



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
碳酸盐工厂与浅水碳酸盐岩台地: 研究进展与展望*
[颜佳新1, 2  , 孟琦
, 孟琦1, 2 , 王夏3 , 刘志臣1, 4 , 黄恒1, 5 , 陈发垚1, 2 , 郭全鼎1 ]
, 孟琦|
|

第一作者简介 颜佳新,男,1962年生,教授,博士生导师,主要从事沉积地质学研究工作。E-mail: jaxy2008@163.com。
碳酸盐岩具有广泛的时空分布,主要为生物成因,而该生物成因属性注定了其与海洋生物、海洋化学条件等密切相关。海相碳酸盐岩沉积相模式对于描述、表征和解释碳酸盐岩地层结构成因具有重要的指导意义。本文回顾了近 70年来碳酸盐岩沉积相模式从二维水体能量模型到三维地形模型的发展变化。如何深入分析不同形态特征和内部构成的碳酸盐岩台地的成因机制,解读其相应的地球生物—环境演化信息,是当前和下一步需要努力的方向。为此本文介绍了碳酸盐工厂和碳酸盐岩台地成因方面的研究进展,强调了碳酸盐岩地层的时代特殊性,并以滇黔桂地区为例,从碳酸盐工厂分析的角度剖析了该区二叠纪吴家坪期碳酸盐岩沉积学研究现状,展望了下一步的研究方向,旨在引起同行重新认识生物和海洋环境条件在碳酸盐岩台地演化中的作用,重视全球空间尺度范围内同时代碳酸盐岩的相似性与差异,并从古生态角度入手开展碳酸盐工厂分析,发掘碳酸盐岩台地的生物学和海洋学意义。
, Meng QiAbout the first author Yan Jia-Xin,born in 1962,is a professor in China University of Geosciences(Wuhan). He is mainly engaged in sedimentology. E-mail: jaxy2008@163.com.
Carbonate rocks are widely distributed in spatial and temporal. Most carbonates are biogenetic,which depend highly on marine biological and chemical and other intra-basinal environments. Such a biogenetic nature provides great potentials in deciphering marine biology and chemical conditions. Depositional facies model plays a key role in describing,characterizing and interpreting of ancient carbonate rocks. This paper briefly reviewed the development of the facies model in the past 70 years,from the 2D water-energy profile model to the 3D topographic profile model. How to deeply analyze the formation mechanism of the carbonate platform with different external shape and internal structures and to interpret the biological and environmental signals from them will be the next research focus. On this basis,this paper firstly introduced the progress in the formation mechanism of the carbonate factories and the carbonate platform,and emphasized the age-sensitive pattern of the carbonate platform. Then a case study from the Wuchiapingian(Permian)carbonate platform in the Yunnan-Guizhou-Guangxi region is used,in the view of carbonate factories,to scrutinize the current research status and propose the future research direction.This paper aims to recall geologists' attention to the role of the biotic evolution and marine environment in the development of the carbonate platform,to the similarities and differences of time-equivalent carbonate rocks in a global scale,and to explore the biological and oceanic significance of carbonate platform through the carbonate factory analysis.
在沉积地质学中, 古代“ 大套浅水碳酸盐沉积序列” 组成的三维沉积地质体(Tucker and Wright, 1990; Flü gel, 2004)称为“ 碳酸盐岩台地” , 当代碳酸盐沉积发育的二维浅水区域称为“ 碳酸盐陆棚” (与陆源碎屑沉积对应)或者“ 碳酸盐岩台地” (Read, 1982, 1985; Allaby and Allaby, 1991)。
碳酸盐岩具有广泛的时空分布, 其特征和成因研究在油气地质、矿产地质甚至地下水环境地质、工程地质中均具重要意义。20世纪60年代开始的碳酸盐岩沉积学研究取得飞跃发展, 已经建立了一系列的碳酸盐岩台地沉积相模式, 奠定了碳酸盐岩沉积学研究的基础。直至现在, 浅水碳酸盐岩台地相模式仍然受到广泛关注(顾家裕等, 2009; 王龙等, 2018), 这是因为碳酸盐岩的组分、结构、岩相、台地类型和内部构成及其成岩作用在时间上和空间上均表现出极大差异, 以致描述、表征和解释这些花样繁多的碳酸盐岩的工作既复杂又常常较为困难, 而沉积相模式在工作实践中却具有重要指导意义(Pomar and Hallock, 2008)。
在解读地球表层系统演化过程中的大气、海洋和生命演化特征方面, 盆内成因碳酸盐岩也是极为重要的载体。碳酸盐岩的生物属性(Carbonates are born, not made)(James, 1977), 注定了碳酸盐岩的成因与海洋生物、海洋化学条件等密切相关。将碳酸盐工厂的理念融入碳酸盐岩沉积相和古地理学研究, 从机理上深入认识碳酸盐岩的成因, 还原碳酸盐沉积形成时的海洋环境— — 生物和海洋化学条件, 将极大地推进碳酸盐岩沉积学的发展。本文在回顾海相碳酸盐岩沉积相模式研究历史的基础上, 介绍碳酸盐岩生产工厂和碳酸盐岩成因研究方面的进展, 展望下一步研究方向, 期望引起更多同行对碳酸盐岩台地成因研究的关注和重视。
