
{kind=link}
{kind=link}
{kind=link}
{kind=link}
层孔虫研究进展与展望*
[董俊彦 , 龚一鸣]
, 龚一鸣]
, 龚一鸣]
|
|

第一作者简介 董俊彦,男,1995年生,中国地质大学(武汉)硕士研究生,专业为古生物学与地层学,主要从事生物礁和沉积学研究。E-mail: junyandong@outlook.com。
层孔虫( stromatoporoid)一词由德国地质学家 Goldfuss在 1826年创立,因其表面呈纹层状而得名。层孔虫属于海绵动物,是一类营群体生活的海洋底栖固着生物,形态多样,大小从几厘米至数米不等,通常生活在温暖、清澈、盐度正常和光照条件好的浅海中,常与珊瑚、藻类共生而形成生物礁,是重要的造礁生物之一。层孔虫起源于早奥陶世,繁盛于泥盆纪,进入石炭纪基本消失,但在晚三叠世复又出现,并于晚侏罗世得到进一步发展,最终在早白垩世末彻底灭绝。本文在回顾总结了层孔虫研究历史与研究现状,指出层孔虫的研究历史大体经历了零星研究阶段( 1826— 1950年)、系统研究阶段( 1951— 1971年)和集成创新阶段( 1972年至今),主要介绍了层孔虫的形态与构造、系统分类、起源与演化、生物间的共生关系和层孔虫在古环境重建中的应用,在此基础上提出了目前层孔虫研究中存在的 3个主要问题: 缺少明确、完善且统一的生物分类方案;起源的时代存在争议,演化过程中存在奇怪的“间断”现象,早石炭世至晚三叠世的地层中未见相关化石记录;与各种共生生物之间的关系不甚清楚。层孔虫在古生物学的研究中是不可缺少的一个门类,它的各种形态、习性特征在恢复古地理、重建古环境的过程中具有重要意义。
About the first author Dong Jun-Yan,born in 1995,is a master degree candidate of China University of Geosciences(Wuhan),and majors in palaeontology and sedimentology. E-mail: junyandong@outlook.com.
The stromatoporoid,a term proposed by the German geologist Goldfuss in 1826,was named for its laminar structure. As a type of sponge,the stromatoporoid is a marine organism leading a colonial life. Stromatoporoids have various shapes and the size of them ranges from a few centimetres to several metres. Living in a shallow sea with warm,clear,normal salinity and good light conditions,they are often associated with corals and algae to form reefs and are one of the most important reef-building organisms. The evolution of stromatoporoids is a mystery because they originated in the Early Ordovician,prospered in the Devonian,and disappeared into the Carboniferous,but reappeared in the Late Triassic,and further developed in the Late Jurassic,eventually became completely extinct in the end of the Early Cretaceous. The research history of stromatoporoids has gone through three stages: The sporadic research stage(1826-1950),the systematic research stage(1951-1971),and the integrated innovation stage(since 1972). This paper mainly introduces the morphology and structure,systematic classification,origin and evolution,the symbiotic relationship between organisms and the application of stromatoporoids in palaeoenvironmental reconstruction. Three main problems in the current research of stromatoporoids were proposed: (1)the lack of clear,complete and unified taxonomical schemes;(2)the origin and the“blank”phenomenon of evolution;(3)tangible relationships with symbiotic organisms. Based on those,prospects of stromatoporoids’ research have been described. Stomatoporoids are an indispensable part of palaeontology research,and the various morphological and habit characteristics of them are of great significance in the process of restoring palaeogeography and reconstructing the palaeoenvironment.
层孔虫(stromatoporoid)最初由德国学者Goldfuss于泥盆纪地层中发现并命名(Goldfuss, 1826)。它是底栖固着生活的海洋动物(Kershaw and Brunton, 1999; Selden, 2015), 由层状生长的碳酸钙质骨骼构成, 属于海绵动物门(Stearn et al., 1999; 董得源, 2001)。层孔虫是重要的指相化石, 它主要生活在热带至亚热带的水体清澈、盐度正常、水动力较强、光照较好的常氧浅海环境中, 常与珊瑚和藻类等构成生物礁或生物层。在古生代尤其泥盆纪, 层孔虫是重要的造礁生物(Copper, 2002), 它能指示礁体的类型和环境, 而且其形成的生物礁具有良好的油气勘探前景(夏明军等, 2008; 杨海军等, 2012)。
