





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
鄂尔多斯盆地苏里格气田中奥陶统马家沟组五段凝块石类型与古盐度初步分析
[乔亚斌1  , 张林
, 张林1 , 王予1 , 张一果1 , 马君霞2 , 姜红霞3 ]
, 张林]
|
|


第一作者简介 乔亚斌,男,1975年生,高级工程师,长庆油田分公司第五采气厂总地质师,从事气田开发技术管理工作。E-mail: qyb_cq@petroChina.com.cn。
微生物碳酸盐岩是近年来地学界研究的一个热点,其时空分布非常广泛,并可形成重要的油气储集层。在鄂尔多斯盆地北部苏里格气田的中奥陶统马家沟组马五 1+ 2段和马五 4段,笔者分别发现了非钙化微生物成因的凝块石微生物岩。依据显微结构,将凝块石划分为蠕虫状凝块石、网状凝块石和房室孔洞状凝块石 3种类型。通过与基里巴斯现代超盐湖微生物碳酸盐形成环境的对比,推测在华北地台云坪广布、坪中有湖的大背景下,凝块石是在相对海平面上升、盐度大约在 60‰~149‰之间的条件下形成的,而含石膏结核碳酸盐岩是在相对海平面下降、盐度达到 149‰以上的条件下形成的。进一步研究发现,蠕虫状凝块石泥粉晶白云岩的白云石晶间孔和溶蚀孔非常发育,网状凝块石和房室孔洞状凝块石的溶蚀残余孔较发育,推测凝块石微生物岩具有较好的物性,是未来油气勘探的重要目标岩性之一。
About the first author Qiao Ya-Bin,born in 1975,senior engineer of gas field development,chief geologist of Gas Production Plant 5 of PetroChina Changqing Oilfield Company. He works on gas field development technical management. E-mail: qyb_cq@petroChina.com.cn.
Studied on well cores of the 5th Member of Ordovician Majiagou Formation in the Ordos Basin,it reveals that the microbialites in the 2nd layer of $Ma^{4}_{5}$,the 2nd layer of $Ma^{2}_{5}$,the 4th layer of $Ma^{1}_{5}$ can be divided into three distinct types: wormlike thrombolite,bacterial netlike thormbolite and chamber-patches thrombolite. By comparing with formation conditions of the modern microbial carbonate in the hypersaline lakes in the Republic of Kiribati,it is inferred that (1)the studied thrombolites formed in hypersaline lakes with salinity of 60‰~149‰ as the relative sea level rose,and(2) the