在20世纪70年代, 沉积学家已经清楚地认识到, 绝大多数碳酸盐岩的形成和分解直接或者间接地与生物过程的影响或者控制有关(如Wilson, 1975)。从结构组分角度看, 碳酸盐沉积包括颗粒、灰泥和胶结物。颗粒主要包括内碎屑、生物碎屑(骨屑)颗粒和非生物碎屑(非骨屑, 如鲕粒、核形石)。毫无疑问, 骨屑颗粒是通过真核生物控制矿化作用(biologically-controlled mineralization, BCM)形成的。生物矿化不仅形成了传统意义上的“ 灰岩” 中的主体— — 碳酸盐矿物, 还可以形成与碳酸盐岩相伴出现的硅质矿物和磷质矿物等(Weiner and Dove, 2003; Pé rez-Huerta et al., 2016)。
非骨屑颗粒中, 核形石的微生物成因已经得到广泛认同(Mei, 2007)。钙质鲕粒的形成时代和地域分布极为广泛, 也常常被当作水体超饱和条件下钙质化学沉淀的代表。现代钙质鲕粒内含有大量有机质, 其中约1/3属蛋白质类, 并富含氨基酸。从有机质含量和特征看, 与骨屑颗粒极为相似(Mitterer, 1968, 1972; Davies et al., 1978)。虽然钻孔藻活动及其后续泥晶充填给鲕粒结构和地球化学特征留下了深深的印记, 以致具有与非生物成因颗粒极为相似的特征(Duguid et al., 2010; Trower et al., 2018), 但是不可否认的是, 在钙质鲕粒的形成过程中, 微生物或者有机质仍发挥了重要作用, 如大量微生物胞外聚合物(EPS)的出现(Li et al., 2013)、与海水相似的稀土元素特征(Li et al., 2017)、与多种微生物成因沉积相伴出现(Riding et al., 1991; Woods, 2013)以及发育多种生物矿化作用的微生物功能群(Diaz et al., 2014)。
生物作用可以形成体量巨大的厚层块状生物礁、分布广泛的微生物岩和大套细粒碳酸盐岩序列。生物礁(碳酸盐建隆)和微生物岩自不必说。在细粒泥晶碳酸盐沉积中, 生物同样发挥了重要的作用, 如早已识别出的骨屑颗粒解体和泥晶化。在热带浅水地区常常出现白色碳酸盐物质悬浮于水体中(称为whiting), 这是浅水地区细粒碳酸盐沉积的主要来源(Cloud, 1962; Robbins et al., 1997), 却曾一度被误以为是化学成因(碳酸盐的超饱和结晶)或者物理成因(搅动再悬浮)的典型代表。实际上, 它们与藻类繁盛(Shinn et al., 1989)或者与鱼类生活有关, 如在巴哈马地区, 平均约14%的细粒碳酸盐沉积物(灰泥)产率与鱼类代谢作用有关(Perry et al., 2011)。
海水化学成分在显生宙经历了巨大的变化, 尤其是海水Mg/Ca值(有关参考文献参见颜佳新和伍明, 2006; 刘喜停和颜佳新, 2009)。主要是因为生物成因的碳酸盐岩也将受到海洋化学条件的影响, 生物演化与海水化学成分演化相互作用, 联合控制着生物骨屑颗粒的原生矿物成分。对40类研究程度较高的后生动物骨骼的原生矿物成分统计发现, 其中25类生物的骨骼原生矿物成分与其起源时的海水成分完全对应; 37类生物碎屑的矿物成分与起源时海水成分相关(Porter, 2007, 2010)。因此, 深入理解碳酸盐岩与生物和海洋化学条件之间的密切联系, 是深入解读碳酸盐岩成因的关键。
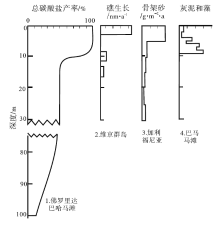
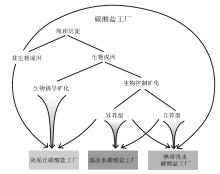
由于大部分碳酸盐沉积物是由分泌碳酸盐的生物产生, 其中不少是光合作用的副产物, 因此碳酸盐的生产过程主要取决于光照程度(图 1)。在海水上部100 m的水层中, 特别是表层10 m, 悬浮着大量能进行光合作用的生物, 是碳酸盐沉积物的主要生产场所(图 1, 图 2)。这个具有高生物产率的浅水区域被称为“ 碳酸盐工厂” (James, 1977)。自20世纪90年代以来, “ 碳酸盐工厂” 这一概念得到重视和深化, 已经确立了热带浅水碳酸盐工厂、灰泥丘工厂和温凉水碳酸盐工厂各自的识别特征(Schlager, 2003)(图 3)。
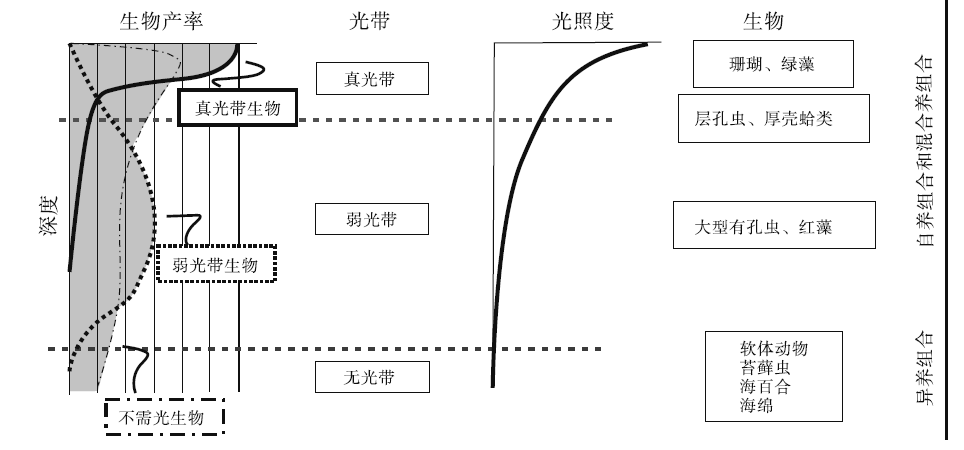 | 图 1 水柱中碳酸盐生产(工厂)与光照控制的主要生物类群的水深梯度分布关系(据Pomar, 2001b)Fig.1 Carbonate production vs. bathymetric gradients of main groups of organisms controlled by their dependence on light penetration in water column(after Pomar, 2001b) |
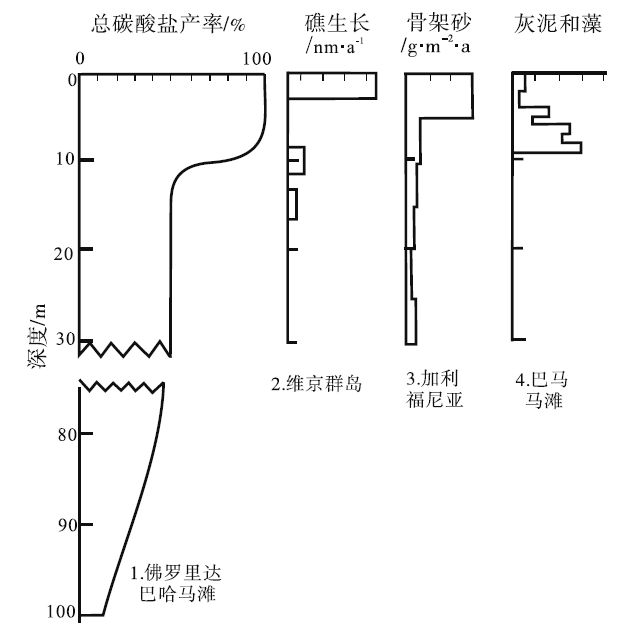 | 图 2 碳酸盐产率与水深的关系(据Sarg, 1988)Fig.2 Carbonate production vs. water-depth(after Sarg, 1988) |
 | 图 3 海洋碳酸盐沉淀方式与碳酸盐工厂(据Schlager, 2003)Fig.3 Patterns of marine precipitation and carbonate factories (after Schlager, 2003) |
热带浅水碳酸盐工厂所处环境水体溶氧浓度高、营养物质较贫乏, 但是光照条件好, 处于南北纬30° 之间, 水体温暖。该生产工厂光能自养型生物繁盛, 如藻类和与营光合作用的藻类共生的动物, 包括造礁珊瑚、底栖有孔虫和一些软体类; 不依赖光照的异养生物常见, 但不是标志特征(图 1)。因此, 热带浅水碳酸盐工厂以生物控制的碳酸盐矿化为特征, 在台地边缘生物格架和生物建隆较为普遍。也可以发育钙质鲕粒等非骨屑颗粒(图 3)。
现代大洋低纬度地区的一些珊瑚礁岛是该类碳酸盐工厂的典型代表, 它们具有惊人的碳酸盐生产效率。图 4展示的是南太平洋地区一个发育环状珊瑚礁的火山岩岛, 构成一个中心为火山岩岛屿及潟湖、周边为镶边碳酸盐岩台地的混积沉积体系。台地边缘生物礁和生物碎屑滩相主要由珊瑚和珊瑚藻构成; 在内侧浅水潟湖(水深< 5 m)沉积物中, 非骨屑颗粒含量可以达到30%~50%; 较深水潟湖(15~40 m)内, 主要为含软体和有孔虫等异养生物的文石针状沉积, 文石质灰泥来源主要与绿藻和颗石藻有关。在该障壁生物礁— 火山岩岛沉积体系中, 陆源碎屑沉积体系不发育, 靠近火山岩岛的沉积物中陆源碎屑物质平均含量少于10%, 热带暖水碳酸盐沉积物的分布占据绝大部分的水域; 环礁中的潟湖相主要被浅水碳酸盐沉积物充填, 潟湖较深水区域位于近火山岩基岩海岸线一侧, 清楚地展示出碳酸盐工厂的生产效率远远超过火山岩基岩风化剥蚀的输入(Gischler, 2011)。
 | 图 4 Bora Bora岛碳酸盐岩沉积相分布(据Gischler, 2011)Fig.4 Distribution of carbonate sedimentary facies of Bora Bora(after Gischler, 2011) |