一般认为, 层孔虫于早奥陶世开始发生和发展, 在泥盆纪晚期数量急剧减少, 进入石炭纪基本消失。中生代的地层中也有层孔虫的化石记录(Leinfelder et al., 2005), 其于晚三叠世开始又复现, 但属种不多, 生物群的面貌也比较单调。早、中侏罗世很少见到层孔虫, 这是层孔虫演化过程中的一个衰退期。从晚侏罗世开始, 层孔虫无论是属种数量还是生物群的面貌都得到进一步的发展, 其分布地区主要是特提斯海域。到早白垩世尚有一些层孔虫, 但自此以后就从地球上永远地消亡了(董得源, 2001)。
层孔虫发现至今已有近200年的时间, 然而与之有关的一些重要的科学问题还未解决, 这也是层孔虫成为“ 冷门” 化石的原因之一。首先是层孔虫的生物分类位置, 从最初的海绵动物(D’ Orbigny, 1850; Twitchell, 1928)到后来的水螅动物(Carter, 1877; Nicholson, 1886; Lecompte, 1951, 1956; Yavorsky, 1955; Galloway, 1957; Mori, 1970), 再到现在将之划分至海绵动物门的一纲(Stearn, 1972, 1975; 董得源, 1988), 这是一个认识不断深化和更新的过程。其次是层孔虫的起源与演化问题, 最早的海绵动物出现于寒武纪, 甚至前寒武纪, 而最早的层孔虫化石记录仅出现在早奥陶世的地层中(Li et al., 2017)。石炭纪至中三叠世是层孔虫演化中的“ 空白期” , 这一阶段的地层中未曾有过层孔虫化石的报道, 这是该生物独特的演化规律还是受制于环境条件?亦或是内外因共同作用的结果?解决该问题在层孔虫的起源与演化的研究中至关重要。另外, 古生代的层孔虫与其他海洋无脊椎动物之间存在广泛的共生现象, 这种共生的实质以及它对生物演化带来的影响同样值得进一步关注和研究。
Stock(2001)曾对1926— 2000年间与层孔虫有关的研究成果进行了统计。笔者基于Stock(2001)的统计, 将层孔虫首次发现作为研究的时间起点, 简要梳理了层孔虫的研究历史, 大致概括出3个阶段, 分别是零星研究阶段、系统研究阶段和集成创新阶段。
零星研究阶段(1826— 1950年):层孔虫的研究工作相对较少。这一阶段以研究层孔虫的生物分类位置为特征。自1826年Goldfuss命名了层孔虫并对其进行描述之后, 只有少部分学者对层孔虫进行过相关工作, 且研究重点主要集中在层孔虫的生物分类位置上(D’ Orbigny, 1850; Salter, 1873)。Nicholson于1886— 1892年间发表的关于英国层孔虫的专著, 是首次有关层孔虫的系统论述, 对后来层孔虫的研究有着深远的影响。此后, 层孔虫与其他生物的亲缘关系、分类位置以及地区性层孔虫的报道与描述相继展开(Nicholson, 1874, 1886; Carter, 1877, 1878; Parks, 1907, 1908, 1909, 1935; Kü hn, 1927, 1939; Riabinin, 1929, 1932; Hickson, 1934)。在这100多年的时间里, 古生物学家们对于层孔虫的认识更多的是一个从无到有的过程, 对于层孔虫的研究主要集中于形态的描述和生物的分类, 突出其生物学特征, 而很少关注它的地学意义。而后, 由于第二次世界大战, 层孔虫的研究出现“ 停滞” , 几乎没有任何研究报道和进展(Stock, 2001)。
系统研究阶段(1951— 1971年):二战过后, 层孔虫的研究进入一个新的发展阶段。这一阶段的特征是层孔虫的微细构造以及生物与环境的关系逐渐被重视, 研究内容开始丰富和拓展, 研究进程加快。层孔虫的研究内容不仅限于其微细构造和分类位置(Lecompte, 1951; Yavorsky, 1955, 1957, 1961, 1963, 1967; Galloway, 1957; St. Jean, 1962; Stearn, 1966), 有关层孔虫的生物亲缘性与古生态恢复的重要性在这一时期逐渐凸显出来, 相关的研究成果数量也逐渐增多(Mori, 1968, 1970)。此外, 中生代层孔虫的研究在这一阶段也有较大进展(Hudson, 1954; Schnorf-Steiner, 1960)。得益于整体科学技术的发展, 该阶段较过去取得了重要突破, 但是一个最基本而又最重要的问题— — 层孔虫的分类位置仍悬而未决, 学界还未形成一致的观点, 导致不同分类体系下的研究成果不便于对接与交流。
集成创新阶段(1972年至今):硬海绵(sclerosponge)的发现(Vacelet, 1964; Hartman and Goreau, 1970)是层孔虫研究进程中一个重要的转折点。此后, 层孔虫研究进入集成创新阶段。Stearn(1972)专门论述了层孔虫与硬海绵之间的关系, 提出层孔虫属于海绵动物, 这使得人们对层孔虫的归属认识发生了从水螅动物(Carter, 1877; Nicholson, 1886)到海绵动物的转变。尽管有学者提出不同的认识, 如Kazmierczak(1976)认为层孔虫属于藻类, 但是人们对于层孔虫属于海绵动物这一认识一直持续到现在。这一阶段的特点是层孔虫的研究不再局限于个别特征, 而是涉及其诸多方面, 主要包括: (1)层孔虫的系统分类(Stock, 1994; Stearn et al., 1999); (2)层孔虫的形态结构, 如Stearn和Anmarie(1987)讨论了古生代层孔虫的微细构造, Stock(1989)专门论述了微细网构造等; (3)层孔虫的起源与演化, 包括层孔虫的发生、发展与消亡(董得源, 1989; Mistaien, 1994; Stock, 1994, 2005; Webby, 2004, 2012; Li et al., 2017)、重大生物灭绝事件对层孔虫演化的影响(Stearn, 1987, 2010; Nestor and Stock, 2001); (4)层孔虫在分析古地理(Kö nigshof and Kershaw, 2006; Da Silva et al., 2011a, 2011b)、古环境及古生态(Kershaw, 1998; 吴义布等, 2013; Macneil and Jones, 2016; Park et al., 2017)中的应用等。
3.1.1 宏观构造
层孔虫的构造有内部和外部之分, 不同尺度下可分为宏观构造和微观构造, 大多构造形态可以用图像分析软件来模拟(Wolniewicz, 2010)。与层孔虫有关的构造术语较多, 各学者对术语的解释不统一, 现简要介绍一些意见较为统一的, 具有分类意义或者环境指示意义的构造。