gypsum-bearing carbonates formed in the hypersaline lakes with salinity of above 149‰. The wormlike thrombolites have high contents of intercrystaline pores and dissolution pores,and the netlike and chamber-patches thrombolites have high contents of relict pores, which makes thrombolites as an important exploration target.
微生物碳酸盐岩是指由底栖微生物群落通过捕获和粘结碳酸盐岩碎屑颗粒而成的沉积物, 或由微生物诱发形成的碳酸盐沉淀物, 故是一种由生物媒介作用沉积的产物。微生物碳酸盐岩一般包括叠层石、凝块石、树形石、均一石4种类型(Riding, 2000), 吴亚生等(2018)在总结了前人对微生物岩的各种定义和分类后, 对微生物岩的分类给出了一个新的方案。微生物碳酸盐岩是近年来地学界研究的一个热点, 其时空分布非常广泛并可形成重要的油气储集层, 因而成为油气储层研究的一个新领域。
凝块石(thrombolite)的概念最初是由Aitken(1967)提出来的, 指由非钙化藻类(隐藻)通过粘结或分泌作用形成的沉积物(岩), 外观多具有穹状隆起或者不规则波状起伏的宏观形态, 与围岩的边界明显, 内部无纹层状构造。形成凝块石的微生物主要是原核微生物(Riding, 1991, 2000), 也可能有真核生物参与(Feldmann and Mckenzi, 1998), 或含有钙化的生物或者微生物化石(Riding, 2000)。在吴亚生等(2018)的分类方案中, 凝块石是由非钙化底栖微生物形成的内部具有凝块结构的微生物岩, 一般具有丘状、柱状、面状等构造。现代凝块石通常发育在潮下带(Aitken, 1967; Feldmann and Mckenzi, 1998), 中生代凝块石出现在向海一侧的深水珊瑚— 海绵生物礁环境中(Mancini et al., 2004), 古生代已报道过的不同构造的凝块石既可能出现在浅滩或者较深水的低能潮下带(肖传桃等, 2018), 也可能出现在具较高能量水动力的微生物丘滩复合体中(熊鹰等, 2016), 前寒武纪的凝块石出现于水体动荡的浅潮下带(余家仁等, 1998)。上述所有的研究结果均认为, 凝块石形成的水深应该比叠层石大, 但是水体能量的高低却因为研究区域的环境不同而不一样。
目前, 国内学者已经在塔里木盆地、鄂尔多斯盆地奥陶系中发现微生物岩, 均具有重要的油气储层价值(贡云云, 2016; 熊鹰等, 2016; 王玉伟等, 2019)。鄂尔多斯盆地奥陶系马家沟组是长庆油田天然气主力产层, 许多学者围绕该储集层成因开展了大量研究, 确定其主要是奥陶纪末古风化壳型的。但最近在鄂尔多斯盆地东北部奥陶系马家沟组马五1+2段中识别出的微生物碳酸盐岩(熊鹰等, 2016), 为深化认识马家沟组储集层成因提供了新的思路。
笔者最近在对鄂尔多斯盆地苏里格气田奥陶系马家沟组的研究中, 发现马五1+2段和马五4段中均发育微生物岩, 具有较丰富的储集空间, 进一步昭示了微生物岩对奥陶系马家沟组储层的贡献。文中介绍了笔者对这套储层的微生物岩的结构分类和形成环境方面的研究成果。
