阐述还有什么其他因素对微生物岩的形成起重要作用。他们的微生物岩形成模式图(该文中的图10-C)中, 微生物被画成树形的, 但他们文章中描述的钙质微生物化石只包括同心层状的和小球状的。同心层状的微生物化石, 以前已有学者报道(王永标等, 2005, 该文中的图5), 不仅个体小, 而且数量很少。在老龙洞1、2号剖面的大量薄片中, 同心层状微生物化石数量很少, 说它们是形成微生物岩的主要生物或主要因素是不符合实际的。小球状微生物化石在老龙洞剖面中的含量不到10%, Wu等(2016)对之做了专门研究。目前, 尚没有任何学者在P-T界线地层微生物岩中发现树枝状的钙质微生物化石, 所以Wu等(2017)提出树枝状钙质微生物是P-T界线地层微生物岩的主要形成者这一观点是缺乏依据的。
笔者认为, Wu等(2017)一文中的图5-B(本文的图15-H)中的浅色部分就是与本文描述的老龙洞剖面的树枝状体一样的结构。尽管因为去白云石化作用的破坏, 其中的亮晶充填体难以辨认, 但还是能识别出一些(图15-H: 黄色虚线内)。正如姜红霞和吴亚生(2007)描述的一样, 江西修水剖面0.5im厚的树枝状微生物岩也是主要由树枝状体组成的, 树枝状体主要由亮晶充填体组成, 也就是由蓝细菌微囊菌的模铸化石组成的, 但模铸化石大多数已经被成岩作用破坏了。
P-T界线地层中含有钙化微生物化石, 但是数量少。主要有3种钙化微生物化石: (1)丝状蓝细菌, 见于1号剖面的第1-3层中部(图4-E, 4-F); 一些学者把它们解释为有孔虫, 但是, 它们的弯曲表明生物体活着的时候具有一定的柔性, 而这不是有孔虫的特征; 另外, 有孔虫有房室结构, 第1-3层的丝状化石无房室结构; 再者, 有孔虫的壁比较复杂和精致, 而钙化蓝细菌的壁有一定程度的不规则, 特别是钙化壁的表面不像有孔虫的壁表面那样清晰和规则; (2)小球状钙化化石, 出现在微生物岩段的中上部, 下部没有, 含量为0~10%; 笔者已经进行了专门的系统古生物学研究, 将它们鉴定为蓝细菌门色球菌纲的Stanieria(Wu et al., 2016); (3)具同心状房室的某种钙化微生物化石, 已经有学者进行了鉴定。以上3种钙质微生物化石都没有被成岩作用破坏, 形态和结构保存很好。所以, 没有理由假定存在一种未保存为化石的钙化微生物, 并用它来解释这套微生物岩的成因。
1)微生物岩指主要由微生物作用形成的沉积体或者主要由微生物的骨骼组成的沉积体, 包括叠层石、凝块石、纹层石、核形石、均一石、微生物骨架岩、微生物粘结岩、模铸岩、微生物颗粒灰岩、微生物泥粒灰岩等。凝块石指底栖微生物群落吸附或挡积泥晶沉积物形成的内部不均一的碳酸盐沉积体。
2)P-T界线地层微生物岩, 指具有斑点状、网状、树枝状构造的一套岩石。绝大多数的斑点状、网状、树枝状构造, 由亮晶充填体和其间的微亮晶组成。亮晶充填体是指某种生物成因的孔洞, 被准同生和成岩期形成的一种或多种矿物充填形成的结构。
3)亮晶充填体形状不规则, 没有壁, 所含的矿物包括: (1)针状方解石, 可能由针状文石转变来, 是最早期的胶结物; (2)黄色粒状白云石, 形成于针状方解石之后; (3)白色粒状方解石, 可能是粒状白云石去白云石化形成的; (4)大块状方解石。成岩作用类型除了包括形成以上矿物的事件之外, 还包括溶蚀作用。去白云石化作用和溶蚀作用可能都发生在表生成岩期, 可能是由成因上有联系的淡水造成的。
4)通过与所有现生微生物属种的比较, 发现蓝细菌门色球菌纲微囊菌属的群体在形态和大小上与亮晶充填体一致, 并且这种一致性不在任何其他微生物属种中存在。另外, 微囊菌群体的胶鞘能够在沉积物中保存十多年的特性使之具有保存为模铸化石的可能性。所以, 笔者再次确认亮晶充填体是微囊菌群体的模铸化石。微囊菌是一种漂浮蓝细菌, 该群体沉入海底被泥晶沉积物掩埋, 在沉积物半固结或固结之后, 其胶鞘才降解消失, 留下的孔洞被后来的多期矿物充填, 最终形成现今所见的亮晶充填体。
5)亮晶充填体没有定向性, 所以没有证据表明其前身微生物是底栖或附着的。亮晶充填体没有壁, 不具有任何钙化部分, 所以不属于钙质微生物。微生物岩中有少数小球状钙化蓝细菌化石, 但其含量少, 不是在整个微生物岩段都有分布。同心层状钙质微生物含量很少。所以, 钙化微生物不能作为微生物岩的形成机制。
6)斑点状、树枝状、网状结构是成岩流体对沉积物进行改造形成较粗晶体造成的现象, 形成过程没有微生物直接参与, 也不是泥晶成分的, 所以不应该叫凝块石。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|