







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
辽东半岛复州湾剖面寒武系第二统光合作用生物膜建造的核形石*
[梅冥相1, 2  , MuhammadRiaz
, MuhammadRiaz1 , 刘丽1 , 孟庆芬1 ]
, MuhammadRiaz|
|

第一作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作。E-mail: meimingxiang@263.net。
作为一种经常大于鲕粒的包覆颗粒类型,核形石以其不平滑的圈层被解释为微生物成因,区别于成因存在较大争议的鲕粒,而且常与鲕粒和其他类型的碳酸盐颗粒相互共生;作为一种在围绕着生物碎屑和非生物碎屑核心的序列式纹层化作用过程中形成的球形或假球型生物沉积构造,核形石还常常单独产出和分布,所以又被解释为微生物碳酸盐岩,或者被归为球状叠层石。在辽东半岛寒武系第二统碱厂组和馒头组之中,厘米级别大小的核形石密集发育在三级层序的顶部,成为一种时间特化的相。另外,以下重要特征将辽东半岛寒武系第二统的核形石特征化,包括: (1)与凝块和微凝块共生;(2)多为球状和椭球状;(3)由不均一的非纹层状致密泥晶和微亮晶构成;(4)核形石皮层以及核形石间凝块中发育特别的蓝细菌鞘钙化化石等。尽管穿越成岩作用过滤器去解释古代核形石复杂的形成机理存在着巨大的挑战,也尽管形成这些核形石的复杂生物膜钙化作用细节需要更加深入地研究才能得到更好地了解,但是,辽东半岛第二统碱厂组和馒头组核形石中直接的微生物化石证据,尤其是核形石内较为丰富的钙化蓝细菌鞘化石,使其成为一个了解光合作用生物膜建造核形石的典型实例。
About the first author Mei Ming-Xiang,born in 1965, is a professor at School of Earth Sciences and Natural Resources,China University of Geosciences(Beijing),and is engaged in sedimentology and stratigraphy. E-mail: meimingxiang@263.net.
As a type of coated grains with the size of more than that of ooids,oncoid is marked by non-smooth circles and is commonly interpreted as a microbial origin,so different from ooid whose origin remains uncertain. Also as a variety of spherical or pseudo-spherical biosedimentary structures formed as successive laminations around both biogenic and nonbiogenic nuclei,its widespread distribution and development by oneself imply that the oncoid can be grouped into a type of microbial carbonates or a kind of spherical stromatolites. Within the Jianchang and Mantou Formations of the Series 2 of Cambrian in the Liaodong Peninsula,oncoids with the size of centimeter level as a time-specific facies concentrate in the top part of third-order sequence. In addition,several features characterize these oncoids of the Series 2 of Cambrian in the Liaodong Peninsula: (1)the symbiosis with clots or microclots;(2)the spherical and ellipsoid morphology;(3)the inhomogenous or non-layer composition of dense micrite and microspar;(4)particular calcified sheath fossils of the cyanobacteria within the cortex of oncoids and the clot among oncoids and so on. Although the greatest challenge that remains is the interpretation of ancient oncoids and their detail forming processes through the filter of the diagenesis,also the detail of the calcification of biofilms dominated by the cyanobacteria has still not got better understanding,the richness of direct evidence represented by microbial fossils especially for sheath fossils of the cyanobacteria within oncoids of both the Jianchang and the Mantou Formations of the Series 2 of Cambrian in the Liaodong Peninsula make them become a good example of oncoids built by non-layer photosynthetic biofilms.