鄂尔多斯盆地位于华北台地西缘, 为一矩形构造盆地。马家沟组沉积时期, 由于西部贺兰裂谷强烈的沉降扩张以及肩部翘升导致形成一个“ L” 形的中央古隆起, 并在古隆起东侧形成以米脂— 延安为沉降中心的陕北坳陷(何江等, 2009)。
马家沟期是鄂尔多斯盆地奥陶纪海相沉积层形成的主体时期, 海侵— 海退旋回明显发育, 形成碳酸盐岩与蒸发膏盐岩交替发育的局限海台地相沉积序列。马家沟组可分为6段, 对应的沉积期分别为马一亚期— 马六亚期, 指示3次海退— 海进旋回。
鄂尔多斯中东部在马一亚期的海退期时就开始明显的拗陷沉降, 由于中央古隆起等高地的障壁作用, 使中东部的坳陷区逐渐形成与秦祁广海分隔的局限海洼地沉积环境, 在炎热干旱的古气候环境及海平面周期下降等因素配合下, 形成强烈蒸发的巨厚膏盐岩沉积。后历经马三亚期、马五亚期2次海退旋回, 中东部的沉积范围渐次加大, 到马五末期已基本覆盖至中央古隆起所在的区域(图 1)。马五段总体以含硬石膏结核泥粉晶白云岩为主, 局部有微生物岩发育(图 1)。
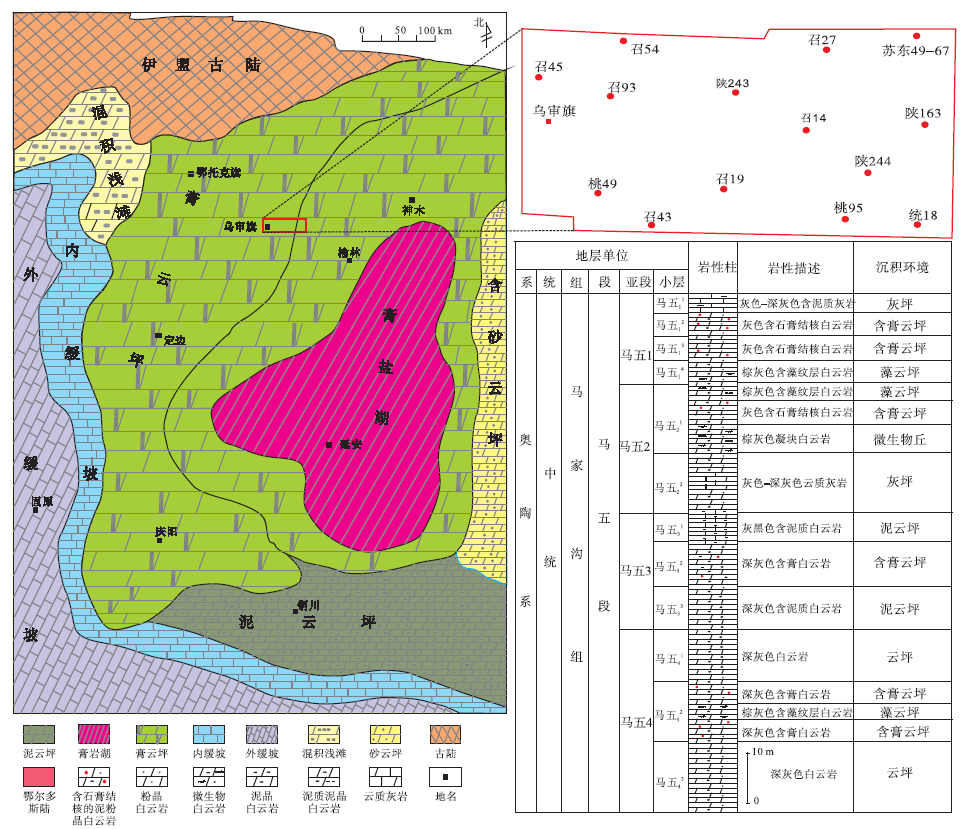 | 图 1 鄂尔多斯盆地中奥陶统马家沟组五段地层柱状图及岩相古地理(据邵东波等, 2019, 有修改)Fig.1 Lithofacies palaeogeography map(modified from Shao et al., 2019) and stratigraphic column of the 5th Member of Middle Ordovician Majiagou Formation in Ordos Basin with distribution of study wells |
鄂尔多斯盆地马家沟组长期被归于中奥陶世达瑞威尔阶晚期, 但冯增昭等(2004)认为其应属于下奥陶统, 目前仍有学者将其归入下奥陶统(沈杨等, 2018; 孙玉景和周立发, 2018; 王国亭等, 2018; 邵东波等, 2019)。王志浩等(2016)通过对牙形刺的研究, 将其修订为中奥陶统(王志浩等, 2016)。Meng等(2017)对马家沟组的地层进行了碳、硫同位素测试, 测试结果也表明马家沟组属于中奥陶统。文中采用王志浩等(2016)的意见, 将马家沟组归入中奥陶统达瑞威尔阶。
苏里格气田是长庆油田所属鄂尔多斯盆地的主要大气田, 位于北起鄂托克后旗敖包加汗、南至陕西定边、东到乌审旗桃利庙、西达鄂托克前旗的区域内, 在构造区域上主要属于伊陕斜坡北部。文中研究的4口取心井苏东49-67、召54、召14、陕243井位于苏里格气田北部(图 1)。
根据岩心和薄片观察的结果, 对4口井取心段的岩石学特征及沉积环境做简要描述。
该井奥陶系马五段取心段限于马五
 | 图 2 鄂尔多斯盆地中奥陶统马家沟组五段取心段岩性和沉积微相柱状图 A— 苏东49-67井; B— 召54井; C— 召14井; D— 陕243井Fig.2 Lithology and sedimentary microfacies of the 5th Member of Middle Ordovician Majiagou Formation in Ordos Basin |
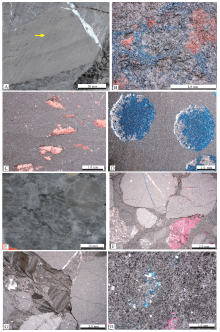