1)共骨(coenosteum)。是层孔虫的构造骨架, 主要由虫体不断分泌钙质碳酸盐而成, 一层骨架形成后又继续向上生长, 这样周而复始, 最后形成大小不同的各种形状的骨架(董得源, 2001)。骨骼成分主要为高镁方解石。共骨形态多样, 常见的形状包括层状— 板状、低丘状、高丘状、球根状和树枝状(Kershaw and Riding, 1978), 大小可从数毫米级至十余米, 通常层状— 板状和丘状的可达数米宽, 而球根状的一般不会超过30 cm, 树枝状的只有几厘米宽。层孔虫的不同属种形成不同的骨架, 虽然共骨的大小没有分类上的意义, 但是共骨的形态是区别层孔虫科、属、种的重要依据(董得源, 2001), 例如树枝状的Stachyodes Bargatzky和Amphipora Schulz可凭借其外形鉴定到属。此外, 共骨的形状还受到生物自身行为功能和生活环境的影响, 因此, 共骨的形状对于层孔虫的古环境与古生态分析具有重要意义(Kershaw and Riding, 1978; 吴义布和龚一鸣, 2013)。
一个理想的层孔虫对称截面可用3个参数表征, 分别是垂直高度(V)、基底长度(B)和对角线距(D)。对于基底不平直, 两侧边缘呈锯齿状的(如图 1, D=(D1+D2)/2), 增加参数RV(RV1与RV2的平均数)和RH(RH1和RH2的平均数), 分别代表锯齿的垂向高度和水平宽度(Kershaw, 1984), 用来描述锯齿状边缘的形态。Ł uczynski(2005)在原有的基础上增加了一些参数, 使层孔虫外形的定量分类更加完善。
 | 图 1 以垂直高度(V)、基底长度(B)和对角线距(D)为端点的层孔虫形态三角分类图(据Kershaw, 1984)Fig.1 Triangular classification figure of stromatoporoids with vertical height(V), base length(B)and diagonal distance(D)(after Kershaw, 1984) |
以V、B和D这3个参数作为三角图的3个端元, 可模拟出层孔虫各种可能的外形轮廓(图1)。图1能很好地显示出层状— 板状、丘状和球根状这3种主要的形态类型, 实际情况中常见的形态在图 1中的虚线范围内, 该区域可分为L、D、B共3部分, 分别代表层状— 板状、丘状和球根状3种形态类型。其中丘状根据V/B值可大致分为低丘状、高丘状等。这样的分类方式一方面可以使外形分类变得简单高效, 便于统计, 但另一方面, 由于形态多样复杂, 树枝状以及一些具有特殊不规则的外形不能呈现在BVD三角图中。
2)星根(astrorhizae)。是在共骨表面上或是弦切面中见到的呈放射状或星状分布的沟槽, 它们的排列呈规则或不规则状(董得源, 2001)。人们最初认为星根是由层孔虫的生殖根(stolon)或虫芽(zooid)形成的, Kazmierczak(1969)根据现代海绵动物和腔肠动物与其他生物存在的共生特性, 提出星根构造可能是某种植物(如藻类)或动物(如无骨骼的腔肠动物)与层孔虫共生而留下的痕迹。但是这一观点遭到后来学者的反对(Mori, 1970; Stearn, 1975)。Labarbera和Boyajian(1991)以Murray定律为模型, 运用定量分析方法对层孔虫的星根构造进行研究, 提出该构造属于流体传输系统的假说, 并且可能与生物体的滤食作用有关。后来的研究指出, 层孔虫化石中的星根构造特点与Murray定律有很高的吻合度(Labarbera, 1993)。尽管星根作为与生物体滤水系统有关的构造已被广泛接受(Stearn, 1975), 但其功能和生物学属性仍然不能完全确定。另外, 星根是层孔虫的重要构造之一, 它的形状和大小可作为鉴别种的依据。
3)细层(laminae)。为橫向骨素, 有层状和板状2种主要类型。通过细层表现出来的不同特征对层孔虫属、种鉴定很重要。
4)支柱(pillars)。为纵向骨素, 其形状和排列多样。支柱构造在分类中有重要作用, 根据支柱的特征可鉴别层孔虫的各个属。
5)虫室(galleries)。2个细层和支柱之间的空间, 是虫体软组织居住的地方(图 2)。
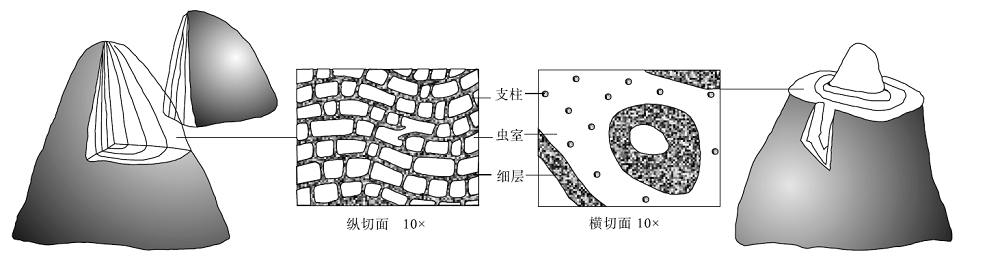 | 图 2 层孔虫的丘状共骨形态及内部主要构造(切面以Simpelxodictyon属为例)(横、纵切面改自董得源, 2001)Fig.2 Mound-like coenosteum and internal structure of the stromatoporoid(the section of the genus Simpelxodictyon is taken as an example)(Horizontal and longitudinal sections were modified after Dong, 2001) |
6)管状珊瑚管(caunopora tubes)。是一种常见的管状构造, 有自己的体壁及横板, 一般认为属于笛管珊瑚(Syringopora)。对于管状珊瑚管的认识, 不同时期的学者有不同的认识, 曾有人认为它是层孔虫自身的一种管状构造, Kirkpatrick(1912)认为这种构造是某种环节动物在共骨上留下的痕迹。但是大多数学者认为管状珊瑚管是珊瑚与层孔虫共生而形成的(Mori, 1970; Mistiaen, 1984; Kershaw, 1987)。
7)厚层(latilaminae)。常由许多细层组成, 界线清楚, 是层孔虫中的层状构造。厚层的形成可能与生长过程中的环境因素有关(Mori, 1968, 1970; Meyer, 1981; Young and Kershaw, 2005)。
3.1.2 微细构造
层孔虫的微细构造在研究早期已为一些学者所注意, 但未曾进行过系统分类。虽然针对层孔虫微细构造的系统研究较少, 但有学者指出微细构造在生物分类和地层划分上具有一定意义(Galloway, 1957; St. Jean, 1962; Mori, 1970)。笔者结合Stearn等(1999)和董得源(2001)对微细构造的描述, 对其中几个比较重要的术语进行介绍和解释。
1)细孔构造(cellular)。是充填在骨骼组织中的小空隙。这些空隙排列紧密, 不规则, 呈亚球形。
2)致密构造(compact)。是用以描述结构元素中致密均匀分布的细点。放射层孔虫目和网格层孔虫目的属种常具有此种微细构造。
3)纤维状构造(fibrous)。由排列成行的细点组成, 行之间相互平行并横穿构造骨素, 但在支柱中, 它可以在喷泉状或羽毛状结构中从轴线向上和向外弯曲。
4)黑球构造(melanospheric)。是指结构元素中小点集中在不规则分布的、球形的、不透明的区域中所形成的一种暗黑色的点。斑点(maculae)可能与此同义。斑点是层孔虫目的主要特征。
5)条纹构造(striated)。是结构元素中细点集中成短棒状的部分, 是Stachyodes属的特有微细构造。
3.1.3 生长形态
1)层孔虫的外饰(ornament)。是在共骨表面形成的, 例如柱状突起(papillae)、乳头状突起(mamelons)等。柱状突起是共骨表面小而圆的隆起, 在鉴定属种方面有一定的意义(董得源, 2001), 但在有些属种上出现是任意的, 没有分类价值(Prosh and Stearn, 1996)。乳头状突起是规则或不规则分布在共骨表面的隆起, 比柱状突起宽大, 高2~10 mm, 其顶端常见星根构造。乳头状突起可能有助于生物体内外的水流交换, 但在生物分类上并不重要, 控制其生长的因素仍不确定。
