

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
生物碎屑灰岩差异成岩及储集层特征:以伊拉克HF油田白垩系Mishirif组为例*
[余义常1  , 宋新民
, 宋新民1 , 郭睿1 , 高兴军1 , 林敏捷1 , 衣丽萍1 , 韩海英1 , 陈军2 , 邓亚1 , 李峰峰1 , 刘航宇3 ]
, 宋新民, 林敏捷|
|

第一作者简介 余义常,男,1991年生,中国石油勘探开发研究院博士研究生,主要研究方向为碳酸盐岩开发地质。E-mail: yuyichang@petrochina.com.cn。
中东地区白垩系 Mishirif组以生物碎屑灰岩为主,其形成于温暖潮湿的环境中。综合利用岩心、铸体薄片、全岩分析、常规物性及高压压汞等资料,以伊拉克 HF油田 Mishrif组为例,开展生物碎屑类型、分布特征、差异成岩及储集层特征等研究。 Mishrif组灰岩中生物碎屑以底栖有孔虫、非固着类双壳类、厚壳蛤和棘皮动物为主,含少量苔藓动物、藻类与海绵动物,其含量、类型及大小对沉积环境有重要指示意义。沉积环境决定岩石组分与结构的差异,在此基础上成岩作用控制岩石的孔隙结构与物性特征。生物碎屑主要经历了不同程度海水环境的泥晶化和生物钻孔、大气淡水环境的溶蚀和胶结、埋藏环境的压实压溶和颗粒破裂作用。以底栖有孔虫和非固着类双壳类碎屑为主的低能沉积环境具有“弱溶蚀、强胶结、强压实”的成岩特征,主要发育微孔、晶间孔及粒内孔,孔喉分布呈偏细态细微喉单峰型,物性较差;以厚壳蛤和棘皮动物碎屑为主的高能沉积环境具有“强溶蚀、弱胶结”的成岩特征,主要发育铸模孔、粒间孔及粒间溶孔,孔喉分布呈偏粗态中粗喉极宽峰型,是 Mishrif组最有利储集层。以 HF油田 Mishrif组为代表的白垩系生物碎屑灰岩在中东地区发育广泛,故上述成果对于该地区生物碎屑灰岩油气开发具有重要意义。
, Song Xin-Min, Lin Min-JieAbout the first author Yu Yi-Chang,born in 1991,is a Ph.D. candidate of Research Institute of Petroleum Exploration & Development,PetroChina. He is mainly engaged in research of carbonate development geology. E-mail: yuyichang@petrochina.com.cn.
The Cretaceous Mishrif Formation in the Middle East was developed in a warm and humid environment,mainly consisting of bio-clastic limestone deposits. Based on core observations,cast thin sections,whole rock analysis,petrophysical properties,and high-pressure mercury injection tests,bio-clastic types,distribution,composition,differential diagenesis and reservoir characteristics of the Mishrif Formation in HF Oilfield were systematically studied to clarify the relationship between bio-clastic and intrinsic heterogeneity. The Mishrif Formation is mainly composed of benthic foraminifera,non-fixed bivalves,thick crustaceans,and echinoderms,containing a small amount of bryozoans,algae and sponge animals. The distribution characteristics of biological debris are consistent with the living environment. The content,type,composition and size of biological debris are important signs of the depositional environment. The original composition of these bio-clastics includes unstable aragonite,high-magnesium calcite, and relatively stable medium-low magnesium calcite,which mainly experienced various degrees of soft decay,micritization, and biological drilling in the seawater environment,dissolution and cementation in the atmospheric fresh water environment,compaction pressure dissolution and particle disruption in the buried environment. The differences in sedimentary environment,particle size,morphological characteristics,and composition of various types of bio-clastics have led to differential diagenesis,which controlled pore type,physical properties,and pore-throat structure of bio-clastic limestone,forming a complex pore-penetration relationship. As the content of bio-debris increases,the physical properties of the reservoir become better.The low-energy depositional environments mainly developed benthic foraminifera and non-fixed bivalve detritus are characterized by weak dissolution and strong cementation. Micropores,intercrystalline pores, and intragranular pores were mainly developed. The physical properties are poor,and pore throat distribution shows a fine micro-throat single peak pattern. The high energy sedimentary environment with rudist and echinoderms debris is characterized by strong dissolution and weak cementation. The main development of mold pores,intergranular pores, and intergranular dissolution pores with good physical properties,pore throat distribution shows a very broad range mainly with coarse pore throat,which is the most favorable reservoir type in the study area. The Cretaceous bio-clastic limestone,represented by the Mishrif Formation in HF Oilfield,was widely developed in the Middle East. The above study is of great significance to the development of bio-clastic limestone oil and gas in the Middle East.