 | 图 3 鄂尔多斯盆地苏东49-67井和召54井中奥陶统马家沟组五段岩心及显微特征 A— 苏东49-67井, 3134.5 m, 石膏结核泥粉晶白云岩, 黄色箭头指示被方解石交代的球状的石膏结核, 岩心照片; B— 苏东49-67井, 3133.37 m, 蠕虫状凝块石泥粉晶白云岩, 蓝色为铸体, 红色为染色的方解石, 单偏光; C— 苏东49-67井, 3132.0 m, 具鸟眼构造的泥晶白云岩, 鸟眼孔洞多数被方解石充填, 方解石被染色成红色, 单偏光; D— 苏东49-67井, 3122.14 m, 石膏结核泥粉晶白云岩, 球状的石膏结核中石膏大部分被溶蚀, 溶蚀孔被蓝色铸体溶蚀充填, 单偏光; E— 召54井, 3284.0 m, 角砾岩, 岩心照片; F— 召54井, 3284.4 m, 角砾岩, 单偏光; G— 召54井, 3282.3 m, 角砾岩, 单偏光; H— 召54井, 3289.0 m, 网状隐藻凝块石泥晶白云岩, 单偏光Fig.3 Microphotographs and core of the 5th Member of Middle Ordovician Majiagou Formation from Wells Sudong 49-67 and Zhao 54 in Ordos Basin |
根据上述分析, 苏东49-67井马五
该井奥陶系马五取心段同样只有马五
因此, 召54井马五
该井奥陶系马五段取心较多, 包括马五
 | 图 4 鄂尔多斯盆地召14井和陕243井中奥陶统马家沟组五段岩心及显微特征 A— 召14井, 3198.6 m, 房室孔洞状凝块石泥粉晶白云岩, 红色区域为方解石染色, 单偏光; B— 召14井, 3196.1 m, 云质灰岩, 红色区域为方解石染色, 单偏光; C— 召14井, 3187.3 m, 含石膏结核的角砾白云岩, 黄色箭头指示较规则的球状石膏结核, 岩心照片; D— 陕243井, 3229.5 m, 云质灰岩, 单偏光; E— 陕243井, 3226.5 m, 含石膏晶体的泥晶白云岩, 石膏晶体溶蚀后多数被方解石充填溶蚀孔, 方解石被染色, 单偏光; F— 陕243井, 3225.3 m, 含隐藻条带碎块的泥粉晶白云岩, 单偏光; G— 陕243井, 3223.0 m, 房室孔洞状凝块石泥粉晶白云岩, 单偏光; H— 陕243井, 3220.0 m, 云质灰岩, 单偏光Fig.4 Microphotographs and core of the 5th Member of Middle Ordovician Majiagou Formation from Wells Zhao 14 and Shan 243 in Ordos Basin |
综上可知, 召14井马五
该井奥陶系马五段取心不连续, 自下而上有马五
根据上述分析, 陕243井马五
主要依据4口井岩心薄片的微观形态特点, 笔者将凝块石进一步分为3种类型: 蠕虫状凝块石, 网状凝块石和房室孔洞状凝块石。
1)蠕虫状凝块石。见于苏东49-67井(图 5-A), 为凝块石泥粉晶白云岩。在镜下, 呈不规则短棒状的暗色区域分割亮色白云石微胶结物区域(图 5-A1)。在局部放大区域的清绘图上(图 5-A2), 暗色区域的短棒状形态表现为不规则断续条带状— 斑块状展布。由于边缘极度不规则, 伸展长度也短, 整体弯弯曲曲, 用条带状描述并不妥当; 同时, 由于凝块部分的大小、宽窄、长短都差别太大, 用斑块状描述也有些不妥。基于这种形态好似蠕虫聚集或者分散的样子, 故命名为蠕虫状凝块石。
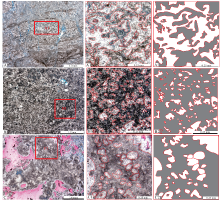