核形石(Oncoids), 一个由Heim于1916年所提出的术语, 原指发现在瑞士侏罗系陆架灰岩中的泥晶颗粒, 且被认为是微生物岩的一种。作为一种包覆颗粒类型, 核形石具有扰动的和不平滑的圈层以及较大的粒径, 表明其具有明显的微生物成因(Tucker and Wright, 1990; 梅冥相等, 1997; 梅冥相, 2007; Flü gel, 2010; Han et al., 2015; Wilmeth et al., 2015; 齐永安等, 2016), 从而区别于成因还存在争议的鲕粒。核形石常与凝块石共生, 前苏联地质学家将二者统称为微植石(Microphytolite)以与叠层石相区别, 而杜汝霖(1992)将凝块石(Thrombolite)等同于花纹石(Catagraphite), 并认为微植石是核形石和花纹石的总称。古生物学家曾经采用林奈法将核形石划分为3个亚类, 并进一步划分为10个群(详见杜汝霖(1992)的总结), 3个亚类分别是具同心纹层构造的核形石(以Osagia为代表)、具放射状构造的核形石(以Asterrospheroides为代表)及具同心纹层和放射状构造的核形石(以Pilisphaeroides为代表); 但是, 随着蓝绿藻的生物学属性被修订为蓝细菌(梅冥相等, 2007, 2017; 梅冥相, 2011a, 2012, 2014; 梅冥相和孟庆芬, 2016), “ 微植石(Microphytolite)” 的概念应该被抛弃了。核形石还被归为球状叠层石, 并成为微生物碳酸盐岩的一种类型(Riding, 2000; 曹瑞骥和袁训来, 2006; 梅冥相, 2007), 具有与叠层石相似的微生物碳酸盐岩的沉积学属性(Riding, 2000, 2011a; 梅冥相, 2007; Flü gel, 2010; Bosak et al., 2013; 梅冥相和孟庆芬, 2016; Peters et al., 2017); 但是, 与叠层石不同的是, 核形石经常作为保存着钙化微生物化石的球形可迁移颗粒, 又具有与鲕粒相似的包覆颗粒的沉积学属性(Tucker and Wright, 1990; 梅冥相等, 1997; 梅冥相, 2007; Vé drine et al., 2007; Flü gel, 2010; Jones, 2011; 杨仁超等, 2011; Liu and Zhang, 2012; Mei and Gao, 2012; Zaton et al., 2012; 张文浩等, 2014; Han et al., 2015; Pederson et al., 2015; Wilmeth et al., 2015; Lozano et al., 2016; Pretković et al., 2016; 齐永安等, 2017a, 2017b)。如同鲕粒形成于复杂的微生物膜钙化作用, 也类似鲕粒产出和分布在前寒武纪至今天的海水和淡水环境之中, 核形石本身确切的形成机理以及其与鲕粒形成机理和产出特征之间的差异, 还没有得到完全的了解。
辽东半岛寒武系第二统碱厂组和馒头组之中, 厘米级别大小的核形石密集发育在三级层序的顶部并成为一种时间特化的相, 构成特别的厚度数米的核形石颗粒滩。特殊的宏观产出特征及微观现象, 尤其是在核形石内部发现的葛万菌之类的丝状蓝细菌鞘化石(Riding, 1991, 2000)以及特殊的胶须菌(Rivulariaceae)之类的钙化蓝细菌化石(Riding, 1991; Whitton and Mateo, 2012; Liu et al., 2016a, 2016b, 2017), 表明核形石形成于较为典型的光合作用生物膜, 从而使其成为一个较为壮观的沉积学现象, 为了解形成核形石复杂的生物膜钙化作用(Decho, 2010; Flemming and Wingender, 2010; Flemming et al., 2016; Decho and Gutierrez, 2017)提供了一个较为典型的实例。
