






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
光合作用生物膜建造的凝块: 来自于辽东半岛芙蓉统长山组凝块石生物丘中的一些证据*
[梅冥相1, 2  , Khalid Latif
, Khalid Latif1 , 刘丽1 , 孟庆芬1 ]
, Khalid Latif|
|

第一作者简介 梅冥相,男,1965年生,教授,博士生导师,主要从事沉积学和地层学研究工作。E-mail: meimingxiang@263.net。
作为一种以非纹层状组构而区别于叠层石的微生物碳酸盐岩类型,中凝块( mesoclots)是产生凝块石凝块组构关键的微结构。构成这种中凝块的离散性凝块状泥晶,似乎代表了细胞外聚合物质( EPS)复杂的钙化作用,但是,由于常常缺乏直接的微生物证据,造成其所涉及的准确成因和特殊过程还不太清楚。在辽东半岛大连金州湾和本溪田师傅剖面芙蓉统下部的长山组顶部,发育较为典型的凝块石生物丘,组成三级层序的强迫型海退体系域沉积,代表着较为壮观的沉积学现象;在这些凝块石生物丘中,毫米级别大小的暗色离散性凝块状泥晶是其最为基本的沉积组构,而且在这些凝块之中以较高密度保存的丝状蓝细菌鞘化石反映的微生物构成,意味着它们具有明显的光合作用生物膜成因。尽管穿越成岩作用过滤器去解释古代凝块石复杂的形成机理将存在着巨大的挑战,也尽管形成这些凝块石复杂的生物膜钙化作用细节需要更加深入的研究才能得到更好的了解,但是,辽东半岛芙蓉统长山组的凝块石,尤其是在凝块内较为丰富的钙化蓝细菌鞘化石所表征的直接的微生物活动证据,使其成为一个了解光合作用生物膜建造凝块的典型实例;再者,与凝块共生的三叶虫化石、底栖鲕粒和皮壳粒,以及围绕着这些多样化颗粒特别的放射纤维状方解石结壳,说明这些凝块石可能属于较厚的微生物席内多重状生物膜以及构成这些微生物席和微生物膜的细胞外聚合物质复杂的钙化作用建造物。
About the first author Mei Ming-Xiang,born in 1965, graduated from China University of Geosciences in 1993,and obtained Ph.D. degree. Now he is a professor at School of Earth Sciences and Resource,China University of Geosciences(Beijing),and is engaged in sedimentology and stratigraphy. E-mail: meimingxiang@263.net.
Thrombolites are marked by non-layer fabrics,which are distinct from stromatolites;the key microstructure of thrombolites are mesoclots that produce clotted fabrics. Diffusely clotted micrite that commonly make up the mesoclots likely represent extracelluar polymeric substances(EPSs)calcification. However,its precise origin and formative processes are not clear due to absence of direct evidence of microbial activities. Thrombolite bioherms occur in the top of the Changshan Formation of the Cambrian Furongian at the Jinzhouwan and Tianshifu sections in the Liaodong Peninsula. Thrombolite bioherms were deposited in the forced regressive system tract of a third-order depositional sequence. Diffusely clotted micrites with the size of micrometers are fundamental fabrics of these thrombolites. Importantly and particularly,the sheath fossils of filamental cyanobateria are preserved,suggesting a biological origin of photosynthetic biofilms. A great challenge is to interpret the ancient clots and their detail formative processes through the filter of diagenesis. Calcification of cyanobacteria biofilms also remains poorly understood. The preservation of cyanobacteria sheath fossils within thrombolite colts in the Furongian Changshan Formation in the Liaodong Peninsula suggests that the colts were formed by photosynthetic biofilms. Furthermore,the clots within thrombolites are commonly symbiosis with other types of carbonate grains such as benthic ooid,cortoid,and trilobite fossil fragment,as well as radiated calcite fabric around these carbonate grains,which indicate that the thrombolites of the Changshan Formation probably resulted from complex calcification of EPSs of multiple biofilms within the relative thick microbial mat.