 | 图 5 鄂尔多斯盆地中奥陶统马家沟组五段凝块石类型及凝块形态 A— 苏东49-67井, 3133.7 m, 蠕虫状凝块石, A1、A2为A局部放大照片, 深灰色为凝块, 不规则弯曲状, 形态曲折不一; B— 召54井, 3289.0 m, 网状凝块石, B1、B2为B局部放大照片, 深灰色为凝块, 被大小不一的亮晶区域分割成网状, 亮晶部分就像网眼一样分布; C— 召14井, 3198.1 m, 房室孔洞状凝块石, C1、C2为C局部放大照片, 深灰色为凝块, 被很多像房室一样的内部亮晶区域将凝块分割成斑状。A2、B1、C2为A1、B1、C1的清绘图Fig.5 Types and clot morphology of thrombolites from the 5th Member of Middle Ordovician Majiagou Formation in Ordos Basin |
2)网状凝块石。见于召54井(图 5-B), 为凝块石泥晶白云岩。镜下看, 明暗区域形成网状结构(图 5-B1), 暗色部分比较细碎, 亮色区域好似网眼, 网眼大小不规则, 局部形成大的网洞。在局部放大的清绘图上(图 5-B2), 网眼的大小不同, 网眼之间网络结构也不是很规则, 以多数网眼之间不连通的特征区别于另外两种类型, 因此命名为网状凝块石。
3)房室孔洞状凝块石。见于召14井(图 5-C), 为凝块石泥粉晶白云岩。镜下可见, 凝块结构如同肾形菌的房室叠加, 内部为亮晶充填(图 5-C1), 多数圈层并不连续。放大后的清绘图(图 5-C2)显示, 其与肾形菌还是有差别的:蓝细菌中的肾形菌的形态通常是由中空、膨大的囊形成的群体, 大小变化甚大, 从由几个小囊组成的简单群体至由许多小囊组成的复杂的姜状或葡萄状群体, 从中心向周围呈叠层状生长, 并有分枝(刘丽静等, 2011); 而图 5-C中虽然有房室状的形态, 但是并不具有肾形菌特征的外壁, 并且叠层状囊群体不明显, 在其他薄片中观察到的更多的是非圆形的孔洞形态。所以, 不能将该微生物岩归为肾形菌成因。但是, 此种结构与前两种凝块石也不相同, 具有疑似房室状的结构, 因此命名为房室孔洞状凝块石。
3.2.1 蠕虫状凝块石
主要出现在苏东49-67井马五
 | 图 6 鄂尔多斯盆地苏东49-67井和召54井中奥陶统马家沟组五段微生物岩显微特征 A— 苏东49-67井, 3133.7 m, 蠕虫状凝块石泥粉晶白云岩, 暗色为凝块, 亮色为凝块之间的胶结物, 蓝色铸体充填白云石晶间孔和晶间溶扩孔, 铸体薄片; B— A的局部放大; C— 苏东49-67井, 3133.7 m, 蠕虫状凝块石泥粉晶白云岩, 黄色箭头指示凝块石溶蚀后被渗流粉砂充填; D— 召54井, 3289 m, 网状凝块石泥粉晶白云岩, 暗色为凝块, 形态为断续的网状, 左下方可见似丝体状凝块(红色箭头所指), 亮色为凝块之间的胶结物, 黄色箭头指示不规则状石膏结核; E— 召54井, 3289 m, 网状凝块石泥晶白云岩, 可见隐藻纹层状凝块(红色箭头所指), 交代石膏结核的块状连晶方解石染色成红色; F— 召54井, 3289 m, 网状凝块石泥粉晶白云岩, 黄色箭头指示柱状的石膏晶体, 红色箭头所指右下方的暗色区域为比较规则的网状凝块结构, 薄片多数区域为不规则的断续的网状结构Fig.6 Microscopic characteristics of the microbialite the 5th Member of Middle Ordovician Majiagou Formation from Wells Sudong49-67 and Zhao54 in Ordos Basin |