自寒武纪第二世至中奥陶世, 华北地台是一个典型的陆表海台地(冯增昭等, 1990, 2004; Meng et al., 1997; Kwon et al., 2006; Lee et al., 2014)。该地台为一个稳定的克拉通, 即中— 朝地块, 在早古生代大致位于接近古赤道的冈瓦纳大陆的边缘(Yang et al., 2002; McKenzie et al., 2011)。华北地台在寒武纪接受沉积的时间较晚, 大致从寒武纪第二世晚期开始接受沉积, 而且第二世的地层超覆于不同时代的前寒武纪地层之上, 在二者之间形成一个类似于北美地台的“ 巨型不整合面” (Peters and Gaines, 2012)。所以, 在寒武纪华北地台形成一个特别的沉积序列(图1): 从寒武纪第二世红层与碳酸盐岩混合沉积(Meng et al., 1997; 冯增昭等, 2004; Lee and Chough, 2011)开始, 经苗岭世鲕粒滩主导的碳酸盐岩台地序列(马永生等, 2017), 变化到芙蓉世碳酸盐泥所主导的碳酸盐岩台地沉积序列(梅冥相等, 2017)。
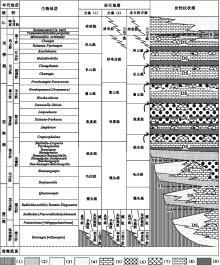
 | 图1 华北地台寒武系沉积序列 三叶虫生物地层序列引自彭善池(2009); 岩石地层划分方案(1)引自冯增昭等(1990, 2004)、卢衍豪等(1994)、 Meng等(1997), 方案(2)引自项礼文等(1999), 本文采用的方案沿用方案(1)并基于沉积相序列的层序地层划分予以修改, 即华北地台寒武系大致可以划分为9个三级层序(DS1至DS9), 包含着2种类型的层序界面即暴露间断面构成的层序界面(SB)和淹没不整合型层序界面(箭头所指)。 图例的含义分别是: (1)地层缺失, (2)海岸萨布哈相红层泥岩地层, (3)潮上砂泥坪相砂泥岩地层, (4)潮坪相白云岩地层, (5)浅缓坡相灰岩地层, (6)鲕粒滩相灰岩地层, (7)微生物碳酸盐岩生物丘, (8)中至深缓坡相泥灰岩和灰岩地层, (9)陆棚相钙质泥岩Fig.1 Sedimentary succession of the Cambrian in North-China Platform |
华北地台寒武纪最早期的地层, 在山东一带称为李官组和朱砂洞组(Lee and Chough, 2011; Lee et al., 2014), 河南一带被称为辛集组, 北京一带被归为昌平组, 在辽东半岛则划分为大林子组与碱厂组。这些地层在寒武纪第二世(彭善池, 2009; Peng et al., 2012)的辛集组沉积时期(Meng et al., 1997; 冯增昭等, 2004)围绕着西北部的大片古陆分布, 大致构成3个相带(图2): (1)海岸萨布哈相, (2)局限潮坪相, (3)开阔潮坪相。在寒武纪第二世晚期的馒头组沉积时期(Meng et al., 1997; 冯增昭等, 2004), 沉积格局的最大变化是随着海侵的继续(Meng et al., 1997; 冯增昭等, 2004; Lee and Chough, 2011), 古陆范围逐渐变小(图3), 也大致由类似于图2所示的早期的3个相带所构成。总的来讲, 局限潮坪相带中的寒武系第二统, 与开阔潮坪相带比较, 表现出以下变化(图4):(1)海岸萨布哈相的红色泥岩地层厚度变大; (2)局限潮坪相中的白云岩地层相变为开阔潮坪相中的灰岩地层, 而且在辽东半岛的碱厂组和馒头组顶部集中发育核形石滩相地层而成为文中的研究对象, 它们分别组成三级层序DS1与DS2的晚期高位体系域(图1; 图4; 梅冥相, 2010, 2011b; Catuneanu et al., 2011)。
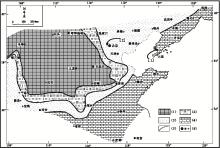
 | 图2 华北地台寒武纪辛集组沉积期的沉积格局 图中的[1]代表开阔潮坪相的辽东半岛复州湾剖面的位置, [2]所指为北京西郊下苇甸剖面的位置; 修改自冯增昭等(1990, 2004)。图例代号的含义分别是: (1)古陆, (2)海岸萨布哈相, (3)局限潮坪相, (4)开阔潮坪相, (5)相变线, (6)城市Fig.2 Depositional setting of the sedimentary period of Xinji Formation in North-China Platform |
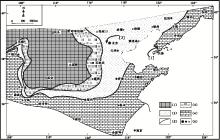
 | 图3 华北地台寒武纪馒头组沉积期的沉积格局 图中的[1]和[2]所指的剖面位置以及图例代号的含义与图2相同; 修改自冯增昭等(1990, 2004)Fig.3 Depositional setting of the sedimentary period of Mantou Formation in North-China Platform |