在温跃层之下, 热带浅水碳酸盐工厂转换为温凉水碳酸盐工厂。这里为弱光或无光区, 水温低, 营养物质一般比热带浅水碳酸盐工厂区丰富。温凉水碳酸盐工厂的生物以异养生物为主, 上部可以发育红藻并伴生大型有孔虫。沉积物中缺少浅水生物礁和钙质鲕粒(图 1, 图 3)。水温更低的冷水碳酸盐工厂, 主要出现在南北纬30° 到极地的范围内。在低纬度地区上升流发育的地方, 热带浅水碳酸盐工厂/温凉水碳酸盐工厂的转化可以发生在浅水区。因此, 温凉水碳酸盐工厂的水深范围可以从浅海延伸到半深海甚至深海。
灰泥丘碳酸盐工厂中沉积物颗粒非常细, 为原地沉淀, 是生物和非生物因素在细菌参与下共同形成的, 包括有机质的降解(矿化)。典型代表有古生代和中生代的灰泥丘, 所以称之为“ 灰泥丘工厂” 。其典型环境为弱光或透光带, 营养物质丰富但贫氧。在元古代和显生宙生物大灭绝之后的特殊时期, 该工厂可以占据热带浅水生产工厂的位置(图 3, Schlager, 2003)。
将海洋生态系的概念融入到碳酸盐沉积物的生产(工厂)中, 为碳酸盐工厂的进一步分析提供了方向。随着研究的深入, 碳酸盐工厂已经呈现出快速发展的态势, 频频出现在碳酸盐岩沉积学研究成果中, 如鲕粒碳酸盐工厂、微生物工厂等, 值得进一步关注。
碳酸盐岩地层的沉积古地理编图研究离不开碳酸盐岩沉积相模式研究。早期的碳酸盐岩沉积学研究, 主要关注化石、岩性特征和地层时空分布特征的描述。在石油工业的推动下, 20世纪50年代, 开始了大规模现代碳酸盐沉积系统的调查研究, 以探索碳酸盐沉积的形成机理及其空间分布规律, 并依据“ 将今论古” 的原理将其用于古代石灰岩研究中(Ahr, 1973)。自20世纪60年代起, 开始建立相应的碳酸盐岩沉积相模式, 直至最近仍不断有新的研究成果出现(王龙等, 2018)。
碳酸盐岩沉积相模式实际上是一个阶段人们对碳酸盐岩成因认识的最好总结; 半个世纪碳酸盐岩沉积相模式的演变, 映射了人们对碳酸盐沉积(岩)成因认识的心路历程。但是碳酸盐岩沉积相模式之多在其他沉积岩类中是不多见的。回首已有研究, 可以将碳酸盐岩沉积相模式的发展大体分为2个阶段。
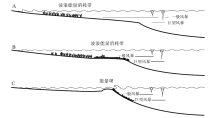
在20世纪50年代之前, 对碳酸盐岩沉积环境的认识相当肤浅, 几乎全是十分笼统的“ 浅海” 二字(冯增昭, 1989)。Shaw(1964)发现现代碳酸盐沉积环境主要是陆缘海, 古代碳酸盐岩主要发育在陆表海环境, 因此提出陆表海的概念,
论述了陆表海的水能量特征, 并将陆表海碳酸盐沉积物的分布(沉积相)与水能量分带联系起来, 从此奠定了陆表海碳酸盐岩沉积环境分析的理论基础(转引自冯增昭, 1989)(图 5)。这个水体能量分带模型经Irwin(1965)修订为X、Y和Z带(由海向陆), 之后Laport(1967)和Young(1972)等将碳酸盐沉积环境水能量特征及分带与潮汐作用相联系, 提出了相应的潮坪碳酸盐岩沉积相模式(转引自冯增昭, 1989)。类似于当时的碳酸盐岩岩类学研究, 这些模式重视碳酸盐沉积与水动力之间的关系, 同时也反映了碳酸盐沉积与陆缘碎屑沉积的差别, 清楚地体现了清澈、浅水环境条件对碳酸盐沉积形成的重要性(如Irwin, 1965)(转引自冯增昭, 1989)。
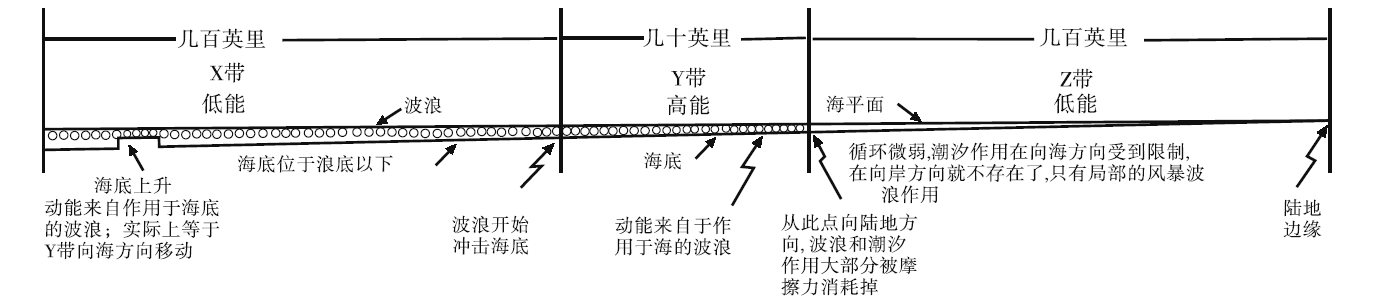 | 图 5 陆表海的能量带(据Shaw, 1964, 转引自冯增昭, 1989)Fig.5 Energy belts of epeiric sea(after Shaw, 1964, in Feng, 1989) |
对应陆源碎屑浅海陆棚, 发育碳酸盐沉积的浅海陆棚区被称为碳酸盐陆棚(carbonate shelf)。由于这些浅水碳酸盐沉积往往呈现顶面平坦、外侧具陡峻斜坡的地形, 因此也被称为碳酸盐岩台地(carbonate platform)。Ginsburg和James(1974)在总结对比全新世8个碳酸盐陆棚(北美东南部陆棚、佛罗里达西部陆棚、尤卡坦陆棚、澳大利亚北部陆棚、大堡礁陆棚、贝利兹陆棚、南佛罗里达陆棚和巴哈马滩)碳酸盐沉积物分布特征的基础上, 发现它们可以分为2种类型: 镶边陆棚(rimmed shelves)和开放陆棚(open shelves)。
但是有些碳酸盐沉积环境水动力高能带位于向陆地一侧, 随着水深向海增加、水体能量逐渐降低, 类似于现代的波斯湾, 它们明显不同于上述顶部平坦、外侧发育高能沉积相带的碳酸盐陆棚。据此, Ahr(1973)提出另外一种陆棚碳酸盐沉积相模式— — 缓坡(ramp)模式。在当时以现代沉积研究为主的背景下, 这些沉积相模式均为二维模型。
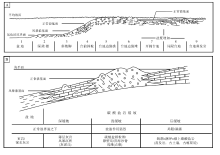
经过20多年的调查研究后, 全球大部分现代典型碳酸盐沉积区已不再是未经开发的处女地, 人们开始转向研究古代灰岩(Ahr, 1973)。在稳定的克拉通地区和被动大陆边缘地区, 浅水成因碳酸盐岩可以形成厚达数千米、分布达数百甚至数千千米的巨大沉积体, 描述、测量和采样也极为方便。Wilson(1975)首次在概念上将这些巨型三维碳酸盐沉积体(相)与地貌— 环境单元概念的“ 陆棚” 区分开, 陆棚(shelf)仍指发育浅水碳酸盐沉积的一个二维区域(地貌单元), 而碳酸盐岩台地和缓坡作为碳酸盐沉积相则指二维“ 陆棚” 区域之下的巨型三维碳酸盐沉积体(图 6-A)。
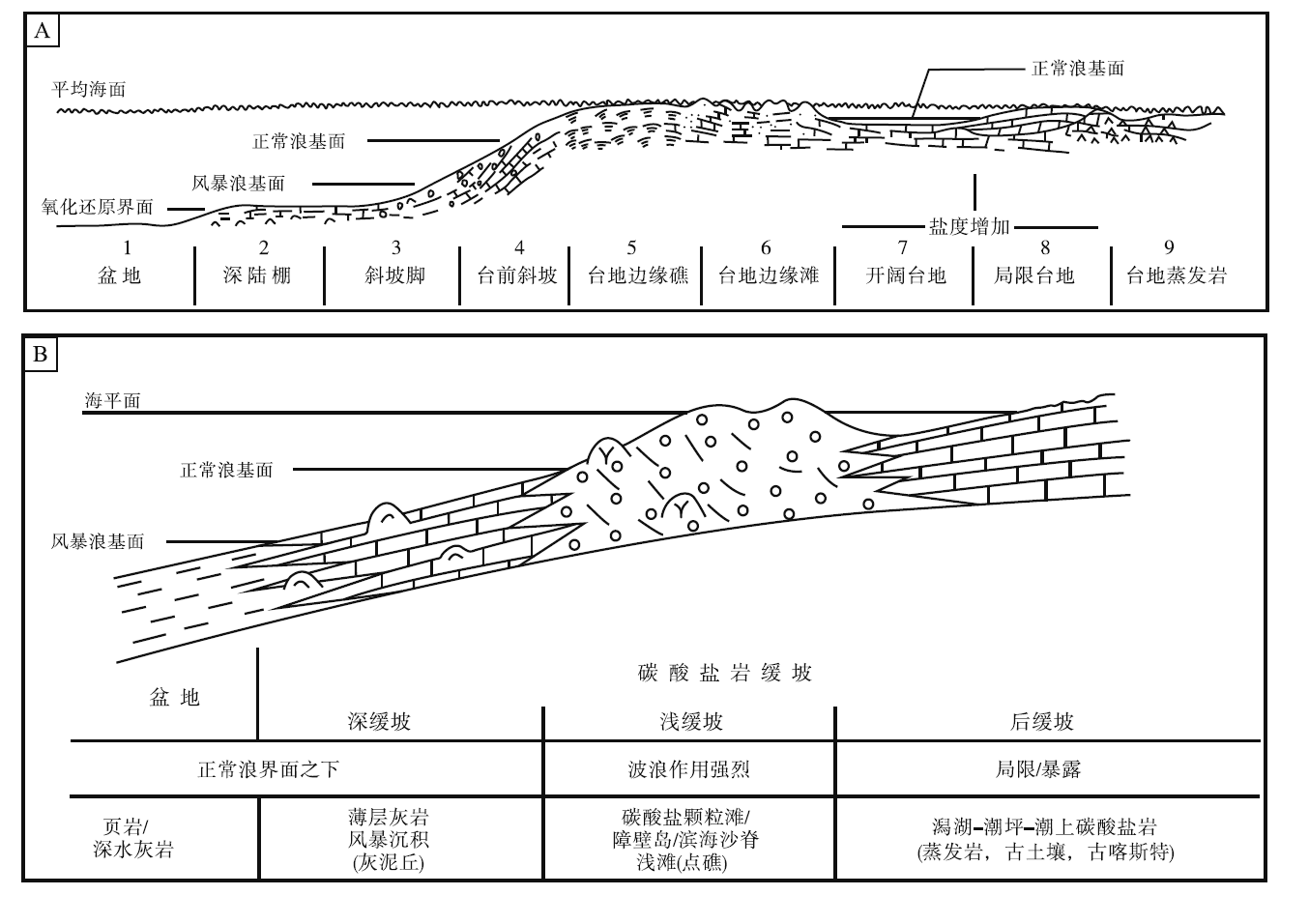 | 图 6 碳酸盐岩台地相模式(A)(据Wilson, 1975)和碳酸盐岩缓坡模式(B)(据Tucker and Wright, 1990)Fig.6 Carbonate platform model(A)(after Wilson, 1975) and carbonate ramp model(B) (after Tucker and Wright, 1990) |
依据岩石的结构、构造和颗粒类型等特征, Wilson(1975)将9个在地质历史中反复出现的碳酸盐岩沉积相, 构成一个完整的碳酸盐岩沉积相模式。9个标准相包括台地蒸发相、局限台地相、开阔台地相、台地边缘滩相、台地边缘礁相、台前斜坡相、斜坡脚相、深陆棚相和盆地相(图 6-A)。在此之前, 碳酸盐岩因为其变化多样的结构、构造特点和复杂的颗粒类型, 加上非均一化的成岩作用, 常常使人眼花缭乱; 自此以后, 碳酸盐岩沉积环境分析不再令人生畏。该模式不仅适用于整个显生宙, 稍加修改还可以用于前寒武纪(Schlager, 2000)。虽历经30余年的实践检验, 且经常被修订, 但是仍难以撼动Wilson(1975)碳酸盐岩台地相模式在碳酸盐沉积相研究中的地位。
Wilson(1975)9个标准相带由海向陆的排列呈现了镶边陆棚的特点, 因此常被人认为是“ 镶边陆棚” 碳酸盐岩沉积相模式。实际上, 可以将其标准相作为基本模块组合成新的相模式, 包括缓坡沉积相模式(Schlager, 1997)。不过, 碳酸盐岩缓坡沉积相模式的修订和发展, 主要还是得益于Tucker和Wright(1990)与Burchette 和 Wright(1992)的贡献(图 6-B)。
