{kind=link}
{kind=link}
{kind=link}
{kind=link}
从水体到沉积物: 探寻有机质的沉积过程及其意义*
[蔡进功1  , 曾翔
, 曾翔1 , 韦海伦1 , 宋明水2 , 王学军2 , 刘庆2 ]
, 曾翔|
|

第一作者简介 蔡进功,男,1961年生,1983年获南京大学学士学位,2003年获同济大学博士学位,现为同济大学海洋与地球科学学院教授、博士生导师。目前主要从事沉积学和石油地质学研究。E-mail: jgcai@tongji.edu.cn。
自然界中有机质分布广泛、类型多样且性质各异,不论在水体还是在沉积物(岩)中都扮演着重要的角色。因此,探讨各类型有机质聚集和沉积过程的差异性,对深化认识有机质沉积特征和演化规律具有重要的意义。生物体在生长过程中产生了许多有机质,包括生物体自身、生物残体、动物排泄物以及生物分泌的有机分子等,各类型有机质的性质差异极大。由于研究方法的不同,可将海洋中有机质划分为颗粒有机质( particulate organic matter,POM)和溶解有机质( dissolved organic matter,DOM),而DOM又可划分为胶体有机质( colloidal organic matter, COM)和真溶解有机质;从聚合体形式上看,又有海雪和悬浮体等存在形式。进一步分析发现有机质的形态包括生物体、生物残体、排泄物和有机质聚合体等,并且各类型有机质与无机矿物以不同的形式共存。此外,DOM与POM间存在一定的转化关系。这些性质和特征架起了不同类型有机质间聚集和沉积的桥梁。在沉积物(岩)中通过粒度或密度分级分离及孢粉相分析,也发现不同类型的有机质,如生物体、生物残体和无定形等,它们常与特定的无机矿物共生,如无定形多富集在黏粒级颗粒中,而生物体和生物残体多富集在粗颗粒中。进一步溯源发现沉积物(岩)中的各类有机质与生物有机质或海洋中有机质的类型极具相似性,这些特征展现各类型有机质在沉积过程中的差异性。综合有机质的形态、性质以及与矿物共生关系,认为有机质可通过机械沉积、化学沉积和生物沉积等3种不同的方式沉积保存,其中机械沉积的有机质以惰性的生物残体为主,化学沉积的有机质以活性极强的无定形为主,生物沉积有机质以活性较强的微生物和粘附物为主。有机质沉积方式及有机质特征、矿物—有机质间关系和保存条件的差异,决定了有机质的演化命运的不同,进而对有机质生烃以及碳循环产生重要的影响,因而应引起人们的高度关注。
About the first author Cai Jin-Gong,male,born in 1961,graduated and obtained his Bachelor's degree and Ph.D. degree from Nanjing University and Tongji University in 1983 and 2003,respectively. Now he is the professor and doctoral supervisor of the School of Ocean and Earth Science of Tongji University. He is currently engaged in sedimentology and petroleum geology. E-mail: jgcai@tongji.edu.cn.
Organic matter of various types and different properties is widely distributed in nature,and it plays an important role in both water body and sediments. Therefore,exploring its accumulation and depositional processes is significant for better understanding the sedimentation characteristics and evolution laws of organic matter. Organisms produce abundant organic matter during the process of growth,including the bodies,remains,excreta of organisms and molecular scale of their secretion. The properties of different types of organic matters vary greatly. Organic matter can be divided into particulate organic matter(POM)and dissolved organic matter(DOM)according to the size,and DOM can be further divided into colloidal organic matter(COM)and true DOM. From the point view of aggregation form,there are marine snow and suspension matter. Further analysis shows that morphology of organic matter includes organisms' body,biological residues,excreta and organic matter aggregates in water body,and various types of organic matter can co-exist with inorganic minerals in different forms. In addition,DOM and POM can transform mutually in appropriate conditions. These properties and characteristics bridge the accumulation and deposition of different types of organic matter. It is also found that different types of organic matter,such as organisms,biological residues and amorphous organic matter,are often associated with specific inorganic minerals,such as amorphous mostly concentrated at clay