 | 图4 华北地台寒武系第二统不同相带的剖面对比 剖面图中, 左边为北京西郊下苇甸剖面, 代表局限潮坪相, 右边为辽东半岛复州湾剖面, 代表开阔潮坪相, 2个剖面的位置如图2和图3所示。 寒武系第二统大致可以划分为2个三级层序(DS1至DS2), 均由海侵体系域(TST)和高位体系域(HST)所构成。 照片所代表的沉积现象分别是: 在北京西郊下苇甸剖面, (a)毛庄组底部的海岸萨布哈相红色泥岩, (b)DS2的顶界面(箭头所指), (c)昌平组下部的海岸萨布哈相红色泥岩, (d)昌平组下部的潮坪相白云岩地层, (e)昌平组与前寒武系井儿峪组之间的平行不整合面, 代表大于3× 108年的沉积间断; 在辽东半岛复州湾剖面, (f)DS2的顶界面(箭头所指), (g)馒头组顶部的块状核形石灰岩地层, (h)馒头组下部与灰岩互层的局限潮坪相红层沉积, (i)DS1的顶界面(箭头所指), (j)碱厂组顶部的块状核形石灰岩地层, (k)碱厂组中部的厚层块状灰岩地层。 图例的含义分别是: (1)海岸萨布哈至局限潮坪相红色泥岩地层, (2)潮坪相砂泥岩地层, (3)潮坪相白云岩地层, (4)潮间至潮下坪相灰岩地层, (5)核形石碳质地层, (6)夹灰岩层的红层沉积地层, (7)层序界面, (8)相变面Fig.4 Correlation of stratigraphic sections of the Series 2 of Cambrian belonging to different sedimentary facies zones in North-China Platform |
如图4所示, 碱厂组的核形石滩相灰岩地层的厚度为6 m左右, 由块状灰岩组成(图4中的照片i和j所示)。该套核形石灰岩中, 核形石的粒径为厘米级别(图5), 最大可以达到2~3 cm, 多为0.5~1.0 cm。这些核形石的形状多为圆形和椭圆形, 核心多为生物碎屑或形态不规则的凝块, 皮层由不规则纹层状的泥晶或块状泥晶所组成, 核形石之间被凝块和凝块状泥晶所填充。从大小上看, 其属于宏观核形石到巨型核形石的范畴(Flü gel, 2010; Pretković et al., 2016); 从构成上分析, 核形石皮层中的纹层不明显和不发育是碱厂组核形石的基本特征, 它们类似于Vé drine等(2007)所描述的由泥晶主导的核形石, 也类似于Han等(2014)所描述的凝块状皮层核形石和完全皮层(没有核心的)核形石, 还类似于Wilmeth等(2015)所描述的基质支撑的核形石。由于这些核形石的包覆颗粒属性, 以及它们集中产出的特征, 所以形成较为特别的核形石滩。
 | 图5 辽东半岛复州湾剖面寒武系第二统碱厂组顶部核形石灰岩的宏观特征Fig.5 Macroscopic features for oncoid in the top part of Jianchang Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
碱厂组核形石在显微镜下为致密的非均质泥晶皮层(图6-a, 6-c), 核心不明显(图6-a)或以三叶虫化石作为核心(图6-c), 皮层中的纹层不明显或不发育, 表现为泥晶主导的核形石(Vé drine et al., 2007)或凝块状皮层的核形石和完全皮层的(没有核心的)核形石(Han et al., 2015); 在核形石内部, 残留着以谜一样的可能的胞网菌(Bacinella; Vé drine et al., 2007; Schlagintweit and Bover-Arnal, 2013)状组构为代表的钙化蓝细菌(图6-a, 6-b)。这些大型或巨型核形石之间多为凝块所填充, 在填充基质之中还见到可以类比于现代胶须菌(Rivularia)的岩枝菌(Apophoretella)之类的钙化蓝细菌化石(Riding, 1991; Whitton and Mateo, 2012; Liu et al., 2016a, 2016b, 2017)构成的凝块(图6-b)。