在三维形态型碳酸盐岩沉积相模式提出的同时, 人们已经意识到形成巨大的浅水碳酸盐沉积体与其所处的区域构造背景分不开, 而且在沉积盆地的演化发展中, 三维形态的“ 镶边台地” 和“ 缓坡” 沉积相格局可以互相转化, “ 镶边台地” 和“ 缓坡” 可能只是盆地演化发展过程中的一个阶段(Read, 1982, 1985; Bosence, 2005)。在侧向(空间)上也常常可以看到, 同一个浅水碳酸盐沉积体有可能东侧被认为是镶边台地, 西侧被认为是缓坡。将这种空间差异归结为构造背景方面的不同, 如同沉积断裂, 是再自然不过的事情。
从特征上看, Read(1982, 1985)的碳酸盐岩台地相模式应视为三维形态型沉积相模式。但是在这些模式中, 沉积相类型或与沉积发育之初的地形地貌特征相关, 或与盆地发育过程中的区域构造背景有关。较为完善的碳酸盐岩台地构造背景分类体现在Bosence(2005)的研究中, 包括: 断块型(fault-block)、盐丘穿刺型(salt-diapir)、下沉(被动)陆缘型(subsiding or passive margin)、孤立台地型(offshore bank or unattached platform)、火山底座型(volcanic pedestal platform)、前陆边缘型(foreland margin)、逆冲断块顶部型(thrust-top)和三角洲沉积顶部型(delta-top)。中国学者也进行过积极尝试(如顾家裕等, 2009)。细究这些对浅水碳酸盐岩沉积相模式的补充, 可以看出他们关注的重点还主要是单个影响控制因素, 尤其是区域构造和区域古地理方面的, 故内在成因机制仍然需要解读。
对碳酸盐岩台地形态和结构的详细刻画, 是为了深入分析其成因。经过半个多世纪的(古代和现代)碳酸盐岩沉积相描述和资料积累, 浅水碳酸盐岩台地三维形态类型已经基本清楚, 可以视为由同斜缓坡和镶边台地2个端元及一系列介于两者之间的过渡类型(Pomar, 2001b)所组成的群集。如何深入分析不同形态特征和内部构成的碳酸盐岩台地的成因机制, 解读其相应的地球生物— 环境演化信息, 是当前和下一步需要努力的方向。从碳酸盐生产角度分析碳酸盐岩台地的成因, 需要重点考虑沉积物类型和特征、沉积物生产工厂位置以及沉积环境水动力状况(Pomar, 2001b)。在此有必要先介绍一下生态可容空间的概念, 并重新审视台地边缘特征以及对台地形态特征有着重要控制作用的几个主要因素。
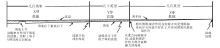
在大陆边缘陆源碎屑沉积体系中, 陆源碎屑物质由岸向海输送, 按粒度由粗到细依次堆积下来。受陆棚平衡剖面控制, 能容纳沉积物的有效空间位于沉积基准面之下(可容空间)。在沉积基准面之上, 受波浪搅动和水流作用, 细粒物质难以堆积下来。因此沉积基准面和海底之间构成一个受水动力控制的物理可容空间。但是在碳酸盐沉积体系中情形有些不同, 形象地说, 物理可容空间不仅仅是码放“ 产品” 的仓库, 也是“ 产品” 的一个重要生产车间(连同沉积基准面之上的水体环境)。在此沉积体系中, 环境条件, 如气候、温度、盐度、水体营养等, 直接影响生物的类型及其兴衰, 生物条件(产品制造者)直接影响碳酸盐沉积物的生产效率和特征。适宜的生态条件可以生产大量的碳酸盐沉积物; 相反在环境条件恶化时, 碳酸盐沉积物的生产将受到抑制甚至(工厂)停止或者发生转换(图 7)。
重要的是, 生物成因的骨架构造可以抗波浪和洋(海)流的改造, 其堆积地形(产品码放)可以向上突破沉积基准面限制直抵海平面附近。虽然未固结的、松散的碳酸盐沉积物仍然可以受到水动力搬运与扩散的影响, 但生物因素仍然在起作用, 如包粘、捕获、障积, 还包括与有机质有关的早期成岩胶结。因此, 碳酸盐沉积物主要生产者(生物)的特征和数量可以改变其沉积物可容空间, 这就是碳酸盐岩沉积体系中生态(控制的)可容空间的理念(Pomar, 2001a, 2001b; Pomar and Hallock, 2008)。碳酸盐岩沉积体系中的沉积基准面取决于碳酸盐工厂生产的沉积物类型以及沉积物产生的位置(Pomar, 2001a)。
 | 图 7 碳酸盐岩台地形成发展过程中的物理和生物因素关系流程图(据Pomar and Kendall, 2008)Fig.7 Flow diagram of the physical and biological relationships that control carbonate accumulation in carbonate platforms(after Pomar and Kendall, 2008) |
生物因素不仅控制着碳酸盐的生产, 密集分布的生物体本身以及生物建造还影响着沉积环境水动力状况, 是重要的水力(波浪)屏障或者消减因素(图 8)。在碎屑岩— 碳酸盐岩混积的陆棚环境中, 往往因为水质浑浊难以形成快速生长的生物建隆, 缺乏远离海岸的水力学屏障, 沉积基准面主要受物理可容空间控制。如果近岸浅水地区有大量的海草繁殖, 它们不仅能消耗广海传递过来的波浪能量, 其共生生物还能产生大量的松散碳酸盐沉积物, 如在地中海地区中— 新生代碳酸盐沉积体系中。这些松散沉积物被周期性的风暴回流输送到真光带下部, 可堆积形成一个远端变陡的缓坡(图 8-A)。在水质较清澈的情况下, 一些具有较强建造能力、也能适应软基底的生物, 如厚壳蛤、红藻及结壳等, 可以在沉积基准面之上形成相应的生物建隆, 并有效地消耗来自广海的波浪能量(图 8-B)。较高建造能力的生物建隆, 如现代的台地边缘珊瑚礁, 可以形成坚固的抗浪格架, 堆积地形可以高至海平面附近, 不仅是一个抵御外海波浪能量的屏障, 也制约着生物礁后侧台地顶部的水体循环(图 8-C)。因此, 碳酸盐沉积物主要生产者(生物)的特征和及发育位置, 将直接影响碳酸盐岩台地的形态。
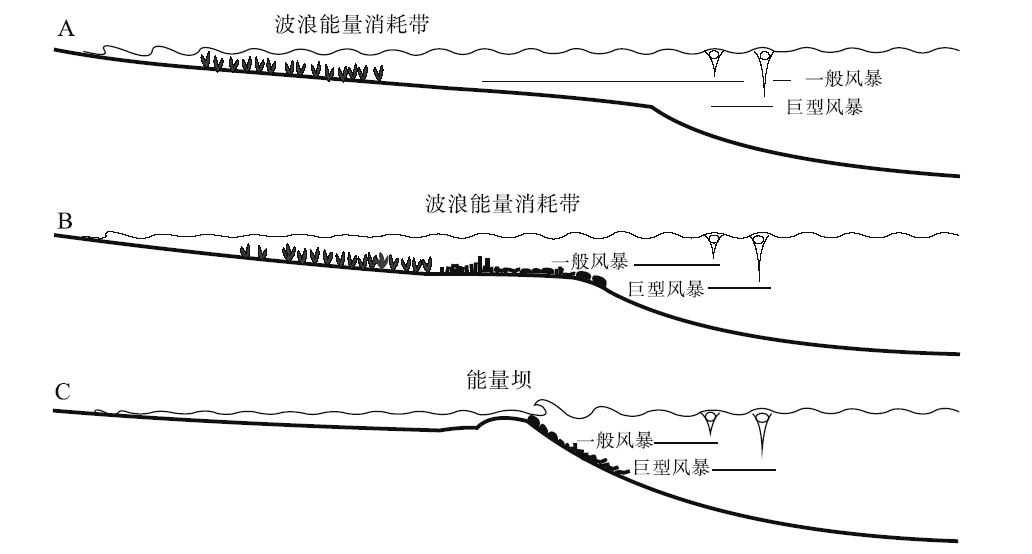 | 图 8 水能的耗散示意图(据Pomar and Kendall, 2008)Fig.8 Sketch map of dissipation of the hydraulic energy(after Pomar and Kendall, 2008) |
如上所述, 台地边缘碳酸盐岩建隆的发育位置和特征是区分碳酸盐岩台地类型的关键。在浅水碳酸盐岩台地发育演化过程中, 真光层内造架生物群的发育往往形成镶边台地。如果真光层内缺乏造架生物, 代之以偏好软基底的生物群, 意味着大量砂砾级碳酸盐沉积物的生产, 它们在水动力搬运作用下易于形成顶部平坦的开阔台地(陆棚); 如果组成生物建隆的生物群繁盛于寡光带, 如新生代的大型底栖有孔虫和红藻, 则极易形成远端变陡的缓坡; 如果缺乏生物建隆形成的环境和生物条件, 生物在真光带和寡光带形成大量的灰泥沉积物, 在水动力作用改造下, 则形成同斜缓坡(图 9)(Pomar and Kendall, 2008)。由此可见, 生物作用在碳酸盐岩台地边缘和台地形态类型中发挥着关键作用。
 | 图 9 不同碳酸盐生产和分散方式形成的沉积剖面图(据Pomar and Kendall, 2008) 右侧粒度刻度: m、s、S、g、b、F分别对应灰泥、粉砂、砂、细砾、巨砾、格架Fig.9 The depositional profiles that result from different types of carbonate production and dispersal(after Pomar and Kendall, 2008) |