中东地区探明可采油气储量占全球探明可采油气储量的43%, 石油产量约占全世界产量的2/3, 且80%的产层位于碳酸盐岩之中(范嘉松, 2005; 张宁宁等, 2014; 马永生等, 2017)。仅就该地区碳酸盐岩油气田的个数而言, 白垩系发育最多, 占到碳酸盐岩大油气田的 29%(Kauffman and Johnson, 1988; 刘政等, 2011)。白垩系 Mishrif组生物碎屑灰岩是伊拉克HF油田乃至中东地区的主力产层, 而生物碎屑主要为白垩纪中晚期大量存在的有壳类碎屑(田泽普等, 2016; 王昱翔等, 2017)。
随着中国在中东地区的油气合作项目日益增多, 亟需对该类生物碎屑灰岩进行系统的认识。前人对于该地区生物碎屑灰岩的研究或侧重于生物的生态、习性特征及壳体成分(Fursich and Oschmann, 1993; 苟宗海, 1996; Carannante et al., 1998; Sadooni, 1998), 或集中于储集层分类及主控因素等(Alsharhan and Nairan, 1997; 邓虎成等, 2014; 郭睿等, 2014; 孙晓伟等, 2017), 缺乏对各类生物碎屑在不同沉积环境及成岩演化过程中的差异性分析, 对生物碎屑与储集层发育的内在联系研究亦不够深入, 限制了对该类储集层的认识及高效开发。文中基于岩心观察、铸体薄片、全岩分析、常规物性及高压压汞测试等资料, 开展伊拉克HF油田Mishrif组生物碎屑灰岩的生物碎屑类型、分布特点、差异成岩演化及储集层特征等研究, 以期加深对生物碎屑灰岩的认识, 为中东碳酸盐岩储集层的勘探开发提供地质依据。
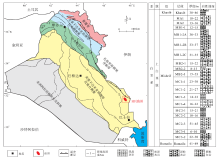
HF油田位于伊拉克东南部米桑省内, 距离首都巴格达约400ikm, 是以生物碎屑灰岩为主的巨型油田。构造上位于美索不达米亚盆地南部的前渊带内, 整体呈北西— 南东向的宽、缓长轴背斜(Aqrawi, 1998)(图 1)。自寒武纪以来, HF油田长期处于古冈瓦纳大陆的北缘, 主要发育地台型沉积。HF油田白垩纪构造活动总体较弱, 处于温暖湿润的低纬度地区, 阳光充足, 含氧量高, 海水循环好, 适宜生物生长(Davies et al., 2002), 有利于形成大规模的生物碎屑灰岩。Mishrif 组是HF油田主力产层, 为森诺曼期海退沉积的生物碎屑灰岩, 其与下伏Rumaila组泥晶灰岩呈整合接触, 与上覆Khasib组呈不整合接触(Aqrawi et al., 2010), 厚度达 400im, 可进一步划分为18个小层。
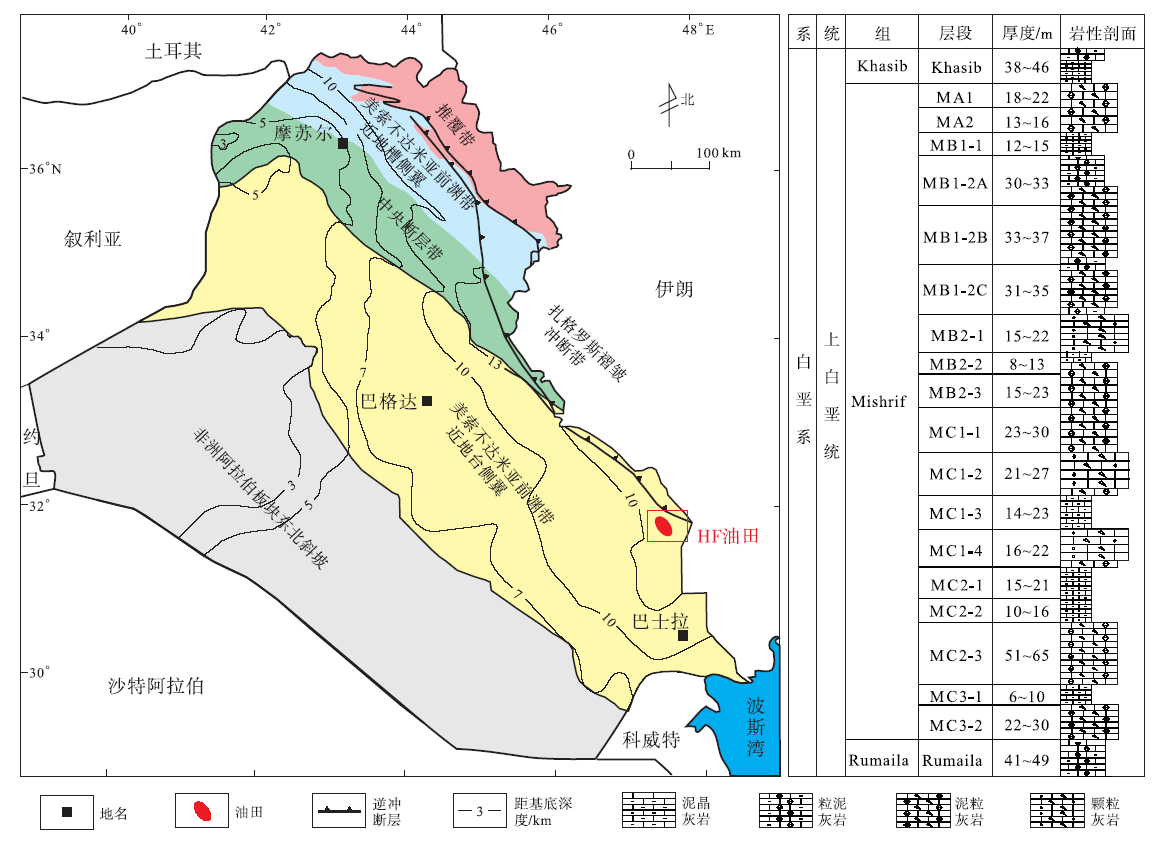 | 图 1 伊拉克HF油田构造位置和Mishrif组地层柱状图Fig.1 Tectonic location and stratigraphic column of the Mishrif Formation in HF Oilfield of Iraq |
Mishrif组的沉积演化受控于Amara古突起的抬升和整个地区海平面的下降(Gaddo, 1971), 前人将HF油田Mishrif组划分为局限台地、开阔台地、台地边缘及斜坡等4个相带(Mahdi et al., 2013, 2014), 并根据沉积特征, 将局限台地细分为灰坪、台内滩与滩间, 开阔台地分为台内滩、滩间和台内洼地, 而储集层集中分布于台地边缘滩和台内滩(王君等, 2016; 王昱翔等, 2017)。目前, HF油田有12口井在Mishrif组油层段取心, 以 M316系统取心井资料最多, 分析化验资料最全。
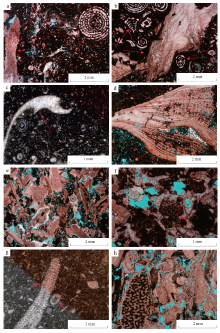
基于12口岩心井的1338块铸体薄片观察, HF油田Mishrif组生物类型丰富, 主要包括底栖有孔虫、非固着类双壳类、厚壳蛤和棘皮动物, 并含有少量苔藓动物、藻类和海绵动物。其独特的外部形态、内部结构及粒径大小是识别生物碎屑类型最重要的标志(表 1)。底栖有孔虫为原生动物, 在低能环境中较多, 主要包括蜂巢虫、栗孔虫、单列有孔虫及双列有孔虫, 外壳常出现比灰泥颜色更深的泥晶套(图 2-a, 2-b)。