凝块石(thrombolite)被认为是一种微生物岩(Burne and Moore, 1987)或微生物碳酸盐岩(Riding, 2000)。Aitken(1967)首先定义和介绍了凝块石, 其被用来指那些“ 与叠层石相关但是缺乏纹层的隐藻(cryptalgal)构造, 而且以宏观上的凝块状组构为特征” , 他还将构成凝块石特征性的中等尺度的构造要素称为凝块(clot)。随着研究的深入, 凝块石被Shapiro(2000)较为确切地定义为“ 凝块状中凝块(mesoclots)组成的微生物岩” , 而中凝块为多形态的(简单球形到多叶片状)、毫米到厘米级别大小的聚合体所组成的离散性凝块状泥晶(Riding, 2000), 为凝块石的关键性组构(Shapiro, 2000; Theisen and Sumner, 2016)。但是, 要注意的是, 凝块还可以作为凝聚碳酸盐颗粒的一种类型(Tucker and Wright, 1990; 梅冥相等, 1997; Flü gel, 2010), 而且在许多凝块石中除了中凝块以外, 常常还发育着其他类型的碳酸盐颗粒。再者, 凝块石结构存在着一个非常宽广的范围, 可以归因为多样化的过程, 这也导致了将凝块石解释为一个微生物碳酸盐岩组合存在困难(Theisen and Sumner, 2016)。
凝块石曾经被认为等同于花纹石(catagraphite), 并认为微植石(microphytolite)是核形石和花纹石的总称, 同时古生物学家也曾经采用林奈法将凝块石划分为多孔状凝块石(Vesicularites等)、凝块状凝块石(Vermiculites等)、网状凝块石(Maecovella)(详见杜汝霖(1992)的总结)。但是, 随着蓝绿藻的生物学属性被修订为蓝细菌(梅冥相, 2007a, 2007b, 2011a, 2012a, 2014; 梅冥相和孟庆芬, 2016; 梅冥相等, 2017), 就像早年的“ 隐藻(cryptalgal)” 概念一样, “ 微植石(Microphytolite)” 的概念应该抛弃。
凝块石的凝块状组构具有各种各样的成因。 (1)通过对现代巴哈马叠层石— 凝块石组合进行研究, 认为凝块石构成的柱状体是由于物理作用和后生动物破坏所造成的, 同时还包括准同生矿化作用、次生胶结作用和局部的碳酸盐溶解作用(Feldmann and Mckenzie, 1998; Planavsky and Ginsburg, 2009); (2)被归因为“ 球状和球状所主导的微生物群落原地钙化作用” , 即叠层石与凝块石的基本差异是前者为丝状细菌所构建, 而后者为球形细菌所构建(Kennard and James, 1986; Flü gel, 2010; Gerdes, 2010)。尽管这一认识符合前寒武纪凝块石的特征(Mei and Tucker, 2013; Tang et al., 2013; 梅冥相, 2014), 但似乎得不到本文将要展示的证据和资料的支持; (3)第3种成因为钙化的丝状体, 诸如葛万菌, 改变了微生物碳酸盐岩的组构, 瓦解了叠层石的纹层化作用过程并促进了凝块石的宏观凝块组构的发育(Riding, 2011)。中元古代早期的凝块石(Mei and Tucker, 2013; Tang et al., 2013; 梅冥相, 2014)又将这一认识复杂化; (4)其他的机制, 涉及到生物扰动构造和生物侵蚀作用, 而且那些大型的巴哈马叠层石中的凝块石的中等尺度构造特征, 常为真核生物繁殖所造成的纹层破坏所致(Dupraz et al., 2009, 2011), 而这样的特征也在寒武纪的凝块石中被观察到(Theisen and Sumner, 2016)。虽然目前取得了一些重要的认识, 即重复性的纹层状沉积被归为叠层石, 而那些非纹层状的凝块状构造则被定义为凝块石(Kennard and James, 1986; Burne and Moore, 1987), 且构成凝块石的中凝块的离散性凝块状泥晶似乎形成于细胞外聚合物质(EPS; Decho, 2010; Flemming and Wingender, 2010; Tourney and Ngwenya, 2014; Flemming et al., 2016; Decho and Gutierrez, 2017)复杂的钙化作用(Riding, 2000, 2011), 但是由于常常缺乏直接的微生物证据与叠层石相对照, 同时凝块石还表现出更高程度的、更加复杂的碳酸盐沉淀作用的非均质性和变化性, 所以说, 这些全然不同的微生物岩组构, 它们精确的微生物机制和过程几乎为一个未知领域(Mobberley et al., 2013, 2015)。
近年来, 针对形成凝块石的微生物席的培养试验和现代实例的研究取得了一些重要的新成果和新认识, 包括: (1)在特别的凝块石方解石沉淀作用中, 表现出一些生物膜的功能与形态, 一系列微生物基团发育而成的多孔状微组构将沉淀物组织化, 这就会在生物膜内产生特征性的凝块石组构, 而且这样的组构明显不同于纹层状的叠层石(Pedley, 2014); (2)形成凝块石的微生物席群落存在着新陈代谢活动的垂向梯度, 而且凝块石还可代表特征性的基因群落(Mobberley et al., 2013, 2015); (3)在西澳大利亚Clifton湖现代凝块石微生物岩研究中取得了1个有意义的发现, 即依据(结晶结构)三八面体的蒙脱石矿物硅镁石的生物膜矿化作用获得了它们的初始构造的坚固性(Burne et al., 2014); (4)生物膜反而形成纹层发育较好的叠层石, 而那些较厚的微生物席则产生粗糙的纹层到高孔隙的不规则状微生物岩, 如凝块石(Arp et al., 2012); (5)球状蓝细菌与丝状蓝细菌如双须菌(Dichothrix)的协调活动, 将有助于在形成凝块石的微生物席内开启碳酸盐沉淀作用(Planavsky et al., 2009; Louyakis et al., 2017, 2018)。