生物建隆与碳酸盐岩台地形态之间的密切关系, 或者说生物建隆对碳酸盐岩台地特征的控制作用, 放在整个显生宙历史中来看更清楚。早古生代, 台地边缘生物建隆主要为古杯和微生物丘, 对应早古生代低起伏的台地边缘; 中古生代, 骨架礁主要由层孔虫和珊瑚构成, 台地往往呈现陡坡边缘; 晚古生代, 台地边缘往往发育钙质海绵、藻类、有孔虫、苔藓虫等建隆, 建隆中异养生物繁盛, 且富含灰泥, 灰泥丘也极为发育, 因此晚古生代期间多为低角度缓坡; 至三叠纪和侏罗纪, 石珊瑚成为建隆主要格架生物, 台地呈现陡峻边缘; 早中白垩世, 固着蛤可形成较为陡峻的台地边缘; 自古新世开始, 石珊瑚重新回归, 碳酸盐岩台地再次出现陡峻边缘直至现在(Moore and Wade, 2013)。
显生宙期间生物建隆与台地形态的密切关系正如Ginsberg指出的, 碳酸盐建隆就像莎士比亚戏剧, 精彩剧目不断重演, 替换的只是演员。但是如果遇到的是蹩脚演员和制片人, 效果就会有相差很远(Kuznetsov, 1990)。因此, 台地边缘生物建隆的生产能力对碳酸盐岩台地形态来说极为关键。
区域构造背景对浅水碳酸盐岩台地的初始形态、台地规模大小、台地类型之间转化的作用是毋庸置疑的, 也一直受到碳酸盐岩沉积学的重视, 有关内容可以参考相关文献, 如Read(1982, 1985)及Bosence(2005)的碳酸盐岩台地构造成因分类讨论。
但是从台地发展演化角度来看, 需要重视在区域性构造下沉或者海平面快速上升过程中, 碳酸盐岩台地和台地边缘生物建隆的表现, 即碳酸盐岩台地和生物建隆的增长潜能是否可以抵消区域性基底沉降或者海平面的快速上升, 考虑碳酸盐岩台地是否会因基底沉降或者海平面上升而淹没。现代珊瑚礁和碳酸盐岩台地的增长潜能平均值为1000 mm/ka, 洋壳下沉速率最大值为250 mm/ka, 盆地下沉速率平均值为10~100 mm/ka, 洋底扩张导致的海平面上升平均值小于10 mm/ka。因此碳酸盐岩台地的增长潜能远远大于海平面上升和盆地下沉带来的影响, 即碳酸盐沉积能够赶上一般的区域性构造下沉和海平面上升(Schlager, 1981, 1999)。
随着时间间隔的延长, 碳酸盐岩台地平均沉积速率会降低。在数十个百万年时间尺度范围内, 台地的增长速率大致与时间的平方根成反比。即使如此, 两者仍将在一个尺度范围内, 纯粹的海平面上升和热冷却下沉在数个至数十个百万年时间尺度范围内不会导致台地淹没, 台地淹没往往与海洋环境条件恶化有关(Schlager, 1999)。如在构造活动频繁复杂、以构造下沉影响为主的南海地区, 新生代碳酸盐岩台地的淹没往往与海平面上升和环境条件恶化相伴出现(Wilson and Hall, 2010)。西沙群岛和南沙群岛的新生代生物礁也没有在较快的构造热沉降中完全淹没(Hutchison and Vijayan, 2010; Wu et al., 2014)。
伴随海平面周期性变化, 台地沉积序列可以出现退积、加积和进积, 台地边缘可以呈现相应的短距离迁移摆动, 层序地层学对此作了极好的总结, 限于篇幅不在此细述。海平面变化对碳酸盐岩台地的影响, 更多体现在台地内部结构方面。单纯的海平面升降变化, 难以改变碳酸盐岩台地的整体特征。如全球石炭— 二叠纪冰期期间, 虽然广泛发育冰川性高频沉积旋回, 海平面变化幅度可以超过100 m(Wang et al., 2013), 但是中国华南地区晚古生代碳酸盐岩台地并未因此发生本质的改变, 就是极好的例证(严雅娟等, 2015)。
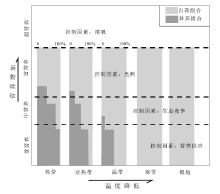
颗粒组合(包括骨屑颗粒和非骨屑颗粒)的概念源于Lees和Buller(1972)对全球68个浅水碳酸盐沉积区(南北纬60° 之内)的调查研究, 因为其特征客观也易于掌握, 现在已经得到了广泛使用。对碳酸盐沉积物颗粒的纬度分布投点表明, 非骨屑颗粒(鲕粒、球粒和团粒)主要限于热带、亚热带暖水环境; 骨屑颗粒(即生物碎屑)的纬度分布呈现2个组合: 低纬度热带、亚热带地区以造礁珊瑚和(或)绿藻发育为特征, 称为绿藻— 珊瑚组合(chlorozoan); 中高纬度地区以缺乏造礁珊瑚和绿藻为特征, 称为有孔虫— 软体组合(foramol)(颜佳新, 1991)。随后在20世纪八九十年代出现一个针对碳酸盐沉积骨屑颗粒组合识别及环境条件研究的小高潮, 大量不同特征的骨屑颗粒组合得以描述和记录(Nelson, 1988)。为方便交流, James(1997)将其归纳为自养光和作用组合(photozoan)和异养组合(heterozoan)2类。
影响碳酸盐沉积颗粒组合特征分布的因素, 既包括古水温条件(以及轻微的与古盐度的相互补偿作用), 还有水体环境营养条件(图 10)。对大多数低纬度地区、盐度正常的浅水碳酸盐沉积来说, 骨屑颗粒组合成为分析水体营养条件和碳酸盐工厂的重要内容(Mutti and Hallock, 2003)。
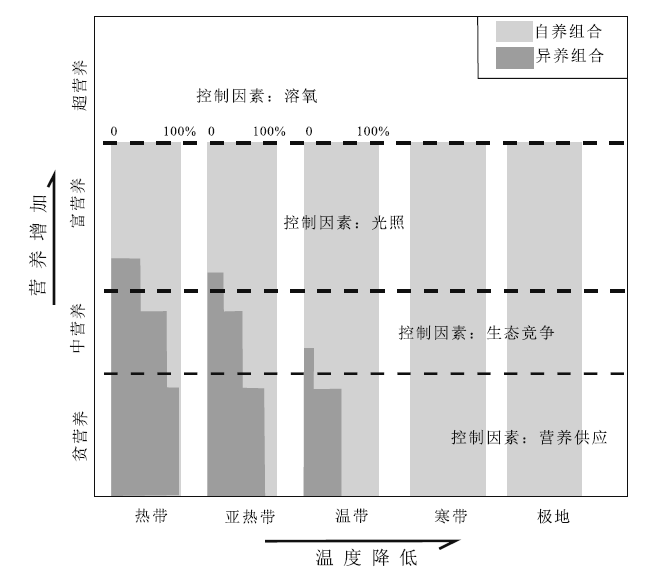 | 图 10 骨屑颗粒组合相对于温度分带和营养梯度的分布 (据Mutti and Hallock, 2003)Fig.10 Schematic representation of distribution of skeletal associations with respect to common temperature zones and nutrient gradient(after Mutti and Hallock, 2003) |
水体营养条件通过生物群落影响和控制碳酸盐沉积骨屑颗粒组合特征。如在尼加拉瓜上升流发育地区, 浅水珊瑚礁的生长适应贫营养的水体(每立方米水中叶绿素浓度不超过0.1 mg); 在中度营养的水中(每立方米水中的叶绿素浓度为0.1~0.2 mg), 占支配地位的是较大的藻类、海绵、软体动物和海胆类, 还有大量的生物侵蚀者, 生物钙化速率较低, 强烈的生物侵蚀抑制了网状生物骨架的生长, 并有可能致使台地边缘被淹没(在高产率的透光带之下); 富营养的水中(每立方米水中的叶绿素浓度大于0.2 mg), 较大的藻生长迅速; 超营养的水(每立方米水中的叶绿素浓度远远大于0.2 mg)则有利于异养生物发育, 钙化率较低。即使是短时间的富营养的上升流, 也可减弱碳酸盐产率(Hallock, 2001)。另外, 水体营养条件也影响了生物侵蚀和生物建隆的发育。
水体营养条件也是浅水碳酸盐岩台地发育的最为重要的影响因素。对当代印度洋— 太平洋和大西洋197个孤立碳酸盐岩台地的调查统计表明, 水体营养条件可能是台地面积的最主要控制因素。大多数碳酸盐岩台地发育在寡营养区域, 水体较浅, 富营养水体中缺乏大的碳酸盐岩台地。在地理分布上, 碳酸盐岩台地明显分为南北两群, 赤道地区和高纬度地区碳酸盐岩台地不发育, 或者面积极小(Vecsei, 2000)。