particles,whereas organisms and biological residues mostly concentrate in coarse ones. Further study reveals that the organic matter of sediments is similar to that of water body or organism. The variation of different kinds of organic matter reflects that they deposited via different processes. The mechanically deposited organic matter is mainly composed of inert phytoclasts,the chemically deposited organic matter is mainly composed of amorphous organic matter,and the biologically deposited organic matter is mainly composed of microorganisms and adherents. Both the differences in organic matter deposition and organic matter characteristics and the mineral-organic matter and storage conditions determine the evolutionary fate of organic matter,which has an important impact on organic hydrocarbon generation and carbon cycle,and thus should be highly concerned.
自然界中的有机质类型多样, 性质各异, 不论在水体还是在沉积物(岩)中都扮演着重要的角色。大量的研究都围绕着有机质的构成、分类及其与矿物的结合特征开展(Bodineau et al., 1998; Weller et al., 2007; Hare et al., 2014; Crawford and Liber, 2015)。然而, 关于有机质的研究, 还有许多悬而未决的问题。首先, 有机质在沉积物中的保存包括降解缩聚、自然硫化、黏土保护和选择性保存等多种方式(Nissenbaum and Kaplan, 1972; Mayer et al., 1988; Damst'E et al., 1989; Schulz et al., 1994; 樊馥等, 2011b), 这些方式直接影响了碳循环过程。众所周知, 有机质保存的多样性主要由其性质决定(Hedges and Keil, 1995; Keil and Mayer, 2014), 然而有机质从形成到保存经历了怎样的命运, 却一直困扰着研究人员。其次, 对富有机质沉积岩成因的研究一直处于争论阶段, 有机质往往被作为独立的部分进行考虑(刘传联等, 2001; Loucks and Ruppel, 2007; 王冠民, 2012; Chen et al., 2016), 但实际研究却发现有机质在沉积岩中与矿物关系密切且形态具有多样性(Love et al., 1984; Al-Otoom, 2008; Wang et al., 2010; 张晶巧等, 2013; Zeng et al., 2018), 因此, 有机质以何种方式沉积成了需要关注的重点。此外, 烃源岩中不同赋存态有机质具有差异生烃的特点(Li et al., 2017)。有机质沉积研究为解开差异生烃之谜提供了新思路。因此, 关注有机质从水体到沉积物的演化过程是极其重要的。
目前, 对于有机质沉积的研究是深入但分散的, 大量研究分别从生物学、海洋学和沉积物(岩)等方面, 用不同的测试手段建立了多种有机质分类标准, 如生物化学关注各类有机质的分子特征(刘诚刚等, 2010; Hansell and Carlson, 2014); 海洋研究中关注溶解有机质和颗粒态有机质的关系(Verdugo et al., 2004; 何伟等, 2016); 沉积物中有机质研究关注有机— 无机组合特征及各来源有机质的形貌学规律(Bergamaschi et al., 1997; Singh and Mahesh, 2015)。如何综合这些研究成果, 建立一个从生物学到沉积物的完整序列, 弄清各类有机质聚集和沉积过程的差异, 阐明沉积过程中有机质和矿物之间的关系, 是深化认识有机质沉积特征和演化规律的难点。
为此, 文中收集海洋、沉积物和沉积岩中有机质的类型和属性以及有机质聚集和沉积作用的特征等研究成果, 综合有机质沉积作用方式和差异以及在有机质生烃和有机碳循环中非一致性的演化特征, 明确有机质沉积在地质过程中的意义。
自然界的有机质都来源于生物体, 而生物体及其降解状态的不同是导致有机质性质差异的重要因素。生物由于生活环境和方式及生物链的传递和分化等差异, 从陆地的动植物、微生物, 到海洋的浮游生物、游泳生物和底栖生物等(Huston and Huston, 1994), 构筑了一副生物圈的完整画面。
陆生植物分为苔藓植物、蕨类植物和种子植物, 它们在生长过程中释放的大量分泌物, 植物凋落物等进入土壤中, 在微生物作用下降解和转化(张勇等, 2005), 一部分能够保留其生物形态(角质和木质等), 一部分则在土壤中形成腐殖质(Poirier et al., 2003; Mikutta et al., 2006), 以无定形的形态保存。植物死亡后, 茎部部分经历机械破碎等过程, 木质部分保存下来, 在遇火灾时会被焚烧成为碳质, 在机械冲刷下进入河流, 并随其输送到海洋中(Hedges et al., 1997)。这些产物木质素含量高, 抗降解性强, 因而能够大量保存下来。海洋中主要的陆源颗粒有机质便是木质和丝炭。植物的孢子和花粉在河流或风的输送下, 也会进入海洋中, 且分布广泛(Simoneit, 2006)。因此, 从有机质形态上看, 进入海洋的陆源有机质便有木质、丝炭、角质、孢粉、陆源可溶有机质等不同的类型。
浮游植物是海洋中的生产者, 浮游动物、游泳动物和底栖动物作为消费者, 一层一层进行碳的传递。浮游植物细胞直接沉降, 尤其是大细胞, 群体的和链状的浮游植物死亡后快速沉降至海底(Billett et al., 1993; 孙军等, 2016)。海洋中还有大型植物, 如海草和大型藻类等, 其死亡后的残片或被动物利用, 或原地沉积, 或运移至更深海域保存(Smith, 1981)。浮游动物、游泳动物和底栖动物在生长过程中, 除了有大量的分泌物外, 还不断排出粪便, 如粪球粒等。这些粪球粒除了被食粪者破碎和利用之外, 能够向海洋深部运输(Turner, 2015)。这些动物死亡后, 有机质的部分大多会被降解矿化或被其他生物利用, 部分生物内衬和残体能够保存下来(Hottinger and Dreher, 1974; Turner, 2015)。此外, 浮游生物和微生物会释放含多糖的黏性透明胞外聚合物, 能够将浮游植物残体、破碎的粪球、藻类残体、有机质屑、无机矿物等聚合起来一同沉降(Dash et al., 2012)。