在微生物礁的概念框架下, Lee 等(2011)对山东寒武系第二统的凝块石及其钙化微生物构成进行了系统的描述与研究; 更为引人关注的是, 山东西部寒武系苗岭统中附枝菌(Epiphyton)形成的特别的凝块石, 得到了许多学者较为系统的研究和关注(Woo et al., 2008; Woo and Chough, 2010; 陈金勇等, 2014a, 2014b; 贡芸芸, 2016; Ezaki et al., 2017; Yan et al., 2017)。这些苗岭统凝块石中主要为附枝菌构成的凝块, 不同于离散性凝块状泥晶构成的中凝块Aitken, 1967; Kennard and James, 1986; Burne and Moore, 1987; Shapiro, 2000)或凝块(Riding, 2000, 2011; Dupraz et al., 2011), 也与Shapiro(2000)的凝块石定义(凝块状中凝块组成的微生物岩)不相符合, 但似乎更加符合树形石的定义(Riding, 2000, 2011; Rowland and Shapiro, 2002; 梅冥相等, 2007a, 2007b; Dupraz et al., 2011), 因为树形石被定义为是一种具有孤立的树形状钙化微生物的束状体所主导的非纹层状中等尺度的构造(Rowland and Shapiro, 2002), 而且具有宏观的厘米级别的灌木状组构, 是较为典型的由钙化微生物(主要是附枝菌)所产生的微生物碳酸盐岩(Riding, 2000, 2011; Dupraz et al., 2011; Howell et al., 2011)。上述这些不同的认识也是正常的现象, 就像树形石被进一步定义和描述为“ 附枝菌” 格架岩(Woo et al., 2008; 韩作振等, 2009; Woo and Chough, 2010)或树枝状凝块石(齐永安等, 2014, 2017a, 2017b)那样, 同时也说明了凝块状体作为一种特征是很宽泛的, 要比纹层状的叠层石体更加不容易定义。因此, 虽然术语“ 凝块石” 就像叠层石那样已变得尽人皆知, 但是关于它的组构类型及其所包含的成因还存在一些混乱(Riding, 2000, 2011)。另外, 与其他类型的微生物碳酸盐岩一样, 凝块石也是最难研究的, 因为其多为微生物的影响或作用所形成的, 而且不总是保存直接的或坚固的成岩作用形成样式的证据。
辽东半岛寒武系芙蓉统长山组顶部块状凝块石灰岩组成的生物丘集中发育在三级层序的顶部, 成为一种时间特化的相, 构成特殊的强迫型海退体系域沉积(梅冥相, 2010)。特别的宏观产出特征, 与特别的微观现象一起, 尤其是在凝块石内部的凝块中普遍发育的钙化蓝细菌化石(Riding, 1991, 2000), 意味着构成凝块石的凝块形成于较为典型的光合作用生物膜, 从而成为一个较为壮观的沉积学现象, 为了解形成凝块复杂的生物膜钙化作用(Decho, 2010; Flemming and Wingender, 2010; Flemming et al., 2016; Decho and Gutierrez, 2017)提供了一个较为典型的实例; 同时, 在这些凝块石内部, 与凝块共生的底栖鲕粒(Gerdes et al., 1994; 梅冥相等, 2011, 2015; 梅冥相, 2012b; 倪胜利, 2017)和围绕着三叶虫化石与凝块的放射纤维状方解石的沉淀与结壳作用, 也为了解较厚的微生物席内(Gerdes et al., 1994; Gerdes, 2010; 梅冥相, 2011a, 2014; Arp et al., 2012; De los Rí os et al., 2015)多重状微生物膜复杂的钙化作用及其诱发的微生物沉淀作用提供了一个较为典型的实例, 尽管许多细节问题还有待进一步研究。
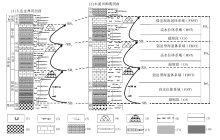
华北地台大致在寒武纪第二世晚期开始接受沉积, 而且寒武纪第二世的地层超覆在不同时代的前寒武纪地层之上, 在二者之间形成一个类似于北美地台的“ 巨型不整合面” (Peters and Gaines, 2012)。所以, 华北地台寒武系形成一个特别的沉积序列(图 1): 从寒武纪第二世红层与碳酸盐岩混合沉积(冯增昭等, 1990, 2004; Meng et al., 1997; Lee and Chough, 2011)开始, 经苗岭世鲕粒滩主导的碳酸盐台地序列(马永生等, 2017), 变化到芙蓉世碳酸盐泥所主导的碳酸盐台地沉积序列(梅冥相等, 2017)。
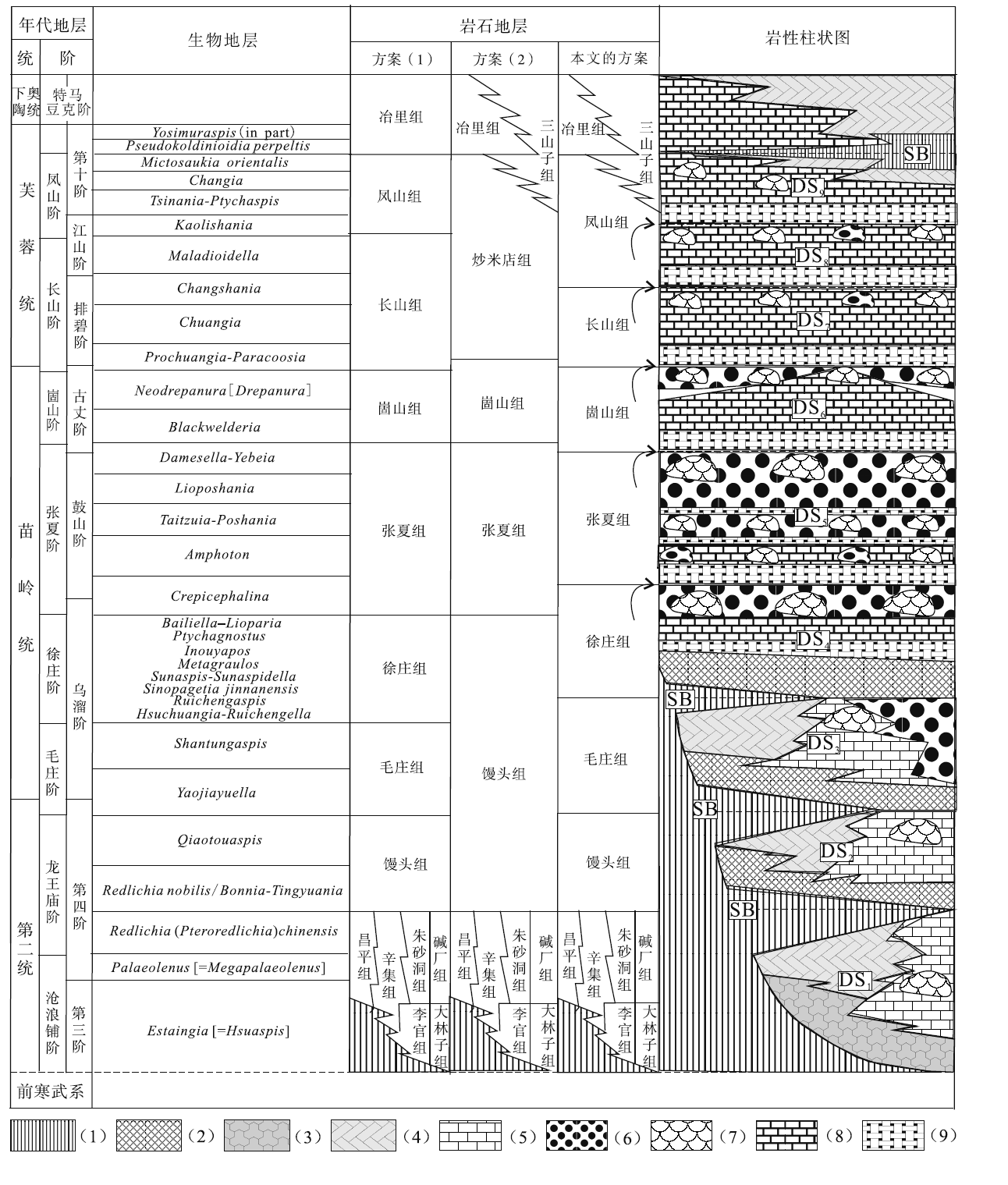 | 图 1 华北地台寒武纪沉积序列 三叶虫生物地层序列引自彭善池(2009); 岩石地层划分方案(1)引自冯增昭等(1990, 2004), 卢衍豪等(1994)、Meng 等(1997), 方案(2)引自项礼文等(1999), 本文采用的方案沿用方案(1)并基于沉积相序列的层序地层划分予以修改, 即华北地台寒武系大致可以划分为9个三级层序(DS1至DS9), 包含着两种类型的层序界面即暴露间断面构成的层序界面(SB)和淹没不整合型层序界面(箭头所指)。图例的含义分别是: (1)地层缺失, (2)海岸萨布哈相红层泥岩地层, (3)潮上砂泥坪相砂泥岩地层, (4)潮坪相白云岩地层, (5)浅缓坡相灰岩地层, (6)鲕粒滩相灰岩地层, (7)微生物碳酸盐岩生物丘, (8)中至深缓坡相泥灰岩和灰岩地层, (9)陆棚相钙质泥岩Fig.1 Sedimentary succession of the Cambrian in North China Platform |
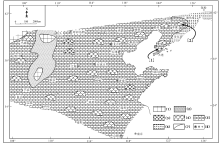
在芙蓉世早期的长山组沉积时期, 围绕着西北部的小片古陆, 在华北地台形成一个碳酸盐泥所主导的缓坡型台地沉积(冯增昭等, 1990, 2004; 梅冥相等, 2011, 2015, 2017; Pratt et al., 2012), 与下伏寒武纪第三世鲕粒主导的碳酸盐台地沉积(梅冥相, 2011b; 陈小炜等, 2013; 马永生等, 2017)形成明显的差异。最为引人注目的是, 在华北地台芙蓉世早期长山组沉积时期的沉积格局中(图 2), 从缓坡(浅海台地)相灰岩一直到较深水的浅海陆棚相钙质泥岩地层中, 均生长和发育着由大大小小的微生物碳酸盐岩主导的生物丘(梅冥相, 2011b; 梅冥相等, 2011, 2015, 2017; Xiao et al., 2018), 而且这些生物丘类似于许多学者描述和定义的“ 微生物礁” (Rowland and Shapiro, 2002; Kiessling, 2009; Pratt et al., 2012; Chen et al., 2014; Lee et al., 2014, 2015; Adachi et al., 2015; Lee and Riding, 2018), 在寒武纪缓坡型碳酸盐岩台地中形成了特别的沉积体, 代表着较为壮观的沉积学现象。
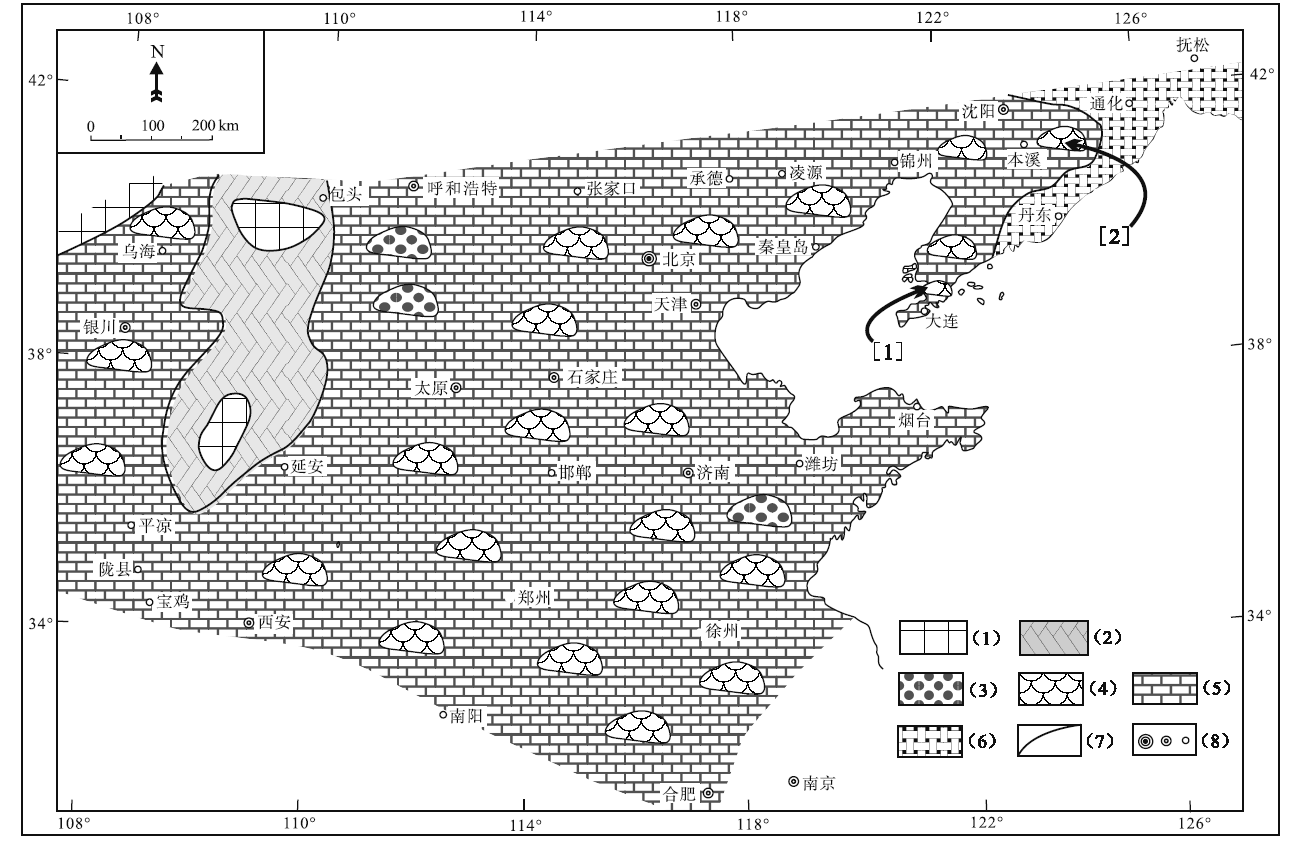 | 图 2 华北地台寒武纪芙蓉世长山组沉积期的沉积格局 图中的[1]代表大连金州湾剖面的位置, [2]所指为本溪田师傅剖面的位置, 注意在浅水台地中还零星分布着小范围的鲕粒滩, 修改自冯增昭等(1990, 2004)。图例代号的含义分别是: (1)古陆, (2)环潮坪相白云岩地层, (3)鲕粒滩, (4)生物丘, (5)中至缓坡相灰岩地层, (6)中至深缓坡相灰岩与泥灰岩地层, (7)相变线, (8)地名Fig.2 Depositional setting of the sedimentary period of the Changshan Formation of the Cambrian Furongian in North China Platform |