 | 图6 辽东半岛复州湾剖面寒武系第二统碱厂组核形石的微观特征 a— 不发育核心的次圆形核形石, 箭头所指为可能的孢网菌(Bacinella)状组构; b— 巨型核形石间的凝块(clot)以及以孢网菌(Bacinella)状组构(直线箭头所指)为核心的小型核形石, 注意可能的岩枝菌(Apophoretella)构成的凝块(内插图所示); c— 以三叶虫为核心的次圆形核形石, 为非均质的暗色泥晶构成核形石的非纹层状皮层; d— 照片c中白色巨型所圈定的暗色泥晶的放大, 表现出为清楚的葛万菌(Girvanella)蓝细菌鞘化石构成; e— 照片的左下部的进一步放大, 其中的黑点可能为黄铁矿晶体的残余物; f— 充填在核形石之间的葛万菌(Girvanella)蓝细菌鞘化石构成的凝块Fig.6 Microscopic features for oncoid of the Jianchang Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
更为特征的是, 在那些在低倍显微镜下表现为暗色泥晶的核形石皮层(图6-c)之中, 发育着较为清楚的可以类比于现代织线菌(Plectonema; Riding, 1991; De los Rí os et al., 2015)的葛万菌(Girvanella)蓝细菌鞘化石和黄铁矿晶体残余物(图6-d, 6-e)。类似的丝状蓝细菌也在核形石间的凝块中发现(图6-f), 而且这种类似丝状的蓝细菌鞘化石还被类比于伪枝菌(Scytonema; 齐永安等, 2017b)。
就像图6-c所示的可以类比于现代胶须菌(Rivularia)的岩枝菌(Apophoretella)之类的钙化蓝细菌化石构成的凝块那样, 在巨型核形石的非均质致密泥晶和微亮晶构成的皮层之中, 也见到较为清楚的岩枝菌(Apophoretella)之类的钙化蓝细菌化石(图7; Riding, 1991; Whitton and Mateo, 2012; Liu et al., 2016a, 2016b, 2017)。以外, 在核形石皮层中还见到三叶虫化石和微凝块。
 | 图7 辽东半岛复州湾剖面第二统碱厂组巨型核形石皮层中的岩枝菌(Apophoretella) a— 无核心的完全皮层巨型核形石, 白色框代表较为典型的岩枝菌(Apophoretella), 箭头所指为三叶虫化石; b, c— 为岩枝菌(Apophoretella)钙化蓝细菌化石的进一步放大Fig.7 Apophoretella within cortex of oncoid of the Jianchang Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
同时, 在巨型泥晶核形石的皮层之中, 也发现较为典型的窗格状孔隙(图8), 其类似于Wilmeth等(2015)在蒙古西部下寒武统Bayan Gol组核形石中所描述的现象。这样的窗格状组构, 已在现代黄石国家公园的蓝细菌微生物席所构成的叠层石中被观察到, 并被解释为光合作用产生的氧气气泡的残余物(Bosak et al., 2010; Pepe-Ranney et al., 2012)。由于这些窗格状孔隙被较细的垂向丝状体所分开, 由此又被Mata等(2012)定义为“ 砂滤状(hourglass structures)” 构造。但与黄石国家公园现代硅质叠层石中的窗格状构造(Bosak et al., 2010; Mata et al., 2012; Pepe-Ranney et al., 2012)不同的是, 碱厂组核形石皮层中的窗格状孔隙, 发育在相互缠绕而且可以类比于现代织线菌(Plectonema; Riding, 1991; De los Rí os et al., 2015)的葛万菌(Girvanella)丝状蓝细菌鞘化石构成的网状体之中(图8-b, 8-c)。
 | 图8 辽东半岛复州湾剖面寒武系第二统碱厂组巨型核形石皮层中的窗格状孔隙 a— 巨型核形石边缘的窗格状孔隙; b, c— 照片a中矩形框所示之处的进一步放大, 表现出窗格状孔隙发育在葛万菌之类的丝状蓝细菌鞘化石构成的网状体内Fig.8 Fenestral pore within cortex of oncoid of the Jianchang Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