海洋中还有底栖生物席, 其绝大多数是菌藻席。类型不同的生物席, 发育的环境也不同。在光合作用带发育的是蓝细菌席, 而在光合作用带之下则发育化能自养或异养菌席(张庆珍等, 2017); 根据氧的含量, 在氧化带发育好氧菌席, 在还原带发育厌氧菌席(梅冥相, 2011)。当发育的环境发生改变时, 可移动的微生物席便会转移到可生存处(梅冥相, 2014), 如在沉积作用事件的间隙, 丝状蓝细菌会滑动到新的沉积面上, 形成新的微生物席(Gerdes and Klenke, 2007)。被微生物席矿化的泥、不能移动的细胞和鞘, 便会被埋藏下来(梅冥相, 2014)。此外, 微生物席分泌的细胞外聚合物(EPS), 可将有机或无机悬浮颗粒粘附, 增加微生物生长表面的类群、结构和新陈代谢的多样性。因此, 微生物的死亡残体、释放的代谢产物及其圈捕的有机质颗粒便是微生物席向沉积物贡献的有机质类型。
从生物化学的角度看, 有机质由蛋白质、碳水化合物、类脂类、木质素和丹宁组成, 且各种有机化合物的性质各异。蛋白质是高分子有机质, 其主要来源于动植物和微生物残体的降解、微生物分泌的胞外酶等(梁小兵等, 2001), 其很容易被生物利用而降解。被无机矿物以化学键连接或被高度脂肪化分子封装的氨基酸, 则能够被保护下来(Collins et al., 1991; Knicker and Hatcher, 1997)。糖类分为结构多糖(如纤维素)和储能多糖(如淀粉)。在没有纤维素降解菌的情况下, 纤维素不易被生物利用而降解; 而储能多糖则不同, 很容易被生物利用, 因而在沉积物中很少发现(Hernes et al., 1996)。类脂类的情况则较为复杂, 其种类多样, 包括各种酮、醛、酸及其衍生物等, 生物化学性质不一。有机质自身稳定性由高到低依次为链烷烃、脂肪酮、甾醇、脂肪酸、烯酮、烯酸, 长链大于短链, 饱和有机质大于未饱和有机质(Wakeham and Canuel, 2006)。从有机质的保护上看, 被无机矿物吸附及大分子有机质封装的有机质能够较好地保存下来(Collins et al., 1991; Knicker and Hatcher, 1997)。木质素和丹宁不溶于水, 其分子结构为网状的大分子结构, 较为稳定, 具有较强的抗降解性, 很少有水生生物和微生物能够利用其作为碳的来源, 因而能够很好地保存于地层中(Wakeham and Canuel, 2006)。以上可以看出, 不同有机质的化学组成和结构不同导致的性质差异, 使其在沉积和埋藏过程中经历不同的生物化学过程。
综上所述, 自然界中有不同类型的生物体, 如动物、植物和微生物等, 也有生物生长过程中的分泌物和排泄物, 以及生物体死亡后的碎片(或残片)和分解或降解的分子化合物等, 这造就自然界中有机质的多样性特征。此外, 自然环境的差异会形成陆源及海源生物群落, 展现了有机质来源的差异特征。由此看来, 有机质虽然源自于陆源和海源等不同环境的生物体, 但生物体在生长、死亡甚至分解和降解的过程中会有不同的有机质样式呈现, 将会造成自然界中有机质的来源、特征和性质的多样性和差异性, 这必将导致有机质聚集和沉积过程的复杂性, 也是有机质聚集和沉积作用差异的重要根源。
海洋中有机质依据粒径大小划分为溶解有机质(dissolved organic matter, DOM)和颗粒有机质(particulate organic matter, POM), 其中溶解有机质可进一步细分出胶体有机质(colloidal organic matter, COM)和真溶解有机质(low molecular weight dissolved organic matter, LMW DOM)。
溶解有机质DOM是海水中通过0.45 μ m滤膜的有机质, 是海洋中最大的有机碳储库, 其总量估计在700 Gt左右(Hedges et al., 1997), 约占海洋中有机碳的90%(Gan et al., 2016)。DOM来源于生物的分泌物和残体碎屑等(何伟等, 2016), 从海洋化学的角度它包含各种类别的细胞生化物质, 如碳水化合物(单糖、寡糖和多糖)、含氮化合物(氨基酸、溶解蛋白质和多肽)、脂类和脂肪酸、有机酸(乙醇酸和三羧酸)、核苷酸、植物色素(叶绿素和类胡萝卜素)、维生素、生长抑制成分(毒素)以及海洋腐殖质等(刘诚刚等, 2010; Hansell and Carlson, 2014)。其中, 腐殖质是构成溶解有机质的重要类型。它是海洋生物的代谢产物和死亡后残留物的简单成分或分解作用的中间产物(如糖类、氨基酸、酚类和类脂物等), 在微生物和其他自然条件作用下, 经过化学和生物化学的合成作用形成的一类性质稳定、不易分解、分子量较高、结构复杂的有机物(Aiken, 1985; Kang et al., 2002)。海洋腐殖质具有分子量较大、结构复杂和性质稳定的特点, 能抵抗细菌进攻和化学氧化作用, 不易被微生物降解或化学分解。海洋腐殖质占溶解有机质总量的40%~80%, 深层水中的含量高于表层(Kitis et al., 2001)。此外, 腐殖质的分子结构中包含多种官能团, 如羧基、羰基、羟基、酚羟基、乙酰基、氨基和醌基等。由于含有多种官能团, 与痕量金属元素络合或螯合(Smith and Kramer, 2000; 李巍巍, 2017), 与矿物之间也有密切关系(吴宏海等, 2003)。溶解有机质的丰富类型决定了其转化和迁移方式的多样性。
胶体有机质(COM)是通过切向超滤等方式截留的相对分子质量超过1000的有机质, 相当于这部分有机质未通过孔径约为1 nm滤膜(Wilkinson et al., 1999; 王江涛等, 2000)。COM是DOM中重要的组成部分, 占据DOM的50%~70%(Aluwihare et al., 1997), 主要成分包括糖类、含氮化合物和脂质等(Zhu et al., 2017)。胶体相介于有机质溶解态和颗粒态之间, 且其表面积大, 在水体中动力活性强, 可通过絮凝— 解絮凝作用作为两者相互转化的重要介质(谭丽菊和王江涛, 1999; 蔡进功等, 2005)。真溶解有机质粒径小于1 nm, 在藻类勃发时, 由于藻类的释放, 其在水体中的含量增长快, 可占据水体总有机碳(TOC)的40%~50%(Gobler and Sañ udo-Wilhelmy, 2003)。真溶解有机质包括单糖、低分子量多糖、低分子量氨基酸和有机质微粒等(Zhu et al., 2017)。研究发现, 真溶解有机质在水体中滞留的时间比COM长(Gobler and Sañ udo-Wilhelmy, 2003), 这是由于有机质降解后留下的惰性部分保存于水体中, 部分可达千年之久(Giorgio and Duarte, 2002)。