由于碳酸盐沉积的生产与生物有关, 地质历史中的生物大灭绝事件对碳酸盐岩台地的影响更是不容忽视的。古生代、中生代之交生物大灭绝事件是显生宙最大的生物— 环境事件, 也对全球碳酸盐岩台地演化产生了重要影响。二叠纪末期的高温、海洋缺氧、海洋酸化事件、生物礁生态体系的崩溃等(Clapham and Payne, 2011; Kiessling and Simpson, 2011; Hinojosa et al., 2012; Sun et al., 2012), 使得浅水碳酸盐工厂遭受重创(Pruss et al., 2006); 因而在中生代初, 生物控制的钙化作用让位于生物诱导的碳酸盐沉淀和灰泥丘沉淀, 以低级菌藻类为主的浅水生态系(以碎屑沉积为底质的藻席生态系和以碳酸盐沉积为基底的微生物岩生态系)极其繁盛(Xie et al., 2010; Chen and Benton, 2012)。上扬子地区早三叠世台地边缘缺乏生物建隆, 碳酸盐岩台地转变为典型的碳酸盐岩缓坡(Li et al., 2013, 2019)。
该生物大灭绝事件也直接导致了上扬子碳酸盐岩台地南部边缘在此期间快速向北退却。早三叠世, 海洋环境条件恶劣多变、生物复苏迟缓, 因此三叠纪初期上扬子碳酸盐岩沉积区地形起伏不大。随着生物复苏和环境条件的逐步改善, 早三叠世晚期碳酸盐岩台地也开始恢复, 台地边缘相带逐渐呈现。中三叠世初, 台地开始向外进积(Kelley et al., 2017)。在阿尔卑斯地区, 早三叠世仅发育少量孤立的碳酸盐建隆; 至中三叠世卡尼早期, 发育千米级的、高起伏的孤立台地或陆表海台地; 晚三叠世中后期, 大陆级的碳酸盐岩台地几乎遍布整个西特提斯地区, 很好地诠释了生物复苏与碳酸盐岩台地再造的过程(Caggiati et al., 2018)。
全球性古海洋事件同样对碳酸盐岩台地演化具有重要影响。白垩纪是联合古大陆加速分裂、洋中脊快速增生时期, 由此引发的全球高海平面和高大气二氧化碳浓度, 促进了碳酸盐岩台地的广泛发育; 同时也发育一系列显著的大洋缺氧事件和黑色页岩沉积, 最终导致特提斯洋周边海域的碳酸盐岩台地在白垩纪中期淹没(Phelps et al., 2015)。对应这些缺氧— 碳循环异常事件, 北美南部碳酸盐岩台地沉积相序列、动物群、沉积速率、台地内部结构以及相对海平面变化趋势, 呈现平衡期、危机期、缺氧期和台地恢复期4个阶段的特点:平衡期, 以珊瑚— 厚壳蛤光自养生物组合和进积沉积序列为特征。危机期虽然仍发育光自养生物,
但是生物钙化危机已经开始危及台地, 台地边缘转变为退积序列, 沉积物黏土矿物含量增加。在缺氧阶段, 发育富有机质黑色页岩背景沉积, 牡蛎类异养生物组合繁盛, 台地被淹没; 低碳酸盐产率导致沉积速率降低, 使得幅度仅30im左右的全球海平面变化在台地内部表现非常明显。在台地复苏阶段, 在海岸带开始重新建立异养生物构成的碳酸盐工厂, 并逐渐转变为进积缓坡沉积体系, 沉积速率随着贫氧状况的逐渐消失而缓慢增加(Phelps et al., 2015)。
在特提斯洋北部边缘的阿尔卑斯地区(瑞士), 碳酸盐岩台地在早白垩世也经历了3次以凝缩层、磷酸盐化和海绿石化作用为特征的台地淹没事件。古气候间歇性湿度变化以及随后的水体营养物质增加, 导致了碳酸盐工厂由光自养到异养的转变, 以及持续较长时间的台地淹没(Godet, 2013)。类似海侵及缺氧事件导致的台地淹没也发生在早侏罗世晚期(Toarcian), 如法国南部与缺氧事件相关的富营养事件— — 硬底构造、灰岩泥灰岩韵律、黑色富有机质泥灰岩等(Lé onide et al., 2012)。
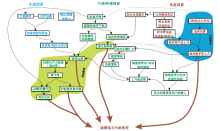
在碳酸盐沉积物的生产不能保持与可容空间的增加同步时, 碳酸盐岩台地形成淹没不整合, 是碳酸盐岩台地演化历史中的关键界面。除海平面上升外, 浅海生态系和浅水碳酸盐工厂的崩溃至关重要。白垩纪中期特提斯洋周缘碳酸盐岩台地对全球缺氧事件的响应具有典型的代表性。一方面大西洋快速打开、新的洋盆形成, 洋中脊增生以及海底火山高原的就位使得全球海平面上升; 另一方面, 洋中脊热液活动给深部海洋提供了丰富营养元素, 增加了水体营养。因此, 台地及台地淹没不整合面蕴含丰富的生物— 环境相互作用信息(Godet, 2013)(图 11)。由此也可以看出, 全球性生物大灭绝事件和环境条件恶化事件对碳酸盐工厂的影响, 或者说对碳酸盐岩台地的影响, 可能远远超过单纯性的海平面变化, 或者一般性沉积盆地基底热沉降。
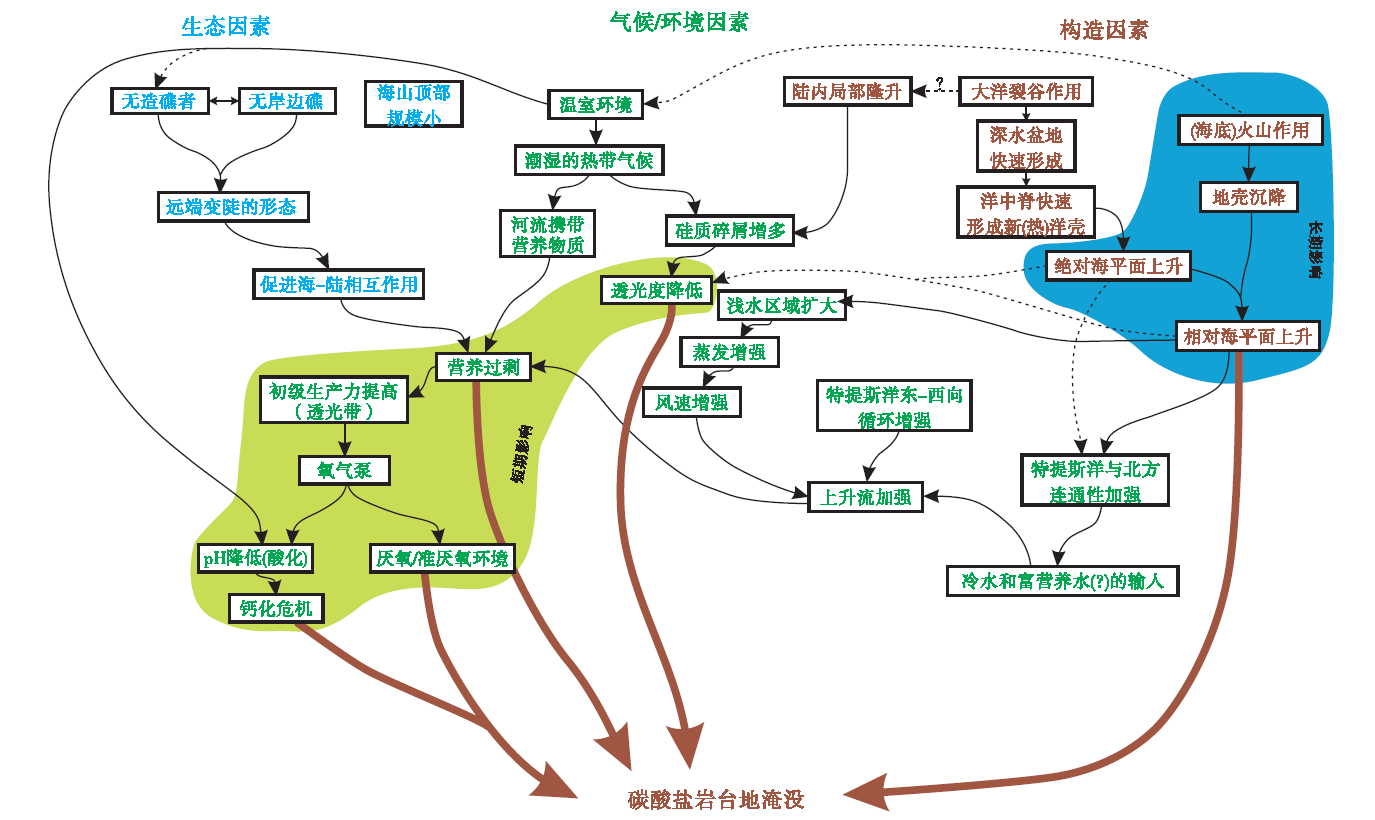 | 图 11 碳酸盐岩台地淹没成因反馈机制(据Godet, 2013)Fig.11 Feedback mechanisms of unfolding of carbonate platform drowning(after Godet, 2013) |
《地质历史中的碳酸盐岩相》一书(Wilson, 1975)的另一个巨大贡献, 是认识到地史时期浅水碳酸盐岩沉积相具有鲜明的时代印记, 如早石炭世的沃尔索蒂相、晚石炭世— 早二叠世叶状藻丘相等, 以及生物群落的重大变革对碳酸盐岩台地边缘生物建隆的控制作用。但是这方面似乎并没有引起人们足够的重视, 甚至是被遗忘的。从碳酸盐岩生物属性的角度来说, 地史时期生物和地球环境经历的显著变化, 势必导致碳酸盐岩沉积体系出现相应的变化。试图用一个沉积相模式概括所有地史时期的碳酸盐岩沉积相特征, 可能是造成上述忽略的原因之一。另一方面, 以目前对显生宙生物— 环境条件的了解和对显生宙浅水碳酸盐岩沉积资料的积累, 已经可以确切地说, 不是显生宙每一个碳酸盐岩沉积体系都特殊到需要一个独立模型来刻画的地步, 碳酸盐岩台地类型的数目是有限的和可知的(Markello et al., 2008)。
碳酸盐岩地层的时代特殊性(age-sensitive pattern or theme)就是特定时间段内生物— 环境相互作用留下的印记。碳酸盐岩中巨大的油气资源潜力给碳酸盐岩沉积学进展带来了巨大动力, 但是碳酸盐岩地层结构和成因的复杂性也给油气资源勘探、开发和生产带来挑战。根据已有认识和实例, 找到适合油气勘探、开发和生产的碳酸盐岩储层模型, 是油气工业的现实需要。区分这些特定的时间间隔、识别该时段内碳酸盐岩沉积系统的关键特征、解读其成因联系, 尽可能使储层预测模型的概念、原理和特征更加适用, 对油气工业来说具有非常重要的实践意义。地史时期碳酸盐岩类比计划(The Carbonate Analogs Through Time, CATT), 就是这样一项围绕显生宙碳酸盐岩沉积体系和储集层特征的宏伟研究计划。CATT计划希望通过总结地史时期碳酸盐岩形成过程和地球演化过程中的环境条件, 构建一系列高可信度、具有时代特征的针对碳酸盐岩沉积系统中储集层构成、位置、地层属性和储层特性的预测模型, 或者称之为样式(age-sensitive patterns or themes)(Markello et al., 2008)。计划的核心思想包括: (1)在显生宙期间碳酸盐岩形成过程(生物演化和原生矿物演化等)和地球过程(大地构造、古气候、海平面变化、海洋化学和海洋循环)经历了哪些显著变化; (2)在生物演化和地球环境变化的相互作用下, 显生宙特定时期碳酸盐岩成因具有哪些可类比的、独特的特征。
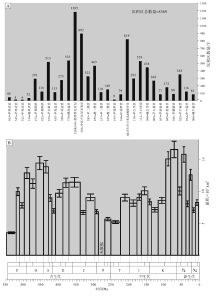