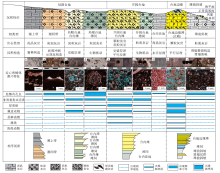
| 表 1 伊拉克HF油田Mishrif 组各类生物的生态特征、沉积相带及碎屑镜下识别标志 Table1 Ecological characteristics of various types of organisms, sedimentary facies and debris identification marks in the Mishrif Formation, HF Oilfield of Iraq |
 | 图 2 伊拉克HF油田Mishrif组各类生物碎屑的镜下特征 a— M316井, 2888.13im, 具有圈层辐射状房室形态的蜂巢虫及具生长条纹的厚壳蛤碎屑, 单偏光; b— M316井, 2894.11im, 盘旋状房室的栗孔虫、对称状房室的双列有孔虫及具生长条纹的厚壳蛤碎屑, 单偏光; c— N137井, 3133.32im, 具平滑弯曲及向铰合部位壳体变厚特征的非固着类双壳类碎屑, 单偏光; d— M316井, 2934.13im, 具有原始生物壳壁生长条纹及隔壁分开的有序多孔构造的厚壳蛤碎屑, 单偏光; e— M316井, 3059.13im, 见类似微裂缝的节理、等轴粒状的具有深色生屑与浅色圈层的棘皮动物碎屑, 单偏光; f— M316井, 3057.82im, 外形呈网状, 保存着具椭圆状泡孔分隔的虫室的苔藓动物碎屑, 单偏光; g— N137井, 3060.11im, 大致呈串珠柱状体, 藻类内部可见排列整齐植物细胞的藻类与棘皮动物碎屑, 单偏光; h— M316井, 2987.13im, 外形呈近椭圆状, 内部具暗色脑纹状骨架的海绵动物碎屑, 单偏光Fig.2 Microscopic characteristics of various types of biological debris in the Mishrif Formation, HF Oilfield of Iraq |
非固着类双壳类动物在水动力较弱及中等的局限台地和开阔台地中出现较多, 具有平滑弯曲的外形和向铰合部位变厚的壳体(图 2-c)。厚壳蛤存在于波浪淘洗作用较强的台地边缘部位(Kauffman and Johnson, 1988; Pascualcebrian et al., 2016), 其壳壁厚度大, 呈现深浅相间的生长纹, 部分可见到排列有序的多孔状构造(图 2-a, 2-b, 2-d)。棘皮动物生活在与广海连通性好的开阔台地和台地边缘环境中(Bryan et al., 1997), 横切面上有深色的轴腔和浅色的骨板(图 2-e, 2-g)。苔藓动物内部有多个横切面为圆形或椭圆形的虫室(图 2-f), 藻类大致呈略弯的串珠柱状体, 内部可见排列整齐的深色椭圆形植物细胞(图 2-g), 海绵动物内部发育不规则的暗色脑纹状骨架(图 2-h), 这3类生物多存在于开阔台地和台地边缘环境中。
生物碎屑原始成分亦存在较大差异。底栖有孔虫的壳体为中高镁方解石, 房室内部为有机软体; 非固着类双壳类为文石质; 厚壳蛤壳体为混合矿物组分, 壳壁及窗格的隔壁为低镁方解石, 多孔状的窗格处为高镁方解石(Schulze et al., 2004; Jacob et al., 2008), 棘皮动物为中— 低镁方解石, 整体稳定性较好; 苔藓动物的壳体为低镁方解石, 虫室内部为有机软体; 藻类的藻体为文石质, 植物细胞内为同沉积期的泥晶方解石; 海绵动物整体为低镁方解石(肖勒和厄尔默— 肖勒, 2010; Fö llmi, 2012)。
与陆源碎屑岩相比, 生物碎屑灰岩的形成地点与最终沉积地点较为邻近, 它们含有沉积时期的生态学信息, 即沉积环境决定了生物种类、含量、颗粒大小、形态特征、成分甚至分布方式(肖勒和厄尔默— 肖勒, 2010; 陈洪德等, 2014; 田泽普等, 2016; 文龙等, 2017)。HF油田Mishrif 组不同沉积环境的生物碎屑差异较大:灰坪和斜坡主要为底栖有孔虫及非固着类双壳类碎屑; 潟湖和滩间靠近台内滩与台地边缘存在少量厚壳蛤和棘皮动物碎屑, 其中厚壳蛤破碎程度低, 粒径相对较大; 台内滩中非固着类双壳类和棘皮动物碎屑占主要部分; 台地边缘滩则以厚壳蛤、棘皮动物碎屑为主, 生物碎屑破碎程度较高, 粒径相对较小(图 3)。