POM是未通过0.45 μ m滤膜的颗粒有机质, 其包括陆源输入的高等植物碎屑, 也包括海洋的动植物及其残体、动物分泌的粪球粒, 还包括有机质絮凝体等, 类型非常丰富。陆源植物碎屑随着河流搬运入海, 包括木质和丝碳等高等植物碎屑, 其性质稳定而不易被生物利用(Hedges et al., 1997); 海洋生物体包括浮游动植物、游泳动物和底栖生物, 它们通过食物链将碳固定并转移至海底; 动植物残体包括未被分解的动物内衬和藻残片等; 粪球粒是动物的分泌物, 易于破碎, 且可被食粪生物利用; 有机质还能通过絮凝作用形成大颗粒的有机质聚合体而沉降。POM是有机质沉降的主力军, 在沉降过程中不断地被食用以及降解、矿化等作用, 最终进入到沉积物保存的量是很少的(Hedges et al., 1997)。
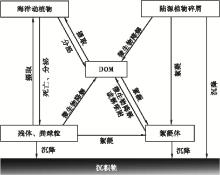
水体中的有机质类型是多样的, 且在一定条件下, DOM与POM可以相互转化(图1), 具体可分为非生物过程、生物过程及两者的复合过程(何伟等, 2016)。溶解有机质和胶体有机质具有溶解性、电离性、胶体性和较大的比表面积等化学活性, 研究也发现COM等凝胶相是溶解有机质转化为颗粒有机质的中间过程(Verdugo et al., 2004); 首先, 异养生物在生长过程中需食用DOM等, 而后转化为其自身组成部分和排出的粪球粒, 即通过食物链的转化(Turner, 2015); 其次, 透明胞外聚合物(transparent exopolymer production, TEP)可将DOM及细颗粒的有机质聚合起来, 形成粒径更大的絮凝体(Macquaker et al., 2010); 第三, 黏土矿物具有较大的比表面积, 可吸附DOM, 并与其他有机颗粒形成絮凝体(蔡进功等, 2005); 第四, POM也可转化为DOM。微生物对POM作用后, POM释放小分子有机质, 生物自身释放可溶的代谢产物(Dyda et al., 2009); 动物食用POM时的物理性破碎作用, 絮凝体在合适外部环境时的解吸附作用等(Aochi and Farmer, 2011), 都可使POM转化为DOM。DOM和POM的相互转化使得其在水体中保持一个动态的平衡。
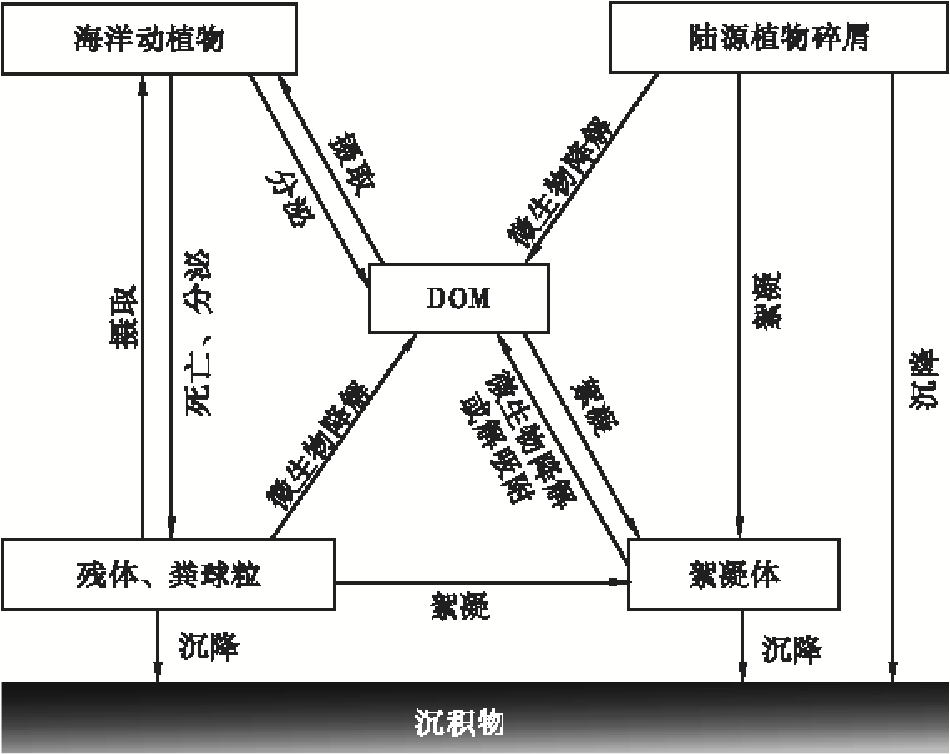 | 图1 海洋中有机质的相互转化 (据蔡进功等, 2005; Turner, 2015)Fig.1 Mutual transformation of organic matter in the ocean (after Cai et al., 2005; Turner, 2015) |
综上所述, 各种形态的有机质在海洋中都有分布, 从分子级的有机化合物到有机质的聚合体或碎屑体等多种类型的特征, 而且可能同时存在生物有机质、分解后的中间产物— — 溶解有机质以及有机质相互结合而成的胶体有机质和聚合有机质等, 不同类型之间相互关联、相互转化, 达到一定的动态平衡, 既体现海洋中有机质的多样性和复杂性, 也为有机质聚集和沉积作用提供了多样性的空间。
海雪是一种颗粒较大的有机质聚合体, 其粒径大于500 μ m, 因其在深海中观察时其具有白色疏松的特点而得名, 在近岸和远海中均有发现(Dash et al., 2012; Turner, 2015)。海雪的形成受控于颗粒丰度、粒径、碰撞及粘结剂(Jackson, 2001)。因而, 含多糖而具有黏性的TEP在海雪的形成过程中便起到重要的作用。海雪中含有的有机质来源多样, 类型丰富, 包括海洋藻类残体、矿物颗粒、高等植物碎屑、生物住囊、粪球粒和微生物等(Shanks and Trent, 1979)。在不同区域, 其主要类型并不一致(Turner, 2015)。因此, 根据主要有机质类型差异, 将海雪分为浮游植物型、住囊型和粪球粒型等(Dash et al., 2012)。
2.2.1 浮游植物型
浮游植物型海雪主要由浮游植物组成。在浮游植物繁盛时, 水体或有机颗粒上的微生物释放TEP, 增加颗粒黏性, 并将浮游植物与其他有机无机颗粒聚合起来而稳定存在(Decho, 1990; Heissenberger and Herndl, 1994)。微生物在此类型海雪形成过程中起着非常重要的作用(Grossart et al., 2006)。在美国圣胡安岛的密度不连续的沉积物中发现硅藻型海雪, 其硅藻含量高, 出现不同的厚度和含量, 显示出其季节性或年度变化的特征(Alldredge et al., 2002)。
2.2.2 住囊型
住囊型海雪是动物住囊占据主要成分的海雪。住囊是住囊虫产生的壳体, 多孔且具有生物黏液(Ohtsuka et al., 1993; 李开枝等, 2010)。尾海鞘纲动物在其生命历程中会产生50个以上的废弃住囊, 这些住囊是浮游植物、细菌、鞭毛虫、粪球粒、矿物颗粒和其他颗粒的重要集中场所(Alldredge and Gotschalk, 1988), 住囊上各有机质类型的含量较周围水体高许多(Steinberg, 1994), 比如部分海域硅藻和粪球粒含量分别是其在周围水体含量的1000倍和100 000倍(Hansen et al., 1996)。Alldredge(2005)在部分海域和沿岸区域的POM中, 尾海鞘纲动物住囊占据12%~83%; 在波罗的海的薄地层中也发现挠足类生物住囊是POC的主要部分(Mö ller et al., 2012)。这些住囊带着其上附着的有机和无机颗粒一起快速沉降。
2.2.3 粪球粒型
部分海洋区域的生物繁盛, 因而生物排出的粪球粒也丰富, 该区域海雪的主要类型便是粪球粒。在远洋及深海区域, 粪球粒通常都经历食粪生物利用和降解、机械破碎等过程, 使得该区域的粪球粒颗粒细小(Small et al., 1979; Lampitt et al., 1990)。在亚得里亚海北部收集到的海雪中, 粪球粒含量为31~353 000粒/L, 其中细微颗粒的粪球(< 105 μ m3)占海雪的68%以上(Bochdansky and Herndl, 1992)。大的粪球粒自由沉降, 破碎细小的粪球粒被TEP粘结或者黏土矿物吸附, 并与其他有机颗粒集聚沉降(Turner, 2015)。
海雪分布广泛, 在各个海域都有发现, 由于所含有的主要类型有机质差异, 除了上述3种之外, 还有其他类型, 包括植物碎屑型等(Dash et al., 2012)。此外, 其吸附性、微生物含量和动物食用等存在差异, 因而其沉降速率、降解作用和被食用等是存在差异的(Dash et al., 2012), 这对有机质的沉降和保存具有重要影响。