推测暗色的泥粉晶蠕虫状凝块可能是微生物引起的钙化作用形成的并且遭受了白云石化作用, 粉晶组成的浅色蠕虫状结构是暗色凝块之间的孔洞被文石或者方解石胶结物充填后遭受白云石化形成的。粉晶白云岩和粉细晶白云岩晶间孔发育。另外, 在成岩过程中凝块石发生溶蚀, 溶蚀孔洞被粉砂级的白云石充填(图 6-C), 粉砂级的白云石之间发育晶间溶扩孔。
3.2.2 网状凝块石
网状凝块石出现在召54井马五
3.2.3 房室孔洞状凝块石
房室孔洞状凝块石出现在召14井马五
 | 图 7 鄂尔多斯盆地召14井和陕243井中奥陶统马家沟组五段微生物岩显微特征 A— 召14井, 3198.1 m, 房室孔洞状凝块石泥粉晶白云岩, 中央为大的斑块状凝块石角砾, 角砾外侧为溶蚀成因的粉晶屑白云岩, 可见蓝色的铸体充填胶结物的溶蚀孔隙, 铸体薄片; B— 召14井, 3198.1 m, 房室孔洞状凝块石泥粉晶白云岩, 凝块的结构与照片A稍有不同(黄色箭头所指), 呈圆形或椭圆形的房室形态, 染色后红色的是方解石, 充填溶蚀孔洞, 蓝色的铸体充填溶蚀孔, 铸体薄片; C— 召14井, 3198.1 m, 脑纹斑块状凝块石泥粉晶白云岩, 大面积的溶蚀区域填充自形细粉晶白云岩, 白云岩在成岩后期又遭受溶蚀, 溶蚀孔洞被方解石(红色)充填, 还有部分剩余孔隙被铸体充填, 铸体薄片; D— 陕243井, 3223.0 m, 房室孔洞状凝块石泥粉晶白云岩, 被溶蚀后的凝块石角砾大小不一, 角砾内部有溶蚀孔洞残留, 铸体薄片; E— 陕243井, 3222.85 m, 房室孔洞状凝块石泥粉晶白云岩, 黑色的凝块呈圆形或椭圆形的房室形态(黄色箭头所指)以及斑状形态, 凝块石内部有残留孔隙被铸体充填, 铸体薄片; F— 陕243井, 3222.85 m, 房室孔洞状凝块石泥粉晶白云岩, 残留的溶蚀孔隙多分布在凝块石内部, 被铸体充填, 铸体薄片Fig.7 Microscopic characteristics of microbialites of the 5th Member of Middle Ordovician Majiagou Formation from Wells Zhao 14 and Shan 243 in Ordos Basin |
召14井的房室孔洞状凝块石由于溶蚀作用强烈, 局部的凝块石完全被溶蚀, 自形和半自形的细粉晶白云岩充填了溶蚀区域(图 7-C)。细粉晶白云岩的原始晶间孔较发育(图 7-B), 后期又发生溶蚀, 溶蚀空间部分被方解石充填。细粉晶白云岩的晶间孔和溶蚀孔的发育, 使岩石具有针孔状结构。凝块石内部的亮晶胶结物也发生溶蚀, 形成溶蚀孔洞(图 7-A, 7-D)。丰富的针孔状溶蚀空间表明, 该种凝块石具有良好的油气储集性能。
陕243井采样部分虽然没有观察到针孔状结构, 但是凝块结构以及凝块角砾的溶蚀成因都是一致的, 不同的是后期成岩改造作用影响较小。由于鄂尔多斯盆地东部马家沟组马五1+2的古风化壳非常发育(沈杨等, 2018), 故可以推测在其他位置应有溶蚀发育区域。
根据岩心观察和薄片研究, 4口井的取心段有球状石膏结核白云岩、凝块石白云岩、白云岩以及云质灰岩等。