辽东半岛的芙蓉统包括长山组和凤山组, 可以识别出3个三级层序(图 3中的DS7至DS9所示; Mei et al., 2005; 梅冥相, 2011b; 梅冥相等, 2011, 2015, 2017), 这些三级层序的底界面均为淹没不整合型层序界面(Gó mez and Ferná ndez-Ló pez, 1994; 梅冥相, 1996, 2010, 2011b; Schlager, 1999), 发育在这些界面上的淹没不整合型层序由“ 凝缩段(CS)+高位体系域(HST)+强迫型海退体系域(FRST)” 序列所构成, 不符合经典和标准的层序地层模式(Vail et al., 1977; van Wagoner et al., 1990), 而更加符合Schlager和Warrlichw(2009)所倡导的碳酸盐岩层序的下降阶段体系域模型。
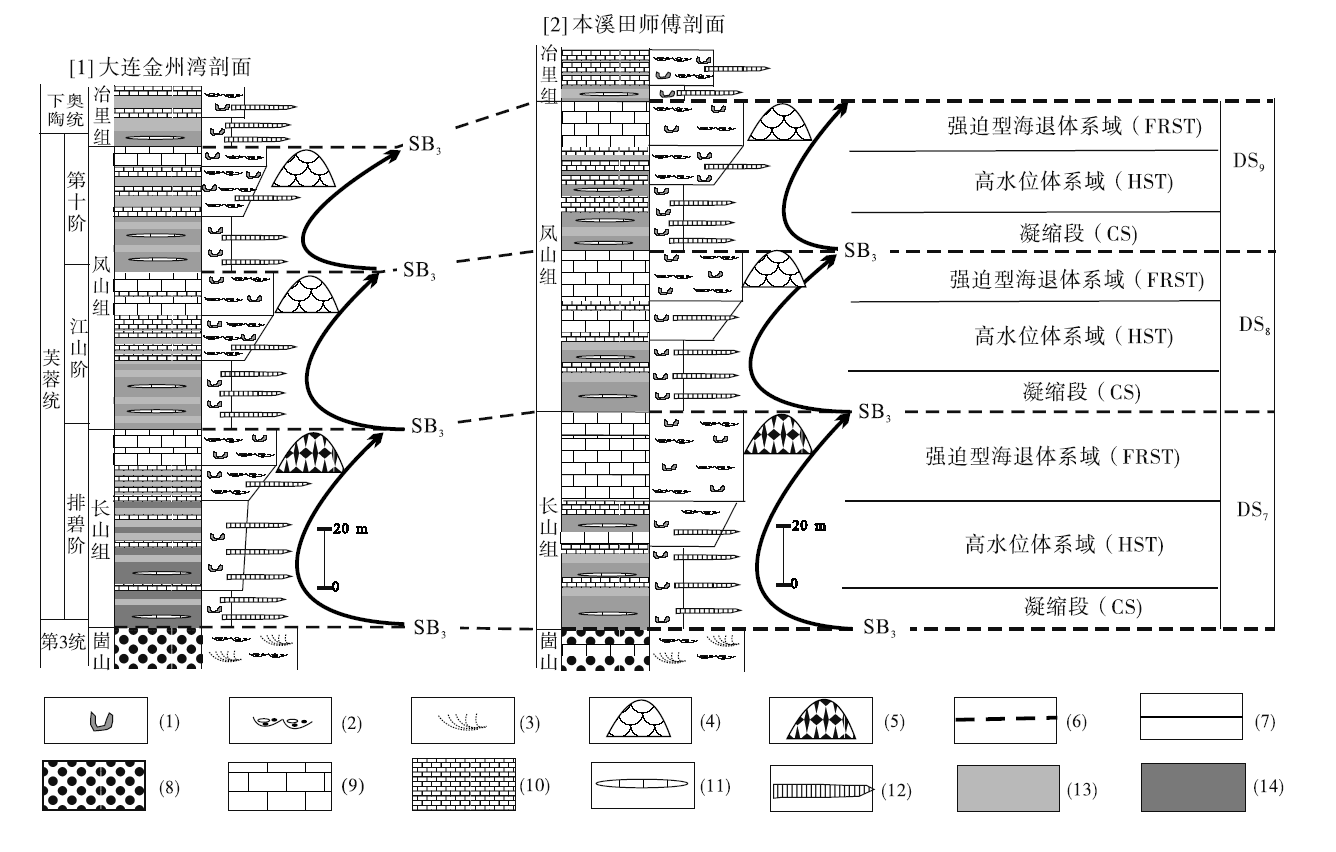 | 图 3 辽东半岛芙蓉统剖面对比以及长山组凝块石生物丘发育的层序地层位置 图中2条剖面的分布位置见图 2; DS1至DS3代表在芙蓉统中识别出的3个三级沉积层序, SB3代表淹没不整合型层序界面, CS代表凝缩段, HST代表高水位体系域, FRST代表强迫型海退体系域; 图例中的符号的含义分别是: (1)生物潜穴, (2)冲刷面, (3)交错层理, (4)叠层石生物丘, (5)凝块石生物丘, (6)层序界面, (7)体系域分界面, (8)鲕粒灰岩, (9)浅缓坡相灰岩, (10)中至深缓坡相泥质条带泥晶灰岩, (11)泥晶灰岩透镜体, (12)竹叶状砾屑灰岩构成的风暴沉积, (13)深缓坡相泥灰岩, (14)陆棚相钙质泥岩Fig.3 Correlation of stratigraphic sections and the sequence-stratigraphic position of thrombolite bioherms in the top part of Furongian Changshan Formation in the Liaodong Peninsula |
最为重要的是, 在大连金州湾和本溪田师傅剖面长山组构成的三级层序的强迫型海退体系域(Hunt and Tucker, 1992; Hell-Hansen and Gjelberg, 1994; 梅冥相, 2010; Catuneanu et al., 2011)中, 集中发育着凝块石(Riding, 2000, 2011; 梅冥相, 2007a, 2007b; Dupraz et al., 2011)组成的生物丘。这些生物丘特别的层序地层位置, 代表了较为典型的相对海平面下降阶段的沉积记录(Schlager and Warrlichw, 2009; Desjardins et al., 2012; Samanta et al., 2016), 不仅反映了在相对慢速的三级相对海平面下降阶段较高的碳酸盐生产作用以及较慢的侵蚀作用的基本特征(Schlager and Warrlichw, 2009), 而且为深入理解寒武纪微生物碳酸盐岩造礁和成丘作用提供了宝贵的实际材料。
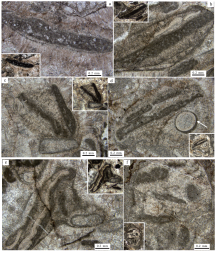
辽宁本溪田师傅剖面位于本溪市东部40ikm的田师傅镇附近的采石场中(图 2)。在该剖面(图3)中, 一套厚度为30余米的块状凝块石生物丘灰岩地层(图 4-a, 4-b)组成了长山组三级层序的强迫型海退体系域(图 3中DS7的FRST; Hunt and Tucker, 1992; Hell-Hansen and Gjelberg, 1994; 梅冥相, 2010; Catuneanu et al., 2011), 与上覆凤山组底部的陆棚相钙质泥岩地层之间形成1个较为典型的淹没不整合型层序界面; 同时在凝块石生物丘的顶部发育1层厚度为30 cm左右的鲕粒滩相灰岩(图 4-a, 4-c), 表明随着三级海平面下降, 沉积环境变浅到潮间至潮下的高能鲕粒滩环境, 代表着特别的海平面下降阶段的沉积记录(Schlager and Warrlichw, 2009; Desjardins et al., 2012; Samanta et al., 2016), 也进一步表明凝块石生物丘发育在中高能的潮下环境之中。这些凝块石生物丘由形态极端不规则的斑块和致密的泥晶构成(图 4-d), 类似于前人的“ 中凝块” 概念, 其特别的块状构造表现出“ 微生物礁” (Rowland and Shapiro, 2002; Kiessling, 2009; Pratt et al., 2012; Chen et al., 2014; Lee et al., 2014, 2015; Adachi et al., 2015; Lee and Riding, 2018))的特点。
 | 图 4 本溪田师傅剖面长山组顶部凝块石生物丘及其鲕粒滩相灰岩覆盖层的宏观特征 a— 长山组浅缓坡相厚层块状灰岩地层突然被凤山组陆棚相钙质泥页岩地层覆盖所形成的淹没不整合型层序界面(箭头所指), 长山组顶部的块状灰岩就是凝块石生物丘灰岩, 在凝块石生物丘灰岩顶部发育1层厚度为20~30 cm的鲕粒滩相灰岩帽; b— 长山组顶部的块状凝块石生物丘灰岩远照; c— 长山组顶部的鲕粒滩相灰岩帽近照, 发育冲刷面(箭头所指); d— 长山组凝块石生物丘灰岩近照Fig.4 Macroscopic feature of thrombolite bioherm and its cap of the ooid-grain bank in the top part of the Changshan Formation at the Tianshifu section in Benxi |
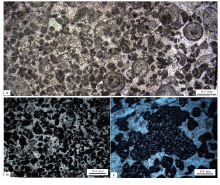