海洋悬浮物包括陆源输入颗粒物和海洋自生颗粒物及其混合体, 其不断向海底沉降, 形成沉积物; 同时在一定条件下也会再悬浮而进入水体, 形成新的悬浮物(Che et al., 2003)。海洋中悬浮的颗粒物, 如浮游动植物、细菌、动植物残体和无机碎屑等, 在生物化学等作用下形成絮凝体。此外, 这些悬浮物含有多种不同类型的有机质及无机颗粒, 且区域差异明显(李洪波和邵魁双, 2011)。
在太平洋西北海域, 通过沉积物采集器获取的悬浮物中, 含有丰富的硅藻、粪球粒、碎屑矿物及细颗粒的絮凝体, 颗粒物粒径大多都小于50 μ m(Hung et al., 2012)。这是开放海域悬浮物粒径特征。而在太平洋西南海域, 透明无定形的有机质聚合体是悬浮物有机质的主要部分; 不同深度上, 硅藻、粪球粒和有孔虫等的百分含量此消彼长, (Nodder, 1997)。方建勇等(2015)对南极普里兹湾海域研究发现, 悬浮物中含有生物颗粒包括硅藻、微型藻、有机包膜、矿物颗粒和混杂集合体。其中, 有机包膜是能够捕获矿物颗粒和生物碎屑的膜状胶体物质, 混杂集合体主要是由生物碎屑、矿物颗粒在布朗运动和生物黏液作用下形成的集合体。生物颗粒的含量在70%~90%, 粒径在1~400 μ m之间, 由于颗粒物质来源、海洋环流和海底地形的影响, 在不同区域分布不均匀。在动物繁盛的近海区域, 悬浮物中的有机质在类型、含量和粒径上与南极不相同。研究发现, 台湾岛及其邻近海域的悬浮物中, 矿物组分以石英、长石等轻矿物为主, 有机质组分则包括生物住囊、粪球粒、硅藻、微型藻、有机包膜和混杂集合体; 生物颗粒含量在20%~85%, 粒径介于1~100 μ m之间, 且不同深度和区域上, 分布不同(方建勇等, 2010)。由此可知, 悬浮物由有机颗粒和无机矿物组成, 其含量和粒径在不同区域和深度上是有明显差异的, 除了源区影响, 还受海洋环流的再分配、沉降过程中的破碎以及絮凝等物理化学作用的控制。
海洋中的有机质类型丰富多样, 性质各异, 因而其形态、与无机矿物的结合方式以及沉降与保存等都有明显的差别。从不同的角度来看, 可以全面了解有机质在水体中聚集、搬运和沉降等各个阶段的存在状态, 这对其沉积后的保存与演化有重要的意义。
沉积物和沉积岩是有机质最终的富集场所。水体中的有机质在沉积过程中消耗巨大, 仅有少量的有机质能够保存下来(Hedges and Keil, 1995; Hedges et al., 1997; Zonneveld et al., 2010), 而且这些有机质在埋藏过程中会发生降解缩聚、选择性保存、自然硫化和黏土保护等作用得以稳定存在于沉积物中(Hedges and Keil, 1995; 樊馥等, 2011b)。有机质在沉积物中的演化导致其特征与沉积时期性质往往大不相同。然而, 沉积物中残存的有机质仍然保留了大量原始有机质的信息, 通过这些信息(如有机质丰度、形态特征与化学性质等)能够准确还原有机质的来源与沉积历史。在沉积物和沉积岩的分析中, 常常关注的是沉积物中有机质的丰度, 其既反映了水体的初级生产力(Vetö et al., 1997), 也反映了有机质的保存情况(Zonneveld et al., 2010)。然而, 有机质丰度只能体现沉积物中有基质的整体特征, 而不能了解不同类型有机质各自的演化路径。因此, 大量研究更关注有机质屑的特征和性质, 由此也衍生出了多种的有机质分离与分类方案, 用于恢复沉积物中原始有机质的组成和特征。下面对它们分别介绍。
矿物和有机质的粒径与自身特性有很大的差异, 这也导致在沉积过程中不同粒径矿物和有机质之间的不同结合关系(Thompson and Eglinton, 1978; Oades, 1989; Christensen, 1992; Hedges and Oades, 1997)。基于这种原因, 粒度分离是研究沉积物中有机质特征的常用方法。沉积物的颗粒分级常采用筛析法、沉降法和SPLITT分级等方案进行粒度分离(冯晓萍和蔡进功, 2010)。在研究沉积物中有机质分布特征时, 会根据样品的特征, 如黏土的含量、沉积物的胶结程度等因素(陆现彩等, 1999; 王德杰等, 2003), 选择合理的分析方法进行粒度分离。目前, 沉积物的粒度主要以250 μ m、63 μ m和2 μ m为界, 划分出砂、粉砂和黏粒级组分, 并对各组分进行更细致的粒度分级, 用以揭示每种粒度组分中有机质的特征(Keil et al., 1998; 冯晓萍和蔡进功, 2010)。
研究发现, 有机质在沉积物各粒级中具有差异性分配的特征。这种差异性分配首先表现在不同粒度组分的有机质对总有机碳含量的贡献率上, 其中, 粉砂级和黏粒级组分(< 63 μ m)中赋存的有机质对沉积物TOC的贡献率达到了80%以上, 而保存在砂粒级沉积物中的有机质贡献率则较低(Stemmer et al., 1998)。这说明了在沉积过程中, 细粒组分中的有机质沉积量更大。
同时, 有机质的差异性分配也表现在有机质类型与粒度组分关系上。蔡进功等(2006, 2011)对浙闽沿岸、东海和长江口区域的表层沉积物进行粒度分级后发现, 表层沉积物中不同粒级组分中有机质赋存态存在显著差别(冯晓萍和蔡进功, 2010)。其中, 粗粒级组分富集木质和黑色不透明团块等有机屑; 小于2 μ m的黏粒级组分富集无定形有机质。Keil 等(1994b)对华盛顿沿岸地区沉积有机质特征进行粒度分离分析并进行碳同位素分析发现, 大于63 μ m的砂粒级沉积物组分中, 95%以上的有机质都来自于外源输入的维管植物; 而黏粒级组分(< 2 μ m)的沉积物主要富集水体自生的有机质。卢龙飞等(2011)分析东海陆架泥质区表层沉积物组成与分布特征发现, 超细颗粒物中脂肪酸以海洋内源藻类来源为主(> 70%)。各粒级中有机质性质和形态的差异说明了沉积过程中有机质和矿物的关系, 也说明了有机质沉积方式的差异。此外, 各粒级中有机质的分布和沉积区域有一定关系。Keil 等(1994a)、Hedges 和 Keil(1995)通过对比表层沉积物各粒级组分内有机质特征后发现, 海洋陆架区域的沉积物明显更加富集砂粒级的陆源植物碎屑颗粒, 而陆坡区域的有机质则富集与黏土矿物紧密联系的复合有机质。大量研究发现, 不同粒级沉积物有机质保存规律的巨大差异, 都预示了不同性质的有机质在水体中就发生了差异性沉积。
自然界中, 矿物密度约为2.6 g/cm3, 而纯有机质颗粒的密度一般在1.0 g/cm3左右, 二者结合的比例不同导致沉积物密度的差异(熊毅, 1985)。因此, 在分析沉积物有机质的特征研究中, 常采用密度分离方法对沉积物进行分级。密度分离一般以1.6 g/cm3和2.2 g/cm3为界划分密度等级, 用以分析不同类型有机质的差异(樊馥等, 2011a)。