 | 图 3 伊拉克HF油田Mishrif 组沉积相类型及生物碎屑分布 a— M316井, 2879.6~2879.8im, 具窗格构造的潮上带泥晶灰岩; b— M316井, 2888.1~2888.35im, 具示顶底构造的潮间带粒泥灰岩, 可见不规则的底冲刷界面; c— M316井, 2927.7~2927.9im, 具低角度交错层理的局限台地台内滩泥粒灰岩; d— M316井, 2924.4~2924.65im, 具生物扰动构造的局限台地滩间粒泥灰岩, 可见粒径较大的底栖有孔虫碎屑; e— M316井, 3062.2~3062.4im, 具平行层理的开阔台地台内滩颗粒灰岩; f— N137井, 3033.4~3033.6im, 具生物扰动构造的开阔台地滩间粒泥灰岩; g— Y115井, 3067.4~3067.55im, 具水平层理的台内洼地泥晶灰岩; h— M316井, 2983.4~2983.6im, 具平行层理的台地边缘滩颗粒灰岩; i— M316井, 3070.2~3070.5im, 具滑塌撕裂构造的滩前斜坡粒泥灰岩, 可见外形不规则的滑塌角砾Fig.3 Sedimentary facies and contents of bio-clastic in the Mishrif Formation, HF Oilfield of Iraq |
HF油田Mishrif组中泥晶灰岩、粒泥灰岩、泥粒灰岩和颗粒灰岩均有分布, 其中粒泥灰岩和泥粒灰岩约占总厚度的65%。
泥晶灰岩形成于水动力条件弱的沉积环境中, 主要发育在局限台地潮上带和开阔台地台内洼地环境。泥晶灰岩中颗粒含量低于10%, 主要为底栖有孔虫碎屑, 即有孔虫泥晶灰岩(图 4-a)。发育灰白色的斑状生物扰动构造, 铸体薄片中可见毫米级大小、亮晶方解石充填的窗格构造(图 3-a), 这种构造主要是潮上带的碳酸盐沉积物干燥收缩形成的, 是良好的环境标志。台地洼地内的有孔虫泥晶灰岩则发育典型的水平层理, 镜下可见较为明显的白云石化现象(图 3-g)。
 | 图 4 伊拉克HF油田Mishrif 组岩石类型 a— N195井, 3001.4~3001.6im, 具水平层理的泥晶灰岩; b— N195井, 3034.5~3034.7im, 具生物扰动的粒泥灰岩; c— Y161井, 2934.4~2934.6im, 具低角度交错层理的泥粒灰岩; d— Y161井, 3002.5~3002.7im, 具平行层理的颗粒灰岩Fig.4 Rock types in the Mishrif Formation, HF Oilfield of Iraq |
粒泥灰岩形成于水动力条件较弱的沉积环境中, 主要发育在潮间带、滩间和滩前斜坡环境。粒泥灰岩中颗粒含量为10%~50%, 底栖有孔虫和非固着类双壳类碎屑占主要部分, 即有孔虫— 双壳类粒泥灰岩(图 4-b)。发育大量的生物扰动构造(图 3-d, 3-f), 铸体薄片中可见示顶底构造, 孔隙下部为沉积阶段暗色的灰泥, 上部为成岩阶段浅色的亮晶方解石, 两者界面平直(图 3-b)。滩前斜坡存在滑塌角砾和滑塌撕裂构造, 可见滑塌产生的次棱角状泥晶角砾(图 3-i)。
泥粒灰岩形成于水动力条件较强的沉积环境中, 主要发育在局限台地台内滩和开阔台地台内滩环境。泥粒灰岩中生物碎屑的含量为50%~80%, 主要是双壳类和棘皮动物碎屑, 含有少量苔藓动物和藻类, 即双壳类— 棘屑泥粒灰岩(图 4-c)。其粒间具有较多灰泥, 生物碎屑的排列具有一定的方向性, 发育交错层理和平行层理(图 3-c)。其中局限台地台内滩的泥粒灰岩横向和纵向规模较小, 泥粒灰岩和粒泥灰岩频繁互层分布, 而开阔台地台内滩的泥粒灰岩横向和纵向规模较大, 底部与粒泥灰岩互层, 顶部逐渐演变为颗粒灰岩(图 3)。