本溪田师傅剖面的块状凝块石生物丘灰岩在显微镜下表现为由大大小小的凝块构成(图 5)。这些暗色泥晶组成的形状极端不规则的凝块, 大小在0.1~0.2 mm之间, 多为次棱角状和棱角状(图 5-a, 5-b), 基本都属于微凝块(microclot; Tucker and Wright, 1990; 梅冥相等, 1997)的范畴。也含有较多的中凝块, 在凝块之间填充着微亮晶胶结物, 其究竟是原生沉淀的还是次生填充的还难以准确判断; 但是, 由于缺乏明显的世代, 这些微亮晶胶结物可能是形成于主导者凝块石的蓝细菌微生物席的昼夜循环中, 是硫化物氧化反应和需要呼吸作用产生的碳酸盐溶解作用所致, 从而区别于后生的化学胶结物。与这些凝块共生的还有泥晶放射鲕粒(图 5-a), 大小在0.2~0.42 mm之间, 不太明显的圈层以及在外部圈层明显的暗色泥晶套表明了这种鲕粒可能的微生物膜成因(Gerdes et al., 1994; Woods, 2013; Siahi et al., 2017), 也可能是属于形成在微生物席内部的一种底栖鲕粒(Gerdes et al., 1994; 梅冥相等, 2011, 2015; 梅冥相, 2012b; 倪胜利, 2017)。另外, 也见到大小超过0.5 mm的葡萄石, 其由暗色泥晶薄膜包覆的几个微凝块和泥晶等构成。除了微凝块之外, 还见到较大的凝块(图 5-c), 其为毫米级别, 大小超过2 mm, 也是形态极端不规则的颗粒, 属于中凝块(Aitken, 1967; Kennard and James, 1986; Burne and Moore, 1987; Shapiro, 2000)的范畴。最为重要的是, 在这些较大的凝块之中, 还残留着类似于基座菌(Hedstroemia)以及孢网菌(Bacinella)之类的钙化蓝细菌化石残余物。
 | 图 5 本溪田师傅剖面长山组顶部凝块石的微观特征 a, b— 凝块石普遍由微凝块构成, 还见到底栖鲕粒(曲线箭头所指)和葡萄状凝块(直线箭头所指); c— 可能为附枝菌(Epiphyton)或基座菌(Hedstroemia)之类的钙化蓝细菌化石构成的凝块(照片中部)Fig.5 Microscopic feature of thrombolite in the top part of Changshan Formation at Tianshifu section in Benxi |
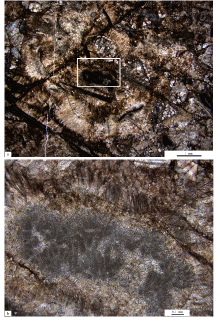
幸运的是, 在一些较大的暗色泥晶组成的凝块中, 发现了较为典型的钙化蓝细菌(图 6), 从而成为了解凝块形成的直接的微生物证据。主要包括以下类型:
 | 图 6 本溪田师傅剖面长山组顶部凝块石的凝块中的钙化蓝细菌 a— 凝块中残留的基座菌(R), 窗格状组构中的亮晶胶结物中的很薄的泥晶薄膜(微米级别)代表着这些微亮晶构成了一种特殊的孢网菌状的组构(B); b— 管状微生物的低倍照片; c— 照片(b)中的矩形框所示部分的管状微生物的高倍照片Fig.6 Calcified cyanobacteria fossil within clots of the thrombolite in the top part of the Changshan Formation at Tianshifu section in Benxi |
1)基座菌(Hedstroemia; 图 6-a)。直径可以达到15~20 μ m的暗色泥晶构成的管形成扇状形态的束状体, 放射状直立生长或排列, 相邻的管可能共享着共同的壁, 或者是共享着共同的隔膜。这是一种可以类比于现代胶须菌(Rivularia; Whitton and Mateo, 2012)的钙化蓝细菌化石(Riding, 1991; Liu et al., 2016; Yan et al., 2017)。
2)孢网菌(Bacinella; 图 6-a)。其在最初命名的时候被作为一种藻。作为一种存在疑问的钙化蓝细菌, 孢网菌多出现在中生代, 以具有明显厚度与延伸的暗色泥晶壁的网状物结壳为特征。Schlagintweit和Bover-Arnal(2013)基于详细的类比和观察认为, 胞网菌(Bacinella)状的组构代表着一种不规则的多孔状结壳而表现出与现代微生物席的一些相似性, 并进一步被解释为丝状蓝细菌及其气孔状的结壳, 尽管卷入它们形成的微组构特征和微生物过程还需要进一步的调查和研究。这样的组构常与基座菌共生, 从而成为一种特别的窗格状组构。
3)管状钙化蓝细菌(图 6-b, 6-c)。这是一种类似于葛万菌(Girvanella)的特殊钙化蓝细菌鞘化石, 以较大直径(20~30 μ m)、较厚的泥晶壁(2~5 μ m)、毫米级别(可以达到2 mm)的长度以及不分叉而近似平行排列的管状体(图 6-b, 6-c)而区别于葛万菌之类的钙化蓝细菌化石。其又类似于山东寒武系第二统的凝块石中发现的管状形式的微生物(Tubiform microbe; Lee et al., 2014), 然而不分叉的特征使其与后者又存在着差异。由于形态学特征和大小(长度与宽度)类似于现代的第谷菌(Tychonema; De los Rí os et al., 2015), 因此, 这种管状钙化蓝细菌是否可以归到葛万菌(Girvanella)组合还有待于进一步研究。
大连金州湾剖面位于大连市金州区西北约30 km的滨海公路旁(图 1)。在该剖面长山组顶部厚度近20 m的块状灰岩与上覆凤山组底部的陆棚相泥页岩之间, 为1个跳相并代表着下伏地层向上沉积环境变浅、上覆地层的沉积环境突然快速加深的淹没不整合型层序界面(图3, 图 7-a; Gó mez and Fernà ndez-Ló pez, 1994; 梅冥相, 1996, 2010; Schlager, 1999; Catuneanu et al., 2011)。长山组顶部的块状灰岩由凝块石生物丘灰岩所组成(图 7-b), 形成较为典型的相对海平面下降阶段的沉积记录(Schlager and Warrlichw, 2009; Desjardins et al., 2012; Samanta et al., 2016), 组成了长山组三级层序的强迫型海退体系域(图 3中DS1的FRST; Hunt and Tucker, 1992; Hell-Hansen and Gjelberg, 1994; 梅冥相, 2010; Catuneanu et al., 2011)。在凝块石生物丘灰岩中, 肉眼可见的形态极端不规则的斑块类似于中凝块(Aitken, 1967; Kennard and James, 1986; Burne and Moore, 1987; Riding, 2000, 2011; Shapiro, 2000; Dupraz et al., 2011), 而且特别的块状构造表现出类似于“ 微生物礁” (Rowland and Shapiro, 2002; Kiessling, 2009; Pratt et al., 2012; Chen et al., 2014; Lee et al., 2014, 2015; Adachi et al., 2015; Lee and Riding, 2018)的基本特点。
 | 图 7 大连金州湾剖面长山组顶部凝块石生物丘的宏观特征 a— 长山组浅缓坡相厚层块状灰岩地层突然被凤山组陆棚相钙质泥页岩地层覆盖所形成的淹没不整合型层序界面(箭头所指), 这些块状灰岩为凝块石生物丘灰岩; b— 长山组顶部的块状凝块石生物丘灰岩近照Fig.7 Macroscopic feature for the thrombolite bioherm in the top part of Changshan Formation at Jinzhouwan section in Dalian |