同粒度分级一样, 大量研究通过对口岸、表层海洋和湖泊的表层沉积物以及沉积岩进行颗粒分密度检测发现, 各密度组分沉积物中的有机质对总有机碳的贡献率差异很大, 其中, 高密度组分沉积物中的有机质对TOC的贡献率达到90%, 而低密度组分中的有机质则贡献了不足10%(Arnarson and Keil, 2001; 樊馥等, 2011a)。对总有机质贡献率的差异说明了高密度组分中的有机质沉积量更大。除了总有机质的贡献率, 各密度组分的有机质类型差别也很大。Wang 等(2015)对长江口的沉积有机质进行密度分离后发现, 低密度组分的有机质主要为外源输入的有机质, 而高密度组分则主要为浮游藻类等水体自生的有机质。Dickens 等(2004, 2006)对华盛顿湖的沉积物进行密度分离发现, 具有海源特征的有机质富集在细粒级、高密度的组分中, 与矿物的关系非常密切。另一部分具有陆源特征的有机质富集在粗粒级、低密度组分中, 与矿物的关系并不密切。沉积过程中矿物具有保护作用。叶文青等(2009)尝试通过密度分离的方法分离沉积岩中不同密度的有机质, 分析发现岩石中大量有机质都与黏土矿物等无机物共生且紧密结合, 只有少量的生物碎屑类有机质往往单独保存, 不易与无机矿物结合。Bock 和 Mayer(2000)分离出密度为2.0~2.3 g/cm3级别的沉积物后发现, 这一密度范围的沉积矿物几乎全都是黏土矿物, 几乎所有的有机质都以有机— 黏土复合体的形式保存于沉积物中。总的来说, 轻组分有机质来源于陆生植物等颗粒, 重组分有机质为水体自生且与矿物关系密切, 这些特征展现了有机质沉积的差异性。
粒度分离和密度分离往往联合使用。结合前文分析可知, 水体自生有机质通常具有细粒径、高密度特征, 陆源植物则具有粗粒径、低密度特征。前人研究分析发现, 富集陆源植物的组分中木质素的含量很高, 木质素是维管植物所特有的高分子聚合物, 往往和纤维素共生(Burdige, 2007; Keil et al., 1998)。富集自生有机质的组分中糖类、醇类、氨基酸和脂类等含量较高(段毅等, 1997)。对表层沉积物的研究也发现, 陆源高等植物碎片具有高相对分子质量的烃类(低C21-/C22+), 而低等水生植物则具有中等相对分子质量的正构烷烃(高C21-/C22+)(蔡进功等, 2006; 郭志刚等, 2001)。各类有机质独特的化学性质都预示了在沉积过程中有机质与矿物的不同关系及独特的沉积方式。
研究发现, 黏土矿物富集的沉积物中有机质往往也十分丰富(Keil et al., 1994a; Bergamaschi et al., 1997; Kaiser and Guggenberger, 2000; Gelaw et al., 2015), 这说明沉积物中的一些有机质与矿物关系密切, 在前文论述中细颗粒或高密度组分中有机质富集的特征也证实了这一点(Salmon et al., 2000; Kennedy et al., 2002)。前人研究发现, 沉积物或岩石中矿物结合态有机质普遍存在(Schulten et al., 1996; 蔡进功, 2004; 蔡进功等, 2007; Rahman et al., 2017), 最常见的矿物是黏土矿物。黏土矿物是层状硅酸盐矿物, 其自身具有粒径细、具荷电和比表面大的特征(Bergaya et al., 2011)。这种特征决定了矿物对其他物质具有更快的吸附速率、更大的吸附容量和更强的吸附稳定性(Weaver, 1989; Huang et al., 2014)。黏土矿物的这种特性是沉积时期有机质和矿物结合的必要条件。
在沉积物(岩)中, 黏粒级组分的元素地球化学特征和其他粒级组分特征有非常明显的差异。Xiong等(2010)对土壤和沉积物的研究发现, 黏粒级组分更富集Cr、Cu、Zn、Co、Ni和V等微量元素(Acosta et al., 2011), 而这些微量元素也是生物生长所必备的营养元素(Hinds and Lowe, 1980)。这些发现说明沉积时期黏粒级组分为有机质的结合提供了物质保障, 同时也为微生物在沉积过程中和黏粒级组分结合提供了有力证据。
沉积物(岩)中黏粒级组分的有机质主要来源于溶解有机质和藻类等微生物(卢龙飞等, 2011), 这说明沉积过程中有机质和矿物就已经有了密切联系(Turner, 2015)。矿物和有机质的结合包括了配位基交换、离子交换、阳离子桥联、范德华力、氢键联结以及聚合作用等方式(Keil and Mayer, 2014)。卢龙飞等(2013)在对结合态有机质分析研究中发现, 有机质能够以物理吸附和化学吸附2种方式保存于矿物的外表面或内表面。有机质以多种方式与矿物结合, 包括以物理方式吸附于矿物表面以及以化学结合方式保存于矿物边缘破键处或矿物层间(Zhu et al., 2016, 2018)。
通过以上分析可知, 自然界中无机矿物(黏土矿物等)可以吸附有机质、特别是可溶有机质等, 进而聚集和沉积, 这将对有机质的沉积和保存意义巨大。
生物成因的岩石主要包括非钙化微生物和矿化微生物构成的岩石, 其中非钙化微生物岩包括叠层石、纹层石、核型石、凝块石、均一石和模铸岩等; 而矿化微生物岩则包括骨架微生物岩、微生物粘结岩、生物颗粒岩和硅藻土等(吴亚生等, 2018)。总的来说, 这些岩石都是生物体经过各种矿化作用而形成的(Characklis and Wilderer, 1989; Neu and Marshall, 1990; 梅冥相, 2014)。在生物或有机矿化的过程中, 有机质往往被矿物晶体闭塞包裹并保存起来(Keil and Mayer, 2014)。这类有机质的形成伴随着矿物的产生, 反映了与其他有机质截然不同的沉积方式。
综合沉积物(岩)中有机质分离检测结果可以发现, 沉积物(岩)中的有机质不是以单一方式出现, 粗粒级和黏粒级组分分别富集了性质各异的有机质, 说明沉积物中有机质的聚集和沉积方式都可能存在差异; 细颗粒富集的有机质以无定形或可溶有机质为主, 展现出矿物与有机质间的相互作用关系的存在; 生物体在生长过程中经过矿化作用形成生物岩, 表明生物可通过自身作用聚集沉积而形成沉积物(岩); 当把沉积物(岩)中无机矿物除去后发现, 有机质的类型极其丰富, 并与生物体分解或降解的产物和类型是极其相似的, 展现了来源于生物体的各类有机质都可以通过不同的方式而沉积。
在生物体、水体和沉积物(岩)等领域的研究成果, 充分展现了有机质的多样性特征。孢粉相分析是去除沉积物中的矿物组分后, 对残留的颗粒有机质进行形貌学分析, 直观地判识有机质的形态和结构, 这可以将生物体、水体和沉积物(岩)的有机质进行关联。孢粉相分析被广泛应用于沉积有机质溯源和古环境分析研究(Batten, 1996; Sebag et al., 2006; Schneebeli-Hermann et al., 2012; 张晶巧等, 2013)。目前孢粉相分析普遍使用Batten(1996)和Tyson(1995)等人的分类方案, 将沉积物中的颗粒有机质划分为无定形、孢型有机质和结构有机质。湖相泥页岩作为典型的富有机质沉积物, 既包含了类型丰富的有机质, 也包含了碎屑、黏土和碳酸盐等多种矿物类型, 为全面认识有机质的类型提供了良好的基础条件(曾翔等, 2017)。因此, 可通过湖相泥页岩的孢粉相分析为例, 详细阐明从生物到沉积物中有机质的联系。