2)层孔虫内部的生长纹层。包括细层和厚层。边缘是否为锯齿状与层孔虫生活的沉积环境有关, 锯齿状的边缘一般为沉积物所充填, 一定程度上反映了当时的水动力条件及沉积物特征。因此, 对于同一个层孔虫, 随着生长过程中沉积环境的变化, 其边缘形态也会发生变化。厚层可分为包裹和非包裹2种形式。层状— 板状和丘状的层孔虫, 一般光滑的外表面对应着包裹状的厚层, 锯齿状外表面对应非包裹状的厚层, 而树枝状层孔虫的厚层通常是非包裹状的(Kershaw, 1998)。
3)年生长条带。是珊瑚、层孔虫等生物为适应年周期环境变化而在骨骼中出现的沿生长方向周期性重复的条带状构造单元, 不同条带的厚度一般不一样。Mori(1969)首次提出厚层(latilaminae)的概念, 并根据层孔虫内部厚层之间的界线提出了4类厚层: (1)界线为紧密拥挤的厚层; (2)界线清楚的厚层; (3)界线为或宽或窄的层间间隙厚层; (4)界线中含有细粒沉积充填物的厚层。
一般认为, 厚层的形成与层孔虫生长过程中环境因素有关, 这种环境因素包括周期性环境因素(如温度、光照、洋流和季风等)和随机环境因素(如风暴等)。生物在生长过程中, 以“ 厚层” 的方式记录了其生活环境的变迁, 因此“ 厚层” 这一概念基本等同于层孔虫的“ 生长条带” (growth banding)。通常情况下, 以年作为规律性变化的一个周期是最容易识别和划分的, 大部分可观察到的生长条带都是年生长条带。Young和Kershaw(2005)将层孔虫和群体珊瑚的内部条带分为4种类型: (1)条带缺失; (2)由细层形态及细层间距变化而呈现出的密度条带; (3)指示生长停止和再生的生长中断条带; (4)生物遗体经压实和成岩作用而形成的条带。生长条带是通过细层的分布和排列来体现的, 上述(2)、(3)所描述的条带与活体生物的生长环境有关。针对化石珊瑚和层孔虫内部条带是否是季节性变化所诱导形成的, Young 和Kershaw(2005)提出了4个标准: (1)生长条带的厚度及其特点的一致性; (2)骨骼生长的连续性; (3)边缘特征; (4)成岩蚀变的证据。
层孔虫化石的密度条带是通过骨骼内部微细结构排列的“ 疏密” 程度识别出来的。吴义布和龚一鸣(2013)总结了年生长条带的几种类型, 同时提出年生长条带除了密度条带外, 还存在结构条带。密度条带包括5种:(1)疏密型。 生长条带内部微细结构展现出明显的疏密变化, 周期性重复出现, 交替明显, 这是最典型的密度条带结构。 (2)递变型。 条带依然由细层的疏密变化来显现, 但是单个厚层中, 细层往生长方向变厚或变薄, 呈渐变过渡。 (3)混杂型。 单个厚层中, 细层有宽有窄, 但厚层可以被识别出。 (4)突变型。 单个厚层中细层之间的间隔比较均一, 而厚层与厚层之间以较薄或较厚的细层作为界线。 (5)生长脊型。 厚层之间的界线明显, 与厚层内的细层有明显不同, 易识别。
密度条带是温度的季节性变化在生物生长纹层中的体现, 并被保存在化石之中。然而, 周期性的环境因素不单只是温度的周期性变化, 还有其他因素, 如光照、洋流等, 这些非温度的周期性环境因素可能会以其他类型的条带记录在生物化石之中。吴义布等(2013)提出结构条带, 并发现2类这种年生长条带, 分别是“ 结构差异型” 和“ 单一结构层式” 。结构差异型是指相邻两厚层具有完全不同的2种结构, 并以此为基本单元往上周期性重复; “ 单一结构层式” 含有界线清楚的细层, 与“ 生长脊型” 密度条带的差异在于密度条带是存在细层的疏密变化的, 而结构条带没有。与随机环境因素有关的记录是厚层之间被细粒沉积物所充填, 即生物生长过程中出现了生长间断。生长间断在珊瑚和层孔虫中是较常见的现象(Young and Kershaw, 2005), 分为完全生长间断和部分生长间断, 分别是整个层孔虫停止生长和部分停止生长而形成的。层孔虫的间断结构一般与生长纹层平行, 在化石中体现为厚层之间被细粒沉积物所充填或者在共骨的两侧边缘存在“ 沉积物楔” (sediment wedge)。层孔虫内部出现生长间断, 一般是由沉积速率增快而形成的。当沉积速率大于生长速率, 层孔虫顶部全部或部分被沉积物所覆盖而停止生长, 直到一次沉积事件将其顶部的沉积物被水流带走, 层孔虫重新恢复生长。
一般认为, 层孔虫是海绵动物门的一个纲, 但针对其分类位置, 历史上不同学者有不同的看法。早在19世纪, D’ Orbigny(1850)就提出层孔虫属于海绵动物的观点。虽然后来也有学者坚持这一观点(Twitchell, 1928), 但是基于层孔虫与水螅的相似性, 一段时间内大多数学者将层孔虫归属于水螅类(Carter, 1877; Nicholson, 1886; Lecompte, 1951, 1956; Yavorsky, 1955; Galloway, 1957; Mori, 1970)。此外, 也有不同于上述的见解, 如Hickson(1934)与Parks(1935)根据有孔虫Gypsina plana与层孔虫Actinostroma某些特定种的共骨和微细构造相似, 将层孔虫的分类位置置于有孔虫类; Kazmierczak(1976)将其归属于菌藻类。层孔虫属于水螅类这一观点持续了很长一段时间, 直到Vacelet(1964)、Hartman和Goreau(1970)分别在地中海地区和牙买加周边海域发现了硬海绵(sclerosponge)。Stearn(1972, 1975)将层孔虫的软体组织与硬海绵Merlia和Astrosclera相比较, 提出层孔虫是海绵动物门的一个纲, 这一分类位置如今已被大多数学者所认可(董得源, 1988; Stock, 1994; Stearn et al., 1999)。需要注意的是, 确定层孔虫的分类位置是一项基础而又困难的工作, 目前的分类方式只是一种趋同观念, 并没有最终解决层孔虫的分类位置。
Nicholson(1886)首次提出了层孔虫的分类方案, 这是早期最为重要的一种分类方案。根据虫管的有无, Nicholson(1886)将层孔虫分为Hydractinoidea和Milleporoidea 这2个组合, 每个组合又划分为2科, 即Actinostromatidae、Idiostromatidae和Stromatoporidae、Labechiidae。之后的学者在Nicholson的分类基础上提出许多建议并不断完善其分类方案(Heinrich, 1916; Dehorne, 1920; Kü hn, 1927, 1939; Galloway, 1957; Flugel and Flugel-Kahler, 1968), 同时不少学者也提出了新的分类方案, 这些方案的分类标准存在不一致性。如Lecompte(1951)根据骨骼构造要素的排列和微细构造的特征, 将层孔虫划分成5科。Bogoyavlenskaya(1965)在分类中强调星根的作用, 按照支柱、细层和星根构造的排列特征, 将层孔虫作为水螅纲的一个亚纲, 下分5目14科。Khalfina和Yavorsky(1973)主要根据微细构造来分类。Nestor(1974)提出一个系统发生的谱系图, 基本上把层孔虫分为5超科14科。Stearn(1980)基于层孔虫构造要素的相似性, 将古生代的层孔虫作为一个纲归属于海绵动物门, 划分为5目16科87属。Stearn(1980)的分类方案在众多不同的分类方案中是比较合理和全面的, 但也存在一些不足之处, 因此在之后的研究中对该分类方案进行了补充和完善(Stearn et al., 1999)。董得源(2001)将层孔虫作为海绵动物门的一个纲, 分为7目23科101属(表 1)。