颗粒灰岩形成于水动力条件强的沉积环境中, 主要发育在台地边缘滩中, 开阔台地台内滩也有所分布。颗粒灰岩中的颗粒含量大于80%(图 4-d), 主要是棘皮动物和厚壳蛤碎屑, 还可见到少量海绵动物碎屑(图 3-e, 3-h)。依据主要生物碎屑的类型, 将其分为棘屑颗粒灰岩和厚壳蛤颗粒灰岩。其中厚壳蛤颗粒灰岩与厚壳蛤礁生长、破碎相关, 礁滩均发育在强水动力条件的台地边缘古地貌高部位。颗粒灰岩中平行层理较为发育, 平行层理除通过粒度变化显现外, 还可由长条形的厚壳蛤碎屑定向排列所致。
421块全岩分析表明, Mishrif 组生物碎屑灰岩的成分简单, 方解石占绝对优势, 白云石含量低, 不同岩石类型之间成分差异较小(表 2)。其含有少量石英和黏土矿物, 不含石膏、硬石膏及盐岩, 反映Mishrif 组形成于温暖湿润的气候环境下, 且受陆源碎屑的影响较小。
| 表 2 伊拉克HF油田Mishrif 组不同类型生物碎屑灰岩全岩分析和X衍射统计 Table2 Whole rock analysis and X-ray diffraction statistics of bio-clastic limestones of different rock types in the Mishrif Formation, HF Oilfield of Iraq |
结合1338块铸体薄片综合分析, Mishrif组生物碎屑灰岩沉积后至埋藏期经历了多种成岩作用, 建设性成岩作用包括生物钻孔、大气淡水溶蚀和颗粒破碎作用, 破坏性成岩作用包括泥晶化、胶结和压实压溶作用。其中, 大气淡水溶蚀、胶结、压实和颗粒破碎、压溶作用对生物碎屑影响较大。
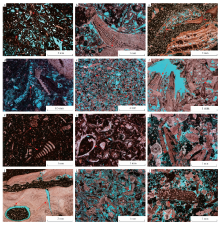
大气淡水溶蚀是储集层关键的建设性成岩作用, 包括选择性溶蚀和非选择性溶蚀作用。选择性溶蚀作用发生的动力学机制是溶解度的差异(肖勒和厄尔默— 肖勒, 2010), 当不稳定文石和高镁方解石质的生物碎屑和稳定的中— 低镁方解石质的生物碎屑共存时, 不稳定的生物碎屑优先溶蚀(图 5-a至5-d)。非选择性溶蚀发生于矿物稳定化之后, 大气淡水溶蚀多不具有选择性, 使得原有孔隙溶蚀扩大, 产生粒间溶孔和溶蚀溶洞(图 5-e, 5-f)。溶蚀产生的碳酸钙在其他地方沉淀, 多于淡水潜流带产生等轴粒状胶结物(图 5-g, 5-h)和共轴增生胶结物(图 5-i), 充填于体腔孔、铸模孔, 或赋存于颗粒周围; 碳氧同位素分析和全岩测定结果显示, 其为淡水— 海水混合成因, 与原始孔隙被大量淡水饱和有关(王昱翔等, 2017)。压实作用是指在上覆地层压力下生物碎屑发生破碎、折断、紧密压实等, 颗粒呈点— 线接触或线接触(图 5-k, 5-l); 随着地层压力的增加, 颗粒接触处局部溶解, 发生压溶作用(图 5-b, 5-i)。
 | 图 5 伊拉克HF油田Mishrif 组成岩作用特征 a— M316井, 3028.37im, 非固着类双壳类和藻类的选择性溶蚀, 棘皮动物未发生溶蚀; b— M316井, 3063.12im, 非固着类双壳类和藻类的选择性溶蚀, 厚壳蛤的生物钻孔和压溶作用; c— M316井, 2915.22im, 厚壳蛤多孔状窗格处发生选择性溶蚀, 壳体和隔壁位置较完好; d— M316井, 3014.15im, 藻类外壳选择性溶蚀, 留下难溶的内部结构; e— M316井, 2984.11im, 非选择性溶蚀形成粒间溶孔, 底栖有孔虫形成较薄的泥晶套; f— M316井, 2989.12im, 厚壳蛤非选择性溶蚀形成溶蚀孔洞; g— M316井, 2900.14im, 小栗虫和单列有孔虫房室内发生等轴粒状胶结, 外壳泥晶化; h— M316井, 2891.11im, 非固着类双壳类发生溶蚀作用后被等轴粒状胶结物充填; i— M316井, 3059.13im, 棘皮动物共轴增生胶结, 下方胶结物多, 呈重力型胶结的大气淡水成岩特征; j— M316井, 2892.33im, 生物对厚壳蛤钻孔, 之后灰泥脱水收缩产生孔隙, 埋藏期压实作用产生裂缝; k— M316井, 2985.14im, 长形厚壳蛤碎屑的压实作用, 并导致颗粒破裂, 后发生压溶作用; l— M316井, 3060.13im, 海绵动物被压实破裂, 棘皮动物发生压溶作用Fig.5 Diagenetic features in the Mishrif Formation, HF Oilfield of Iraq |