如图4所示, 馒头组的核形石滩相灰岩地层厚度为8 m左右, 由块状灰岩组成(图4中的照片f和g所示)。馒头组顶部的核形石, 多为椭圆形, 少数为次圆形, 大小多在0.5~1.5 cm之间, 属于宏观核形石到巨型核形石的范畴(Flü gel, 2010; Pretković et al., 2016)。从构成上分析, 核形石皮层中强烈扰动或不明显的纹层是馒头组核形石的基本特征, 它们类似于Vé drine等(2007)所描述的由泥晶所主导的核形石, 也类似于Han等(2015)所描述的具凝块状皮层的核形石和完全皮层的(没有核心的)核形石。核形石之间填充着较小(0.5~1.5 mm)的凝块之类的颗粒, 类似于Wilmeth等(2015)所描述的基质支撑的核形石。如同图5所示的碱厂组顶部的核形石那样, 由于馒头组核形石的包覆颗粒属性, 以及它们的集中产出特征, 所以亦形成较为特别的核形石滩。
与图6至图8所示的碱厂组核形石相似, 在显微镜下馒头组核形石表现为致密的非均质泥晶皮层, 由暗色泥晶和微亮晶构成(图9-a), 核心不明显(图9-a)或以三叶虫生物碎屑作为核心; 核形石皮层中的纹层不明显或不发育, 表现为泥晶主导的核形石(Vé drine et al., 2007)或凝块状皮层的核形石和完全皮层的(没有核心的)核形石(Han et al., 2015)。在核形石内部, 残留着谜一样的可能的胞网菌(Bacinella; Vé drine et al., 2007; Schlagintweit and Bover-Arnal, 2013)状组构代表的钙化蓝细菌(图9-a, 9-b)。填充在核形石之间的凝块或微凝块, 在低倍显微镜下表现为由形态不规则的暗色泥晶构成, 但一些凝块在高倍显微镜下呈现为较为清晰的葛万菌(Girvanella)之类的丝状蓝细菌鞘化石(图9-f)。
 | 图9 辽东半岛复州湾剖面寒武系第二统馒头组顶部核形石灰岩的宏观特征Fig.9 Macroscopic features for oncoid in the top part of the Mantou Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
最为特别的是, 在馒头组巨型核形石的皮层之中, 除了一些由暗色泥晶构成的微凝块具有明显的丝状蓝细菌鞘化石以外, 较为明显地发育着可以类比于现代胶须菌(Rivulariaceae)的基座菌(Hedstroemia; Riding, 1991; Whitton and Mateo, 2012; Liu et al., 2016a, 2016b, 2017)之类的钙化蓝细菌化石(图9-b, 9-c), 而且还见到成束生长的类似于尖头菌(Acuasiphonoria; Liu et al., 2016a, 2016b, 2017)之类的更像现代胶须菌(Whitton and Mateo, 2012)的钙化蓝细菌化石(图9-d, 9-e)。
就像图6-c所示的碱厂组核形石之间具有可以类比于现代胶须菌(Rivularia)的岩枝菌(Apophoretella)之类的钙化蓝细菌化石构成的凝块那样, 在馒头组核形石之间的凝块基质中, 也见到单独的岩枝菌(Apophoretella)之类的钙化蓝细菌化石构成的凝块(图11-a, 11-b); 而且, 在核形石之间的凝块和微凝块(图11-a, 11-c), 在低倍显微镜下多表现为由暗色泥晶构成, 但在高倍显微镜下则发现较为明显的葛万菌(Girvanella)之类的丝状蓝细菌鞘化石, 还见到黄铁矿晶体的残余物(图11-d, 11-e)。
之所以强调由基座菌组合的钙化蓝细菌化石(Riding, 1991; Liu et al., 2016a, 2016b, 2017)的胶须菌(Rivularia)生物亲和性, 是因为现代胶须菌(Rivularia)具有特征性的支撑它们生长循环的逐渐变细、变尖的香毛簇, 以及具有底端为异型细胞的一类较为高级的固氮蓝细菌, 从而进一步说明形成馒头组核形石的蓝细菌的多样化。
辽东半岛寒武系第二统碱厂组和馒头组顶部的宏观和巨型泥晶主导的核形石, 以颗粒滩的形式集中发育在三级沉积层序的顶部, 构成晚期高位体系域沉积(图4)。这些由暗色泥晶和微亮晶结壳构成的球形和椭球形核形石, 在保留着球状叠层石沉积属性(梅冥相, 2007; Flü gel, 2010; Riding, 2011; 梅冥相和孟庆芬, 2016; Peters et al., 2017)的同时, 最终形成经常保存着钙化蓝细菌鞘化石的可迁移的球形颗粒(Wilmeth et al., 2015), 成为较为壮观的沉积学现象。