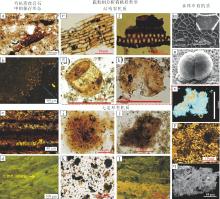
以渤海湾盆地东营凹陷沙河街组湖相泥页岩为例, 前人镜下观察发现(Zeng et al., 2018), 不同类型的有机质在岩石中的保存方式差异很大(图2-a至2-d)。部分植物残片和孢型有机质能够形态完好地保存于岩石中, 从镜下观察和矿物与有机质关系都发现, 这些有机质与矿物没有明显的共生关系(图2-a, 2-b)(Zeng et al., 2018)。岩石中另一部分有机质则以富有机质纹层或特殊构造形态保存(图2-c, 2-d), 研究发现, 这些构造中矿物和有机质具有明显相关性, 说明这类有机质与矿物的关系密切(张水昌等, 2007; 王冠民, 2012; Liu and Wang, 2013)。从孢粉相分析来看, 岩石中保存的有机质都能在水体中找到对应的类型, 结构有机质由于保留了原始形貌特征, 而被确认为来源于高等植物、沉水植物和花粉孢子等(图2-e至2-h); 无定形有机质没有固定的形貌特征, 但前人从有机质残留的信息分析确定了无定形有机质主要来源于海雪、藻类聚合体以及底栖微生物席等(图2-i至2-l)(李国山等, 2014)。结合前文研究可以确定, 沉积物中结构有机质性质稳定, 从形成到最终保存都比较稳定, 以单颗粒形态沉积; 而无定形有机质的最终保存, 经历了从溶解有机质到颗粒有机质的聚合或生物矿化过程, 整个阶段与矿物关系密切。通过对泥页岩中有机质的溯源, 可以确定沉积岩中有机质的多样性可追溯到沉积时期, 其与矿物的共生关系差异性, 展现了不同的沉积方式。
 | 图2 湖相泥页岩中有机质特征及溯源(据Tyson, 1995; Batten, 1996; 张晶巧等, 2013; Zeng et al., 2018) a— 泥页岩中保存的有机质残片; b— 泥页岩中保存的孢型有机质(荧光); c— 泥页岩的暗色有机质纹层; d— 泥页岩中碳酸盐组分, 明显的微生物成因(荧光); e, f— 宏观藻类与高等植物残片; g, h— 保存在泥页岩中的孢型有机质; i, j— 保存在富有机质纹层中无定型有机质; k, l— 保存在碳酸盐组分中的无定形有机质; m— 水体中残留的高等植物残片; n— 现代水体中保存的孢子; o— 水体中的海雪聚合体; p— 水体中已经絮凝的聚球藻; q— 水体底部的微生物席(扫描电镜)Fig.2 Characteristics and origin of organic matter in lacustrine argillutite (after Tyson, 1995; Batten, 1996; Zhang et al., 2013; Zeng et al., 2018) |
通过泥页岩孢粉相分析可知, 从生物、水体到沉积物, 自然界中的有机质是可以溯源的。由此, 总结了不同自然环境有机质的关联性(表1)。
| 表1 各类有机质特征总结 Table1 Summary of characteristics of various organic matters |
生物有机质中的植物残体(如木质和丝炭等), 在水体中也同样出现常呈现颗粒有机质的形态存在, 在沉积物中为粗粒级、低密度组分的结构有机质, 这类有机质结构稳定, 不易被降解(Keil et al., 1994b; 蔡进功等, 2011)。生物体的分泌物或有机质分子, 在水体中以溶解有机质的形式存在, 且多为低分子态有机质, 因其化学活性强而与无机矿物关系密切; 在沉积物(岩)中能见到大量的无定形有机质, 其富集于细颗粒、高密度组分中, 通过有机溶剂的抽提可获得大量的可溶有机质, 部分有机质与黏土矿物等紧密结合(蔡进功等, 2005)。底栖微生物席等生物体可通过矿化作用形成微生物岩等沉积物(岩)。
对生物、水体和沉积物(岩)中有机质类型和性质的溯源对比可知, 自然界中有机质的类型多样, 性质差异巨大, 这将造成有机质聚集和沉积的差异性, 值得人们关注。
通过生物体、水体和沉积物(岩)中各类有机质的形态和性质差异研究, 充分认识到了有机质从水体到沉积物间的关联性。下面将探讨不同类型的有机质从水体沉降至沉积物的过程中沉积方式的差异。
4.2.1 机械沉积有机质特征
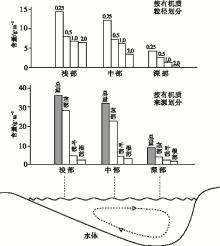
外源输入的陆生植物残片和水体中的部分有机质, 由于自身颗粒较大, 且表面活性较差, 颗粒之间的相互作用也很微弱。Wiley(2014)研究发现, 这些有机质的沉积方式与泥砂颗粒非常相似, 主要受颗粒粒径和水动力条件的影响, 机械沉降方式主导了这类有机质的沉积(Tyson, 1995)。这类有机质主要分布在近岸浅水区域。现代研究中发现, 粒径主导了有机质颗粒的搬运距离, 当有机质粒径大于63 μ m时会快速沉积进入水底; 当有机质粒径较小(< 63 μ m)时, 颗粒悬浮于水体中, 有机质的搬运距离则更远(Gastaldo and Huc, 1992)。此外, 这些机械沉积的有机质并不是一个整体, 植物往往具有木质、角质、皮质以及非木质和非皮质等组织的分化。这些组织由于成分和理化性质的差异, 在水体中的搬运距离和沉积特征都各不相同, 在相同的环境下, 木质组织残片粒径最大, 搬运距离最近, 皮质组织等粒径较小的残片搬运距离则更近(图3; Tyson, 1995)。
 | 图3 水体中外源输入有机质的分布情况(据Tyson, 1995)Fig.3 Distribution of exogenous organic matter in water body(after Tyson, 1995) |
由于自身稳定性较好, 这类有机质的保存能力较强, 埋藏后能够很好地保存下来(Tyson, 1995)。前人研究发现, 在沉积过程中, 这类有机质与外源输入的碎屑矿物共同保存, 但二者的结合关系并不紧密(Zeng et al., 2018)。
4.2.2 化学沉积有机质特征
水体中的微生物、生物分泌物和有机分子聚合体, 由于自身带有活性基团, 在水体能够发生聚合作用并彼此粘合(Turner, 2002; Burd and Jackson, 2009; Turner, 2015)。水体中有机质颗粒间相互聚集是化学沉积过程, 包括电化学沉积与生物化学沉积(Turner, 2015)。电化学沉积作用与有机质颗粒的表面电荷特征密切相关(Elimelech et al., 2013), 有机质颗粒往往具有双电层特性, 在离子的参与下, 有机质颗粒间能够聚集在一起进而形成有机质聚合体(Sharma et al., 2006)。部分微生物表面产生的胞外聚合物, 因具有黏滞性而能够黏合周围的有机质, 进而促使颗粒表面的有机质不断聚合进而形成了以微生物为核心的有机质聚合体(Villacorte et al., 2015)。值得注意的是, 在有机质聚合的过程中, 往往能够吸附部分矿物并共同沉积(Flü gel, 2010, 2012)。