| 表 1 古生代层孔虫的不同分类方案比较 Table 1 Comparison of different classification schemes for Palaeozoic stromatoporoid |
3.3.1 层孔虫的起源
Goldfuss(1826)在发现层孔虫的同时, 对其主要构造进行了描述。人们根据他的描述发现, 具有类似构造的生物从寒武纪到现代一直存在(Kershaw, 1998)。关于层孔虫的起源有多种观点(表 2)。最初的观点认为层孔虫最早出现于寒武纪, 但后来的研究发现, 所谓的寒武纪“ 层孔虫” 实际上是古杯类(Archaeocyath), 与古生代地层中的层孔虫只是形态结构上具有某些相似性(Zhuravlev et al., 1994)。这种形态上的趋同可能是古杯动物自适应能力的体现, 因为在早寒武世短时间内的温暖浅海环境中, 古杯动物门Kazachstanicyathida目的一些分子参与了当时礁的建造。Galloway(1957)曾指出层孔虫可能是由古杯动物演化而来, 即由Exocyathus的有关类群演变而来, 但后来的学者指出, 这些小型的古杯动物群并不是后来古生代层孔虫的祖先(Nestor, 1966; Webby, 1986), 没有任何证据表明层孔虫是由古杯动物演化而来的(Webby, 2012)。另外, Khalfina和Yavorsky(1967)认为, 一些奥陶纪的拉贝希层孔虫来源于诸如寒武纪的被称为Stromatocerium属的种。Khalfina和Yavorsky(1974)报道过西伯利亚西南地区2种寒武纪早期的所谓的“ 种” , 但并未对其进行详细描述, 他们将之归于层孔虫Stromatocerium属。但Zhuravlev等(1993)通过对比发现, Khalfina和Yavorsky(1974)报道的2个寒武纪的“ 种” 与寒武纪早期的类珊瑚Flindersipora具有更多的相似性。而后, 他将Khalfina的2个“ 种” 结合成1个新的分类单位, 即Yaworipora khalfinae。关于层孔虫的起源还有多种说法, 但没有一种能有切实的证据说明层孔虫的祖先是何种生物。寒武系第二统至下奥陶统(特马豆克阶)地层中, 至今未发现有类似于层孔虫骨架的生物, 原因可能是当时海水的化学成分(Mg/Ca值)发生了变化, 导致难以孕育出新的后生动物, 而菌藻类乘虚而入, 使得该时期形成的礁体主要由蓝细菌等菌藻生物构成(Rowland and Shapiro, 2002)。
| 表 2 层孔虫起源的代表性观点 Table 2 Representative views of the origin of stromatoporoids |
现在最普遍的观点是, 最古老的层孔虫出现在奥陶纪, 它们主要是拉贝希层孔虫目的一些分子, 主要生活在赤道附近的浅海环境中(Webby, 2004; Nestor and Webby, 2013; Selden, 2015)。层孔虫的早期演化中, Pulchrilaminida属在早— 中奥陶世扮演了十分重要的角色, 可能是层孔虫演化过程中的一个早期分子(Sayutina, 1980)。Pulchrilaminida通常是指一类个体小、单体生活并参与造礁的钙质海绵, 在地理分布上较为局限。Pulchrilaminida是一个存疑的属种。由于它形态较小, 分布局限, 加之与古生代普遍的层孔虫的关联性不强, 学者很少将它划入层孔虫的分类体系中或者在体系中将它作为一个存疑分子(Stearn, 1980; Bogoyavlenskaya, 1984; Khromykh, 1999)。由于至今仍然无法确定Pulchrilaminida是否能归于层孔虫一类, 因此, 也不能断定它是否能作为层孔虫演化的起始。
目前, 古生物学家所确定的最早的层孔虫是拉贝希层孔虫目的属种。Webby(2004)指出拉贝希层孔虫最初突发式地在中奥陶世晚期出现, 可能与当时全球性的进化事件(奥陶纪生物大辐射)有关, 其他的后生动物, 如珊瑚、苔藓虫和藻类等, 也在这一时期迅速分异。Galloway(1957)曾经提出, 最简单、最古老的层孔虫出现在中奥陶世, 即Cystostroma vermontense Galloway。Oh 等(2015)也报导过中朝板块东缘中奥陶世的Cystostroma, 但在安徽省北部宿县和萧县下奥陶统马家沟组下段的地层中, 出现了Labechia, Rosenella, Aulacera, Cystistroma, Cryptophragmus等层孔虫类群分子, 却没有发现Cystostroma属的分子。Li等(2017)报导了黔东南下奥陶统红花园组的石质海绵Calathium礁中的Cystostroma, 时间上位于弗洛期早期, 可能是目前已发现的最古老的层孔虫, 这也是在华南地区首次发现奥陶纪的拉贝希层孔虫。
3.3.2 层孔虫的演化
一般情况下, 古生物的多样性趋势是通过对类群的计数以及统计科、属、种的数量随时间变化来体现的。层孔虫的整体演化过程可从其在地史中的丰度和分异度演化中体现(图 3)。
 | 图 3 显生宙层孔虫的丰度与分异度演化(数据来自董得源, 2001)Fig.3 Evolution of stromatoporoid abundance and diversity in the Phanerozoic(data from Dong, 2001) |
奥陶纪是层孔虫演化的初始阶段, 其数量和分布较为局限, 如中国的奥陶纪层孔虫主要见于鄂尔多斯盆地南缘、塔里木盆地以及华南的多个地区(王振宇等, 2010; 姜红霞等, 2011, 2018; 马璐等, 2014; 包洪平等, 2016; Li et al., 2017)。奥陶纪以后, 只有拉贝希层孔虫目的多样性减少, 取而代之的是网格层孔虫目、层孔虫目和放射层孔虫目等。从中奥陶世到志留纪的罗德洛世, 层孔虫各个目的属种多样性不断增加, 且趋势平稳。在志留纪末, 属种多样性普遍降低, 原因可能是晚志留世和早泥盆世时期海水分布局限(Stearn, 2010)。整个志留纪属种分异度变化不大, 属的数量将近30。但是在整个古生代, 分异度最高的时期是中泥盆世, 达43属, 直到法门期末, 层孔虫的属种多样性依然较高, 有超过30属存在。