研究计划的基础是全球显生宙古地理图集。考虑到可操作性, CATT依据二级层序概念和划分, 确定出29个古地理编图时间单元(时长5~20 Ma, 平均10 Ma), 以海泛面成图, 圈定了全球显生宙8369个碳酸盐岩沉积区(carbonate field)(图 12-A)。目前项目已经完成(1)显生宙碳酸盐岩台地演化趋势表(图 12-B)和(2)全球碳酸盐岩分布区图集(Markello et al., 2008)。当然, 此项工作仍需要大量人力物力投入才能得以完成。初步成果表明, 全球显生宙碳酸盐岩沉积区的时空分布既不是单一规律控制的, 也不是随机分布的, 因此地史时期的大套碳酸盐岩地层, 不再是孤立和零散的沉积地质体。查明全球显生宙碳酸盐岩地层时空分布格局的工作已经开始, 也将为后续深入研究奠定坚实的基础。
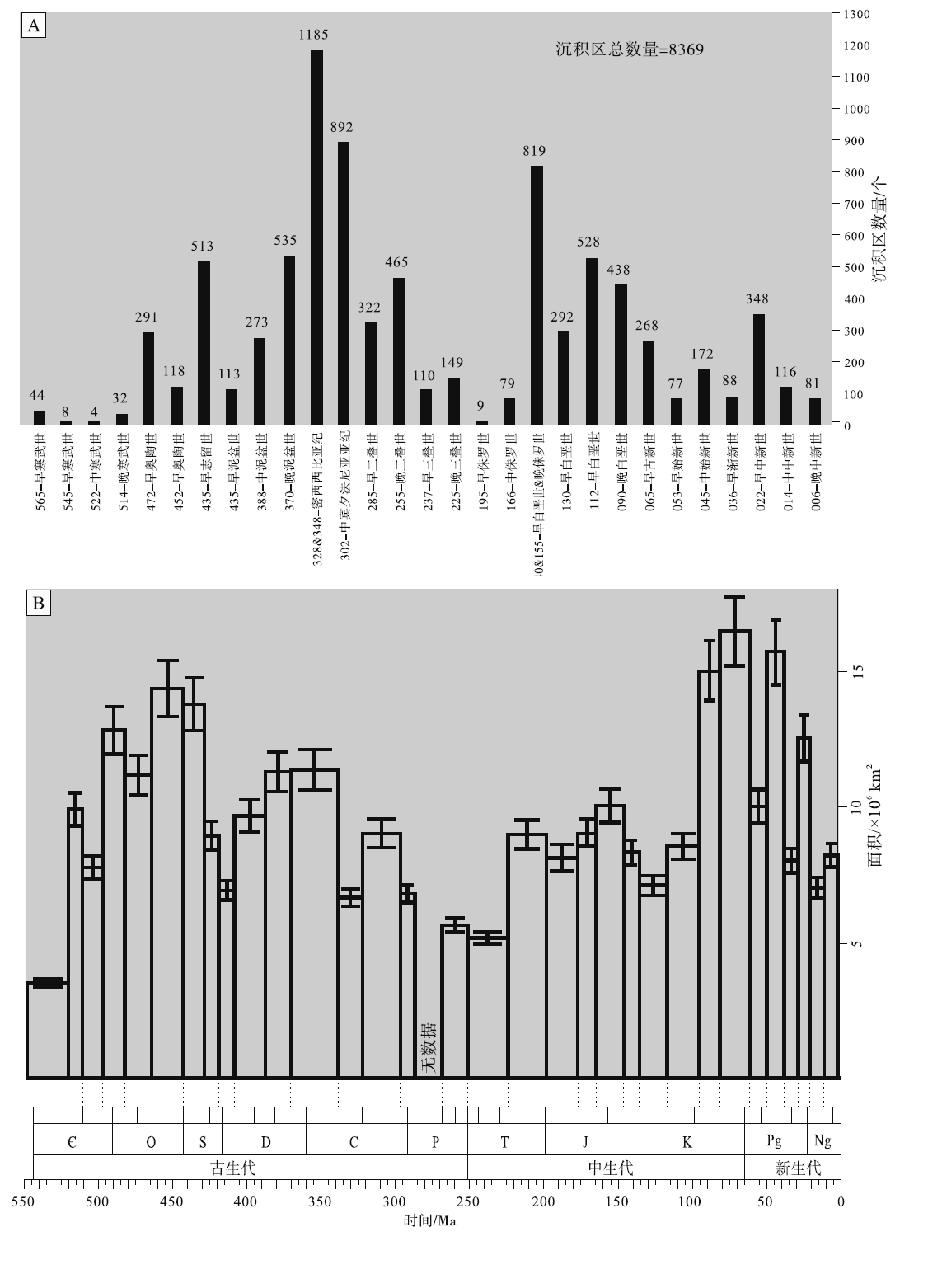 | 图 12 显生宙不同时期碳酸盐岩沉积区频度分布(A)(据Markello et al., 2008)和碳酸盐岩台地面积分布(B)(误差条表示基于总值统计的95%的置信区间, 据Kiessling et al., 2003)Fig.12 Frequency distribution by age of carbonate fields (A)(after Markello et al., 2008) and area distribution by age of carbonate platforms (B) for the Phanerozoic(error bars indicate statistical 95% confidence intervals based on total values, after Kiessling et al., 2003) |
碳酸盐岩地层鲜明的时代特征, 有些类似于沉积学中的“ 时代特征相” (Time-specific facies, TSF), 只不过TSF包括非碳酸盐岩等其他沉积物(Brett et al., 2012)。很多碳酸盐岩沉积特征已经作为TSF得到重点关注, 如“ 方解石海” 、“ 文石海” 、海洋缺氧事件(OAEs)、“ 缺礁(reef gap)” 、“ 缺硅(chert gap)” 等。与TSF相关的另一种广受关注的是在显生宙生物大灭绝之后出现的错时相沉积, 如扁平砾石、皱饰构造、巨鲕等(Li et al., 2013)。大量的碳酸盐岩地层中的时代特征相, 还有待发掘和重视。
碳酸盐岩地层的时代特征甚至可以影响到成岩作用。如北美西部加利福尼亚地区新元古界— 寒武系界线附近鲕状白云岩的成岩组构存在明显差别。新元古代晚期地层白云岩化发生在早期成岩作用阶段, 白云岩化作用彻底且较好地保留了原生矿物组构, 而下寒武统白云岩化作用发生在埋藏成岩作用过程中, 白云岩中的原生矿物和结构信息保存较差。这种成岩作用特征的差别与新元古代末期— 寒武纪早期海洋Mg/Ca值变化有关(Corsetti et al., 2006)。类似现象既具有科学意义, 也具有实践意义, 如上述CATT计划所针对的储集层特征。
依据“ 碳酸盐工厂” 特征, 分析其形成过程中的生物与环境相互作用, 已经成为当前碳酸盐岩台地分类、台地内部结构成因解释、沉积相带展布预测的重要方法(Pomar and Ward, 1995; Insalaco et al., 2000; Pomar, 2001b; Kiessling et al., 2003; Wright and Burgess, 2005; Castell et al., 2007; Schlager and Keim, 2009; Williams et al., 2011; Chesnel et al., 2016; Ł abaj and Pratt, 2016; Lü dmann et al., 2016; Kleipool et al., 2017), 但是在此全面系统介绍和讨论现有浅水碳酸盐岩台地成因研究显然不现实。下面以滇黔桂地区为例, 从碳酸盐工厂分析角度, 剖析该区二叠纪吴家坪期碳酸盐岩沉积学研究现状以及值得进一步研究的问题, 在现今中国碳酸盐岩沉积古地理研究中有一定代表性, 或许会有启发意义。
华南晚古生代碳酸盐岩台地, 尤其是二叠纪的碳酸盐岩台地, 是中国浅水碳酸盐岩沉积相(包括生物礁)模式的发源地(范嘉松等, 1979; 关士聪等, 1980), 对中国碳酸盐岩沉积学和生物礁研究的发展发挥了关键性的示范作用。相关碳酸盐岩地层, 也是中国晚古生代地层学、沉积古地理学等相关学科研究的重要基地。经过多轮古地理编图研究, 华南地区二叠纪沉积古地理轮廓已经较为清楚, 但是如何开展下一步深入研究, 明显缺乏清晰的思路。在晚二叠世吴家坪期, 这种情形更加明显。当时碳酸盐岩台地规模相对较小, 加上同期陆源碎屑岩及含煤沉积的广泛发育, 与其下伏和上覆地层相比, 吴家坪期碳酸盐岩地层沉积学研究几乎处于被忽略的位置。
首先, 吴家坪期碳酸盐岩台地的一些基本问题亟待解决, 如台地边缘的位置。目前广泛认为二叠纪上扬子碳酸盐岩台地南部边缘位于册亨— 望谟— 紫云— 罗甸一带, 但是该格局体现的主要是茅口早期— 中期的情形。随着台地在茅口晚期的萎缩, 台地边缘在向北退缩, 到三叠纪(甚至早至长兴期)台地边缘已经向北退缩至福泉— 青岩— 镇宁— 贞丰— 兴义南一带(贺自爱等, 1980; 1981; 刘宝珺和许效松, 1994)。若仍沿用茅口早期数据推定吴家坪期台地边缘的位置, 显然不合适, 需进一步明确。
其次, 滇黔桂地区吴家坪期碳酸盐岩台地的类型。由于吴家坪期极少发育生物礁, 同期上扬子碳酸盐岩台地南部主体肯定不是镶边台地。二叠纪是全球生物礁发育历史中的一个高峰时期, 但是吴家坪期是其中的一个低谷期(Flü gel and Kiessling, 2002; Kiessling and Flü gel, 2002; Weidlich, 2002; Alvaro et al., 2007)。国外发育较好、研究程度较高的吴家坪期生物礁屈指可数。华南地区虽然有文献记载的吴家坪期生物礁发育地点较多, 如广西西林、平果、天峨、田东、乐业、来宾和贵州紫云等(贺自爱等, 1981; 周怀玲和张振贤, 1995; 周怀玲等, 2014), 但经他人研究证实的生物礁只有广西来宾铁桥剖面(邓占球, 1981; 杨万容, 1987; Huang et al., 2017)。贵州紫云吴家坪期“ 生物礁” 可能只是一些珊瑚层(王生海和范嘉松, 1996), 其吴家坪期“ 生物礁” 属性并未得到后续研究证实(如吴亚生等, 2003; Shen and Xu, 2005)。显然, 缺少台地边缘生物建隆, 滇黔桂地区吴家坪期碳酸盐岩台地类型和特征需要重新认识。
第三, 台地地层结构和成因。二叠纪滇黔桂地区发育多个孤立碳酸盐岩台地, 大部分台地在吴家坪早期被淹没, 如在广西的东兰— 巴马— 凤山台地(刘超, 2017)、乐业台地(王新强和史晓颖, 2008)和来宾— 合山台地东部(姚尧等, 2012)。在吴家坪期中— 晚期, 碳酸盐岩台地上呈现进积序列, 台地范围有逐渐向外扩展的趋势。