虽然核形石常被解释为微生物成因, 但是直接的微化石证据常常较为缺乏(Tucker and Wright, 1990; 梅冥相等, 1997; 梅冥相, 2007; Vé drine et al., 2007; Flü gel, 2010; Jones, 2011; Liu and Zhang, 2012; Mei and Gao, 2012; Zaton et al., 2012; Han et al., 2015; Pederson et al., 2015; Wilmeth et al., 2015; Lozano et al., 2016; Pretković et al., 2016)。在寒武纪第二统碱厂组和馒头组顶部的颗粒滩相核形石中, 相似的中等尺度的致密暗色泥晶和微亮晶组构, 可以类比于现代织线菌(Plectonema; Riding, 1991; De los Rí os et al., 2015)中的葛万菌一类的, 他们的丝状蓝细菌鞘化石保存较高的密度; 也可以类比较为常见的可以归为现代胶须菌(Rivularia; Whitton and Mateo, 2012)的钙化蓝细菌化石(Riding, 1991; Liu et al., 2016a, 2016b, 2017), 包括可能的胞网菌(Bacinella; Vé drine et al., 2007; Schlagintweit and Bover-Arnal, 2013)状组构代表的钙化蓝细菌残余物。他们与图8所示的在丝状蓝细菌之间代表着光合作用产生的氧气气泡(Bosak et al., 2010; Mata et al., 2012; Pepe-Ranney et al., 2012; Wilmeth et al., 2015)的窗格状孔隙一起, 指示着这些核形石具有明显的生物成因。
那些可以类比于现代胶须菌(Rivularia; Whitton and Mateo, 2012)的岩枝菌(Apophoretella)之类的钙化蓝细菌化石, 不仅发育在核形石皮层之中(图7; 图10), 而且还可以构成毫米级别的凝块(图6-b; 图11-a, 11-b), 填充在核形石之间的基质中。最为重要的是, 不管是在核形石皮层中(图6-c, 6-d, 6-e; 图8), 还是在核形石之间的暗色泥晶组成的凝块与微凝块之中(图6-f; 图10-f; 图11-c, 11-d, 11-e), 可以类比于现代织线菌(Plectonema; Riding, 1991; De los Rí os et al., 2015)的葛万菌之类的丝状蓝细菌鞘化石, 均具有较高的密度。这些丝状蓝细菌鞘化石, 具有较为均匀的丝状体直径(10 μ m左右)以及均匀的暗色泥晶壁的厚度(1~2 μ m), 较为充分地表明了这些蓝细菌丝状体应该是活体鞘的钙化作用产物(Arp et al., 2001; Kah and Riding, 2007; Riding, 2000, 2011b)。而蓝细菌活体鞘的钙化作用, 是指一个直接与大气圈二氧化碳浓缩作用机制(CCMs)相联系的生态生理学机制(Kah and Riding, 2007; Riding, 2000, 2011b); 最为重要的是CCMs在蓝细菌中得到诱发的条件是当二氧化碳分压(
 | 图10 辽东半岛复州湾剖面寒武系第二统馒头组核形石的微观特征 a— 暗色泥晶和微亮晶组成的无核心的巨型核形石, 箭头所指代表可能的孢网菌(Bacinella)状组构, 核形石间填充着微凝块; b— 巨型核形石皮层的泥晶和微亮晶构成, 白色框所示为基座菌(Hedstroemia)之类的钙化蓝细菌化石, 箭头所指为可能的胞网菌(Bacinella)状组构; c— 为照片b中白色框所示的基座菌(Hedstroemia)之类的钙化蓝细菌化石的进一步放大; d— 巨型核形石皮层的泥晶和微亮晶构成, 白色框所示为成束生长的尖管菌(Acuasiphonoria)之类的钙化蓝细菌化石; e— 为照片d中白色框所示的基座菌(Hedstroemia)之类的钙化蓝细菌化石的进一步放大, 更像现代的胶须菌(Rivulariaceae); f— 充填在核形石之间的葛万菌(Girvanella)蓝细菌鞘化石构成的凝块Fig.10 Microscopic features for oncoid of the Mantou Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