通过矿物形态、溶解与胶结充填关系分析, 结合区域埋藏史, 建立HF油田Mishrif组生物碎屑灰岩储集层成岩序列, 确定不同成岩事件发生的阶段及其对储集层物性的影响。HF油田Mishrif 组生物碎屑灰岩成岩作用具有两极分化的特征, 开阔台地台内滩和台地边缘等中高能相带处于古地貌的高部位, 容易出露海面接受大气淡水的溶蚀改造, 溶蚀产物也能较快向周围排出, 使得储集层物性变好(金值民等, 2018; Zhong et al., 2018a, 2018b)。局限台地和斜坡等低能相带处于古地貌的低部位, 溶蚀作用发育较弱, 且还接受来自高地貌区溶蚀产物的沉淀, 储集层物性变差(王君等, 2016; 王昱翔等, 2017)。HF油田Mishrif组沉积之后, 经历同生成岩、早成岩和中成岩演化, 其中同生成岩阶段成岩作用较为显著, 根据成岩环境, 可进一步将同生成岩阶段划分为早期和晚期(图 6)。
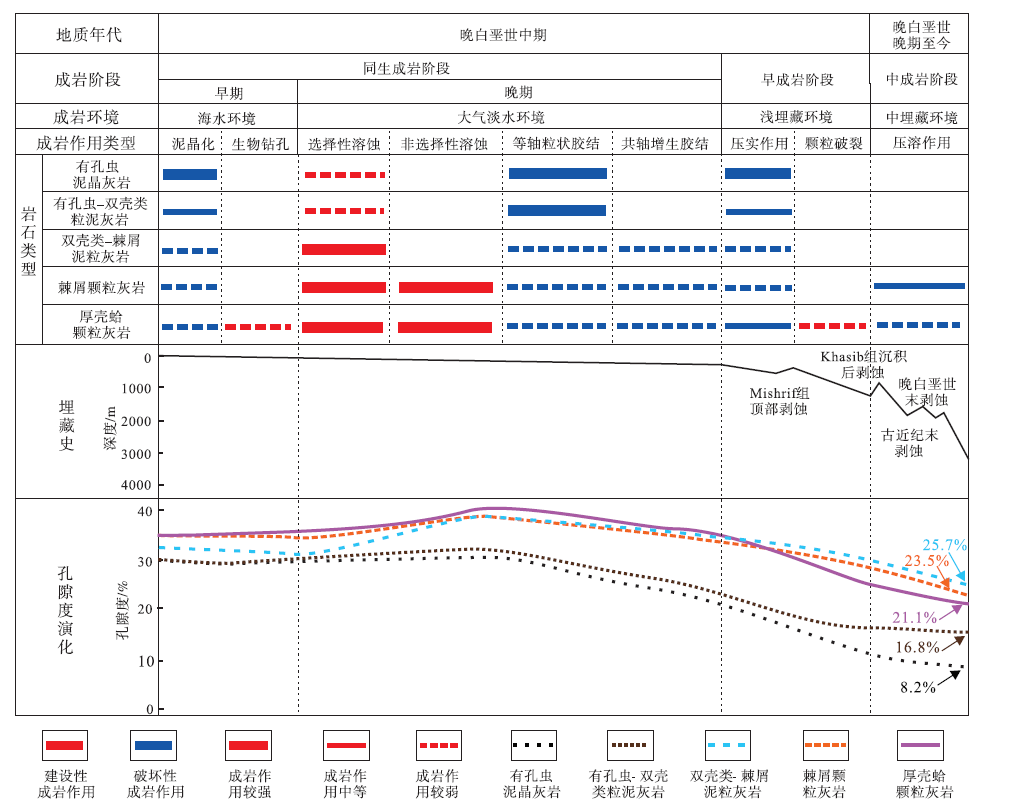 | 图 6 伊拉克HF油田Mishrif 组成岩作用序列Fig.6 Diagenesis sequence in the Mishrif Formation, HF Oilfield of Iraq |
同生成岩阶段早期。在海水环境中, 藻和真菌类对碳酸盐质壳体自外向内反复穿孔, 后又被泥晶充填, 略微降低孔隙度。这类泥晶化作用在有孔虫泥晶灰岩中较为明显, 其次是有孔虫— 双壳类粒泥灰岩。随着底栖有孔虫的变大, 泥晶套厚度增加, 泥晶化程度加深。另外, 微生物对厚壳蛤壳体进行钻孔, 形成近似椭圆形的孤立粒内孔, 虽然后多被灰泥充填, 但孔隙度有一定程度的提升(图 5-j)。
同生成岩阶段晚期。在大气淡水环境中, 有孔虫泥晶灰岩和有孔虫— 双壳类粒泥灰岩处在低部位, 溶蚀作用较弱, 而有孔虫原生体腔孔和双壳类类溶蚀形成的铸模孔, 大部分处于开启状态, 这些孔隙被等轴粒状方解石所充填, 孔隙度有较大规模的下降。双壳类— 棘屑泥粒灰岩在大气淡水环境中遭受短时周期性淋滤, 文石质非固着类双壳类发生选择性溶蚀作用形成铸模孔(图 5-a, 5-b), 孔隙度有较大幅度提升; 少量孔隙在淡水潜流带被等轴粒状方解石充填, 而棘皮动物碎屑多为方解石单晶, 胶结物围绕其外圈共轴增长(图 5-i), 降低储集层孔隙度。棘屑颗粒灰岩和厚壳蛤颗粒灰岩发育于水动力强的环境中, 其沉积速率快, 易暴露于大气淡水环境, 遭受短时周期性淋滤, 不稳定的成分首先发生溶蚀。当不稳定的矿物被溶蚀后, 较稳定的厚壳蛤和棘皮动物碎屑发生溶蚀作用, 颗粒边界凹凸不平, 出现溶蚀港湾, 使得原有孔隙扩大, 形成粒间溶孔(图 5-e), 孔隙度有较大幅度的增加。粒间溶孔进一步发育, 形成直径大于1imm的溶蚀孔洞(图 5-f), 孔隙度继续上升。溶蚀产物为淡水潜流带的再沉淀提供了溶质, 少量孔隙被等轴粒状方解石充填, 棘皮动物碎屑胶结物围绕其外圈共轴增长, 降低储集层孔隙度。
早成岩阶段。在浅埋藏环境中, 在上覆地层的负荷压力下, 压实作用使孔隙流体减少、沉积物密度增加, 细粒的有孔虫泥晶灰岩压实作用明显, 其次是有孔虫— 双壳类粒泥灰岩。双壳类— 棘屑泥粒灰岩和棘屑颗粒灰岩为颗粒支撑结构, 抗压实作用能力较强, 压实作用的减孔效应较弱。而厚壳蛤颗粒灰岩受“ 强溶蚀、弱胶结” 的影响, 胶结物含量低, 难以减弱压实作用。此外, 部分厚壳蛤碎屑为长条形, 且非选择性溶蚀形成不规则的边界, 这些因素进一步导致厚壳蛤颗粒灰岩抗压实能力较弱。压实作用使长条形厚壳蛤碎屑在厚度较小处发生破裂, 并呈现定向排列(图 5-k)。虽然颗粒破裂形成部分颗粒间微裂缝, 但其增孔效果有限, 压实作用一定程度上降低了厚壳蛤颗粒灰岩孔隙度。