4.2.3 生物沉积有机质特征
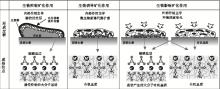
底栖微生物席等的沉积与保存伴随着矿物的形成。与生物生命活动及生物遗体紧密相关的沉积作用称为生物沉积作用。生物的沉积作用可表现为生物遗体直接堆积, 也可表现为间接的方式, 后者表现在生物的生命活动过程中或生物遗体的分解过程中, 引起介质的物理、化学环境发生变化, 从而使某些物质沉淀或沉积。底栖沉积的生物(底栖微生物席等) 的原地沉积往往能够产生生物沉积。底栖宏观藻类基本以原地沉积为主, 在沉积过程中, 底栖微生物席的钙质骨骼经有机或生物矿化作用形成碳酸盐等矿物(图4), 少量有机质在矿物形成的过程中被保存下来(Keil and Mayer, 2014)。这类有机质最显著的特征是在沉积过程中伴随矿物的形成, 矿化过程在沉积过程中起到了重要作用。
 | 图4 不同有机矿化作用下形成的碳酸盐矿物(据Dupraz et al., 2009)Fig.4 Carbonate minerals formed under different types of organic mineralization(after Dupraz et al., 2009) |
在沉积物中, 机械沉积、化学沉积和生物沉积构成了有机质沉降方式的多样性。其中, 无定形有机质由化学或生物沉降方式保存, 这也导致大量的无定形有机质主要存在于黏土矿物中, 在分离检测中表现出了高密度、细粒级的特征; 植物残片等结构有机质通过机械沉积方式进入水底, 且由于自身的稳定性而得以完整保存。可见, 有机质的差异沉积直接影响了其在沉积物中的保存方式。虽然水体环境具有复杂性, 在同一环境下往往由多种作用共同影响了有机质的沉积, 但通过有机质自身性质和特征的研究, 能够确定起主导作用的方式, 从而对沉积物(岩)的形成有更全面地认识。
从前面的分析可知, 在自然界中不同类型有机质的形态和性质都有差异, 由此导致有机质沉积方式的多样性。无定形类有机质受黏土矿物的保护可有效抵御微生物降解与氧化, 能迅速地絮凝和沉降, 缩短了从生物圈、水圈到岩石圈的进程, 致使碳循环周期延长; 机械沉积主导的颗粒有机质, 可以长期保存在沉积物中, 碳循环周期最长(蔡进功等, 2011)。因此, 关注有机质的沉积方式对碳循环周期的影响, 为碳汇研究提供了新思路。以絮凝为主的无定形有机质和以机械沉积为主的颗粒有机质在沉积岩中的保存方式截然不同(Zeng et al., 2018), 这反映了在不同环境下沉积岩成因的差异, 因此, 在充分了解有机质沉积方式的基础上, 将矿物和有机质结合考虑, 对理解泥页岩的形成具有重要意义。另外, 有机质和矿物之间具有不同的关系, 会导致在演化过程中有机质生烃有不同步的可能性。有机质沉积特性的研究, 能够为烃源岩中有机质在演化过程中差异生烃奠定基础。综上所述, 关注有机质的沉积方式对沉积学的研究具有重要意义。
1)自然界中生物(如动物、植物和微生物等)在生长、死亡甚至分解和降解的过程中, 会有不同的有机质样式呈现, 既有生物体形态, 也有生物生长过程中的分泌物和排泄物, 以及生物体死亡后的碎片或残片和分解或降解的分子化合物等, 这造就自然界中有机质的多样性特征。
2)海洋中的有机质可划分为颗粒有机质和溶解有机质, 2种有机质都有丰富的类型和来源, 且彼此间存在一定的转化关系, 这些性质和特征架起了不同类型有机质间聚集和沉积的桥梁。
3)沉积物(岩)中的有机质以多种方式保存, 通过粒度或密度分级分离及孢粉相分析发现, 生物体、生物残体和无定形等各类有机质, 常与特定的无机矿物共生, 如无定形多富集在黏粒级颗粒中, 而生物体和生物残体多富集在粗颗粒中。进一步溯源可知, 来源于生物体的各类有机质都可以通过不同的方式而沉积。
4)综合有机质的形态、性质以及与矿物共生关系, 发现有机质可通过机械沉积、化学沉积和生物沉积等3种不同的方式沉积和保存, 其中机械沉积的有机质以惰性的生物残体为主, 化学沉积的有机质以活性极强的无定形为主, 生物沉积有机质以活性较强的微生物和粘附物为主。有机质沉积方式及有机质特征、矿物— 有机质间关系和保存条件的差异, 决定了有机质的演化命运的不同, 这对碳循环、细颗粒沉积岩的形成和烃源岩生烃演化等方面的研究有重要意义, 因而应引起人们的高度关注。
作者声明没有竞争性利益冲突.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
| 56 |
|
| 57 |
|
| 58 |
|
| 59 |
|
| 60 |
|
| 61 |
|
| 62 |
|
| 63 |
|
| 64 |
|
| 65 |
|
| 66 |
|
| 67 |
|
| 68 |
|
| 69 |
|
| 70 |
|
| 71 |
|
| 72 |
|
| 73 |
|
| 74 |
|
| 75 |
|
| 76 |
|
| 77 |
|
| 78 |
|
| 79 |
|
| 80 |
|
| 81 |
|
| 82 |
|
| 83 |
|
| 84 |
|
| 85 |
|
| 86 |
|
| 87 |
|
| 88 |
|
| 89 |
|
| 90 |
|
| 91 |
|
| 92 |
|
| 93 |
|
| 94 |
|
| 95 |
|
| 96 |
|
| 97 |
|
| 98 |
|
| 99 |
|
| 100 |
|
| 101 |
|
| 102 |
|
| 103 |
|
| 104 |
|
| 105 |
|
| 106 |
|
| 107 |
|
| 108 |
|
| 109 |
|
| 110 |
|
| 111 |
|
| 112 |
|
| 113 |
|
| 114 |
|
| 115 |
|
| 116 |
|
| 117 |
|
| 118 |
|
| 119 |
|
| 120 |
|
| 121 |
|
| 122 |
|
| 123 |
|
| 124 |
|
| 125 |
|
| 126 |
|
| 127 |
|
| 128 |
|
| 129 |
|
| 130 |
|
| 131 |
|
| 132 |
|
| 133 |
|
| 134 |
|
| 135 |
|
| 136 |
|
| 137 |
|
| 138 |
|
| 139 |
|
| 140 |
|
| 141 |
|
| 142 |
|
| 143 |
|
| 144 |
|
| 145 |
|
| 146 |
|
| 147 |
|
| 148 |
|
| 149 |
|
| 150 |
|
| 151 |
|
| 152 |
|
| 153 |
|