 | 图11 辽东半岛复州湾剖面寒武系第二统馒头组核形石间的凝块基质的微观特征 a— 核形石之间的凝块和微凝块基质, 照片中部所示的是岩枝菌(Apophoretella)之类的钙化蓝细菌化石构成的凝块, 大小为2 mm左右; b— 照片a中的岩枝菌(Apophoretella)之类的钙化蓝细菌化石构成的凝块的局部放大; c— 核形石之间的暗色泥晶构成的形态不规则的凝块与微凝块; d, e— 分别代表照片c中白色框所示的暗色泥晶凝块的进一步放大, 表现了在这些暗色泥晶凝块中较为清晰的葛万菌 (Girvanella)蓝细菌鞘化石构成, 其中的黑点可能为黄铁矿晶体的残余物Fig.11 Microscopic features for clot matrix between oncoids of the Mantou Formation of Series 2 of Cambrian at Fuzhouwan section, Liaodong Peninsula |
图5以及图9所示的这些宏观和巨型核形石皮层中普遍的致密暗色泥晶和微亮晶构成(图6; 图7; 图8; 图10), 代表着一个复杂的蓝细菌主导的微生物膜或微生物席的钙化作用过程(Riding, 2000; Dupraz et al., 2009), 因为并非所有蓝细菌所主导的微生物席或微生物膜均能够钙化, 而且今天的绝大多数微生物席确实没有形成坚硬的岩石(Stal, 2012)。再者, 在核形石(图6-d, 6-e)及填充基质的凝块(图11-d, 11-e)中常见到的黄铁矿颗粒与晶体, 说明形成核形石的微生物沉淀作用(Riding, 2000)和早期石化作用(Dupraz et al., 2009)与硫酸盐还原细菌造成的硫酸盐还原反应(Rickard et al., 2017)存在着成因联系, 从而进一步表明在微生物席内碱度发动机(Gallagher et al., 2012)和细胞外聚合物质(Decho, 2010; Flemming and Wingender, 2010; Tourney and Ngwenya, 2014; Flemming et al., 2016; Decho and Gutierrez, 2017)构成的有机基质这2个关键要素的紧密耦合, 涉及到碳酸盐矿物的原地沉淀作用。进一步讲, 由于蓝细菌是最为成功的造席生物(Stal, 2012), 蓝细菌作为原始生产者的微生物活动, 最终造成了细胞外聚合物质(EPS)的生产作用, 并形成特别的生物膜和微生物席, 而且构成生物膜和微生物席EPS的硫酸盐还原细菌之类的异养细菌的降解作用, 又造成了矿物的沉淀作用(Dupraz et al., 2009, 2011; Bosak et al., 2013)。这样一个复杂的作用过程, 形成了寒武纪第二世非纹层状光合作用生物膜(或微生物席)所主导的核形石。因此, 具有叠层石属性而且还具有类似于鲕粒的包覆颗粒属性的核形石, 更加明显地表征着地球历史中微生物与沉积物乃至与流水之间极为复杂的相互作用, 从而具有代表着生命与环境的长时间演变的潜力; 但是, 由于要穿越成岩作用过滤器, 导致其确切的形成机制和过程中的许多细节问题, 还有待于更加深入全面的研究(Dupraz et al., 2009, 2011; Bosak et al., 2013)。
辽东半岛寒武系第二统碱厂组和馒头组中的宏观和巨型核形石, 集中发育在三级层序的顶部而构成晚期高位体系域沉积, 并形成较为独特的核形石滩, 代表着较为壮观的微生物沉积作用现象。这些核形石主要由暗色泥晶和微亮晶构成, 含有多样化的钙化蓝细菌化石所表征的微生物, 充分说明其属于典型的光合作用微生物席或微生物膜的钙化作用(包括微生物沉淀作用或早期石化作用)产物。尽管这些核形石确切的形成机制和生长过程中的许多细节问题, 还需要进一步的研究才能得到更加合理的解释, 但是, 直接的微生物化石证据, 尤其是在核形石内较为丰富的钙化蓝细菌鞘化石, 使其成为一个了解非纹层状光合作用生物膜建造核形石的典型而且重要的实例。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|