晚泥盆世的Kellwasser事件之后, 层孔虫的生物分异度明显下降, 尤其是层孔虫Stromatoporellida、Stromatoporida和Actinostromatida属的数量急剧减少, 几乎灭绝, 但到了法门期, 拉贝希层孔虫目出现复苏现象, 属的多样性增加, 而拉贝希层孔虫以外的层孔虫, 其属的数量从弗拉期的30属减少到11属。在华南地区也存在这样的现象, 华南弗拉期层孔虫有22个属, 但Kellwasser事件后, 在法门早、中期地层中未曾发现过层孔虫化石。晚法门期的地层中(贵州的者王组、广西的额头村组)出现19属层孔虫, 其中5个属(Actinostroma, Anostylostroma, Clathrostroma, Gerronostroma, Stictostroma)是复苏分子, 其余14属是新生分子, 特别是出现了拉贝希层孔虫类(labechiids)和网格层孔虫类(clathrodictyids)分子(董得源, 2001)。层孔虫和床板珊瑚是泥盆纪的主要造礁生物, 这一时期的生物礁表现为弗拉期的珊瑚和层孔虫礁分布有限, 而在Kellwasser事件后数量骤减, 法门期的生物礁主要为钙质菌藻礁(Copper, 2002; 吴义布等, 2010; San Miguel et al., 2017)。
在晚泥盆世, 古生代的层孔虫不论是在丰度还是分异度上都急剧降低, 其属种多样性的降低速率有2个峰值, 一个是在弗拉期和法门期之交, 另一个是在泥盆纪和石炭纪之交。
Stock(2005)比较了世界范围内的7条泥盆系剖面, 统计了在不同地区层孔虫属的变化, 指出从弗拉期到法门期, 属的数量明显减少。在泥盆纪的前4个时期, 即从洛赫考夫期至艾菲尔期, 新生的属多于消失的属, 此后情况与之相反, 无论在数量上还是速率上, 新生属逐渐减少而灭绝属逐渐增加, 并在弗拉期达到最大值。
在早石炭世后至晚三叠世的漫长地质年代里, 至今尚未见有关真正层孔虫的报道。层孔虫在晚三叠世复又出现, 并一直延续到早白垩世, 但在晚侏罗世之前, 其属种数量不多, 生物群面貌单调(董得源, 1989)。中生代层孔虫分布较为局限, 主要集中在特提斯洋南缘以及特提斯洋内的碳酸盐台地(Termier et al., 1985; Wood, 1999)。在晚侏罗世, 层孔虫作为重要的造礁生物, 常与珊瑚构成生物礁。侏罗纪层孔虫礁主要分布地区包括澳大利亚、意大利、北美、西伯利亚、中东等, 这些礁体的位置集中在古纬度25° N区域(Leinfelder et al., 2005; Kano et al., 2007), 此外, 也分布于古纬度较高的日本(Yabe and Sugiyama, 1930, 1935)和巴塔哥尼亚地区(Ramos, 1978)。中生代层孔虫与古生代层孔虫的属种面貌非常不同。以国内报道的层孔虫为例, 中生代层孔虫可分为3超科, 即Actinostromariicae Hudson, 1959, Milleporellicae Hudson, 1959, Burgundiicae Turnsek, 1967, 其中又以Milleporellicae科下的属种数量最多, 分异度最高。与古生代相比, 中生代层孔虫无论在丰度还是分异度上都明显降低, 在最繁盛的晚侏罗世, 也仅有18属。从数据中可以看出, 中生代与古生代层孔虫在演化上存在一定的相似性, 即当它们经过前期长时间的发展演化达到繁盛之后, 在接下来的相对较短的时间内(约30~40 Ma), 数量急剧减少, 最终消失。
要回答层孔虫的起源问题就离不开层孔虫与其他生物的亲缘关系, 同样的, 要探究层孔虫与其他生物的亲缘关系必然涉及到层孔虫的起源问题, 因为2个亲缘关系越近的物种, 越是明确地表明拥有1个共同的祖先。
层孔虫的起源和亲缘关系长期悬而未决主要有2个原因: (1)层孔虫与多种不同的生物都有着相似性(表3), 这样的相似性体现在不同的方面, 如层孔虫共骨的层状生长方式与四射珊瑚具有相似性, 其组织结构的剥落情况与古杯动物(Archaeocyatha)相似(Galloway, 1957); (2)化石记录表明层孔虫于早白垩世末灭绝, 所以仅根据硬体的特征来恢复和重建其软体组织比较困难(董得源, 2001)。目前还未发现能与层孔虫的硬体骨骼作可靠对照的现生生物, 也就是说只能通过一些间接的方法去获悉层孔虫的本质生物学属性。
| 表 3 层孔虫与相关生物之间的主要异同点(据Galloway, 1957; Stearn, 1972, 1975; Wu, 1991; 董得源, 2001) Table 3 Main similarities and differences between stromatoporoids and related organisms(after Galloway, 1957; Stearn, 1972, 1975; Wu, 1991; Dong, 2001) |
在层孔虫属于海绵动物这一认识被普遍认同之前, 大多数层孔虫的研究者认为层孔虫应该属于水螅类。早在19世纪, Carter(1877, 1878)在研究层孔虫与钙质水螅的关系后提出, 层孔虫属于水螅类。后来也有多数学者指出水螅和层孔虫之间的相似性, 尤其是在水螅纲的Hydractiniidae科和层孔虫纲的Actinostromatiidae科之间(Nicholson, 1886; Lecompte, 1951, 1956; Yavorsky, 1955; Galloway, 1957; Mori, 1970)。
古杯动物出现的时间早(寒武纪), 且在某些方面类似于层孔虫, 因此, 它被认为是层孔虫的起源之一。
好的生物分类方案应该能反映亲缘上的关系(phylogenetical relationship), 由于缺少其他的参照, 层孔虫的分类只能依靠钙质骨骼来进行解译(表 4)。
| 表 4 层孔虫纲的目、科分类依据选取的原则(据Stearn et al., 1999; 董得源, 2001) Table 4 Classification principles of stromatoporoid’ s order and family(after Stearn et al., 1999; Dong, 2001) |
从表 4可以看出, “ 目” 级的分类依据主要是层孔虫横向构造中的细层和纵向构造中的支柱。细层和支柱是层孔虫骨架最基本的构造要素。在缺乏软体化石记录的情况下, 细层和支柱的特征是最基本也是最重要的分类依据。在“ 科” 级的分类中, 细层和支柱仍然是主要的分类依据, 只是相较于上一级更加细化其特征。
在属种的分类中, 生物的特征会被描述得更加详细, 除了细层和支柱的基本描述外, 还包括了在不同切面下的特征。此外, 共骨的形状、星根的有无、微细构造的特征等都会作为属种的分类依据。以网格层孔虫属(Clathrodictyon Nicholson et Murie, 1878)的甘溪网格层孔虫(Clathrodictyon ganxiense Wang)为例, 在描述甘溪网格层孔虫的特征时, 涉及到了共骨的形状和大小、表面纹饰、细层厚度、支柱宽度、不同切面上细层和支柱的特征、微细构造、星根的有无以及共生生物等。