中成岩阶段。中埋藏环境中, 在上覆地层的应力作用下, 压实作用继续进行, 但与浅埋藏环境比, 强度要小。颗粒接触处应力局部集中, 发生压溶作用, 在棘屑颗粒灰岩中较为明显, 其使得颗粒间呈凹凸或缝合接触, 颗粒接触处溶解的方解石又会就近沉淀(图 5-l), 孔隙度降低。
不同岩石类型的生物碎屑灰岩成分和形态特征组成不同, 在海水环境、大气淡水环境、埋藏环境中具有差异成岩特征(图 6)。大气淡水溶蚀作用是储集层的关键性建设作用, 而胶结和压实、压溶作用是储集层主要的破坏作用。有孔虫泥晶灰岩大气淡水环境中体腔孔被等轴粒状胶结物所充填, 埋藏环境中压实作用显著, 具有“ 弱溶蚀、强胶结、强压实” 的成岩特征。有孔虫— 双壳类粒泥灰岩在大气淡水环境中, 体腔孔和铸模孔被等轴粒状胶结物所充填, 埋藏环境中压实作用较显著, 具有“ 弱溶蚀、强胶结、中等压实” 的成岩特征。双壳类— 棘屑泥粒灰岩, 大气淡水环境中选择性溶蚀作用显著, 具有“ 较强溶蚀、弱胶结、较弱压实” 的成岩特征。棘屑颗粒灰岩大气淡水环境中选择性和非选择型溶蚀作用显著, 埋藏环境中压溶作用较显著, 具有“ 强溶蚀、弱胶结、中等压溶” 的成岩特征。厚壳蛤颗粒灰岩大气淡水环境中选择性和非选择型溶蚀作用显著, 埋藏环境中长条形的厚壳蛤壳体发生破裂, 形成生物碎片并定向排列, 具有“ 强溶蚀、弱胶结、中等压实” 的成岩特征。
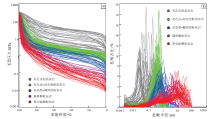
铸体薄片和高压压汞测试结果表明, Mishrif组生物碎屑灰岩孔隙类型和孔喉结构多样, 岩石类型不同, 其发育的孔隙类型也存在较大的差异(图 7)。
 | 图 7 伊拉克HF油田Mishrif 组不同岩石类型的生物碎屑灰岩孔喉结构特征 a— 5类储集层的进汞曲线图; b— 5类储集层的孔喉分布图Fig.7 Pore and throat structure characteristics of bio-clastic limestone of different rock types in the Mishrif Formation, HF Oilfield of Iraq |
有孔虫泥晶灰岩孔隙发育程度低, 主要是生物体腔孔被方解石完全充填后形成晶间孔(图 5-g), 泥晶基质中发育分散状的微孔隙; 排驱压力大于4iMPa, 孔喉分布集中, 以0.03~0.15iμ m的微喉为主, 呈现偏细态单峰型和尖峰型特征。有孔虫— 双壳类粒泥灰岩孔隙空间被方解石部分充填形成晶间孔和粒内孔(图 5-h); 排驱压力为0.65~3.1iMPa, 孔喉分布集中, 以0.25~0.75iμ m细微喉为主, 呈现偏细态尖峰型特征。双壳类— 棘屑泥粒灰岩主要发育大量铸模孔、粒内孔, 但孔隙之间的连通性较差(图 5-a, 5-b); 排驱压力为0.14~0.8iMPa, 孔喉分布相对集中, 以0.5~3iμ m中细喉为主, 呈现偏粗态单峰型特征。棘屑颗粒灰岩以残余粒间孔、粒间溶孔为主, 连通性较好(图 5-l); 排驱压力为0.03~0.14iMPa, 孔喉从0.1iμ m到20iμ m之间均有分布, 以中粗喉为主, 呈现偏粗态宽峰型特征。厚壳蛤颗粒灰岩主要为粒间孔、粒间溶孔, 并含有一定的溶蚀孔洞, 孔隙尺度大, 连通性好(图 5-f); 排驱压力仅为0.01~0.03iMPa, 孔喉从0.1iμ m到60iμ m之间均有分布, 以大于10iμ m的粗喉道为主, 呈现偏粗态极宽峰型特征。
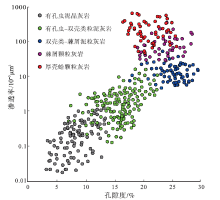
根据常规物性的统计结果, Mishrif 组岩心孔隙度为 3.1%~29.5%, 渗透率为(0.02~656.1)× 10-3 μ m2, 非均质性强(图 8)。
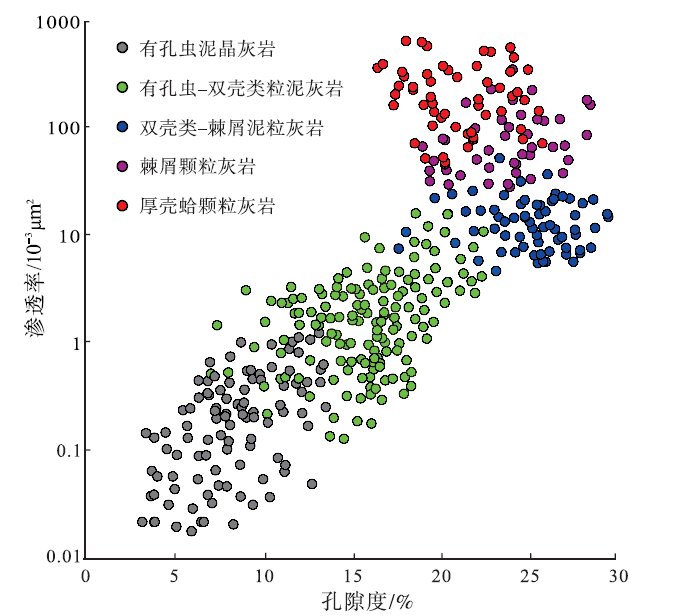 | 图 8 伊拉克HF油田Mishrif 组不同岩石类型的生物碎屑灰岩物性特征Fig.8 Physical property of different rock types of bio-clastic limestone in the Mishrif Formation, HF Oilfield of Iraq |