球状石膏结核白云岩一般为灰色或深灰色含石膏结核的泥粉晶白云岩, 白云岩晶体表面发黄, 中心多含杂质, 为次生加大成因的亮边雾心结构。基于波斯湾现代潮上带环境中有含球状石膏团块的沉积正在形成, 所以具石膏结核的白云岩一般被认为形成于蒸发作用强烈的、高盐度、偏氧化的潮上环境。
对于凝块石, 目前的共识是由微生物群落形成的, 微生物群落主要由蓝细菌组成(Riding, 2000)。由于蓝细菌不可以长时间脱离水体而生活, 因此具有一定厚度的凝块石不可能在潮上带形成。而在寒武纪生物大爆发之后, 奥陶纪海洋中已经有一定数量的后生动物, 它们对蓝藻藻席有破坏作用, 因此凝块石几乎不可能形成于后生动物繁盛的正常浅海环境, 只能在后生动物不发育的环境中发生。
鄂尔多斯盆地在马五段沉积期主要为局限台地环境, 总体特点为“ 陆外为坪, 云坪广布, 坪中有湖, 湖中有膏, 湖中有盐” (冯增昭和鲍志东, 1999; 史基安等, 2009; 王国亭等, 2018), 即主体为局限台地, 台地上广布洼地和云坪。在海平面下降的情况下, 大台地上的洼地变成蒸发潟湖环境, 持续强烈的蒸发作用会导致盐度过高不适合蓝细菌生活, 不能形成微生物岩, 但是可以有蒸发岩形成。在海平面小幅度上升的情况下, 大台地上的洼地中水体盐度降低, 进入到蓝细菌可以生活的范围, 但是尚没有达到正常海洋盐度, 所以蓝细菌等微生物群落得以生活, 形成微生物岩沉积。由于这种非正常盐度仍不适合后生动物生活, 所以微生物群落免遭后生动物的破坏。
这些微生物岩究竟形成于多高的盐度环境?Arp等(2012)对太平洋岛国基里巴斯的现代微生物沉积进行了研究, 发现基里巴斯岛屿上封闭潟湖海水的盐度最高达到132‰ ~149‰ , 是正常海洋盐度的4倍以上(太平洋海水的平均盐度为35‰ ), 温度32.0~33.6 ℃, pH值大于8。在这样的环境下, 湖底生长了1层几毫米厚的黄绿色微生物席, 在微生物席的下面形成了厚达400 mm的多孔状微生物碳酸盐。这一现代实例说明, 微生物群落可以在盐度高达正常海洋4倍的水体中正常生长并形成碳酸盐沉积。由于当海水盐度高于60‰ 后, 后生生物几乎全部消失(卫民, 1999), 所以基里巴斯潟湖的超高盐度环境不适合任何后生动物生活, 微生物席不会遭到后生动物的破坏。
此次研究未在鄂尔多斯盆地北东地区马五段中发现后生动物化石碎片, 说明盐度不适合后生动物生活。后生动物的不发育, 可能指标盐度高于后生动物的忍耐范围, 即高于60‰ 。虽然不清楚盐度多大会使微生物群落消失, 但是, 由基里巴斯潟湖的实例可以推测鄂尔多斯盆地北东地区马五段微生物岩形成环境的盐度最大可以达到正常海水盐度的4倍。所以, 研究区的海水古盐度范围可能在60‰ ~149‰ 之间。
根据以上分析, 建立了鄂尔多斯盆地北东地区马五段微生物岩和含石膏结核白云岩的形成模式(图 8)。在低海平面时期, 台地的低洼处蒸发作用强烈, 盐度超过149‰ , 微生物群落不发育, 形成含石膏结核的碳酸盐沉积。在相对海平面升高的情况下, 台地的低洼处盐度降低到149‰ 以下, 微生物群落开始发育, 形成微生物碳酸盐沉积。
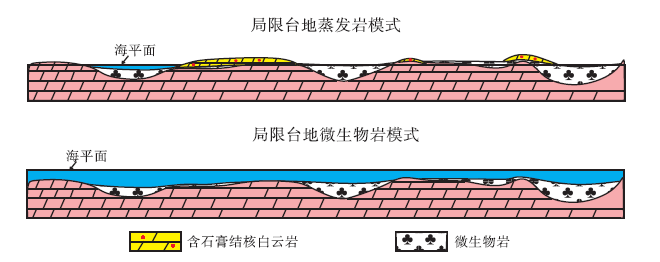 | 图 8 鄂尔多斯盆地中奥陶统马家沟组五段蒸发岩和微生物岩的沉积模式Fig.8 Sedimentary model of evaporite and microbialite of the 5th Member of Middle Ordovician Majiagou Formation in Ordos Basin |