现有研究较为注重盆地演化的构造背景。滇黔桂地区或者右江盆地在二叠纪期间为伸展性区域构造背景(如杜远生等, 2013; Xu et al., 2017; Cawood et al., 2018)。断块式差异升降无疑为右江盆地内孤立碳酸盐岩台地发育提供了有利的区域构造条件; 区域性盆地基底下沉与海平面上升的结合, 看似合理地解释碳酸盐岩台地在吴家坪早期的淹没及后续发展, 细究则可以发现诸多不如意之处, 情形并非如此简单。如乐业台地, 在茅口晚期经历了东吴运动期间的上升剥蚀, 吴家坪组底部发育1层厚度约1 m的铝土质沉积; 之后快速海侵, 几乎在原来(茅口期)台地的中央位置出现泥灰岩和薄层泥晶灰岩, 向外可以相变为薄层硅质岩, 向上逐渐过渡到中层— 厚层灰岩。可以明显感受到浅水碳酸盐工厂在台地顶部的重建和向外扩展, 但是察觉不到控制台地边缘的“ 同沉积断裂” 的存在或者活动; 整个台地出现上升剥蚀、下降淹没, 之后又出现台地明显上升, 既不是台地沉降可以解释的, 也非简单海平面波动变化可以解释的。类似现象同样出现在上扬子碳酸盐岩台地南部和北部边缘地区。
另一方面, 从显生宙地球环境和生命演化角度看, 茅口中— 晚期到吴家坪早期既是一个重要变革时期, 也是一个重要地质事件多发期, 包括峨眉山大火成岩省活动、全球缺氧事件和前乐平世生物灭绝事件(Stanley and Yang, 1994; Wignall, 2001)。以后者最为显著且具全球性影响, 殃及了65%~71%的海生物种(或58%属)(Erwin et al., 2002; Racki and Wignall, 2005; Ota and Isozaki, 2006; Sahney and Benton, 2008; Bond et al., 2010), 其生态危机程度在显生宙主要生物灭绝事件中甚至可以位列第三, 仅次于二叠纪末和白垩纪末的生物大灭绝(Bambach et al., 2004; McGhee et al., 2013; 戎嘉余和黄冰, 2014); 陆生四足类动物的74%~80%灭绝, 灾难程度超过二叠纪末期的生物大灭绝事件(Day et al., 2015)。由此可见, 前乐平生物灭绝事件本身足以对当时的海洋生态系统产生重要影响。伴随生物大灭绝事件的还有海洋环境恶化, 如来宾地区和北美地区硫同位素分析结果等表明, 茅口晚期— 吴家坪早期全球古海洋环境发生过硫化水体的上涌事件(Zhang et al., 2015)。
前乐平生物灭绝事件的最大特征是重创了海洋光合作用生物(包括营光合作用的生物和共生有能进行光合作用微生物的生物)(Weidlich and Bernecker, 2003; Vachard et al., 2010; Groves and Wang, 2013; Bond et al., 2017), 如钙质藻类、(达88%物种)(杨湘宁等, 2000; Groves and Wang, 2013)、造礁珊瑚以及巨型双壳翅蛤类(Alatoconchids)(Chen et al., 2018), 对热带浅水碳酸盐工厂造成了灾难性影响。当时, 浅水生物礁碳酸盐沉积生产损失了88.7%(Flü gel and Kiessling, 2002)。
通过对黔南罗甸纳水和沫阳2个代表性剖面茅口组骨屑颗粒组合统计分析和碳酸盐生产工厂分析, 孟琦等(2018)发现黔南茅口晚期经历了由热带浅水、暖水碳酸盐生产工厂到热带较深水、温凉水工厂的明显转变, 而且这一碳酸盐工厂的转变现象在华南地区普遍存在, 与当时热带浅海碳酸盐工厂受抑制或被破坏有关。在中东阿拉伯板块中— 晚二叠世碳酸盐岩地层序列中, 碳酸盐工厂由中二叠世的生物诱导为主的碳酸盐工厂迅速转变为晚二叠世钙质鲕粒工厂, 与二叠纪末期生物大灭绝事件之后普遍出现巨型钙质鲕粒沉积极为相似(Li et al., 2015), 印证了前乐平生物灭绝事件前后碳酸盐工厂的巨大转变。
因此, 从地球环境和生物演化角度看, 滇黔桂地区(或者华南地区)吴家坪期碳酸盐岩台地发育历史记录了前乐平生物大灭绝事件之后, 也就是台地在中二叠世后期遭到重创之后的生物复苏、碳酸盐工厂调整、台地重建和增长的过程, 是华南晚古生代碳酸盐岩台地演化历史中的重要一环。如此解释上述台地地层结构似乎更加合理。由此可见, 仔细审视古地理图上那些习以为常的“ 台地边缘断裂带” 的“ 差异升降” , 合理评估碳酸盐生产和堆积对沉积地形差异的贡献, 值得在进一步碳酸盐岩沉积古地理研究中加以重视。
第四, 深入研究滇黔桂地区吴家坪期碳酸盐岩台地地层结构及空间展布特征, 对认识当时全球生物和海洋环境也具有非常重要意义。人类正在渴望了解生物在环境恶化后复苏的过程和控制因素。在大灭绝事件后, 生物复苏是生物生命演化进程中不可缺少的重要环节之一, 是一次新的生物辐射的前奏, 也是当前古生物学研究的前沿(戎嘉余和方宗杰, 2004)。生物灭绝事件特征和过程的差别也会体现在生物复苏的过程中, 在生物大灭绝成因机制研究中已开始重视事件后的生物复苏(戎嘉余和方宗杰, 2004)。全面了解生物大灭绝后生态系的组成和结构, 对于正确认识地史转折期, 即圈层耦合结点处的地质作用型式和过程具有重要作用(童金南, 1997; 殷鸿福和童金南, 1997)。
相比传统五大生物大灭绝事件后的生物复苏研究, 前乐平事件后的生物复苏研究成果仍然较少(如Sahney and Benton, 2008; Shen and Zhang, 2008; Wignall et al., 2012)。另外, 还需要注意不同门类生物的复苏过程的明显差别, 如腕足类在吴家坪早期就已经复苏(Shen and Zhang, 2008), 生物礁则可能较晚, 四足类动物的复苏则推迟至晚二叠世末期。灾后的生态系重建是一个从简单到复杂(稳定)、从食物链的底层到顶层的逐步过程, 需要系统地研究(Song et al., 2011; Chen and Benton, 2012)。热带浅水碳酸盐沉积或者工厂是生态系统整体生物面貌的反映, 尤其是食物链金字塔相对靠下层的生物面貌, 因为对碳酸盐沉积物生产贡献最大的生物往往是那些代表着与初级生产力相关的、形态结构相对简单、能够快速繁殖生长的类型, 如石炭— 二叠纪各种钙质藻、中生代的微型浮游生物(如颗石藻)(Stanley, 2006)。碳酸盐岩台地边缘的进积/退积, 连接着碳酸盐岩台地顶部碳酸盐工厂的盛衰, 蕴含着大量生态系底层和中下层生物的整体信息, 是研究生态系复苏的极好素材。
因此, 吴家坪期碳酸盐岩台地边缘演化所承载的不仅是热带浅水碳酸盐岩沉积学问题, 而且其演化必然与当时的(台地)碳酸盐工厂密切相关, 即与当时的生物和海洋环境息息相关。隐藏在吴家坪期碳酸盐岩台地内的热带浅水碳酸盐工厂的重建与增长过程信息, 对深入认识前乐平生物灭绝事件之后的全球生物复苏过程、探讨当时生物与环境之间的相互作用等将具有重要意义。
滇黔桂地区晚二叠世吴家坪期浅水碳酸盐岩台地的实例有一定的代表性, 也有其局限性, 但是其中所涉及到的分析思路值得后续研究者借鉴。如重视碳酸盐岩的生物属性, 重新认识生物和海洋环境条件在碳酸盐岩台地演化中的作用; 针对浅水碳酸盐岩沉积, 视域不能局限于沉积体所处的地理位置和单个盆地范围, 需要从地球环境和生物相互作用维度解读其时代特殊性, 在全球空间尺度范围内, 比较同期碳酸盐岩的相似性及差异, 了解研究对象所处的地球环境— 生物背景; 从生态角度入手开展碳酸盐工厂分析, 来深入剖析碳酸盐岩台地的成因, 发掘蕴藏在碳酸盐岩台地内部的生物学和海洋学意义。
致谢 冯增昭老师等4位审稿专家对本文初稿提出了许多好的、中肯的修改建议, 为本文增色, 在此致以诚挚谢意!
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|