有孔虫泥晶灰岩孔隙度为3.1%~13.2%, 平均值8.2%, 渗透率为(0.02~1.04)× 10-3μ m2, 平均值0.31× 10-3 μ m2, 属于低孔低渗非储集层。有孔虫— 双壳类粒泥灰孔隙度为6.9%~22.4%, 平均值16.8%, 渗透率为(0.24~7.6)× 10-3 μ m2, 平均值2.42× 10-3 μ m2, 属于中低孔低渗储集层。双壳类— 棘屑泥粒灰岩孔隙度为21.8%~29.5%, 平均值25.7%, 渗透率为(5.2~32.8)× 10-3 μ m2, 平均值14.8× 10-3 μ m2, 属于高孔中渗储集层。
棘屑颗粒灰岩孔隙度为19.9%~27.8%, 平均值23.5%, 渗透率为(28.1~352.8)× 10-3 μ m2, 平均值97.5× 10-3 μ m2, 属于高孔中高渗储集层。厚壳蛤颗粒灰岩孔隙度为16.5%~25.8%, 平均值21.1%, 渗透率为(51.7~656.1)× 10-3 μ m2, 平均值248.66× 10-3 μ m2, 是Mishrif 组物性最好的储集层。
从泥晶灰岩到颗粒灰岩, 生物碎屑含量增多, 由泥晶支撑转变为颗粒支撑, 储集层物性逐渐变好, 但孔渗变化不一致(表 3)。其中双壳类— 棘屑泥粒灰岩的孔隙度高于厚壳蛤颗粒灰岩, 但渗透率低, 这是由于其中易溶的非固着类双壳类、苔藓动物和藻类碎屑等占比高, 但生物碎屑之间存在一定的泥晶, 孔隙为连通程度不高的铸模孔, 渗流能力不强。棘皮动物和厚壳蛤颗粒灰岩都位于台地边缘滩高能环境, 早期成岩作用特征相似, 棘皮动物碎屑多呈等轴粒状, 易于形成粒间孔, 但共轴增生胶结和压溶作用使得其喉道缩小; 而厚壳蛤在埋藏过程中压实作用显著, 降低了储集层的孔隙度, 但等轴粒状胶结和压溶作用较弱, 喉道缩小程度低, 使得棘屑颗粒灰岩孔隙度虽高于厚壳蛤颗粒灰岩, 但渗透率较低。
| 表 3 伊拉克HF油田Mishrif 组不同岩石类型生物碎屑灰岩差异成岩和储集层特征 Table3 Diagenesis and reservoir characteristics of bio-clastic limestone of different rock types in the Mishrif Formation, HF Oilfield of Iraq |
伊拉克HF油田Mishrif组生物类型多样, 包括底栖有孔虫、非固着类双壳类、厚壳蛤与棘皮动物, 以及少量苔藓动物、藻类与海绵动物。沉积环境决定了生物种类、含量、颗粒大小、形态特征、成分甚至分布方式的差别, 进而决定了成岩环境、成岩途径、成岩作用类型、成岩强度以及成岩序次上的差异。大气淡水溶蚀作用是储集层的关键性建设作用, 而胶结和压实、压溶作用是储集层主要的破坏作用。有孔虫泥晶灰岩具有“ 弱溶蚀、强胶结、强压实” 的成岩特征, 有孔虫— 双壳类粒泥灰岩具有“ 弱溶蚀、强胶结、中等压实” 的成岩特征, 双壳类— 棘屑泥粒灰岩具有“ 较强溶蚀、弱胶结、较弱压实” 的成岩特征, 棘屑颗粒灰岩具有“ 强溶蚀、弱胶结、中等压溶” 的成岩特征, 厚壳蛤颗粒灰岩具有“ 强溶蚀、弱胶结、中等压实” 的成岩特征。
沉积背景决定岩石的组分与结构, 在此基础上的相控成岩作用决定岩石的物性与孔隙结构特征。以底栖有孔虫和非固着类双壳类碎屑为主的泥晶灰岩和粒泥灰岩, 主要发育微孔、晶间孔及粒内孔, 为细微喉中低孔低渗储集层; 双壳类— 棘屑泥粒灰岩易溶的生物碎屑占比高, 以弱连通铸模孔为主, 为中细喉高孔中渗储集层; 棘屑颗粒灰岩和厚壳蛤颗粒灰岩均以连通性好的粒间(溶)孔为主, 前者受共轴增生胶结及压溶作用导致喉道缩小, 为中粗喉高孔中高渗储集层, 后者为粗喉中高孔高渗最优质储集层, 主要分布于台地边缘滩中。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|