大连金州湾剖面的块状凝块石生物丘灰岩(图 7), 在显微镜下表现为更加复杂的大大小小的暗色泥晶构成的凝块(图 8)。这些形状极端不规则的凝块, 大小在0.1~1.0 mm之间, 多为次棱角状和棱角状, 属于微凝块(microclot; Tucker and Wright, 1990; 梅冥相等, 1997)到中凝块(Aitken, 1967; Kennard and James, 1986; Burne and Moore, 1987; Riding, 2000, 2011; Shapiro, 2000; Dupraz et al., 2011)的范畴(图 8-a, 8-c, 8-d)。这些凝块常与三叶虫等生物碎屑共生并发育在致密泥晶基质之中(图 8-c), 而且多个凝块或微凝块通过泥晶和微亮晶的粘结作用形成更大而且形态更加不规则的聚合型凝块(图 8-a)。
 | 图 8 大连金州湾剖面长山组顶部凝块石的微观特征 a— 泥晶与微亮晶基质中的凝块和复合型凝块; b— 可疑的饼状菌(Tarthinia), 多被特别的白云石胶结物填充; c— 发育在致密泥晶基质中的暗色泥晶构成的凝块以及三叶虫生物碎屑; d— 围绕着凝块的放射纤维状方解石结壳(实线箭头所指)以及可能的胞网菌(Bacinella)状的组构, 还见到少量雾心亮边的白云石晶体Fig.8 Microscopic features of thrombolite in the top part of Changshan Formation at Jinzhouwan section in Dalian |
较为特别的是, 在致密泥晶基质中还可见到饼状菌(Tarthinia; 图 8-b), 这是一种在大小和形态上类似于肾形菌的可能的钙化蓝细菌化石(Lee et al., 2014), 但是其以较厚的微亮晶壁而区别于肾形菌(Riding, 1991); 另一方面, 还曾经认为Tarthinia(饼状菌)是原生构成为泥晶的钙化微生物菌落遭受成岩改造后的产物(Ezaki et al., 2003), 其确切的生物亲和性还难以确定; 再者, 这些可疑的饼状菌发育着雾心亮边的白云石晶体构成的白云石胶结物(Choquette and Hiatt, 2008), 而且是富有机质微成岩环境中的白云石沉淀物(Roberts et al., 2013), 而且在局部也见到胞网菌(Bacinella)状的组构(Schlagintweit and Bover-Arnal, 2013; 图 8-d)发育在凝块之间。
更加重要的是, 围绕着那些大小不一、形态极端不规则的暗色泥晶构成的凝块, 普遍发育着放射纤维状方解石胶结物(Richter et al., 2011)的沉淀作用形成的结壳(图 8-d)。这种特别的海相放射纤维状和光学上呈束状的方解石结壳, 尽管确切的形成机理还不那么清楚, 但是, 就像Castanier等(1999)在培养试验中观察到的富营养条件下的放射状鲕粒的前体那样, 其也可能代表着特别的富有机质环境中的生物成因的产物(Richter et al., 2011), 类似于Kirkham和Tucker(2018)在凝块石中所描述和解释的受微生物影响的沉淀作用产物。
金州湾剖面凝块石中的凝块, 在高倍显微镜下表现出较为明显的丝状蓝细菌鞘化石构成(图 9), 并成为了解凝块形成的直接微生物证据。这些丝状蓝细菌鞘化石多由较长的(可以延伸为毫米级别)丝状体组成(图 9-a), 由较均匀的泥晶壁(小于2 μ m)所包围的较细的管(小于10 μ m)所构成, 由于多呈束状密集分布, 应该归为束线菌(Subtifioria)而区别于缠绕状的葛万菌(Girvanella), 还可以进一步类比于今天的微鞘菌(Microcoleus)(Riding, 1991; Liu et al., 2016)或鞘线菌(Coleofasciculus; Frantz et al., 2015)。与这些丝状蓝细菌化石组成的凝块共生的还有:附枝菌构成的凝块、放射鲕粒组成的底栖鲕粒和三叶虫化石(图 9-b)。
 | 图 9 大连金州湾剖面长山组顶部凝块石内部的凝块中的丝状蓝细菌鞘化石 a— 较大的拉长状凝块, 明显的丝状蓝细菌鞘化石构成以及围绕着凝块的放射纤维状方解石结壳; b— 照片a的低倍显微镜照片, 表示了凝块的暗色致密泥晶构成; c— 对应于照片d的低倍显微镜照片, 表示了凝块的暗色致密泥晶构成; d— 分布在致密泥晶和微亮晶中的丝状蓝细菌鞘化石组成的凝块, 还见: (1)附枝菌; (2)放射鲕粒; (3)三叶虫化石Fig.9 Sheath fossils within clots of thrombolite in the top part of Changshan Formation at Jinzhouwan section in Dalian |
就像图 9-b所示的那样, 尽管较为罕见, 在金州湾剖面长山组凝块石内的凝块中, 还见到附枝菌构成的凝块(图 10)。与丝状体构成的凝块一样, 在低倍显微镜下这样的凝块总是表现为由致密的暗色泥晶构成(图 10-a), 而且被特殊的放射纤维状方解石结壳所包覆(图 10-b)。这样的附枝菌以泥晶质的而且狭窄的树形状丝状体形成明显的房室状体(图10-b), 具有直立或悬挂式的生长形式, 但由于缺乏现代类比物还被归为红藻(Luchinina, 2009)、可疑的钙化蓝细菌(Riding, 1991; Liu et al., 2016)、非蓝细菌(Adachi et al., 2014)而产生了不同的认识, 不过主导性的认识还是将其归为可以类比于现代大型丝状蓝细菌形成的菌落(Riding, 1991, 2000, 2011; Woo et al., 2008; Woo and Chough, 2010; S $\check{a}$s $\check{a}$ran et al., 2014; Adachi et al., 2015; 梅冥相等, 2017; 齐永安等, 2017a, 2017b), 就像Laval等(2000)的认识那样, 即 “ 一个可能的附枝菌(Epiphyton)与葛万菌(Girvanella)的现代类比物, 表现为一种树形状的微结构而且为灌木丛状结构所组成, 这样的结构实际上为向上展开的大型的钙化丝状体(颤菌(Oscillatoria sp.)和眉菌(Calothrix sp.))聚合体” 。
 | 图 10 大连金州湾剖面长山组顶部凝块石内部的凝块中的附枝菌 a— 低倍显微镜照片, 致密暗色泥晶构成的形态不规则而且大小不一的凝块及其周围的放射纤维状方解石结壳, 矩形框内所示部分进一步放大为照片b, 注意其中局部发育的雾心亮边的白云石晶体; b— 附枝菌组成的凝块及其周围的放射纤维状方解石结壳, 也见到少量雾心亮边的白云石晶体Fig.10 Epiphyton within clots of thrombolite in the top part of Changshan Formation at Jinzhouwan section in Dalian |