由此可归纳出, 一般情况下, 在高级别的分类单位中(目、科), 细层和支柱是主要的分类依据, 而在低级别的分类单位中(属、种), 分类依据更加多样化及微观化。在众多的分类依据中, 细层和支柱的特征和微细构造具有明显的生物学属性。而共骨的形状、层厚与柱宽则更具有环境属性, 原因分别为: (1)共骨的形态组合在不同环境下表现出有规律的交替, 这一点将在下文详细讨论; (2)厚层的形成方式可以与珊瑚、叠层石等层状生长的生物或生物成因的集合体联系起来, 层的厚度受到温度、光照、海水盐度等外界环境条件的影响。此外, 星根构造也常作为一项分类依据出现在属种的描述中。星根构造体现了生物学属性还是环境属性, 取决于其形成的条件, 然而, 星根构造的本质尚未十分明确。目前资料显示, 多数学者认为星根构造的作用是渗滤食物(Stearn, 1975; Labarbera and Boyajian, 1991), 更有实验数据显示层孔虫的星根构造与Murray定律具有很高的吻合度, 很可能属于流体传输系统(Labarbera, 1993)。笔者认为星根构造更具有生物学属性, 作为滤食系统, 这一构造是层孔虫本身具有的。此外, 前人在描述中涉及星根构造时, 多注重星根的有无而忽略不同属种间的特征差别。星根构造作为一种分类依据, 除了星根的有无还应考虑其形貌和结构特征。
在晚泥盆世, 古生代的层孔虫不论是在丰度还是分异度上都急剧降低, 属种多样性的降低速率有2个峰值, 一个是在弗拉期和法门期之交, 一个是在泥盆纪和石炭纪之交。前人曾报道过早石炭世早期(杜内期)的层孔虫, 但后来的学者将前人报道的早石炭世层孔虫归于晚泥盆世法门期, 也有观点认为所谓的早石炭世的层孔虫其实是某类珊瑚, 因为层孔虫中的泡沫板构造也出现在横板珊瑚和四射珊瑚中, 仅凭生物碎片容易将其归为拉贝希层孔虫类(Webby, 2012)。目前, 法门期的层孔虫是否延续到了早石炭世还不能完全确定。根据化石记录, 层孔虫一般被认为在泥盆纪末灭绝。然而, 层孔虫的演化并没有止步于泥盆纪, 在晚三叠世, 层孔虫又复现, 并在晚侏罗世再次达到它的一个繁盛期, 在早白垩世尚有一些层孔虫, 之后完全消亡了。中国的中生代层孔虫礁主要发育于西藏北部的侏罗系, 成礁方式主要为造架作用和障积作用, 中侏罗世层孔虫礁主要见于藏北索县— 巴青地区, 晚侏罗世层孔虫分布相对较广, 尤其是藏北安多— 巴青地区的上侏罗统地层的礁灰岩中(肖传桃等, 2000, 2001, 2014; 赵兵, 2005; 龚文平等, 2006)。北羌塘盆地北部上侏罗统索瓦组中产出有6个种, 分归4属: Parastromatopora sp., P. amdoensis Dong, P. compact Turnsek, P. xizangensis sp. nov, Cladocoropsis sp., C. hybridina Dong, Xizangstromatopora nagquensis Dong, Milleporidium lamellatum Yabe et Sugiyama, 其中Parastromatopora compact Turnsek在南斯拉夫出现于晚牛津期至早钦莫利期, Milleporidium lamellatum Yabe et Sugiyama 在日本上侏罗统的Torinosu灰岩中发现过(赵兵, 2005)。在藏北安多— 巴青地区侏罗系沙木罗组产Cladocoropsis mirabilis层孔虫化石带, 其主要分子同样见于上侏罗统牛津阶— 钦莫利阶(龚文平等, 2006)。川西甘孜— 理塘构造带内发育的侏罗系浅海台地相碳酸盐岩产有层孔虫: Milleporidum sp., M. gracile, Actinostromaria sp., Parastromatopora cf. Simplex, 基本上可以与西藏羌塘地区的沙木罗组进行对比(王全伟等, 2009)。
众所周知, 生物演化具有前进性和不可逆性, 已灭绝的生物不会再次出现。层孔虫的化石记录却出现了消失又重现的奇怪现象。为解释该现象, 笔者试图从内、外因2个角度考虑: (1)这是层孔虫本身具有的不同于其他生物的独特演化规律; (2)层孔虫的消失和重现与环境的演变有关。
Mistiaen(1984, 1994)提出随着环境的改变, 层孔虫仅仅只是丧失了分泌钙质骨骼的能力, 直到中生代遂又开始分泌钙质骨骼。他指出, 是晚泥盆世变化多样的海洋环境引起了法门期层孔虫的骨骼密度降低, 最后导致在泥盆纪之后、中生代之前的地层中找不到层孔虫骨骼的化石。值得一提的是, 在显生宙海水地球化学成分变化的过程中, 奥陶纪至泥盆纪、侏罗纪至白垩纪的海洋属于方解石海, 石炭纪至三叠纪属于文石海(Arp et al., 2001; Horita et al., 2002; 颜佳新和伍明, 2006), 这与层孔虫在地层中的化石记录有明显的一致性。
此外, 笔者通过对比古生代与中生代的层孔虫, 发现两者之间在目一级的分类单位上就已经不存在连续性, 也就是说, 虽然古生代与中生代的层孔虫有着一个共同的名字(stromatoporoid), 但它们之间的关联性并不是直接和明确的。依据生物演化的不可逆性和层孔虫分类位置的扑朔迷离, 古生代与中生代的层孔虫是否是2类不同的生物?这一问题的答案还需要进一步的研究。
层孔虫的生长过程中伴随着许多带壳生物的生长, 在合适的条件下, 带壳生物的壳体会嵌入层孔虫共骨里(Stearn, 2011)。层孔虫中出现共生生物是较常见的现象, 尤其是生物礁内的层孔虫常见笛管珊瑚、枝状的四射珊瑚等。探讨层孔虫在生长过程中与其他生物的共生关系, 对于层孔虫古生态的研究具有重要意义(Kershaw, 2013)。共生生物中最常见的是珊瑚(包括横板珊瑚和四射珊瑚), 另外, 在层孔虫的共骨化石中也存在许多可能是蠕虫类的管状化石(Tourneur et al., 1994; Zhen and West, 1997)。与层孔虫化石共生生物有关的专门研究较少, 一些学者展示了某些类群与共生生物之间的特定关系, 其中最常见的是笛管状珊瑚(Da Silva et al., 2011a)。在大多数情况下, 层孔虫生长不受共生生物的影响(Kershaw, 1987), 但也有例外, Zapalski和Hubert(2011)在法国中泥盆世的地层中发现了层孔虫共骨中的螺旋管构造, 他们把这种构造解释为某种外来生物寄生于层孔虫体内而形成的, 两者之间存在寄生关系, 因为层孔虫的生长纹层在螺旋管周围出现了下凹现象。Vinn(2016)指出, 在奥陶纪、志留纪和泥盆纪, 与层孔虫共生的生物也有所不同。在奥陶纪, 与层孔虫共生的的只有寄生的竹节石纲的Cornulitida目的一些分子, 但在志留纪和泥盆纪, 可见多种无脊椎动物与层孔虫共生, 如横板珊瑚和竹节石等。在泥盆纪的地层中, 与层孔虫化石一同出现的除了珊瑚(常见笛管珊瑚Syringopora)、竹节石、环节动物Streptindytes等无脊椎动物实体化石之外, 还有遗迹化石Torquaysalpinx、 Chaetosalpinx等。一般与层孔虫共生的生物得益于层孔虫, 可保护自己免受天敌的伤害, 且方便觅食, 并拥有相对稳定的底质。这种共生现象多发生于层孔虫Clathrodictyon和Ecclimadictyon这2个属的分子中(Lee et al., 2016; Vinn and Toom, 2016)。Vinn和Toom(2016)对爱沙尼亚地区末志留世鲁丹阶中层孔虫与四射珊瑚共生关系的研究发现, 层孔虫生长形成的钙质骨骼可以作为横板珊瑚生长的稳固基底, 为其提供更方便的觅食, 使其能在浅水高能环境下生长, 同时层孔虫也可能从珊瑚的纵向骨骼中获益, 使自己更加坚固以抵御外界的物理破坏。