凝块石(Aitken, 1967)指缺乏纹层而具凝块结构的一类岩石, 凝块的大小可以从厘米级到毫米、微米级(罗平, 2013)。前人对凝块石进行进一步划分, 为凝块石的深入研究提供一种新的思路。如熊鹰等(2016)对鄂尔多斯盆地奥陶系马家沟组马五1+2段微生物岩特征做了细致描述, 将微生物岩划分为叠层石、菌纹层白云岩、肾形菌凝块石以及与微生物相关的颗粒岩4种类型。但从该文的图版可以看出, 其肾形菌凝块石形态与肾形菌中空、膨大的囊形成的葡萄状群体的特点差别太大, 不应定为肾形菌凝块石, 而是非钙化蓝细菌成因的凝块石。
王玉伟等(2019)研究了塔里木盆地下奥陶统鹰山组和中奥陶统一间房组微生物岩的类型和储集层特征, 将微生物岩划分为葛万菌凝块石和隐藻凝块石, 其中隐藻指非钙化蓝细菌。这个分类的问题是藻类是指真核生物, 而蓝细菌是原核生物。由于前寒武纪和早古生代的微生物岩主要是原核生物形成的, 所以叫隐藻凝块石比较不妥。本次研究的4口井中未见葛万菌化石, 均为非钙化蓝细菌成因凝块石。基于目前文献中缺乏对塔里木盆地奥陶纪凝块石的科学分类, 故鄂尔多斯盆地马家沟组凝块石与塔里木盆地的对比工作难以深入进行。
肖传桃等(2018)研究了湖北松滋地区下奥陶统凝块石, 认为其形成于开阔台地和局限台地交替的潮间— 潮下带环境, 其形态与后生生物扰动有关, 并根据形态的不同进行分类命名。而本文研究的凝块石微生物岩中未发现任何后生生物的碎片, 可见与后生动物无关, 且其云坪广布的局限台地形成环境与肖传桃等(2018)研究的微生物岩环境差别较大, 也不易进行对比。
Meng等(2017)研究了鄂尔多斯盆地奥陶系榆9井马家沟组的蒸发岩和叠层石微生物岩, 发现榆9井马家沟组五段的叠层石微生物岩最发育, 其形成于高盐度卤水的蒸发岩环境中。然而, 该文对叠层石和蒸发岩形成时蒸发卤水的盐度差别没有探讨。
从上述对比可以看出, 鄂尔多斯盆地马五段独特的强蒸发的高盐度沉积环境, 造就了现今能够观察到的不同显微结构的凝块石微生物岩, 其与地史时期中已报道过的凝块石微生物岩形成环境差别太大。由于目前对古盐度的测试一般仅限于海陆相环境海水或者淡水的确定(蒋一鸣等, 2020), 或者只是定性地判断是否为正常海水或者高盐度咸水环境(高飞等, 2019), 定量地测量古盐度的方法还未见报道, 故文中未进行相关的地球化学指标测试。因此, 对研究区域内古盐度的推测暂以现今太平洋岛国基里巴斯现代微生物沉积的研究为依据, 以为将来更深入的研究凝块石的形成环境提供参考。
另外, 熊鹰等(2016)认为凝块石微生物岩可发育成具有一定规模的微生物丘, 具有一定的储集空间和储集性能, 是油田未来的勘探目标。笔者的研究也发现, 凝块石白云岩具有丰富的溶蚀空间, 是较好的油气储集层。
1)通过对鄂尔多斯盆地中奥陶统马家沟组马五段岩心的研究, 认为马五
2)结合现代微生物岩的研究成果, 推测在华北地台云坪广布、坪中有湖的大背景下, 凝块石微生物岩和含石膏结核碳酸盐分别形成于相对海平面升高和降低的时期, 海水古盐度范围分别为60‰ ~149‰ 和大于149‰ 。
3)对岩石薄片显微结构的分析还发, 现蠕虫状凝块石泥粉晶白云岩的白云石晶间孔和溶蚀孔非常发育, 网状凝块石和房室孔洞状凝块石的溶蚀残余孔较发育, 因此凝块石微生物岩具较好的油气储集空间, 是未来油气勘探的重要目标之一。
(责任编辑 张西娟; 英文审校 陈吉涛)
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|