就像图 8-a所示的那样, 在金州湾剖面长山组凝块石内的凝块中, 常常见到丝状蓝细菌、三叶虫化石、泥晶与微亮晶共同构成的聚合型凝块(图 11), 可能正是这些聚合型凝块才产生了图 7-b所示的凝块石的宏观特征。以下几个特征将这些聚合型凝块特征化: (1)大型的丝状蓝细菌组成的凝块, 发育着典型的放射纤维状方解石结壳饰边, 组成一种特别的大型聚合型凝块(图 10, 图11-a); (2)更多的情况是, 几个丝状蓝细菌鞘化石组成的小型凝块, 通过微亮晶方解石胶结物和泥晶粘聚成形态极端不规则的毫米级别的聚合型凝块(图 11-b至11-e)。如果将单个形态不规则的丝状蓝细菌鞘化石构成的凝块理解为典型的光合作用生物膜复杂的钙化作用形成的凝块的话, 这些凝块通过方解石微亮晶胶结物和泥晶而粘聚成较大的聚合型凝块, 确实反映了极为复杂的微生物沉淀作用和微生物胶结作用。
 | 图 11 大连金州湾剖面长山组顶部凝块石内部的聚合型凝块 a— 具有放射纤维状方解石结壳饰边的较大的拉长状凝块, 凝块中发育丝状蓝细菌鞘化石; b— 几个较大的而且为丝状蓝细菌鞘化石组成的拉长状凝块, 与三叶虫化石碎片、放射纤维状方解石结壳等组成毫米级别的不规则聚合型凝块; c— 共享着放射纤维状方解石结壳的复合型凝块, 包含着2个丝状体构成的凝块和1个泥晶凝块; d— 被放射纤维状方解石结壳和泥晶粘聚而成的聚合型凝块, 其中的2个拉长状凝块为丝状蓝细菌鞘化石构成, 可见到三叶虫骨刺被丝状蓝细菌鞘化石构成的泥晶膜所包覆; e— 三叶虫化石碎片、丝状蓝细菌鞘化石构成的泥晶凝块粘聚而成的不规则大型聚合型凝块; f— 主要为微亮晶所粘聚而成的大型不规则聚合型凝块, 其中包含着6个小型的丝状蓝细菌鞘化石组成的凝块。每一张照片中的内插照片为相应的低倍显微镜照片, 表示了在低倍显微镜下凝块的暗色泥晶构成的基本特点Fig.11 Aggregated clot within thrombolite in the top part of Changshan Formation at Jinzhouwan section in Dalian |
此外, 在金州湾剖面的长山组凝块石中可见到孤立的放射鲕粒所代表的底栖鲕粒(图9-d)。实际上, 在金州湾剖面的长山组凝块石中, 局部密集发育着较为典型的底栖鲕粒(图 12; Gerdes et al., 1994; 梅冥相等, 2011, 2015; 梅冥相, 2012b; 倪胜利, 2017), 至少包括3种类型: (1)较为典型的放射鲕粒(图 12-a中的箭头(1)所示); (2)较薄的(1~2 μ m)泥晶套包覆的放射鲕(图 12-a中的箭头(2)所示); (3)暗色泥晶薄膜所粘聚的单晶方解石(可能的棘皮动物碎屑)而形成的多晶鲕粒(图 12-a中的箭头(3)所示)。这些底栖鲕粒, 大小在0.5~2.0 mm之间, 多为圆形, 呈漂浮状产出在致密泥晶基质中。另外, 就像图 8-b所示的可疑的饼状菌一样, 还见到了第4种类型的底栖鲕粒, 即被成岩作用改造成白云石构成的底栖鲕粒(图 12-b, 12-c), 其不仅为雾心亮边的白云石晶体构成的白云石胶结物(Choquette and Hiatt, 2008), 而且是富有机质成岩微环境中的白云石沉淀物(Roberts et al., 2013), 其原生形式可能为富有机质的泥晶鲕粒。由于部分鲕粒发育在凝块石之中, 而且常常被泥晶套包覆, 如图 12-a所示的放射状底栖鲕粒区别于球状粒(spherulite; Chafetz et al., 2018; Kirkham and Tucker, 2018), 因为这些建设性的泥晶套可能代表着生物膜的钙化作用产物(Riding, 2000, 2002)。
 | 图 12 大连金州湾剖面长山组顶部凝块石内部的底栖鲕粒 a— 发育在致密泥晶基质中的3种类型的底栖鲕粒: (1)没有泥晶套的放射鲕粒; (2)具有泥晶套放射鲕粒; (3)通过暗色泥晶或放射状微亮晶方解石粘聚单晶方解石而形成的假鲕粒, 这些单晶方解石可能为棘皮动物碎屑; b— 被改造为白云石构成的圆形鲕粒; c— b图中鲕粒的局部放大, 表现出典型的雾心亮边白云石晶体的构成Fig.12 Benthic ooid within thrombolite in the top part of Changshan Formation at Jinzhouwan section in Dalian |
辽东半岛芙蓉统长山组的凝块石构成了一种特别的生物丘, 集中发育在三级层序的强迫型海退体系域(图 3, 图 4, 图 7; Hunt and Tucker, 1992; Hell-Hansen and Gjelberg, 1994; 梅冥相, 2010; Catuneanu et al., 2011)之中, 组成特别的海平面下降阶段的沉积(Schlager and Warrlichw, 2009; Desjardins et al., 2012; Samanta et al., 2016), 代表着壮观的沉积学现象。
长山组凝块石多由较为典型的凝块和微凝块组成(图 5, 图 8), 符合Shapiro(2000)的定义, 即凝块石为“ 凝块状中凝块组成的微生物岩” 。构成这些凝块石的凝块, 尽管在低倍显微镜下多表现为由暗色泥晶组成的形态极端不规则而且大小不一的离散性泥晶凝块, 但是, 在高倍显微镜下可较为普遍地见到钙化蓝细菌鞘化石, 包括: (1)类似于现代第谷菌(Tychonema; De los Rí os et al., 2015)的管状钙化蓝细菌(图 6-b, 6-c); (2)普遍发育着可以进一步类比于今天的微鞘菌(Microcoleus)(Riding, 1991; Liu et al., 2016)或鞘线菌(Coleofasciculus; Frantz et al., 2015)的束线菌(Subtifioria)化石(图 9, 图 11); (3)基座菌(图 6-a, 图 9-d); (4)附枝菌(图 10); (5)可疑的饼状菌(Tarthinia)(图 8-b); (6)胞网菌(Bacinella)状的组构(图 8-d)。因此, 长山组凝块石中的凝块确实是蓝细菌, 而且是丝状蓝细菌主导的光合作用生物膜(Decho, 2010; Flemming and Wingender, 2010; Flemming et al., 2016; Decho and Gutierrez, 2017)复杂的钙化作用产物, 并是构成生物膜的细胞外聚合物质(EPS; Decho, 2010; Flemming and Wingender, 2010; Tourney and Ngwenya, 2014; Flemming et al., 2016; Decho and Gutierrez, 2017)复杂的钙化作用产物, 这可能进一步说明了这些凝块的形成可以被看作是蓝细菌黏结和捕获泥晶质球粒(micritic peloids)的结果, 而且发生在快速的沉积作用期间(Castro-Contreras et al., 2014)。因此, 主要为丝状蓝细菌主导的光合作用生物膜所建造的凝块, 代表着一个独特的沉积学现象, 为了解凝块复杂的成因机理提供了一个较为典型的实例。