泥盆纪是地史上生物礁最为繁盛的时期之一, 尤其在吉维特期和弗拉期极为繁盛, 全球范围内分布广泛, 其造礁生物主要是层孔虫与珊瑚。在晚泥盆世的Kellwasser事件后, 造礁生物的数量和多样性明显减少(Scrutton, 1997; Webb, 2002)。法门期出现的生物礁主要由钙质微生物形成(Aretz and Chevalier, 2007)。泥盆纪末期的Hangerberg事件之后, 无论是后生动物形成的生物礁, 还是微生物形成的生物礁, 都全部消失(Webb, 2002)。
泥盆纪生物礁达到鼎盛的时期, 也是层孔虫最为繁盛的时期。在Kellwasser事件中, 层孔虫遭到重创。然而, 层孔虫在法门期出现复苏现象, 尤其是拉贝希层孔虫目的一些分子(Stock, 2005)。泥盆纪末期的Hangerberg事件后, 层孔虫遭到灭绝(Webb, 2002)。
Stearn(2010)简单总结了关于晚泥盆世灭绝事件的不同成因机制及其对层孔虫演化的影响。他指出各种成因机制都有反对与支持的证据, 气候变化、海水缺氧、海平面升降和二氧化碳含量降低等导致环境的不稳定, 这种不稳定超出了暖水海洋底栖生物的承受能力, 其中就包括层孔虫。至于是否是地外天体撞击而引发的环境剧变仍需要更多切实的证据。从吉维特期到法门期, 海洋环境的恶化和温度缓慢下降影响了层孔虫的出现, 在F-F(弗拉阶— 法门阶)和D-C(泥盆系— 石炭系)界线附近变化尤为明显, 使得生物的宜居栖息地减少, 地区性的缺氧海水涌入浅海。到了泥盆纪末期, 层孔虫无法承受突变恶化的环境, 而后消失(Stearn, 2010)。
从化石记录来看, 层孔虫在漫长的地质年代中经历了消亡与复苏, 最终在早白垩世末灭绝。与古生代末期的消失现象一样, 层孔虫于中生代发生的彻底消亡目前并没有完全合理的解释。一个重要特征就是, 层孔虫的消亡往往伴随着珊瑚— 层孔虫礁的消亡, 因此生物礁减少甚至消失的原因或许可以给层孔虫最终灭绝的原因带来启示。捕食、生物侵蚀、生态空间竞争、光合作用和生物丰度等促使生态系统的动态改变(王建坡等, 2014)。海洋水体的营养水平(Wood, 1993)、海洋化学条件的变化(Stanley and Hardie, 1998)、生物的侵蚀作用与生态空间的竞争(龚一鸣等, 2005; 徐冉等, 2006; Kiessling, 2009; 吴义布等, 2013)等都会影响生物礁的发生、发展与消亡。类似地, 层孔虫灭绝可能是多种因素共同作用的结果。
董得源(1986)根据生态环境适应性的不同, 区分出3类层孔虫: (1)只能在特定环境中生长的层孔虫, 可以指示某一特定的生态环境。如Amphipora指示礁后较封闭的潟湖环境; (2)能在不同环境中生长的层孔虫, 即同种层孔虫在不同环境中有不同的生态反映, 大部分层孔虫属于这一类。一般来说, 块状、球状或共骨较大的层孔虫指示清澈、高能、常氧、盐度正常的浅海环境, 而板状、片状的层孔虫指示清澈、中— 低能、常氧、盐度正常的浅海环境; (3)能在多种环境里生长的层孔虫, 这类层孔虫通常不能指示生态环境, 如Stachyodes可以在礁前生长, 也可以在礁后与Amphipora共同生活在较封闭的潟湖环境。因此, 主要可以从层孔虫的3个方面来推断当时生物生长的古生态与古环境, 即属种分异度与丰度、生长形态以及与其他生物的共生关系。
生长形态是由外界环境因素和内在基因决定的(Galloway, 1957; Mori, 1968, 1970; Cockbain, 1984; Kershaw, 1984, 1990; Kano, 1989)。层孔虫共骨的形状和大小与古生态之间的关系非常密切。基于环境与形态之间的关系, 短期内的沉积事件也可以被保存下来, 尤其是生物在生长过程中沉积环境与水动力条件发生了变化, 生长方向会发生倾斜。层孔虫能够持续生长离不开稳定的基底条件, 因此, 越稳固的形态越能使层孔虫长期生长。例如, 个体较大的层孔虫反映环境因子和生物的生长周期稳定而持续(Wood et al., 1992)。Nestor(1984)总结了控制层孔虫生长的多种因素, 包括底质类型、底质稳定性、沉积特征等。另外, 层孔虫中生长条带的形成普遍认为是以年作为周期单位的(Gao and Copper, 1997), 但是还没有十分确凿的证据来证明这个观点(Young and Kershaw, 2005)。一个主要问题是层孔虫条带与沉积物的沉积速率之间存在一定关系。通过层孔虫的边缘形态及结构或许可以探究两者之间的关系, 因此边缘的存在对于这方面的研究是十分必要的, 以查明是否有沉积事件可能造成的带状结构(Young and Kershaw, 2005)。
众多研究表明, 层孔虫的外部生长形态很大程度上受到生长环境的影响。西沙哈拉摩洛哥南部中泥盆世生物礁的研究表明, 锯齿状和圆丘状的层孔虫相对减少可能指示了在生物礁生长历程中沉积速率的减小(Kö nigshof and Kershaw, 2006)。对比利时南部上泥盆统弗拉阶的层孔虫研究显示, 在该碳酸盐岩台地中, 不同生长形态的层孔虫分布在不同的沉积相带(Da Silva et al., 2011b)(图 4)。
 | 图 4 碳酸盐沉积盆地中层孔虫生长形态与沉积相带的关系(改自Da Silva et al., 2011b)Fig.4 Relationship between growth forms of stromatoporoids and facies in the carbonate sedimentary basin(modified after Da Silva et al., 2011b) |
通过对层孔虫研究历史的梳理发现, 虽然层孔虫的分类与亲缘关系存在多种争议, 但层孔虫属于海绵动物门这一认识目前已基本取得共识, 并且有相对可靠的证据来支持这一观点(硬海绵)。在恢复古生态与古环境方面, 层孔虫亦是良好的指相化石, 它的形态、大小、结构与构造都能体现特定的生长环境。
综观层孔虫的研究历史与现状, 目前尚存在3个方面的问题: (1)层孔虫的系统分类。层孔虫的分类位置及分类方案是一个基础而又复杂的工作, 需要长期的积累, 尽管从发现层孔虫至今已有一百多年的时间, 但随着新属种不断被发现, 其分类方案也在不断地被更新。 (2)层孔虫的起源与演化。层孔虫最早的化石记录出现在早奥陶世的地层中, 但其真实的起源时间目前并未确定。层孔虫的演化中最大的特点是在石炭纪和二叠纪的地层中缺少与之相关的化石记录, 这种现象显然与我们对于生物演化不可逆性的认知相矛盾。因此, 探究该现象出现的原因是层孔虫研究中一个重要的内容。 (3)层孔虫与其他生物的共生关系。从化石保存的结果来看, 层孔虫与多种浅海无脊椎生物的共生现象十分普遍, 尤其是古生代的层孔虫, 常与珊瑚、竹节石及各种藻类共生。层孔虫与其他生物之间的关系是互利、偏利、互抗还是彼此不产生影响, 需要进一步研究才能给出确切的答案。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|