在大连金州湾剖面的长山组凝块石中, 不仅发育多个丝状蓝细菌化石组成的凝块通过微亮晶胶结物和泥晶粘聚成更大且形态更加不规则的大型聚合状凝块(图 11), 而且围绕着丝状蓝细菌化石的凝块常发生放射纤维状方解石的沉淀作用或结壳作用(图 8-d, 图 10, 图 11-a), 后者可能代表着富营养条件下的放射纤维状方解石的微生物沉淀作用(Castanier et al., 1999; Richter et al., 2011)或受微生物影响的沉淀作用(Kirkham and Tucker, 2018)。在局部, 类似的放射纤维状方解石的微生物沉淀作用(Castanier et al., 1999; Richter et al., 2011)形成了特殊的放射鲕构成的底栖鲕粒(图 12-a), 其中一些放射状底栖鲕粒的泥晶套以及将棘皮动物碎屑粘聚成假鲕粒的暗色泥晶薄膜, 可能代表着小型的微生物膜的钙化作用产物(Riding, 2002)。
在凝块石中见到的特别的雾心亮边的白云石胶结物(图 8-b, 8-d; 图 10; 图 12-b, 12-c; Choquette and Hiatt, 2008), 代表着富有机质成岩微环境中的白云石沉淀物(Roberts et al., 2013)。更为重要的是, 这些特别的白云石胶结物, 可能代表了微生物席或微生物膜中的细胞外聚合物质(EPS)基质在钙化作用之前曾经经历过的镁— 硅质矿物(如硅镁石, Burne et al., 2014)、非晶质镁硅酸盐(Pacton et al., 2012)、复杂结壳的坡缕石和海泡石纳米纤维构成的微生物席(Cody and Noel, 2012; Perri et al., 2018)的矿化作用, 并在钙化作用之后以及在后来的成岩作用期间形成特别的白云石(以及燧石)残余物(Kruse and Reitner, 2014), 尽管类似的现象也发现在寒武纪浮游鲕粒中(郭芪恒等, 2018), 而且被不太恰当地解释为回流渗透白云石化作用的产物。
上述特征表明, 辽东半岛长山组的凝块石还可能是较厚的微生物席的建造物(Arp et al., 2012), 主要表现在组构性极强的非均质性上。另外, 除了钙化蓝细菌尤其是丝状蓝细菌主导的凝块以外, 还普遍发育各种各样的底栖鲕粒, 从而表明了微生物席内生物成因的碳酸盐颗粒的多样性(图 12; Gerdes et al., 1994; 梅冥相等, 2011, 2015; 梅冥相, 2012b; 倪胜利, 2017), 也进一步说明了在较厚的微生物席内的多重状微生物膜的钙化作用特征, 而且还印证了在方解石海时期(Sandberg, 1983; Wilkinson et al., 1985; Tucker and Wright, 1990; Hardie, 1996; 梅冥相等, 1997; Stanley et al., 1998; Ries et al., 2008; Bots et al., 2011; Balthasar and Cusack, 2015; Kiessling, 2015; Vulpius and Kiessling, 2018)构成微生物席与微生物膜的细胞外聚合物质的降解作用所激发的碳酸盐沉淀作用以方解石为特征这一基本特点(Ries et al., 2008)。
尽管穿越成岩作用过滤器去解释古代凝块石复杂的形成机理将存在着巨大的挑战(Dupraz et al., 2009), 也尽管形成这些凝块石复杂的生物膜钙化作用细节需要更加深入的研究才能得到更好的了解, 但是, 辽东半岛芙蓉统长山组的凝块石, 尤其是在凝块中内较为丰富的钙化蓝细菌鞘化石所表征的直接的微生物活动证据, 使其成为一个了解光合作用生物膜建造凝块的典型实例, 因为蓝细菌是最为成功的造席生物(Stal, 2012), 作为原始生产者的微生物活动最终造成了细胞外聚合物质(EPS; Decho, 2010; Flemming and Wingender, 2010; Tourney and Ngwenya, 2014; Flemming et al., 2016; Decho and Gutierrez, 2017)的生产作用, 并形成特别的生物膜和微生物席。再者, 与凝块共生的三叶虫化石、底栖鲕粒和皮壳粒, 以及围绕着这些多样化颗粒特别的放射状纤维状方解石结壳, 说明这些凝块石可能属于较厚的微生物席内复杂的钙化作用(沉淀作用和早期石化作用; Dupraz et al., 2009, 2011; Arp et al., 2012; Stal, 2012; Bosak et al., 2013)建造物, 从而造成了凝块石内部组成高度的非均质性和变化性(Shapiro, 2000; Planavsky et al., 2009; Mobberley et al., 2013, 2015; Theisen and Sumner, 2016; Louyakis et al., 2017, 2018; Perri et al., 2018)。确实, 这些凝块石中的特别的凝块构成的形成机理, 还存在许多未得到深入了解的细节问题。
辽东半岛芙蓉统凝块石生物丘, 集中发育在三级层序的顶部而构成较为典型的海平面下降阶段的沉积。在辽东半岛芙蓉统长山组凝块石内部的凝块中, 离散性暗色泥晶是这些凝块的基本构成, 但是这些凝块中普遍的丝状蓝细菌鞘化石所表明的蓝细菌微生物构成, 表明这些凝块为丝状蓝细菌主导的光合作用生物膜复杂的钙化作用产物。再者, 以下现象不但将辽东半岛长山组凝块石特征化, 而且还进一步说明了这些凝块石形成于较厚的微生物席内多重状生物膜的复杂的钙化作用, 并表现出高度的非均质性, 即: (1)包覆着蓝细菌化石的暗色泥晶组成的凝块的放射纤维状方解石结壳作用; (2)局部发育的形成于富营养条件下的底栖鲕粒; (3)凝块石内部形成于富有机质微环境中的较为特征的白云石胶结物。尽管还存在着许多细节问题有待于进一步研究, 但是辽东半岛芙蓉统长山组顶部的凝块石为了解较厚的光合作用微生物席内的多重状微生物膜钙化作用形成凝块石及其凝块状组构的复杂过程提供了一个较为典型的实例, 同时也说明了光合作用蓝细菌确实是较为成功的造席生物。希望我们的研究和发现, 为今后的研究提供一些有益的线索和思考途径。
作者声明没有竞争性利益冲突.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|